
Efectos de la Recuperación Activa vs. la Recuperación Pasiva sobre la Producción de Potencia durante Series Repetidas de Ejercicio de Alta Intensidad y de Corta Duración
Declan A Connolly1, Kevin M Brennan1 y Christie D Lauzon1
Human Performance Laboratory, Room 212, Patrick Gymnasium, University of Vermont Burlington, Estados Unidos.
Artículo publicado en el journal PubliCE del año 2003.
Publicado 24 de marzo de 2011
Resumen
Palabras clave: potencia anaeróbica, ejercicio suave, lactato, producción de potencia
INTRODUCCION
La ventaja de una recuperación activa versus una recuperación pasiva sobre el rendimiento subsiguiente en ejercicios de alta intensidad y corta duración ha sido bien documentado (Ainsworth et al., 1993; Stanley et al., 1988). Además, se ha sugerido que el trabajo de baja intensidad de aproximadamente 20-40 minutos es apropiado para evitar la reducción en la producción de potencia en series repetidas de ejercicio de alta intensidad y corta duración (Bangsbo, 1994). El ejercicio de alta intensidad resulta en un incremento en los niveles de lactato, tanto intramusculares como circulantes (McLoughlin et al., 1991; Rowell et al., 1986). Además se ha demostrado, que estos incrementos en las concentraciones de lactato, que a su vez reflejan la concentración de iones hidrógeno, inhiben el rendimiento contráctil y provocan fatiga prematura. La remoción del lactato puede producirse a través de varios órganos , aun cuando los estudios llevados a cabo con trazadores que han utilizado lactato marcado han mostrado que una proporción significativa del lactato es absorbida por el músculo esquelético y subsiguientemente metabolizada a través de la reconversión en piruvato y la entrada en el ciclo de Krebs (Brookes, 1986). De esta manera, es aparente que la remoción del lactato es beneficiosa en términos del mantenimiento de los niveles de rendimiento, y en la literatura se han reportado numerosos y variados protocolos que son efectivos para reducir los niveles de lactato.
Los estudios en este tópico han utilizado períodos de recuperación en el rango de los 30 segundos a 40 minutos de duración (Bangsbo et al., 1994) y la mayoría de las investigaciones parece haber utilizado períodos de recuperación mayores a 5 minutos. Sin embargo, la literatura es equívoca con respecto a si la reducción de los niveles de lactato bajo ciertas circunstancias resulta en una mejora del rendimiento (Ainsworth et al., 1993; Bangsbo et al., 1994; Rowell et al., 1986; Stanley et al., 1988a). Con frecuencia los atletas entrenan, particularmente para mejorar la potencia y la velocidad, con intervalos de recuperación que son mucho más cortos que 5 minutos. Asimismo, los atetas con frecuencia entrenan anaeróbicamente hasta casi el agotamiento durante períodos cortos de entre 15-30 segundos. Teóricamente, esto se relación con la liberación y reposición del trifosfato de adenosina (ATP). Interesantemente, pocos estudios parecen haber considerado los efectos del modo de recuperación sobre la producción de potencia en relación a la utilización del ATP y el tiempo necesario para su reposición. Luego de un ejercicio agotador, las reservas de trifosfato de adenosina se reponen en aproximadamente un 90-95% en tres minutos. Esta reposición es crucial para la realización de trabajos de corta duración y alta intensidad (Signorile et al., 1993). Para nuestro conocimiento, pocos estudios parecen haber utilizado específicamente ese intervalo de tres minutos en la investigación acerca de los modos de recuperación y su influencia sobre la remoción de lactato y más importante aún sobre la producción de potencia. Sería interesante investigar este período de recuperación de 3 minutos particularmente para determinar si la recuperación activa puede remover lactato en tan poco tiempo. Ratel et al. (2002) examinaron los efectos de la duración de la recuperación sobre la producción pico de potencia luego de la repetición de esprints de 10 s realizados en un cicloergómetro y hallaron que en los hombres, los incrementos en las concentraciones de lactato se reducían con un período de recuperación pasiva de 5 minutos en comparación con un período de recuperación de 30 s o 1 min. Asimismo, Hebestreit et al. (1993) en su investigación acerca de la producción de potencia muscular luego de sesiones de recuperación pasiva de 1, 2 y 10 minutos subsiguientes a la realización de Tests de Wingate de Potencia Anaeróbica, hallaron que aun luego de 10 minutos de recuperación pasiva los sujetos no habían retornado a los niveles basales de potencia. Signorile et al. (1993) reportaron incrementos significativos en el pico de potencia y en el trabajo total realizado durante 30 segundos luego de una recuperación activa y en comparación con la recuperación pasiva. Desafortunadamente, no se reportaron los niveles de lactato pro lo que no se sabe si el corto período de recuperación facilitó el incremento en el clearance de lactato.
