
Efectos de la Altura y de la Hipoxia Aguda sobre el VO<sub>2</sub> máx
IE Faria Exercise Physiology Research Laboratory, Department of Kinesiology and Health Science, CSU Sacramento, 6000 J Street, Sacramento, CA 95819R.B.
Artículo publicado en el journal PubliCE, Volumen 0 del año 2004.
Publicado 1 de septiembre de 2004
Resumen
Palabras clave: ácido-Base, equilibrio, hídrico, volumen muscular, ejercicio
HIPOXIA Y CAMBIOS EN EL VO2 MAX.
Primeros Estudios
Si bien es importante destacar que West (49) realizó una crónica acerca del interés del hombre en los efectos de la altura sobre la fisiología humana desde los días de Aristóteles, el real empuje para entender los efectos de la altura sobre el rendimiento deportivo comenzó a mediados de 1960 con el anuncio de los Juegos Olímpicos de México (2250 m). En ese momento no eran claros los efectos que esta altura moderada tendría sobre los resultados de los juegos, si es que en realidad había alguno. Uno de los primeros estudios realizados por Balke, Nagle y Daniels (3), en Red River, NM (2300 m) indicó que, en efecto, esta moderada altura tendría un efecto sobre la capacidad cardiovascular y el rendimiento en la carrera. Una vez que arribaron a la altura estos investigadores observaron una inmediata reducción en el VO2 máx. (~6 %) y en el rendimiento en una prueba de una milla (~9 %). Luego de diez días en la altura el rendimiento era todavía 4.5 % menor con respecto a los valores a nivel del mar. Los resultados de este estudio sugirieron que aun con diez días de aclimatación para recuperar el VO2 máx., el rendimiento en la altura estaba todavía reducido. En un esfuerzo para entender mejor los cambios metabólicos producidos por la exposición a la altura, Consolazio y cols. (9) midieron los gases espirados en tres grupos diferentes de soldados que ascendieron a varias alturas. Estos investigadores reportaron una caída del 8 % en el VO2 máx. desde el nivel del mar hasta los 1610 m y una caída en el VO2 máx. del 14 % desde los 1610 mts hasta los 3475 m. Esto representó un mayor porcentaje de disminución que el reportado por Balke y cols. (3). Durante los 19 días en la altura Consolazio y cols. documentaron una disminución en la deuda de O2 (EPOC) y hallaron que esta era la principal adaptación a la altura con muy poco cambio en el VO2 máx. Los hallazgos de Consolazio y cols. sugirieron que la aclimatación tendría poco efecto sobre la recuperación del VO2 máx. a nivel del mar. Este hallazgo esta en desacuerdo con los hallazgos de Balke y cols. (3).
Para examinar el efecto de la hipoxia sobre el VO2 máx. y sobre la capacidad de trabajo, Dill y cols. (13) realizaron una serie de evaluaciones progresivas en cicloergómetro a 535, 485 y 455 Torr en una cámara hipobárica. Su grupo halló que el VO2 máx. cayó 10, 14, y 19 % respectivamente. La declinación en el VO2 máx. observada por Dill y cols. fue similar a las declinaciones reportadas por otros investigadores (3, 9), sin embargo, el menor cambio en la máxima carga de trabajo que en la disminución en el rendimiento máximo en la carrera, fue un hallazgo sorprendente. Dill y cols. atribuyeron las diferencias observadas al menor tiempo de exposición que sus sujetos tuvieron en la cámara hipobárica en relación con los estudios realizados con carreras en la altura. Para determinar la tasa de adaptación y para observar si un período más prolongado de exposición afectaría el rendimiento en la carrera al volver al nivel del mar luego del entrenamiento en la altura, Buskirk y cols. (8) llevaron a entrenar a un grupo de atletas de pista a una altura de 4000 m. El entrenamiento involucró una combinación de intervalos, repeticiones de carreras, ejercicios de calistenia, y fútbol. Una vez que arribaron a la altura se redujeron la intensidad y duración del entrenamiento al 40 % del que realizaban al nivel del mar. Durante el tiempo de estadía los atletas fueron capaces de incrementar la intensidad y la duración hasta el 75 % del entrenamiento que realizaban al nivel del mar. Buskirk y cols. reportaron que con esta altura se observó una caída del 26 % en el VO2 máx. y reportaron que se requirió de un período de cuatro a cinco meses para que los atletas pudieran competir en fútbol, con los residentes, en iguales condiciones. Sin embargo, el descubrimiento más importante de este trabajo fue que, mientras que los atletas fueran entrenados durante su estadía en la altura, estos no sufrirán una pérdida apreciable en el rendimiento en la vuelta al nivel del mar. El hallazgo de Buskirk (8) de que la estadía en la altura no perjudicaba el rendimiento en la vuelta al nivel del mar fue un punto importante a considerar en el entrenamiento de los atletas para los juegos a realizarse en la ciudad de México. Sin embargo, Faulkner, Daniels y Balke (16) teorizaron que si los atletas entrenaban a una altura moderada durante un período de tiempo suficiente, serían capaces de competir a una altura moderada como si estuvieran al nivel del mar. Utilizaron dos grupos de atletas, corredores y nadadores, cada uno de los cuales tomo parte en un campamento de entrenamiento a una altura moderada (~2300 m). Lo que hallaron fue similar a lo que habían hallado en su investigación original en Red River, NM. Primero, no hallaron reducciones en el rendimiento en distancias menores a los 800 m. Segundo, hallaron que en distancias mayores a los 800 m, el rendimiento se reducía significativamente en la altura y se mantenía significativamente reducido aun después de un período de aclimatación de 21 días. Finalmente, hallaron que luego de 21 días el VO2 máx. se recuperaba hasta el valor obtenido al nivel del mar. Este hallazgo acerca del VO2 máx. estaba en contraste con su investigación original, sugiriendo que el VO2 máx. podría recuperarse en 10 días a una altura moderada. Aunque el hallazgo recibió poca atención, fue interesante que los autores reportaran que la altura no tuvo casi efectos sobre el rendimiento en la natación, sugiriendo que las respuestas a la natación diferían de las respuestas a la carrera.
Si bien los Juego Olímpico realizados en la Ciudad de México sirvieron de impulso a estas primeras investigaciones, es difícil delinear una conclusión real a partir de los mismos. La mayoría de las investigaciones no tuvieron un objetivo definido y por ello no estuvieron bien controladas. Muchos de ellos fueron solamente estudios descriptivos con muestras pequeñas haciendo que sea aun más difícil extraer conclusiones. Sin embargo hubo algunas semejanzas en los hallazgos de los estudios. Primero, al parecer la declinación en el VO2 máx. y en el rendimiento se intensifican durante la primer semana en la altura hasta que comienza el proceso de adaptación. Segundo, aun cuando los atletas puedan recuperar el VO2 máx. del nivel del mar en la altura es improbable que recuperen el rendimiento que tenían al nivel del mar. Tercero, debido a que hay muchos reportes conflictivos acerca de la magnitud en la reducción del VO2 máx. a una determinada altura, parece obvio que hay una gran variabilidad de persona a persona en la magnitud de declinación que se experimenta.
CAMBIOS AGUDOS
Debido a la falta de control en los primeros estudios muchos científicos comenzaron a realizar estudios más controlados utilizando cámaras hipobáricas y sistemas que utilizan gases hipóxicos. Estos estudios determinaron la caracterización de la respuesta del VO2 máx. con el incremento de la hipoxia.
