
Las catecolaminas y su rol en el ejercicio intermitente.
Publicado 11 de noviembre de 2013, 14:40
Ya en el estudio de Tremblay y cols. de 1994 se han indicado que la capacidad del músculo activo para utilizar los AGL, a pesar de los niveles de lactato altos, es debida a la naturaleza intermitente del ejercicio intermitente de alta intensidad. Por ejemplo, se ha demostrado que durante el componente intensivo del ejercicio intermitente de alta intensidad, el ATP y el fosfato de creatina es degradado para producir energía y es resintetizado durante los períodos de descanso no intensivos, vía el sistema aeróbico (según Tremblay, en otro estudio de 1990). Se sabe que con períodos de descanso cortos (12 segundos, por ejemplo), es probable que esta resíntesis esté incompleta; así la glucólisis anaeróbica se ha sugerido de proveer el resto de la energía requerida.
El glucógeno es el sustrato para el sistema glucolítico anaeróbico y se ha demostrado que se depleta hasta algún punto durante el ejercicio intermitente de alta intensidad. La investigación que usa la biopsia muscular ha demostrado que la recuperación de la fosfocreatina era incompleta después de 15 segundos de pausa en un turno de ejercicio intermitente de 15 segundos de trabajo y 15 segundos de pausa (Tremblay, 1994). Esos autores indican que el vaciamiento progresivo de la fosfocreatina y el glucógeno, junto con el mayor citrato citosólico, actúan para inhibir a la glucólisis y llevar a una menor acumulación de lactato y una mejor oxidación de ácidos grasos. El uso del oxígeno de la mioglobina durante los turnos de trabajo se ha estimado que contribuye al 44% del déficit de oxígeno y se ha demostrado que se resatura durante los períodos de descanso. Astrand y cols. han indicado que la mioglobina juega un rol contribuyente como reserva de oxígeno apoyando el metabolismo aeróbico y, probablemente, el uso de AGL como sustrato. La fuente de estos AGL es indeterminada, pero se ha indicado que las reservas de triglicéridos intramusculares (IMTG) proveen algo de sustrato.
Otras posibles fuentes de AGL son los AGL en plasma, triglicéridos, lipoproteína de baja-densidad, lipoproteína de muy-baja-densidad, quilomicrones, y adipocitos en plasma.
Asi, Las principales influencias hormonales sobre la glucogenólisis del músculo son las catecolaminas, mientras la lipólisis periférica (del tejido adiposo) es regulada por los cambios tanto de la insulina como de las catecolaminas (Chastiosis 1983).
En el ejercicio intenso agudo, la mayor secreción de catecolaminas resulta de una estimulación directa de los centros de conducción del cerebro hacia las glándulas suprarrenales. Esto explica los cambios repentinos al inicio del ejercicio, incluso antes.
Factores que afectan la magnitud de la respuesta hormonal al ejercicio (Hansen, 2012):
- *Cafeína
- *Familiarización con el modo del ejercicio
- *Postura al realizar el ejercicio
- *Parte del cuerpo activada (tren superior o tren inferior)
- *Duración e intensidad del ejercicio
La ingesta de cafeína genera un nivel sanguíneo de catecolaminas mayor durante el ejercicio; la familiarización al modo de entrenamiento o de prueba, baja el nivel sanguíneo de catecolaminas cuando los sujetos están acostumbrados al modo de entrenarse o evauarse; la postura tiene que ver con un mayor aumento en el contenido de catecolaminas sanguíneas al ejercitarse parado que recostado; el ejercicio de tren superior o el ejercicio de tren inferior provoca un mayor aumento en el contenido de catecolaminas sanguíneas al ejercitarse con los brazos vs las piernas; la duración del ejercicio produce un mayor aumento en el contenido de catecolaminas sanguíneas al ejercitarse durante un período prolongado; y la intensidad produce un mayor aumento en el contenido de catecolaminas sanguíneas al ejercitarse a una intensidad superior.
La respuesta de las catecolaminas a protocolos de ejercicio intermitente es típica del ejercicio de alta intensidad, ya que los niveles de post-ejercicio son significativamente superiores que los valores de pre-ejercicio.