Bond et al. (1991) hallaron que, luego de 20 minutos de recuperación activa o pasiva subsiguiente a la realización de una serie supramáxima de trabajo de 60 segundos de duración, los valores del torque pico isocinético no eran significativamente diferentes entre los modos de recuperación activa o pasiva, pero que los valores de lactato eran significativamente menores luego de 20 min de recuperación activa en comparación con la recuperación pasiva. Estos hallazgos derivan en interesantes cuestiones acerca de la tasa de reconversión metabólica, ya que se relaciona con el modo de recuperación a corto plazo. Por lo tanto, el propósito de este estudio fue investigar los cambios en el pico de potencia, en la potencia promedio y en la acumulación de lactato durante series repetidas de ciclismo de alta intensidad y corta duración utilizando modos de recuperación activa y pasiva de 3 minutos de duración
METODOS
Sujetos
Siete ciclistas saludables de sexo masculino (edad 21.8±3 años, masa 73.0±3.8 kg, talla 177.3±3.4 cm) fueron voluntarios para participar en este estudio. Todos los sujetos completaron un cuestionario acerca de su historial de salud y firmaron un consentimiento informado antes de su participación. Todos los sujetos eran ciclistas recreacionalmente activos, pero no estaban altamente entrenados. Todos los procedimientos fueron aprobados pro el Comité de Revisión para la Utilización de Sujetos Humanos de la Universidad de Vermont antes de que se realizaran las evaluaciones.
Protocolo Experimental
Todos los sujetos realizaron un protocolo de recuperación activa y pasiva, y por lo tanto sirvieron como sus propios controles. Cada prueba estuvo separada por un período de al menos siete días. El orden de las evaluaciones fue aleatorio y ambos protocolos fueron idénticos excepto en lo concerniente al procedimiento de recuperación. Durante la recuperación pasiva los sujetos se quedaron quietos sobre el cicloergómetro durante dos minutos y cincuenta segundos. Durante los diez segundos previos a la fase de trabajo los sujetos pedalearon suavemente. Durante la recuperación activa, los sujetos pedalearon a 80 rpm con una resistencia de 1 kg durante los 3 minutos (Ainsworth et al., 1993; Stanley et al., 1988). El protocolo completo del test anaeróbico fue el siguiente: Los sujetos realizaron una entrada en calor de 3 minutos con una carga de 1 kg a 80 rpm. Luego de esto los sujetos realizaron seis sprints máximos de quince segundos sobre el cicloergómetro. Hubo 3 minutos de recuperación entre cada serie de trabajo. La serie de trabajo requirió que todos los sujetos pedalearan contra una carga estándar de 5.5 kg de acuerdo con los estudios previos relevantes (Ainsworth et al., 1993). Las cargas de trabajo también fueron estandarizadas debido a la gran variación en las cargas de trabajo previamente reportadas para lograr un clearance del lactato óptimo. Nosotros pensamos que la utilización de este protocolo facilitaría la comparación con algunos trabajos previos. Los procedimientos del Wingate fueron administrados en concordancia con los descriptos por Lakomy (1986). Se les dio a los sujetos el comando de 3, 2, 1, Ya! Todas las pruebas comenzaron con los sujetos en posición estacionaria para que todas las series fueran consistentes. Los sujetos fueron estimulados verbalmente para que pedalearan lo más rápido y duro que pudieran hasta que se les diera la orden de detenerse. No se les proporcionó a los sujetos retroalimentación acerca de las diferencias de rendimiento entre los protocolos o entre las pruebas. Luego de la finalización la carga fue ajustada para realizar el modo de recuperación. Este procedimiento fue repetido para las seis series. Todos los datos anaeróbicos fueron recolectados utilizando un cicloergómetro Monark (modelo 864) y todos los datos sobre la potencia fueron registrados utilizando el programa Sports Medicine Industries Power software V3.02 (SMI, St. Cloud, MN, Estados Unidos). Esto permitió la determinación del pico de potencia (PP), de la potencia promedio (AP), del tiempo hasta alcanzar el pico (TPP), y del índice de fatiga expresado como un porcentaje a partir del PP y del PP más bajo, característicamente observado como el último punto de los datos.