Elliot y Atterbom (15) documentaron la reducción en el VO2 máx. con condiciones hipobáricas equivalentes a 1576 m y hasta 3962 m durante una ergometría realizada con los miembros inferiores en sujetos que residían a moderada altura (~1500 mts). Tanto los hombres como las mujeres mostraron una reducción en el VO2 máx. del 18 %. La caída del 18 % observada en este estudio fue ligeramente más baja comparada con la caída del 26 % observada a 4000 m, reportada por Buskirk (8), pero pudo deberse a la aclimatación de los sujetos a una altura moderada. Squires y Burkirk (45) documentaron los cambios en el VO2 máx. en una cámara hipobárica a 730, 681, 656, 632, y 574 Torr. Estas presiones son equivalentes a alturas de 362, 914, 1219, 1524 y 2286 m respectivamente. Squires y Buskirk (45) hallaron que la primer caída, estadísticamente significativa, en el VO2 máx fue con una condición barométrica equivalente a 1219 m. A 1219 m hallaron una caída del 5 % en el VO2 máx. y una caída del 12 % a 2286 m. El patrón de reducción en esta investigación sugirió que la declinación en el VO2 máx es lineal con el descenso de la presión barométrica. Levitan y cols. (28) obtuvieron hallazgos similares utilizando gas inspirado con presiones de O2 de 20.9 % y 15.5 % (equivalentes a 2440 m). Levitan observó una declinación en el VO2 máx. del 8.3 %, un hallazgo similar a la declinación en el VO2 máx. del 12 % observada por Squires y Buskirk (45).
A través de los datos obtenidos por Andersen y cols. (2) se pudo obtener una mejor comprensión acerca del patrón de declinación en el VO2 máx. Andersen y cols. realizaron una serie de test en cicloergómetro con condiciones barométricas equivalentes al nivel del mar, 2500, 3750, 4375, 4690, y 5000 m. Estos investigadores descubrieron que el VO2 máx. disminuía de manera curvilínea mas que lineal, como lo habían reportado Squires y Buskirk (45). La extrapolación de los datos reportados por Andersen y cols. (2) sugiere que las reducciones más severas en el VO2 máx. comienzan con 525 Torr (3050 m). Los autores sugirieron que esto se debió a una limitación en la ventilación, ya que estos notaron una meseta en la ventilación máxima a una PIO2 de 93 Torr, lo cual está cerca de la PIO2 de 100 Torr a 3050 m. Dill y cols. (12) han especulado también con que la caída en la función pulmonar desempeña un rol en la disminución del VO2 máx. Utilizando datos recolectados a lo largo de años de estudio en las instalaciones de investigación de White Mountain, Dill y cols. hallaron que con la altura había una caída del 20 % o más en la capacidad vital de sujetos ancianos (edad 58-71). Dill y cols. especularon además que esta caída en la capacidad vital podría derivar en mayores disminuciones del VO2 máx. en sujetos ancianos. Sin embargo, hallaron que la tasa de declinación en el VO2 máx. era similar en sujetos jóvenes y ancianos. Sugiriendo que los cambios en la capacidad vital no estuvieron relacionados con la declinación en el VO2 máx.
Para comprender mejor las diferencias sexuales de la respuesta del VO2 máx. durante la hipoxia. Paterson y cols. (36) estudiaron a cuatro mujeres y a tres hombres utilizando gas inspirado con concentraciones de O2 de 20.93 %, 17.39 %, 14.4 % y 11.81 % (equivalentes al nivel del mar, 1500, 3000 y 4500 m). Paterson halló un 24 % de declinación en la mujeres y un 29.5 % de declinación en los hombres con una concentración de O2 de 11.81% equivalente a 4500 m. Si bien estos valores sugieren que los hombres y las mujeres responden de manera diferente a la altura, es importante señalar que los valores son muy similares a la caída del 26 % demostrada por Buskirk y cols. (8) a 4000 m.
La mayoría de estos estudios fueron realizados a alturas y con gases con presiones parciales equivalentes a alturas donde la mayoría de las personas viajan por recreación y competición. Sin embargo, se ha hecho poco por examinar los ambientes más extremos de las grandes alturas. Por ello, un grupo de científicos se propuso la tarea de examinar la caída del VO2 máx. durante un ascenso simulado al Monte Everest. El estudio se conoció como Operación Everest II (10). Los hallazgos de esta investigación indicaron una caída del 70 % en el VO2 máx. con un equivalente hipobárico a ~8932 m.
Finalmente, Maresh y cols. (30) examinaron el efecto de permanecer en la altura a corto y largo plazo sobre el VO2 máx. Los investigadores observaron que no hubo diferencias significativas en el VO2 máx. entre la permanencia a corto y largo plazo. Este hallazgo sugiere que el proceso de aclimatación solamente contribuye a la respuesta al ejercicio submáximo y no al ejercicio máximo.
Los estudios durante este período demostraron además la variabilidad de las respuestas del VO2 máx. a la hipoxia. Sin embargo, estos estudios proporcionaron una descripción del patrón de cambio en un ambiente más controlado que no estuvo confundido por el entrenamiento, el viaje, u otras condiciones ambientales. Si bien estos estudios proporcionaron una caracterización de la declinación en el VO2 máx., había todavía un debate considerable acerca de los factores que influencian el VO2 máx. en la altura.
FACTORES INDIVIDUALES QUE AFECTAN EL VO2MAX EN HIPOXIA
Uno de los cambios más notables en el proceso de aclimatación es el incremento en la masa de glóbulos rojos. Robertson y cols. (40) especularon con que la policitemia aumentaba el VO2 máx. en la altura. Cinco escaladores realizaron una evaluación para medir el VO2 máx. a nivel del mar y bajo condiciones hipóxicas (fracción de O2 inspirado =13.5 % equivalente a 3566 m). Luego de las evaluaciones se les extrajeron muestras de sangre para el subsiguiente almacenamiento de sus glóbulos rojos. Ocho semanas después se les realizó a los escaladores la reinfusión de sus glóbulos rojos, por lo cual se incrementó su masa de glóbulos rojos. La reinfusión produjo un incremento en el hematócrito del 26.5 % y de la hemoglobina en un 27.7 %. Estos cambios hematológicos incrementaron el VO2 máx. al nivel del mar en un 20.9 % y en un 9.8 % bajo condiciones hipóxicas. Estos hallazgos sugieren que las adaptaciones de los glóbulos rojos desempeñan un rol en los cambios en el VO2 máx. en la altura. Sin embargo, en contraste con este artículo, Young y cols. (51) realizaron la reinfusión de glóbulos rojos a 16 hombres saludables y a pesar del incremento de la concentración de hemoglobina por la reinfusión, la misma tuvo poco efecto sobre la declinación en el VO2 máx. en comparación con un grupo control al que se le realizó una reinfusión con un volumen igual de solución salina. Estos hallazgos sugieren que la masa de glóbulos rojos tiene poco efecto sobre la respuesta del VO2 máx. a la exposición crónica a la altura. Los hallazgos de estos dos artículos también sugieren que no es claro que rol desempeñan los glóbulos rojos con respecto a la influencia sobre el VO2 máx. en la altura.