Para graficar esto, podemos citar algunos estudios. Un estudio de M.A. Christmass, (1999), reclutó 7 sujetos activos (24 años; 180 cm altura; 73 kg PC; 56 ml·min-1 VO2máx, (valores promedio) realizaron 2 protocolos de carrera intermitente en cinta ergométrica. Tenían una similar velocidad de carrera (17.5-22.0 km·h-1 igual al 109±5% del VO2pico) y una similar duración (40 minutos) e igual relación trabajo y pausa 1:1,5. La Velocidad se ajustaba para elicitar un RQ de 0.96±0.99. Hubo un protocolo intermitente 'Largo' (IL, intermitente largo: 24”:36“) y otro Corto (IC, intermitente corto: 6”:9”). Ambos protocolos fueron realizados con al menos 5 días de diferencia.
La respuesta de las catecolaminas se puede observar en la Figura 1. No hubo ninguna diferencia en reposo en la concentración de la noradrenalina en plasma venosa en el intervalo largo [3.6 ±0.2 NM] e intervalo corto [3.1±0.3 NM]. Los niveles de la noradrenalina aumentaron del reposo en ambos protocolos (P <0.01, intervalo largo; P <0.05, intervalo corto), aunque la respuesta al ejercicio no fue diferente [11.2±1.5) NM, intervalo largo; 11.0±1.5) nM, intervalo corto]. La adrenalina en plasma en reposo no fue diferente en cualquiera de los protocolos [1.1±0.2) NM, largo; 1.3±0.2) NM, corto]. No hubo ningún cambio en la adrenalina desde el nivel de reposo en el protocolo largo o corto, y no hubo ninguna diferencia, por consiguiente, entre los protocolos para la respuesta de la adrenalina al ejercicio (Fig. 1).
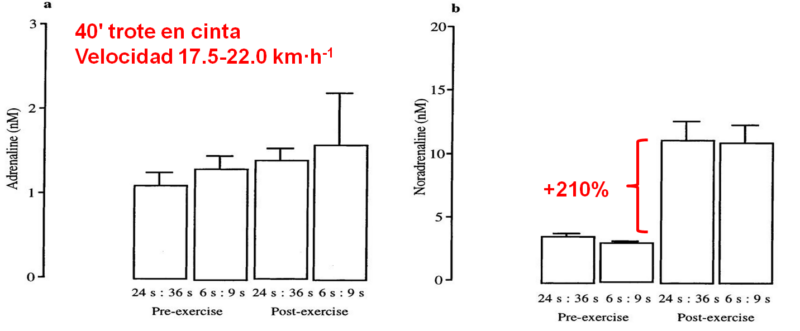
Figura 1.Christmass, (1999)
El objetivo principal de ese estudio era comparar las tasas de oxidación de sustratos en dos protocolos de ejercicio intermitente, con la velocidad en la cinta ergométrica idéntica y la duración de trabajo total para minimizar el efecto de diferencias en los factores como la activación del tipo de fibra muscular, respuestas hormonales, consumo de glucosa del músculo y disponibilidad de los AGL sobre la comparación de la utilización de sustratos. Estos factores confunden una explicación, en base a la regulación metabólica, de resultados previos que demuestran una oxidación de grasas inferior y la oxidación de los hidratos de carbono superior durante un ejercicio intermitente intenso comparado al ejercicio submáximo continuo (Christmass y cols. 1999).