Recolección de las Muestras de Sangre
Todas las muestras de sangre fueron recolectadas a través del método estándar de punción de la yema de los dedos. En cada muestreo se recolectaron 30 µl. Las muestras fueron analizadas inmediatamente como sangre total y en duplicado utilizando un analizador de lactato portátil Accusport (Boehringer Mannheim, Indianapolis, IN, Estados Unidos). Por cada test se extrajeron siete muestras de sangre. Una muestra fue extraída 2 minutos después de cada serie de trabajo de 15 segundos y una muestra adicional fue extraída 5 minutos después de la última serie. No se controló la dieta de ninguno de los sujetos antes de la evaluación, no obstante se les pidió que consumieran las mismas comidas en los días de las evaluaciones.
Análisis Estadísticos
Los datos fueron analizados utilizando el análisis de varianza para mediciones repetidas de 2 x 6 (SPSS, V11.). Se estableció un nivel alfa a p<0.05 para todos los análisis.
RESULTADOS
Los datos del lactato son reportados en mmol/L. Los datos no mostraron diferencias significativas en la media de potencia pico entre los modos de recuperación a través de las seis series, 775±11.2 Watts versus 772±33.4 Watts, para los protocolos de recuperación activa y pasiva, respectivamente (p=0.785, F 0.08). No se observaron diferencias en la potencia promedio, 671±26.4 Watts versus 664±10.0 Watts, entre la recuperación activa y pasiva respectivamente. La Figura 1 muestra los valores de potencia pico y promedio dentro y entre las pruebas.
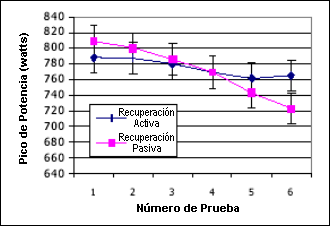
Figura 1. Pico de potencia para las pruebas con recuperación activa y pasiva.
Como uno podría esperar se observaron diferencias significativas dentro de cada prueba tanto para el pico de potencia como para la potencia promedio (p<0.004, F 4.47) con ambos valores reduciéndose con el tiempo. Con el protocolo de recuperación pasiva se observó una reducción significativamente mayor (p<0.002, F 4.78). La potencia promedio fue significativamente diferente entre las series, y se redujo con el tiempo (p<0.008) pero no fue diferente entre los protocolos (p=0.57). Estos cambios en la producción de potencia no parecen poder explicarse a través de los cambios en los valores del lactato ya que las concentraciones del lactato no fueron significativamente diferentes entre los protocolos (p=0.37), 9.09±2.37 mmol/L versus 10.05±2.84 mmol/L, para las recuperaciones activa y pasiva respectivamente. Sin embargo, los valores fueron diferentes dentro de cada protocolo entre las series ya que los valores de lactato se incrementaron con el tiempo. Los datos del lactato se presentan en la Figura 2.