Tucker y cols. (47) examinaron la saturación arterial de oxígeno en la altura en 12 miembros del equipo de atletismo del la Universidad del Estado de Colorado. Tucker y cols. hallaron que desde 760 Torr a 635 Torr (~1500 m) la SaO2 caía en promedio un 9.4 % y caía un 10.8 % adicional desde 635 Torr a 525 Torr (~3000 mts). En relación a estas reducciones en la SaO2, Tucker halló también una caída del 6.5 % y del 22 % en el VO2 máx. a 635 y 525 Torr, respectivamente. Estos hallazgos indicaron que cuanto mayor es la reducción en la SaO2 mayor es la declinación en el VO2 máx. El resultado de esta investigación indica que la desaturación arterial también contribuye a los cambios en el VO2 máx. bajo condiciones hipóxicas. Posteriormente Lawer, Powers y Thompson (27) quienes evaluaron a sujetos utilizando gas inspirado con una concentración de O2 de 21 % y 14 % confirmaron estos hallazgos. Las mayores reducciones en el VO2 máx. fueron observadas en aquellos sujetos con la mayor desaturación arterial. La importancia de una alta VE y por lo tanto una alta SaO2 fue reforzada por el hallazgo de Sutton y cols. (46) que examinó el transporte de O2 durante los estudios realizados en la OEII y por Shepard y cols. (42) quien de manera similar halló que una alta VE limitaba la reducción en la SaO2. La relación entre la presión barométrica y su efecto sobre el VO2 máx. y la SaO2 en el VO2 máx. puede observarse en la Figura 1.
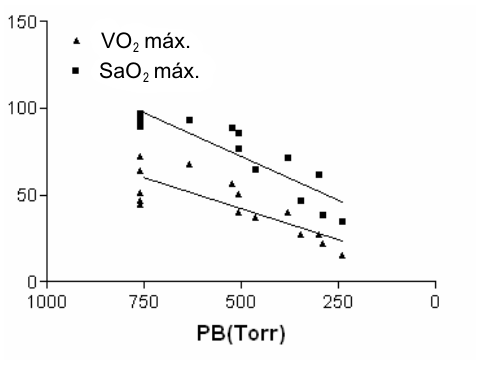
Figura 1. Los datos muestran el % de reducción en la saturación arterial de O2
y en el VO2 máx. con el descenso de la presión barométrica (PB). Los
datos fueron obtenidos de Cymerman y cols. (10), Lawer y cols. (27), Sutton y
cols. (46) y Tucker y cols. (47); y muestran una reducción paralela. Las
pendientes de cada recta no son estadísticamente diferentes (p=0.14).
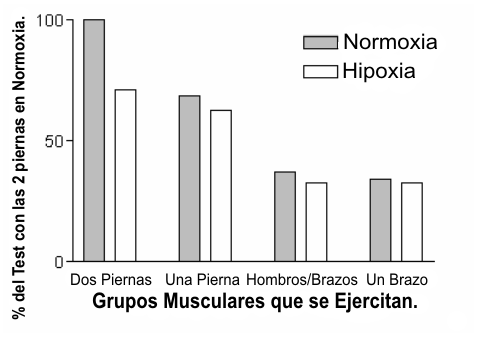
Figura 2. Porcentaje de reducción en el VO2 máx. cuando se realizó
ejercicios con diferentes volúmenes musculares. Obsérvese que el efecto de la
hipoxia sobre el VO2 máx. es mucho menor cuando se involucra un menor
volumen muscular. Los datos fueron obtenidos de Shepard y cols. (41) y para
simplificar el gráfico se agruparon los datos de hombres y mujeres.
Para comprender mejor el rol de la función hormonal en la determinación del VO2 máx. bajo condiciones hipóxicas, Boissou y cols. (7) estudiaron ocho hombres que realizaron una cicloergometría al 40, 60, 80, y 100 porciento del VO2 máx. Estos autores no hallaron diferencias en los índices insulina/glucagón, ∆-4 androstenediona/testosterona, y tampoco hallaron diferencias en las catecolaminas con la hipoxia. Estos hallazgos indicaron que la función hormonal no desempeña un rol en la determinación del VO2 máx.
Para determinar el efecto de la masa muscular activa sobre la declinación del VO2 máx., Shepard y cols. (41) realizaron evaluaciones del VO2 máx. en ocho hombres y en ocho mujeres. Cada sujeto completó un test hasta el VO2 máx. con dos piernas, con una pierna, con hombros/brazos, y con un solo brazo. Cada test fue realizado tanto bajo condiciones de normoxia como bajo condiciones de hipoxia (12 % de O2). Cada reducción en la masa muscular activa resultó en una reducción más pequeña en el VO2 máx. En promedio la hipoxia provocó un 30 % de reducción en el VO2 máx. durante el test realizado con las dos piernas, mientras que la hipoxia en el test realizado con un solo brazo provocó una reducción de tan solo el 2 % en el VO2 máx. Asimismo, cada reducción en la masa muscular resultó en una reducción más pequeña correspondiente (Figura 2). Estos datos muestran que la masa muscular es el factor que más contribuye a la reducción del VO2 máx. bajo condiciones hipóxicas, y puede explicar porque Faulkner, Daniels y Balke (16) hallaron una caída muy pequeña en el rendimiento en natación en la altura. Las investigaciones en natación han documentado que la mayor parte de la propulsión es realizada con los brazos y podría por ello representar una menor masa muscular activa.
La más prominente de todas las variables en la determinación de la caída del VO2 máx. bajo condiciones hipóxicas es el VO2 máx. a nivel del mar. Youn y cols. (52) fueron los primeros en mostrar que a medida que se incrementa el VO2 máx. individual a nivel del mar el porcentaje de caída en el VO2 máx. será mayor bajo condiciones hipóxicas. Este hallazgo fue algo imprevisible, ya que se esperaría que una persona con un alto VO2 máx. al nivel del mar muestre una menor y no una mayor declinación ya que su cuerpo tiene una capacidad mejorada para utilizar el oxigeno. Shepard y cols. (42) mostraron que un alto VO2 máx. de hasta ~48 ml.kg¹.min¹ al nivel del mar, era en efecto protector contra la caída del VO2 máx. bajo condiciones de hipoxia. Por encima de ~48 ml.kg¹.min¹ un alto VO2 máx. fue asociado con una mayor declinación del mismo bajo condiciones de hipoxia. Los efectos del VO2 máx. al nivel del mar sobre la reducción del VO2 máx. durante la hipoxia fueron respaldados por los trabajos de Lawler y cols. (27) y Martin y McKroy (31) donde cada uno demostró mayores caídas en el VO2 máx. durante la hipoxia con un mayor VO2 máx. a nivel del mar.
Estos estudios demostraron que son varios los factores que se relacionan con la declinación en el VO2máx. La desaturación arterial, la masa de glóbulos rojos, la masa muscular activa, y el VO2 máx. al nivel del mar, contribuyen a la reducción del VO2 máx. Conjuntamente estos factores explican el amplio rango de reducciones en el VO2 máx. mostrados en los primeros estudios sobre el mismo durante la hipoxia.
MODELOS DE CAMBIOS EN EL VO2 MAX.
Debido a que se ha demostrado que un número de variables desempeñan un rol en la reducción del VO2 máx. bajo condiciones hipóxicas, los investigadores comenzaron a modelar estos factores para determinar como interactuaban unos con otros.