Los resultados de ese estudio de Christmass del año 1999, demuestran que la oxidación de grasas es aproximadamente 3 veces menos y 1.3 veces mayor la oxidación de los hidratos de carbono durante el ejercicio intermitente con una duración proporcionalmente más larga de trabajo y de recuperación (es decir, 24":36" comparado a 6":9"). Esto es a pesar del hecho de que la velocidad de la cinta ergométrica (es decir, la intensidad del período de trabajo, equivalente al 109 ±5% VO2pico y la duración del trabajo total (es decir, 16 minutos) eran idénticas en los dos protocolos de ejercicio. Las diferencias en la oxidación de sustratos parecen estrechamente a la tasa 3 veces inferior de oxidación de grasas y de 1.2 veces superior a la tasa de oxidación de los hidratos de carbono observadas en comparación del ejercicio intermitente y el continuo (Christmass y cols. 1999), y es soportado por la observación de que el cociente respiratorio aumenta con la duración más larga de trabajo y de recuperación en el ejercicio intenso intermitente (Saltin y Essen 1971). Hacemos la observación, del uso poco fiable del cociente respiratorio en este tipo de esfuerzos no contínuos, como lo hicimos en varias entradas de blogs anteriores (Las condiciones para la interpretación apropiada de las mediciones del intercambio gaseoso respiratorio, y el uso fiable de la calorimetría indirecta para estimar la oxidación de grasas y de los hidratos de carbono en el actual estudio, se ha discutido previamente (Christmass y cols. 1999). Brevemente, se interpretaron datos para el intercambio de gas respiratorio representando un promedio de trabajo y recuperación combinado (es decir global) por un período de medición. Para la calorimetría indirecta, la estimación fiable del VCO2 de los tejidos requiere la presencia de un 'pool' de HCO3- estable (Romijn y cols. 1992))
Durante el ejercicio continuo, un aumento en el VO2, siendo el resultado de un aumento en la intensidad relativa del ejercicio del 65 al 70% VO2máx, se esperaría que sea asociado con un aumento en la oxidación de los hidratos de carbono (Romijn y cols. 1993). En contraste, durante el ejercicio intermitente un VO2 global superior, y de ahí la intensidad relativa del ejercicio superior (65 y 71% VO2pico para 24":36" y 6":9", respectivamente), fue asociado con tasas inferiores de oxidación de los hidratos de carbono. Un VO2 global superior así como un gasto energético superior en la misma velocidad de la cinta ergométrica, hace pensar en una eficiencia inferior del ejercicio durante 6":9". Esta reducida eficiencia puede ser debido en parte al VO2 adicional exigido de parar y arrancar de la cinta ergométrica como consecuencia del comienzo más frecuente de los períodos de trabajo comparado a 24":36". En suma, puesto que la oxidación de grasas comparada a la de carbohidratos requiere más O2 para la producción de la misma cantidad de ATP, las tasas superiores de oxidación de grasas en 6":9" habrían contribuido probablemente a esta eficiencia reducida.
En ese estudio de Christmass (1999), las diferencias en el reclutamiento del tipo de fibra muscular entre los dos protocolos del ejercicio intermitentes no fueron determinadas, pero improbablemente habrá influido en la diferencia observada sustancialmente en la utilización del sustrato. Essen (1978) demostró que el vaciamiento del glucógeno ocurrió en proporción similar en las fibras tipo I y II en ambos ejercicios intensos continuo (100% VO2máx) y ejercicio intenso intermitente (15":15"; la intensidad del período de trabajo era del 100% VO2máx). En contraste, el vaciamiento del glucógeno se observó predominantemente en las fibras tipo I durante el ejercicio submáximo continuo en la mitad de esta intensidad de ejercicio (un promedio del 55% VO2máx).
Estos resultados indican que la intensidad del ejercicio es el determinate principal de la activación del tipo de fibra muscular, mientras que el tipo de ejercicio (es decir, intermitente o continuo) afecta la tasa de vaciamiento del glucógeno en todos los tipos de fibra (Essen 1978). Los resultados de Vollestad y Blom (1985) y Vollestad y cols. (1984) apoyan esta conclusión y confirman la importancia de la intensidad del ejercicio y la duración determinando el tipo de fibra muscular reclutada. Puesto que la velocidad de la cinta ergométrica y la duración del trabajo total eran idénticas en 24":36" y 6":9", las diferencias en la activación del tipo de fibra muscular son improbables de explicar las diferencias observadas completamente en la utilización del sustrato.
Con lo cual, este hallazgo de Christmass indica que no pueden explicarse las diferencias observadas en la oxidación de grasas y de los hidratos de carbono en base a los efectos diferenciales de las catecolaminas circulantes en 24":36" y 6":9". No es probable que la disminución en la oxidación de grasas observada durante el ejercicio de 24":36" sea debida a la disponibilidad de los AGL en plasma limitada (Romijn y cols. 1995). La tasa de lipólisis, estimada a partir del glicerol en plasma, aumentó progresivamente en las dos condiciones de ejercicio y fue aproximadamente un 26% superior durante la prueba de 6":9". A pesar de esto, la concentración de AGL en plasma fue similar en las dos condiciones de ejercicio y fue considerablemente más alta que los niveles indicados para dañar la oxidación de grasas (Romijn y cols. 1995). Es más, un aumento progresivo en los niveles de los AGL ocurrió durante 24":36" con una tendencia hacia un aumento similar en 6":9". Combinado con un R relativamente estable, estos resultados indican que la disponibilidad de los AGL no fue limitante para la oxidación de grasas de todo el cuerpo en cualquier condición de ejercicio.