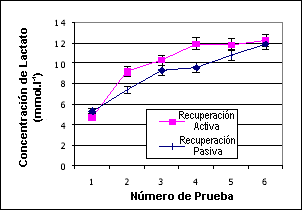
Figura 2. Datos del lactato para las pruebas con recuperación activa y pasiva
DISCUSION
El presente estudio estuvo enfocado en un período de recuperación corto (3 minutos), un protocolo que para nuestro conocimiento no ha sido muy ampliamente evaluado, aunque es ampliamente practicado por los atletas. En concordancia con los hallazgos de Bangsbo et al. (1994), nosotros no hallamos diferencias estadísticamente significativas en la concentración de lactato sanguíneo entre los protocolos de recuperación activa y pasiva con períodos de hasta 3 minutos. Sin embargo, los mismos autores han hallado que cuando el período de recuperación se extiende más allá de los tres minutos hay un mayor metabolismo del lactato dentro de los músculos activos durante la recuperación. Un mecanismo sugerido para esta reducción en la concentración de lactato es la distribución del lactato circulante a sitios del metabolismo tales como el hígado, el corazón y músculos previamente inactivos (Belcastro y Bonen, 1975). Sin embargo, otros investigadores han sugerido que el lactato es absorbido y oxidado por los músculos esqueléticos que realizan una actividad suave a moderada durante los períodos de recuperación (Brookes, 1986; Thiriet et al., 1993). Por mucho tiempo se ha reconocido que el lactato es un metabolito que se acumula durante el ejercicio y contribuye a la fatiga muscular (Gregg et al., 1984; Hetenyi et al., 1983; Jorfeldt, 1970), y que durante la realización de ejercicios el aumento en la concentración de epinefrina provoca un incremento en la acumulación de lactato (Roth y Brooks, 1990; Stanley y Lehman, 1988). Está también bien establecido que luego de la realización de ejercicios intensos, una recuperación de baja intensidad resulta en la reducción de la concentración del lactato circulante (Dodd et al., 1984; Hetenyi et al., 1983).
Los estudios que han examinado los períodos de recuperación y la reducción en el lactato circulante son equívocos (Bangsbo et al., 1994). Bangsbo et al (1994) examinaron la concentración de lactato en muestras de biopsias musculares tomadas de músculos activos e inactivos durante la recuperación y hallaron que la concentración de lactato era similar dentro de los 10 minutos de la recuperación y que las concentraciones arteriales y venosas de lactato mostraban una tendencia similar durante el mismo período de tiempo. Otros estudios han demostrado que el ejercicio de baja intensidad tiene un mínimo efecto sobre el lactato sanguíneo dentro de los 15-20 minutos de la recuperación (Dodd et al., 1984; Hermansen y Stensvold, 1972).
Se ha promovido la recuperación activa, debido a que se piensa que ayuda a la remoción del lactato luego de la realización de ejercicios intensos, y que por lo tanto disminuye la reducción en el rendimiento de las series subsiguientes. Los resultados de este estudio han mostrado diferencias en la producción de potencia en función del modo de recuperación, pero estas diferencias no son fácilmente explicables por medio de los valores del lactato. Nuestros datos han mostrado un patrón consistente y esperado en la disminución de la potencia a medida que los sujetos progresaban desde la primera a la sexta serie. El patrón de nuestros datos acerca del lactato intra prueba también fueron esperados, observándose un incremento en la concentración de lactato a medida que progresaban las series. Sin embargo, nosotros consideramos que deberíamos observar una diferencia en el patrón del lactato entre nuestros protocolos. Sin embargo, dicha diferencia no fue observada. La concentración de lactato circulante es también una función de la intensidad del ejercicio. Está bien establecido que el potencial para la producción de lactato es altamente dependiente de la tasa del flujo glucogenolítico / glucolítico, y es la intensidad del ejercicio la que determina la tasa de flujo en estas vías metabólicas. En el presente estudio se utilizó una resistencia fija de 5.5 kg para cada sujeto independientemente de su masa corporal. La principal razón de esto fue que en la mayoría de los trabajos más relevantes previamente citados se utilizó esta carga de trabajo, lo cual nos permite realizar comparaciones. Además, nuestros valores de masa corporal están dentro de un pequeño rango, con desviaciones estándar relativamente pequeñas, lo cual nos permite disminuir nuestra preocupación sobre esta limitación.