Ferretti (17) observó que la reducción en el VO2 máx. era una imagen en espejo de la curva de disociación de la oxihemoglobina. Esta observación vinculó los trabajos de Squires y Buskirk (45) y de Andersen y cols. (2). Squires y Buskirk habían sugerido que la respuesta del VO2 máx. a la altura era lineal, sin embargo habían evaluado solamente la respuesta en condiciones equivalentes a una altura de 2286 m lo que derivó en la porción relativamente plana de la curva de disociación de la oxihemoglobina. La evaluación por encima del equivalente a los ~3000 m los hubiera puesto en la porción más empinada de la curva de disociación de la oxihemoglobina y podrían también haber observado la respuesta curvilínea observada por Andersen y cols., ya que estos evaluaron hasta una altura equivalente a los 5000 m.
Ferretti también derivó un modelo matemático de los cambios en el VO2 max. con la altura. Los datos para este modelo estuvieron basados en observaciones previas de otros investigadores. En este modelo Ferretti propuso que aproximadamente el 70 % del VO2 max. estaba determinado por el gasto cardiaco máximo a nivel del mar. A medida que se incrementaba la altura hasta aproximadamente 4300 m el gasto cardiaco máximo determinaba solamente el 48 % del VO2 max. y el equilibrio de la respuesta del VO2 max. estaba determinado por el transporte periférico de O2.
De manera similar, Robgers y cols. (39) construyeron un modelo para explicar la varianza en la reducción del VO2 max. Robgers realizó evaluaciones en cicloergómetro en 28 sujetos en condiciones hipobáricas equivalentes al nivel del mar y a 2439 m. Si bien este rango de condiciones hipobáricas no es muy amplio, representa el rango característico de las condiciones para la competición y la recreación. Utilizando análisis de regresión múltiple hallaron que un mayor VO2 max. a nivel del mar, un bajo umbral de lactato, una baja SaO2 máx., un gran masa magra corporal y el sexo femenino derivaban en las mayores reducciones del VO2 max. a 2439 m. Debido a que el umbral de lactato sirve como variable indicadora del transporte periférico de O2, los hallazgos de Robgers acuerdan con el modelo propuesto por Ferretti (17). La mayor cantidad de varianza que explicó la declinación en el VO2 max. fue el VO2 max. a nivel del mar. Nuevamente este hallazgo estuvo de acuerdo con los hallazgos previos de otros investigadores. Young, Cymmerman y Burse (51) como también Shepard y cols. (42) hallaron que los sujetos con un mayor VO2 max. a nivel del mar tenían un mayor porcentaje de reducción en el mismo. En efecto, un alto VO2 max. a nivel del mar es con todo el mayor detrimento para la declinación del VO2 máx. en la altura.
La relevancia de un alto VO2 máx. a nivel del mar en la determinación del porcentaje de declinación en el VO2 máx. en la altura fue esclarecida por Wagner (48). Wagner halló que al nivel del mar el VO2 máx. de los individuos más entrenados (mayor VO2 máx.) estaba limitado por el transporte de oxígeno hacia los músculos, mientras que en los individuos menos entrenados (menores VO2 máx) el mismo estaba limitado por la utilización periférica de O2. Este modelo podría explicar porque aun las más pequeñas reducciones en la SaO2 producirían grandes reducciones en el VO2 máx. en la altura en sujetos bien entrenados. Wagner también confirmó el hallazgo de Robgers y cols. y de Ferretti. Además de demostrar la importancia del transporte de O2 al nivel del mar Wagner también demostró el incremento de la limitación de la utilización periférica de O2 bajo condiciones hipóxicas.
El modelo de Wagner nuevamente sugiere que los individuos con un alto umbral de lactato podrían minimizar la perdida de VO2 máx. en la altura.
Las investigaciones a lo largo de la última década han proporcionado un mayor entendimiento acerca del VO2 máx. en la altura. Las razones detrás de la respuesta lineal y curvilínea reportada previamente, son ahora mejor comprendidas. Principalmente, la varianza en las respuestas individuales que han sido documentadas desde mediados de 1960 han sido ahora reproducidas, verificadas como normales, y explicadas por varios modelos. Además, varias pruebas de laboratorio pueden ser inclusive, adecuadas para identificar quien se desempeñará bien en la altura y quien no, una pregunta que fue formulada originalmente antes de los Juegos Olímpicos realizados en la Ciudad de México.
RESPUESTAS FISIOLOGICAS AGUDAS A LA HIPOXIA
Una vez que se arriba a la altura las dos respuestas fisiológicas más prominentes a la hipoxia son la diuresis renal y el incremento en la ventilación minuto. No está claro cual es el efecto que estas respuestas tendrán sobre el VO2 máx. y sobre el rendimiento en la altura. Se ha demostrado claramente que la respuesta ventilatoria a la hipoxia tiene un pequeño efecto sobre el VO2 máx. en la altura (23, 33), sin embargo puede tener efectos secundarios (i.e., incremento de la perdida de fluidos y aumento del pH) que pueden influenciar el VO2 máx. Además, el incremento en la respuesta diurética a la altura deriva en deshidratación, y podría hipotetizarse que tiene efectos adversos sobre el VO2 máx. Estas respuestas a la altura llevaron a Daniels (11) a sugerir que los atletas deberían viajar a la altura a último momento antes de la competición o arribar con semanas de anticipación para que se produzca la aclimatación, la primera alternativa no es conveniente y la segunda tiene alto costo, algo que muchos atletas siguen todavía en la actualidad. La siguiente sección examina estas respuestas agudas e intenta ajustarlas a los modelos anteriormente mencionados para determinar su valor real o adverso sobre el VO2 máx.
BALANCE HIDRICO EN LA ALTURA
Cambios en los Compartimentos Hídricos en la Altura
La altura tiene un profundo efecto sobre el balance hídrico. A menudo es el efecto más común e inmediato de la altura que puede ser sentido por las personas que viajan a una altitud elevada. Jain y cols. (25) midieron los cambios en los compartimentos hídricos a 200 y 3500 m en 18 hombres saludables. Las mediciones fueron tomadas luego de 3 y 12 días a 3500 m. Lo que estos investigadores hallaron fue una reducción significativa en el agua corporal total (TBW) en los días 3 y 12 (-3.7 y -4.7 %, respectivamente) como así también en el agua intracelular (ICF) (-3.3 % y –4.3 %, respectivamente). El agua extracelular (ECF) también mostró una reducción significativa (6 %) en el 12vo. día. Dentro del compartimento extracelular, el volumen plasmático (PV) se redujo en un 16 %.
El incremento en la diuresis derivará en la deshidratación, e intuitivamente parecería que la perdida de fluidos en la altura podría ser potencialmente perjudicial. Sin embargo, Hackett y cols. (22) especularon con que la perdida de fluidos causada por la exposición aguda y crónica a la altura puede ser beneficiosa y en su estudio realizado con 102 expedicionistas en Nepal hallaron que aquellos que desarrollaron edema pulmonar por las grandes alturas (HAPE) y síntomas de AMS tuvieron una disminución en la frecuencia de excreción urinaria e incrementos en el peso corporal (BW). De manera similar Gunga y cols. (20) también examinaron los cambios en el balance hídrico sobre la densidad de los tejidos a una altura moderada. Utilizando ultrasonido para medir la densidad del tejido en la frente, la tibia y el esternón observaron un incremento significativo en la densidad del tejido de la frente y de la tibia. El incremento en la densidad del tejido estuvo asociado con un incremento en el peso corporal (75.5 a 76.2 kg) y del agua corporal total (44.21 L a 45.01 L). Finalmente Schirlo y cols. (43) llevaron a cabo un experimento similar examinando los cambios en el perímetro del muslo con exposiciones agudas a la hipoxia. Con el incremento de la hipoxia la circunferencia del muslo se redujo significativamente y la reducción en el perímetro fue proporcional al incremento en la diuresis (Figura 3). Esto sugiere, tal como Hackett había especulado (22), que la retención de fluidos o la antidiuresis está significativamente relacionada con el edema.