La idéntica velocidad de la cinta ergométrica y la duración de trabajo total en cada protocolo del ejercicio, combinado con una respuesta de catecolaminas similar y disponibilidad del AGL en plasma entre 24":36" y 6":9", apoya el concepto de que factores extrínsecos a las fibras musculares (es decir, la activación de las fibras del músculo y los niveles de hormonas circulantes) es improbable que sean la base para los resultados en ese estudio. En cambio, los resultados indican que un factor metabólico (o factores) dentro de las fibras musculares oxidativas es (o son) responsable de la diferencia en la utilización del sustrato entre el 24":36" y 6":9".
La reducida disponibilidad de O2 del músculo (Sahlin 1990), produciendo un aumento en la utilización de carbohidratos, puede tener implicaciones para la inhibición de la oxidación de las grasas (Sidossis y cols. 1997) en reverso del ciclo de glucosa-ácido graso clásico (Randle y cols. 1964) durante el ejercicio de 24":36". El protocolo de 24":36" se caracterizó por un 63% de tasa inferior de oxidación de grasas de todo el cuerpo comparada al protocolo de 6":9". Durante el ejercicio continuo, la oxidación de grasas de todo el cuerpo logra los niveles máximos en una intensidad de ejercicio moderado (65% VO2máx), pero disminuye en términos absolutos y relativos en una intensidad alta de ejercicio (85% VO2máx; Romijn y cols. 1993). Las razones para la disminución absoluta cuando la intensidad del ejercicio aumenta, es incierta. Varios estudios han mostrado una disminución en la tasa de aparición de AGL en plasma durante el ejercicio de alta intensidad (Jones y cols. 1980; Romijn et al. 1993). Sin embargo, la disponibilidad de los AGL en plasma no puede responder a la tasa 3 veces menor de oxidación de grasas observada para 24":36".
En realidad, Romijn y cols. (1995) han demostrado que la disponibilidad de AGL en plasma puede, a lo sumo, dar cuenta de aproximadamente el 50% de la disminución absoluta en la oxidación de grasas durante el ejercicio continuo de alta intensidad (85% VO2máx).
SI HAY POCO APORTE SUMINISTRO DE O2 ESO LLEVA A UN MAYOR CONSUMO DE CHO QUE DE GRASAS...POR LO TANTO HAY QUE OBSERVAR LA DISPONIBILIDAD DE OXIGENO EN EL MUSCULO
Otro estudio de Jabbour
G (2011), el objetivo era examinar el efecto del porcentaje de la grasa
corporal en respuesta de las catecolaminas en plasma a un test de sprint de
pedaleo (CST) en chicos adolescentes sedentarios. Para lo cual, se utilizaron
31 chicos adolescentes (9 obesos (% grasa corporal = 31.0% ± 3.0%), 11
sobrepeso (% grasa corporal = 24.0% ± 1.6%), y 11 magros (% grasa corporal =
16.0% ± 1.9%)), igualados para el consumo pico de oxígeno, completaron un CST
(test de sprint en bicicleta) que consiste en 6 sprints máximos de 6" con
2 minutos de pausa pasiva entre cada repetición. El rendimiento de cada sujeto
era determinado como la potencia media (PO (promedio)) desarrollada durante el
CST. Las concentraciones en plasma de lactato, epinefrina, y norepinefrina
fueron consecutivamente determinados en reposo, después del precalentamiento de
10 minutos, inmediatamente después del CST, y después de 20 minutos de
recuperación pasiva. Aunque los grupos no eran diferentes en la edad, altura, o
consumo pico de oxígeno (L·kg-1·FFM·min-1, la concentración de epinefrina
máxima fue significativamente (P <0.05) superior en los magros vs obesos y
se correlacionó negativamente al porcentaje de grasa corporal (r = -0.60, P
<0.05). Los valores máximos de la norepinefrina fueron superiores en los
magros versus sobrepesos y obesos, y una relación negativa se encontró entre la
concentración de norepinefrina máxima y el porcentaje de grasa corporal (r =
-0.60, P <0.05). La concentración máxima de lactato fuesuperior en los
magros vs sobrepeso y obeso (14.7 ± 3.3, 10.4 ± 2.7, y 10.2 ± 2.5 mm en magros,
sobrepeso, y obesos, respectivamente). Se puede observar la Figura 2. Una
relación significativa también se obtuvo entre los valores máximos de
norepinefrina y la epinefrina con ambaos concentraciones máximas de lactato (r
= 0.60 y r = 0.60, P <0.05, respectivamente) y PO (promedio) (r = 0.65 y r =
0.6, P <0.05).