Durante la realización de series de ejercicio de alta intensidad y corta duración los músculos esqueléticos se vuelven grandes productores de lactato y consecuentemente se enlentece el clearance de lactato. Luego, durante la recuperación hay una transición hacia una absorción neta de lactato sanguíneo por los músculos esqueléticos previamente inactivos. El mecanismo responsable de este flujo de lactato durante los primeros minutos del ejercicio es poco claro. Sin embargo, parece que los músculos responsables de incremento inicial en la concentración de lactato durante la realización de ejercicios intensos requieren de un período de recuperación mayor a los tres minutos para modificar el metabolismo intracelular del lactato lo cual resulta en un gradiente de lactato a favor de la absorción por el mismo músculo. La explicación del gradiente de lactato es incorporada en la hipótesis del shuttle o transporte de lactato (Brooks, 1986). Por lo tanto, ¿pueden los niveles de lactato explicar cualquier variación en la producción de potencia?. Nosotros no hallamos diferencias estadísticas en la acumulación de lactato entre las pruebas. Aunque se observaron diferencias significativas, tanto en la potencia promedio como en el pico de potencia dentro y entre las pruebas, nosotros no podemos explicar estas diferencias utilizando las concentraciones de lactato. Esto es interesante, especialmente en relación al pico de potencia, ya que uno podría esperar que mayores niveles de lactato inhibieran una alta producción de potencia. Es aparente que con la recuperación activa el flujo de sangre hacia los músculos se mantiene o se incrementa. Sin embargo, se debería señalar que en el presente estudio no se midieron las concentraciones intracelulares de lactato. De esta manera, aunque las concentraciones de lactato plasmático fueron similares con ambos protocolos de recuperación, el incrementado flujo sanguíneo durante la recuperación activa pudo haber permitido una reducción en la concentración intracelular de lactato sin una reducción concomitante en la concentración plasmática de lactato. Nuestros métodos no estuvieron dirigidos a este punto. Es bastante probable que el flujo sanguíneo desempeñe un papel clave en la reposición del ATP, quizás a través de la resíntesis de fosfato de creatina, derivando en una menor reducción en la producción de potencia. Este no fue una de nuestros objetivos y nuevamente nuestros métodos no estuvieron dirigidos a este punto. Un incrementado metabolismo aeróbico que facilitara el suministro de energía también pudo haber contribuido al mantenimiento de la producción de potencia.
Conclusiones
Los resultados de este estudio sugieren que no parece haber un efecto significativo, en términos de clearance de lactato, de la utilización de períodos de recuperación activa y en comparación con la recuperación pasiva, entre series de ejercicio de alta intensidad y corta duración. Si bien nuestros hallazgos parecen sumarse a la ambigüedad de los hallazgos ya existentes, la naturaleza de nuestro protocolo debería ser considerada. Nosotros no conocemos ningún estudio que haya utilizado este período de recuperación de tres minutos y esto puede explicar cualquier variación en los hallazgos.
Uno debería señalar que una comparación de la producción de potencia entre la primera y la última serie de ejercicio mostró que el protocolo de recuperación pasiva provocó una producción de potencia 2.5% mayor luego de la serie uno y el protocolo de recuperación activa provocó una producción de potencia 5.5% mayor luego de la serie seis. Además se debería considerar que la reducción en la producción de potencia desde la serie uno a la seis durante el protocolo de recuperación pasiva fue del 10.6% en tanto que solo fue del 2.9% para el protocolo de recuperación activa en el mismo período de tiempo. Estos números son hallazgos substanciales en términos, tanto de producción de potencia como de sus implicaciones para el rendimiento. Por lo tanto, si bien la recuperación activa provee ciertos beneficios en términos de incremento de la potencia, nosotros no podemos explicar esto utilizando como fundamento la reducción en la concentración plasmática de lactato. El mecanismo por el cual se produce la mejora en la producción de potencia con la recuperación activa requiere de investigaciones adicionales.