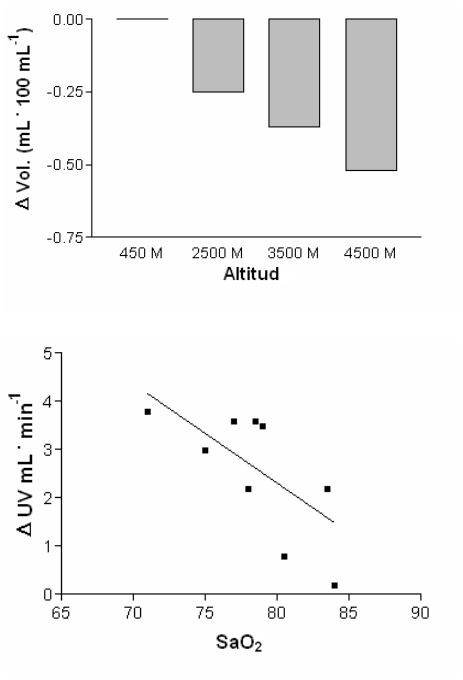
Figura 3. El gráfico superior muestra la disminución del volumen muscular con
el incremento de la altura. El gráfico inferior muestra la reducción en la SaO2
y el incremento en el volumen urinario (∆UV). Los datos fueron obtenidos de
Schirlo y cols. (43).
Con respecto a la AMS y al balance hídrico, Richalet y cols (38) midieron el PV, el BW y la AMS en 9 montañistas a 4950 m durante un período de 4 semanas. Durante este tiempo documentaron reducciones en el PV de 7 % y 10 % y en el BW de 3 % y 5 % en las semanas uno y cuatro, respectivamente. El único individuo que desarrollo AMS mostró incrementos en el BW y en el PV durante la primer semana y reducciones en estas variables en la 4ta. semana a medida que los síntomas disminuyeron. En respaldo de la importancia de la respuesta diurética como prevención de la AMS, Singh, Raul y Tyagi (44) hallaron que un grupo de sujetos aclimatados que fueron reintroducidos a la altura luego de pasar un período previo de 4 a 6 meses a 3500 m tuvieron un 11 % de incremento en el PV y una mayor susceptibilidad a la AMS que en la exposición previa. También se observó en este estudio una reducción significativa en los compartimentos hídricos luego de tan poco como 2 días. El PV se redujo significativamente en un 8 %, siendo similar a la reducción del 7 % observada por Richalet y cols. (38). También se observaron reducciones significativas en el BW, en el agua corporal total, en el fluido extracelular y en el fluido intracelular de 1.6 kg, 1.15 L, 0.49 L y 0.65 L, respectivamente. Bartsch y cols. (6) publicaron un resumen de tres estudios previos. Ellos observaron que el incremento en la ingesta de fluidos incrementaba la producción de orina. Además, más que proteger contra el desarrollo de AMS, el exceso de ingesta de fluidos puede incrementar la incidencia de AMS. También reportaron que la retención de fluidos en aquellos sujetos susceptibles a la AMS se debió a un incremento en la concentración de aldosterona en reposo.
Debido a la variabilidad de los estudios previos, Hoyt y cols. (24) decidieron examinar los efectos de la altura sobre los compartimentos hídricos en cabras. Las cabras fueron elegidas ya que proporcionan un buen modelo del balance hídrico en humanos sin que la AMS complique la interpretación de los resultados. Se examinaron cuatro cabras hembras al nivel del mar y a 380 Torr durante 16 días. El BW, el TBW, el ICF y el PV se incrementaron significativamente. Sin embargo, el volumen sanguíneo permaneció sin cambios debido a un incremento en el volumen de glóbulos rojos. El ECF se incremento significativamente debido a un incremento en el fluido intersticial. La mayor parte de la pérdida de peso observada se debió al ICF (- 4.04 L). Aunque los autores no lo mencionaron los datos parecen indicar que el incremento en el fluido intersticial se debe a cambios de fluido entre el PV y el compartimento ICF. A partir de estos datos los autores concluyeron que la mayoría de la perdida de fluido en las cabras, y probablemente en humanos, en la altura es a partir del compartimento ICF.
CAUSAS DE LA DIURESIS EN LA ALTURA
En la altura la constricción del sistema vascular pulmonar ocurre como un intento de equilibrar mejor la ventilación pulmonar con el flujo sanguíneo. Esta constricción causa un incremento en la carga del lado derecho del corazón y por lo tanto se incrementará el estiramiento de la aurícula derecha. El estiramiento de la aurícula derecha gatillará entonces la liberación del péptido atrial natriurético (ANP) (21). El efecto del ANP es que incrementa el clearance de sodio y por lo tanto lleva a la diuresis (37). Este mecanismo es la causa más probable de diuresis observada a grandes alturas y ha sido demostrada por Bartsch y cols. (4). Bartsch y cols. estudiaron a 25 montañistas (9 semanas/previo HAPE) a 550 m y a 4559 m luego de 6, 18 y 24 horas de exposición. Hallaron que los sujetos sin AMS tuvieron un mayor volumen urinario, una mayor perdida de peso, un hallazgo que fue similar a lo observado por otros investigadores. También observaron en los sujetos sin AMS, luego de 18 horas, reducciones significativas en la renina, la angiostesina II y en la aldosterona. En los sujetos con AMS el ANP fue mayor y la SaO2 menor sugiriendo que el ANP puede, de alguna manera, estar involucrado en el proceso de AMS. Sin embargo, cuando se analizaron los datos utilizando análisis de regresión múltiple, en donde se pudieron controlar las varianzas de otras variables, se observó una correlación positiva y significativa entre el ANP y la SaO2. En respaldo a este hallazgo, Westdrop y cols. (50) administraron dosis bajas de ANP, y, bajo condiciones hipobáricas [456 Torr (4115 m)] hallaron incrementos significativos en la SaO2 (79.6 % vs. 84.7 %), como también una reducción en el gradiente arterial-alveolar (7.3 vs. 3.5 mmHg). Esto sugiere que el ANP es el mecanismo de diuresis en sujetos no susceptibles a la AMS y que está altamente relacionado a la SaO2. Aunque los sujetos con AMS tuvieron una mayor concentración de ANP y una menor diuresis, también tuvieron elevadas concentraciones de catecolaminas, ADH y aldosterona. Bartsch y cols. (4) atribuyeron la falta de diuresis en vista de la elevada concentración de ANP a los efectos predominantes de las catecolaminas, ADH y aldosterona. Estas conclusiones fueron validadas por Koller y cols. (29). Koller utilizó dos grupos de 10 sujetos, unos aclimatados y otros no. Mientras estaban sentados en una silla estos sujetos fueron expuestos a un equivalente a una altura de 6000 m en una cámara hipobárica que fue descomprimida a una tasa de 300 m/min. La descompresión fue detenida cada 10 minutos para la observación. Koller y cols. obtuvieron hallazgos similares a los de Bartsch y cols. (14). Koller y cols. hallaron que el ANP se elevó significativamente en ambos grupos, así como también lo hizo la diuresis. Koller y cols. también observaron un aumento significativo en la ADH en los sujetos no aclimatados. Desafortunadamente debido a la brevedad de la exposición, la diuresis fue mayor en los sujetos no aclimatados, sin embargo debido a su perfil hormonal, si los sujetos no aclimatados hubieran permanecido en la altura por un mayor período de tiempo habrían comenzado a retener agua y hubieran incrementado la susceptibilidad a la AMS. Si bien la aldosterona y la ADH pueden compensar el aumento en el ANP, puede ser necesario re-evaluar el rol de las catecolaminas ya que Grover y cols. (21) le administraron propanolol a sujetos expuestos a una altura de 4300 m y no hallaron efectos sobre el balance hídrico durante las tres semanas a esta altura.