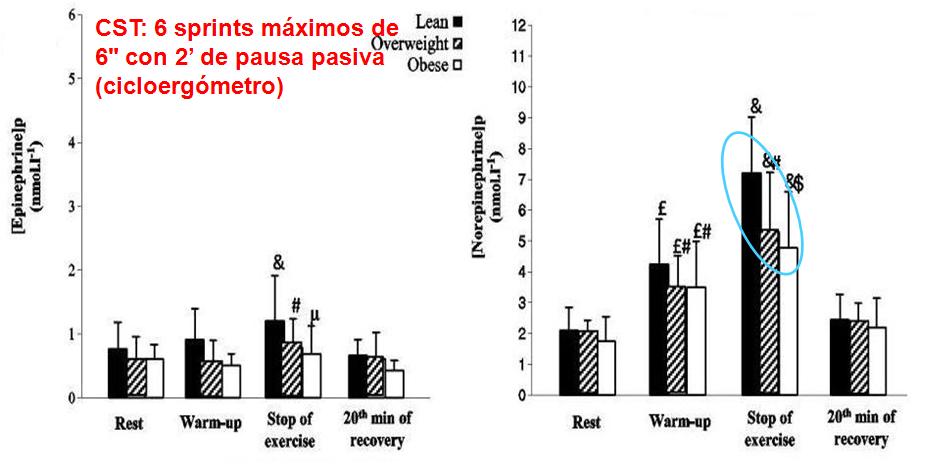
Figura 2.
Los resultados muestran que la respuesta de
las catecolaminas a un CST era afectada por el porcentaje de grasa corporal,
con reducidos valores de epinefrina y la norepinefrina en los adolescentes con
sobrepeso y adolescentes obesos.
Otro estudio sobre este
tema es el de Trapp EG de 2007. La respuesta metabólica a dos formas diferentes
de ejercicio de pedaleo intermitente de alta intensidad fue investigado en
mujeres jóvenes. Los sujetos (8 entrenadas y 8 desentrenadas) realizaron dos
turnos de ejercicio intermitente de alta intensidad: el sprint corto (S)
(sprint de 8 segundos el sprint, 12 segundos de recuperación) y el sprint largo
(LES) (sprint de 24 segundos, 36 segundos de recuperación) durante 20 minutos
en dos ocasiones separadas. La carga de trabajo y el consumo de oxígeno eran
mayores en las mujeres entrenadas pero no eran significativamente diferente
para los grupos S y LES. Las concentraciones de glicerol en plasma
significativamente aumentó durante el ejercicio. Las concentraciones de lactato
subieron durante los 20 minutos y fueron superiores para las mujeres
entrenadas. La concentración de catecolaminas
post-ejercicio también fue superior comparado con el pre-ejercicio para
ambos grupos. Los protocolos S y LES produjeron una respuesta metabólica
similar aunque tanto el lactato como las
catecolaminas fueron superiores después del sprint de 24 segundos.
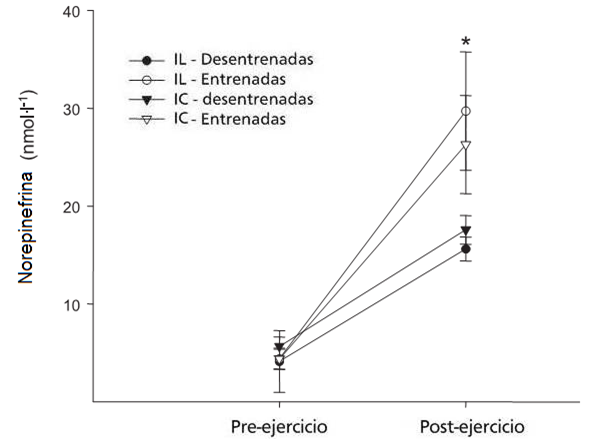
Figura 3.