Referencias
1. Ainsworth, B.E., Serfass, R.C., and Leon A.S (1993). Effects of recovery duration and blood lactate level on power output during cycling. Canadian Journal of Applied Physiology 18, 19-30
2. Bangsbo, J., Graham, T., Johansen, L., and Saltin B (1895). Muscle lactate metabolism in recovery from intense exhaustive exercise: impact of light exercise. Journal of Applied Physiology 77
3. Belcastro, A.N., and Bonen A (1975). Lactic acid removal rates during controlled and uncontrolled recovery exercise. es during controlled and uncontrolled recovery exercise. Journal of Applied Physiology 39, 932- 936
4. Bond, V., Adams, R.G., Tearney R.J., Gresham K., and Ruff, W (1991). Effects of active and passive recovery on lactate removal and subsequent isokinetic muscle function. Journal of Sports Medicine and Physical Fitness 31, 357-361
5. Brooks, G.A (1986). The lactate shuttle during exercise and recovery. Medicine and Science in Sports and Exercise 18, 360-368
6. Dodd, S., Powers, S.K. Callender, T. and Brooks, E (1984). Blood lactate disappearance at various intensities of recovery exercise. Journal of Applied 57, 1462-1465
7. Gregg, S.G., Mazzeo, R.S. Budinger, T.F. and Brooks, G.F (1984). Acute anemia increases lactate production and decreases lactate clearance during exercise. Journal of Applied Physiology 67, 756 764
8. Hebestreit, H., Mimura, K., and Bar-Or, O (1993). Recovery of muscle power after high intensity short-term exercise: comparing boys and men. Journal of Applied Physiology 74, 2875-2880
9. Hermansen, L., and Stensvold, I (1972). Production and removal of lactate during exercise in man. Acta. physiologica Scandinavica. 86, 191-201
10. Hetenyi, G., Perez, G., and Vranic, M (1983). Turnover and precursor-product relationships of nonlipid metabolites. Physiology Review 63, 606-667
11. Jorfeldt, T (1970). Metabolism of L(+)-lactate in human skeletal muscle during exercise. Acta. Physiologica Scandinavica. 388 (suppl.) 55-67
12. Lakomy, H. L (1986). Measurement of work and power output using friction loaded cycle ergometers. Ergonomics 29, 509-517
13. McLoughlin, P., McCaffrey, N. and Moynihan, J.B (1991). Gentle exercise with a previously inactive muscle group hastens the decline of blood lactate concentration after strenuous exercise. European Journal of Applied Physiology 62, 274-278
14. Ratel, S., Bedu, M., Hennegrave, A., Dore, E., and Duche, P (2002). Effects of Age and Recovery Duration on Peak Power Output During Repeated Cycling Sprints. International Journal of Sports Medicine 23, 397-402
15. Roth, D.A., and Brooks, G.A (1990). Lactate transport is mediated by a membrane bound carrier in rat skeletal muscle sarcolemmal vesicles. Archives of Biochemistry & Biophysics. 279, 377-385
16. Rowell, I., Saltin, B.R., Kiens, B. and Christenson, N.J (1986). Is peak quadriceps blood flow in humans even higher during exercise with hypoxemia?. Heart Circulation and Physiology 20, H1038- H1044
17. Signorile, J. F., Ingalls, C., and Tremblay, L.M (1993). The effects of active and passive recovery on short term, high intensity power output. Canadian Journal of Applied Physiology 18, 31-42
18. Stanley, W.C., Wisneski, J.A., E.W. Gertz, E.W., Neese, R.A. and Brooks, G.A (1988). Glucose and lactate interrelations during moderate intensity exercise in man. Metabolism 37, 850-858
19. Stanley, W.C., and Lehman, S.L (1988). A model for measurement of lactate disappearance with isotope tracers in the steady s tate. The BiochemicalJournal 256, 1035-1038
20. Thiriet P., Gozal, D., Wouassi, D., Oumarou, T., Gelas, H., and Lacour, J.L (1993). The effect of various recovery modalities on subsequent performance, in consecutive supramaximal exercise. Journal of Sports Medicine and Physical Fitness 33, 118-129