Mientras que estos artículos documentan los cambios en el balance hídrico y los cambios hormonales asociados con los mismos, Bartsch y cols. (5) sintieron que habían malogrado su intento debido a la alta tasa de ascenso. Ningún montañista podría ascender tan rápido y por ende las respuestas no fueron características de las que podrían observarse en las montañas. Bartsch y cols. teorizaron que con un ascenso más lento el tiempo de aclimatación aliviaría los síntomas de AMS que habían observado en aquellos susceptibles de AMS. En este estudio 15 montañistas varones (con historia de AMS seis semanas atrás) caminaron desde los 1170 m hasta los 4559 m con una parada nocturna a los 2864 m y dos noches a los 3611 m. Nuevamente hubo un incremento significativo en el ANP en los sujetos con AMS y no hubo incrementos en el ANP, ADH, renina o aldosterona en los sujetos sin AMS. Esto llevó a Bartsch y cols. (5) a concluir que un ascenso lento no previene la AMS en aquellos sujetos que son susceptibles.
El mecanismo de liberación de ANP es el estiramiento de la aurícula derecha (22). Sin embargo, es posible que la liberación de ANP estuviera ligada a la respuesta hipóxica. Tanto Bartsch y cols. (5) como Ponchia y cols. (37) examinaron las causas de la liberación de ANP en la altura. Como se esperaba ambos concluyeron que la liberación de ANP estaba relacionada al volumen auricular. Sin embargo, Ponchia y cols. hallaron una correlación significativa con la reducción del volumen auricular izquierdo más que con el volumen auricular derecho como se esperaba. Sin embargo, una examinación más cercana de sus datos reveló un incremento medio del volumen auricular derecho, pero con una variabilidad muy alta, lo cual impediría que hallaran una diferencia significativa.
En oposición a los estudios previos, los cuales demostraron un incremento en la diuresis en la altura, Anard y cols. (1) hallaron un incremento en la retención de fluidos en respuesta a la altura. Anard y cols. examinaron a 10 sujetos a 6000 m durante 10 semanas. Durante este tiempo observaron un incremento en la retención de fluidos en todos los espacios incluyendo un incremento del 33 % en el PV y un incremento del 84.5 % en el volumen sanguíneo. Esta respuesta fue atribuida a una reducción del 37 % en el flujo sanguíneo renal, a un incremento del doble en la concentración de aldosterona, y a ningún cambio en la concentración de ANP. Es difícil decir porque estos resultados son contrarios a los de tantos otros estudios, pero vale la pena señalar que algunos sujetos pueden responder de manera opuesta a la esperada.
A partir de estos artículos es aparente que hay un incremento en la diuresis con el incremento en la altura o en la hipoxia. Este incremento en la diuresis parece además ser causado por un incremento en el ANP relativo al incremento de las hormonas que conservan los fluidos y tiende a tener un efecto sobre todos los compartimentos hídricos. La diuresis es una respuesta importante a la altura y a la hipoxia, no solo debido a que limita los síntomas de AMS sino también porque la SaO2 parece ser mayor con un adecuado efecto diurético. Además, el volumen de los miembros parece disminuir con el incremento en la diuresis y podría teóricamente ayudar a facilitar la difusión de O2 en el músculo. Una elevada SaO2 y una mejora de la difusión han mostrado ser variables importantes para minimizar la reducción del VO2 máx. en la altura (17, 39). Esto sugeriría que la pérdida de fluidos puede ser ventajosa para minimizar la reducción del VO2 máx. en la altura, más que perjudicial tal como los sugiriera Daniels (11).
EQUILIBRIO ACIDO BASICO EN LA ALTURA
Junto con el incremento en la diuresis una de las respuestas más inmediatas a la altura y a la hipoxia es el incremento en la ventilación. El incremento en la ventilación causa una reducción en la presión parcial de CO2 en la sangre. A medida que el contenido arterial de CO2 cae, el pH plasmático se incrementará desarrollando un gradiente entre los tejidos y el plasma y llevando eventualmente a un incremento sistémico en el pH. Luego de un período prolongado de tiempo los riñones comenzaran a excretar bicarbonato y el pH del cuerpo retornara al normal. Esta respuesta ha sido bien caracterizada y resumida con las investigaciones previas de Millege (33).
Para demostrar el efecto de la hipoxia sobre el nivel ácido-básico y su efecto sobre el rendimiento humano, McLellan, Jacobs y Lewis (32) examinaron a cuatro sujetos al nivel del mar y bajo condiciones hipobáricas (445 Torr = ~4200 m). La exposición aguda duró dos horas y durante este tiempo el pH se incremento significativamente desde 7.396 hasta 7.446. En las pruebas de rendimiento que midieron el tiempo hasta la fatiga al 90 % del VO2 máx. específico en la hipoxia, los investigadores observaron un incremento promedio de 3.3 minutos (p<0.05) en la altura. Es imposible decir si en este estudio el incremento en el rendimiento fue resultado de la amortiguación de los iones de hidrógeno o del desplazamiento hacia la izquierda en la curva de disociación de la oxihemoglobina. Una intensidad de esta magnitud (90 % del VO2 máx.) acumularía rápidamente iones hidrógeno y el incremento en la capacidad de amortiguación de estos iones sería beneficiosa para el rendimiento. Sin tener en cuenta la causa, este estudio demuestra que aun una corta exposición a la altura (2 horas) puede tener un gran efecto sobre el pH sanguíneo y sobre el rendimiento en el ejercicio.
Sin embargo, la desviación a la izquierda en la curva de disociación de la oxihemoglobina no puede descartarse como una explicación de la mejora en el rendimiento observada por McLellan, Jacobs y Lewis (32). Gonzales, Zamagni y Clancy (18) utilizaron ratas suplementadas con bicarbonato y las expusieron a condiciones hipóxicas (370-380 Torr). Estos autores observaron un VO2 máx. en promedio 18 % mayor en los animales suplementados con bicarbonato que en los animales de control que recibieron placebo (Figura 4). Los autores atribuyeron el elevado VO2 máx. al incremento en el contenido arterial de oxígeno y hallaron una correlación casi lineal entre el contenido arterial de oxígeno y el VO2 máx. Además los autores sugirieron que el incremento en la alcalinidad sirvió para amortiguar a los iones hidrógeno producidos durante el ejercicio y para atenuar la reducción en el contenido arterial de oxígeno. Esta sugerencia parecería indicar que ambas explicaciones para los hallazgos de McLellan, Jacobs y Lewis (32) son correctas.