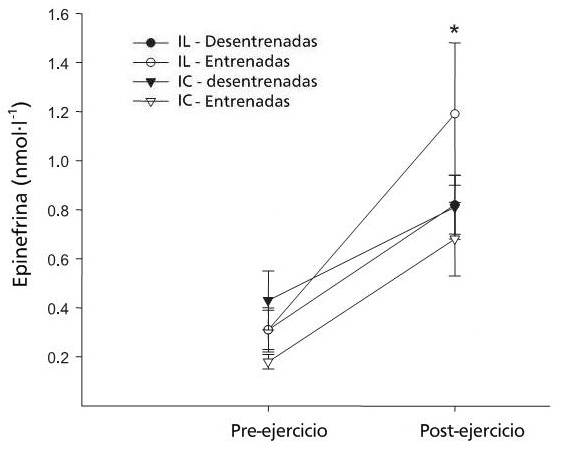
Figura 4.
En conclusión, estos resultados muestran que el ejercicio intermitente de alta intensidad produjo elevaciones significativas en las catecolaminas, lo que parecen estar relacionadas a las mayores concentraciones venosas de glicerol. La mujer entrenada comparada con la mujer desentrenada tendió a mostrar un aumento más temprano en las concentraciones de glicerol en plasma durante el ejercicio de alta intensidad.
Otro estudio muy reciente, es el de AB Abderrahmane (2013), donde se estudió el efecto del modo de recuperación (Activa [AR] vs Pasiva [PR]) sobre las respuestas de las catecolaminas en plasma (Adrenalina [A] y Noradrenalina [NA]) al ejercicio intervalado intenso se estudió durante el entrenamiento intervalado (IT, interval training). Se reclutaron 24 sujetos varones (21.1±1.1 años) asignados al azar a un grupo de control (GC, n=6), grupo de entrenamiento con AR (ARG, n=9) o con PR (PRG, n=9). Los grupos ARG y PRG participaron en una programa de entrenamiento intervalado 3 veces por semana durante 7 semanas (21 sesiones, Ver protocolo en Figura 5).
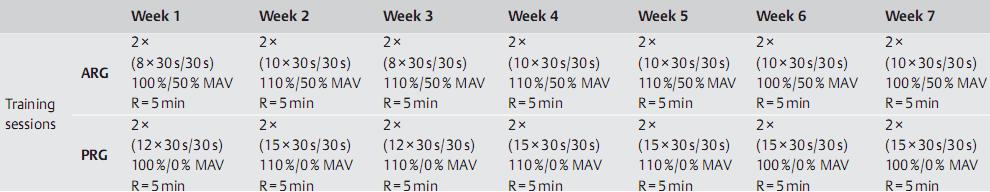
Figura 5. Protocolos.
Antes y después del entrenamiento, se midieron el consumo máximo de oxígeno (VO2máx) y la velocidad aeróbica máxima (MAV) (test de Léger). La A y NA se determinaron en reposo y al final de ejercici intervalado y después de 10 y 30 minutos de recuperación. Este entrenamiento indujo cambios significativos sólo en ARG: un aumento del VO2máx y de la VAM junto con un aumento significativo de A y NA al final de ejercicio (2.82±0.15 vs. 1.03±0.15 nmol/l y 7.22±0.36 vs. 6.65±0.57 nmol/l, respectivamente p <0.05). La proporción que A/NA también medida al final de ejercicio aumentó después del entrenamiento significativamente (0.41±0.11 contra 0.16±0.08, P>0.05).
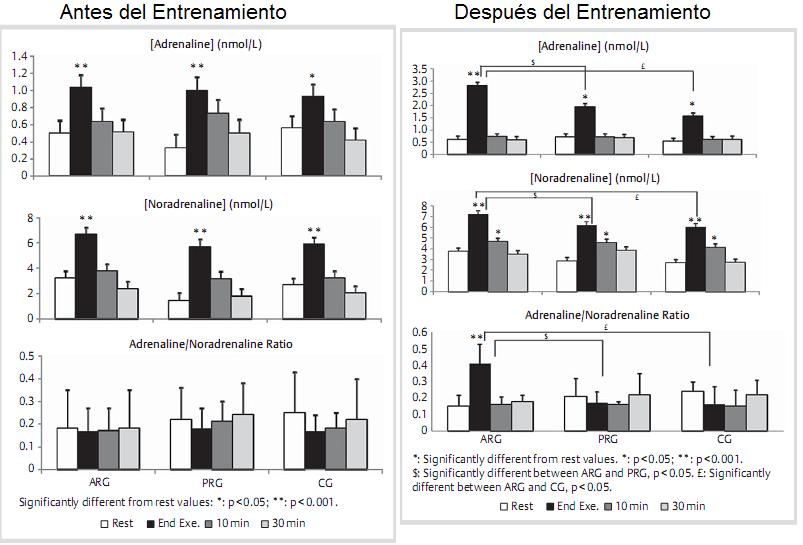
Figura 6. Resultados.