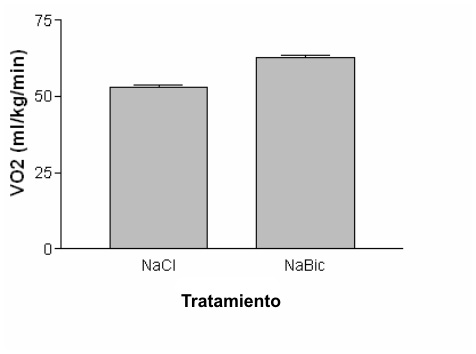
Figura 4. Los datos ilustran el efecto de la suplementación con NaHCO3 (NaBic)
sobre el VO2 máx. bajo condiciones hipóxicas en comparación con el
placebo NaCl. Los datos fueron obtenidos de Gonzales y cols. (18).
Sin embargo, otra explicación para la mejora del rendimiento observada por McLellan, Jacobs y Lewis (32), ha sido sugerida por Loeppky y cols. (29). Loeppky cols. estudiaron a perros anestesiados que fueron ventilados mecánicamente. Estos investigadores hallaron que la presión arterial pulmonar esta relacionada al nivel ácido-básico, tal que a mayor alcalinidad menor presión en la arteria pulmonar. Esta menor presión en la arteria pulmonar permitió una mejor relación ventilación-perfusión. Esto fue verificado por medio de la examinación de las diferencias entre el O2 y el CO2 del volumen corriente y el O2 y el CO2 arterial. Este hallazgo llevo a los autores a creer que la hiperventilación que ocurre en la altura puede facilitar la mejora de la SaO2 reduciendo la presión pulmonar por medio del incremento en la alcalinidad en lugar de la visión tradicional acerca de que la hiperventilación mantiene una alta presión parcial de oxígeno alveolar. Si bien estos datos proporcionan una visión diferente del efecto del nivel ácido-básico en la altura, aun sugieren que los cambios en el nivel ácido-básico luego de la exposición a la altura tendrá un efecto beneficioso sobre la SaO2 y subsecuentemente sobre el VO2 máx.
Los cambios que ocurren en el nivel ácido-básico con la exposición aguda a la altura o a la hipoxia parecen tener influencia sobre la SaO2. Aquí parece haber alguna controversia sobre el mecanismo por el cual el nivel ácido-básico influencia la SaO2. Sin tener en cuenta el incremento en la alcalinidad, el incremento en la SaO2 tiende a tener una influencia positiva sobre el VO2 máx. y por ende teóricamente sobre el rendimiento en el ejercicio.
CONCLUSION
Los primeros estudios sobre el VO2 máx. sugieren que cuanto mayor es la exposición a la altura mayor será la reducción en el VO2 máx. y en el rendimiento en el ejercicio. Aunque pueden observarse mejoras en el rendimiento y en el VO2 máx. con la aclimatación, la mejora nunca vuelve al estado del nivel del mar. Esto sugiere que para cualquiera que realice ejercicios o trabaje en la altura, lo mejor es finalizar la tarea cuanto antes, por lo que se minimizaran los efectos deletéreos de la exposición prolongada a la altura.
Sin embargo, algunas de las respuestas inmediatas a la altura o a la hipoxia incluyendo la diuresis y el incremento en la alcalinidad pueden ser en realidad beneficiosas. Se ha demostrado que estos cambios en el balance hídrico y en el nivel ácido-básico pueden ser ventajosos para la exposición a la altura y para elevar la SaO2. Los modelos de Robgers y cols. (39), Ferretti (17) y Wagner (48) sugieren que una mejor SaO2 resultará en una menor declinación del VO2 máx. De esta manera un exposición de varias horas o incluso pasar la noche en hipoxia puede en realidad mejorar el VO2 máx. más que empeorarlo, tal como se sugirió en los primeros estudios sobre el VO2 máx.
Agradecimientos
El autor desea expresar su gratitud a Robert Robgers, PhD y a David James, MD por su revisión crítica de este manuscrito.
Dirección para el envío de correspondencia
Daryl L. Parker, Dept. Kinesiology and Health Science, CSU, Sacramento 95819. Teléfono: (916) 278-6902; correo electrónico: parkerd@csus.edu
Referencias
1. Anard, IS, Chandrashekhar Y, Rao SK, Malhotra RM, Ferrar R, Chandana J, et al (1993). Body fluid compartments, renal blood flow, and hormones at 6000m in normal subjects. J Appl Physiol 1993;74(3):1234-1239
2. nderson, HT, Smeland EB, Owe JO, Myhre K (1985). Analysis of maximum cardiopulmonary performance during exposure to acute hypoxia at simulated altitude sea-level to 5,000m (760 404 mmHg). Aviat Space Environ Med 1985;56: 1192-1197
3. Balke B, Nagle FJ, Daniels J (1965). Altitude and maximum performance in work and sports activity. JAMA 1965;194(6): 646-649
4. Bartsch P, Shaw S, Franciolli M, Gnadinger MP, Weidmann P (1988). Atrial natriuretic peptide in acute mountain sickness. J Appl Physiol 1988;65(5):1929-1937
5. Bartsch PN, Pfluger N, Audetat M, Shaw S, Weidmann P, Vock P, et al (1991). Effects of slow ascent to 4559m on fluid homeostasis. Aviat Space Environ Med 1991;62: 105-110
6. Bartsch, PN, Shaw S, Weidmann P, Olez O (1993). Does drinking protect against mountain sickness?. Schweiz Z Sportmed 1993;41(1): 7-13
7. Bouissou P, Peronnet F, Brisson G, Helie R, Ledoux M (1986). Metabolic and endocrine responses to graded exercise under acute hypoxia. Eur J Appl Physiol 1986;55:290-294
8. Buskirk, ER, Kollias J, Akers RF, Prokop EK, Reategui EP (1967). Maximal performance at altitude and on return from altitude in conditioned runners. J Appl Physiol 1967;23(2):259-266.
9. Consolozio CF, Nelson RA, Matoush LO, Hanson JE (1966). Energy metabolism at high altitude. J Appl Physiol 1966;21(6):1732-1740
10. Cymerman A, Reeves JT, Sutton JR, Rock PB, Groves BM, Malconian MK (1989). Operation Everast II: maximal oxygen uptake at extreme altitude. J Appl Physiol 1989;66(5):2446-2453
11. Daniels, J (1979). Altitude and athletic training and performance. Amer J Sports Med 1979;7: 370-373
12. Dill DB, Hillyard SD, Miller J (1980). Vital capacity, exercise performance, and blood gases at altitude as related to age. J Appl Physiol: Respirat Environ Exercise Physiol 1980;48(1):6-9
13. Dill, DB, Myhre LG, Phillips EE, Brown DK (1966). Work capacity in acute exposures to altitude. J Appl Physiol 1966;21(4):1168-1176
14. Eldridge FL, Millhorn DE, Kiley JP (1985). Antagonism by theophylline of respiratory inhibition induced by adensoine. J Appl Physiol 1985;59(5): 1428-1433
15. Elliot PR, Atterbom HA (1978). Comparison of exercise responses of males and females during acute exposure to hypobaria. Aviat Space Environ Med 1978;49(2): 415-418
16. Faulkner JA, Daniels JT, Balke B (1967). Effects of training at moderate altitude on physical performance capacity. J Appl Physiol. 1967;23(1):85-89
17. Ferretti G (1990). On maximal oxygen consumption in hypoxic humans. Experientia 1990;46:1188-1194
18. Gonzalez NC, Zagagi M, Clancy RL (1991). Effect of alkalosis on maximum oxygen uptake in rats acclimated to simulted altitude. J Appl Physiol 1991;71(3):1050-1056
19. Grover RF, Selland MA, McCullough RG, Dahans TE, Wolfel EE, Butterfield GE (1998). Beta-adrenergic blockade does not prevent polycythemia or decrease in plasma volume in men at 4300m altitude. Eur J Appl Physiol 1998;77(3):264-270
20. Gunga HC, Kirsch K, Baartz F, Steiner HJ, Wittels P, Rocker L (1995). Fluid distribution and tissue thickness changes in 29 min during 1 week at moderate altitude (2,315m). Eur J Appl Physiol, 1995;70(1):1-5
21. Guyton AC, Hall JE (2000). Textbook of Medical Physiology 10th Ed. New York: W.B. Saunders Company
22. Hackett PH, Rennie D, Grover RF, Reeves JT (1981). Acute mountain sickness and the edemas of high altitude: A common pathogenesis?. Resp Physiol 1981;46:383-390
23. Hopkins SR, McKenzie, DC (1989). Hypoxic ventilatory response and arterial desaturation during heavy work. J Appl Physiol 1989;67(3): 1119-1124
24. Hoyt, RW,. Durkot MJ, Forte VA, Hubbard LJ, Trad LA, Cymerman, A (1991). Hypobaric hypoxia (380m Torr) decreases intracellular and total body water in goats. J Appl Physiol 1991;71(2):509-513.
25. Jain, SC, Bardham J, Swamy YV, Krishna B, Nayar HS (1980). Body fluid compartments in humans during acute high altitude exposure. Aviat Space Environ Med 1980;51(3):234-236
26. Koller, EA, Buhrer A, Felder L, Schopen M, Vallotton MB (1991). Altitude diuresis: Endocrine and renal responses to acute hypoxia of acclimatized and non-acclimatized subjects. Eur J Appl Physiol 1991;62: 228-234
27. Lawler J, Powers SK, Thompson D (1998). Linear relationship between VO2max and VO2max decrement during exposure to acute hypoxia. J Appl Physiol 1988;64(4): 1486-1492
28. Levitan BM, Bungo MW (1982). Measurement of cardiopulmonary performance during acute exposure to a 2440m equivalent atmosphere. Aviat Space Environ Med 1982;53(7):639-642.
29. Loeppky JA, Scotto P, Riedel CE, Roach RC, Chick TW (1992). Effects of acid-base status on acute hypoxic pulmonary vasoconstriction and gas exchange. J Appl Physiol 1992;72(5):1787-1797.
30. Maresh CM, Kraemer WJ, Noble BJ, Robertson KL (1988). Exercise responses after short and long-term residence at 2,200m. Aviat Space Environ Med 1988;59: 333-339
31. McLellan T, Jacobs I, West W (1988). Acute altitude exposure and altered acid-base states II. Effects on exercise performance and muscle and blood lactate. Eur J Appl Physiol 1988;57: 445-451
32. Milledge, JS (1979). Acid-base changes associated with respiratory acclimatization to altitude. Postgrad Med J 1979;55:468-470
33. Milledge (1992). JS Salt and water control at altitude. Int J Sports Med 1992;13(suppl 1): S61-S63
34. Milledge, JS, Beeley, JM, Broome J, Luff N, Pelling M, Smith D (1991). Acute mountain sickness susceptibility, fitness and hypoxic ventilatory response. Eur Respir J 1991;4(8): 1000-1003
35. Paterson, DJ, Pinnington H, Pearce AR, Morton AR (1987). Maximal exercise cardiorespiratory responses of men and women during acute exposure to hypoxia. Aviat Space Environ Med 1987;58(3): 243-247
36. Ponchia A, Noventa D, Zaccaria M, Bertaglin M, Pocher GO, Miraglia G (1995). Body fluids, atrial volumes, and atiral natriuetic peptide during and after high-altitude exposure. Wilder Environ Med 1995;6: 11-19
37. Richalet JP, Rathat C, Keromes A, Herry JP, Larmignat P, Garnier M (1983). Plasma volume, body weight, and acute mountain sickness. Lancet; (March): 525
38. Robergs RA, Quintana R, Parker DL, Frankel CC (1998). Multiple variables explain the variability in the decrement in VO2max during acute hypobaric hypoxia. Med Sci Sports Exerc 1998;30(6): 869-879
39. Robertson RJ, Gilcher R, Metz KF, Skrinar GS, Allison TG, Bahnson HT (1982). Effect of induced erythrocythemia on hypoxia tolerance during physical exercise. J Appl Physiol: Respirat Environ Exercise Physiol 1982;53(2): 490-495
40. Shepard RJ, Bouhlel E, Vanderwalle H, Monod H (1988). Muscle mass as a factor limiting physical work. J Appl Physiol 1988;64(4): 1472-1479
41. Shepard RJ, Bouhlel E, Vanderwalle H, Monod H (1988). Peak oxygen intake and hypoxia: influence of physical fitness. Int J Sports Med 1988;9: 279-283
42. Shirlo C, Bub A, Reize C, Buhrer A, Kohl J, Koller EA (1997). Volume changes in the forearm and lower limb during 2h of acute hypobaric hypoxia in non-acclimatized subjects. Eur J Appl Physiol 1997;75: 124-131
43. Singh MV, Raul SB, Tyagi AK (1990). Body fluid status on induction, reinduction and prolonged stay at altitude of human volunteers. Int J Biometerol 1990;34: 93-97
44. Squires RW, Buskirk ER (1982). Aerobic capacity during acute exposure to simulated altitude, 914 to 2286 meters. Med Sci Sports Exerc 1982;14(1): 36-40
45. Sutton JR, Reeves JT, Wagner PD, Groves BM, Cymerman A, Malconian MK, et al (1988). Operation Everest II: oxygen transport during exercise at extreme simulated altitude. J Appl Physiol 1988;64(4): 1309-1321
46. Tucker A., Stager JM, Cordain L (1984). Arterial O2 saturation and maximum O2 consumption in moderate-altitude runners exposed to sea-level and 3,050m. JAMA 1984;252(20): 2867-2871
47. Wagner, PD (2000). New ideas on limitations to VO2max. Exerc Sport Sci Rev 2000;28(1): 10-14
48. West, JB (1998). High Life: A history of high-altitude physiology and medicine. American Physiological Society, Oxford
49. Westendorp RG, Roos AN, van der Hoeven HG, Tjiong MY, Simons R, Frolich M. (1993). Atrial natriuretic peptide improves pulmonary gas exchange in subjects exposed to hypoxia. Am Rev Respir Dis;148(2): 304-309
50. Young AJ, Cymerman A, Burse RL (1985). The influence of cardiorespiratory fitness on the decrement in maximal aerobic power at high altitude. Eur J Appl Physiol
51. Young AJ, Sawka MN, Muza SR, Boushel R, Lyons T, Rock PB (1996). Effects of erythrocyte infusion on VO2max at high altitude. J Appl Physiol 1996;81(1): 252-259