Los resultados presentes demuestran que el ejercicio intervalado con pausa activa (AR) induce un aumento significativo de las concentraciones de A y de NA en respuesta al ejercicio máximo. El estudio es más, muestras que un programa de entrenamiento intervalado con pausa activa puede inducir más estrés que el mismo programa con pausa pasiva.
De acuerdo a estos estudios en los que se analizaron la respuesta hormonal, dicha respuesta fue similar para todos los protocolos (24":36" y 6":9", y 15":15", 30”:30”). En el estudio de Trapp (bicicleta) la respuesta de las catecolaminas fue mayor que en el estudio de Christmass (en cinta). El aumento del lactato se ve acompañado del aumento de las catecolaminas.
En general, aumentan tanto el glicerol (en sangre) como las catecolaminas. Aún así, en el estudio de Christmass se hace incapié en el disminuido aporte de O2, lo que lleva a un mayor consumo de CHO que de FAT. Esto podría explicar las tasas mayores de oxidación de CHO que de FAT, ya que esa menor disponibilidad de O2 se asocia mayores tasas de glucogenólisis y de glucólisis por los aumentos de el ADP libre, AMP y Pi (Shalin, 1990; Ren, 1992; Christmass, 1999).
Durante el ejercicio de alta intensidad hay allí un fallo del tejido adiposo para liberar AG suficientes hacia el músculo (que es emparejado con una incapacidad de los músculos para usarlos, incluso cuando se proporcionan AG artificialmente). Esta limitación del tejido adiposo para el aporte de AGL puede reflejar alguna inhibición de la retroalimentación de la lipólisis, quizás vía el lactato, o inhibición posiblemente α-adrenérgica de la lipólisis en concentraciones de catecolaminas muy altas.
Pero…¿Por qué son signficativas las catecolaminas?
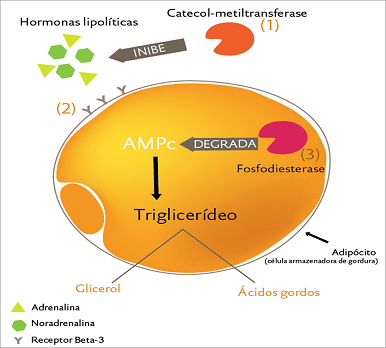
El primer paso en la quema de la grasa corporal es consiguiéndolo de sus células grasas. Este podría ser el paso más importante puesto que, si uno no puede sacarlo de la célula grasa, no puede quemarlo fuera de ella. Repasando acerca de que la grasa corporal se guarda en los triglicéridos principalmente, con una cantidad pequeña de agua y alguna ‘maquinaria’ enzimática y celular, movilizar la grasa corporal requiere que las personas degrademos triglicéridos guardados primeramente en tres ácidos grasos y una molécula de glicerol. La tasa que limita el paso en este proceso es una enzima llamada lipasa hormona sensible (HSL).
¿Qué regula la HSL? Aunque varias hormonas como la testosterona, el cortisol, estrógenos, y hormona de crecimiento tienen efectos moduladores sobre la actividad de la HSL (aumentando o disminuyendo principalmente los niveles totales de HSL en la célula grasa), las únicas hormonas que nosotros necesitamos que se involucren en términos de actividad de la HSL, son la insulina y las catecolaminas. El inactivador primario de la HSL es la hormona insulina y sólo toma cantidades muy diminutas (dependiendo de la sensibilidad de la insulina) para tener un efecto. Los niveles de insulina incluso en ayunas son suficientes para volver inactiva a la HSL por casi el 50%. Aumentos pequeños en la insulina (por la ingesta de carbohidratos o proteínas) vuelven aún más inactiva a la HSL. Adicionalmente, la presencia no más de triglicéridos en el torrente sanguíneo (vía infusión o sólo comiendo grasas de la dieta simplemente) también inhibe la actividad de la HSL de tal forma que esto no es tan simple como culpar a la insulina simplemente. Una manera u otra, cuando uno quiera comer, la HSL va hacerse inactiva, o por el aumento en la insulina a partir de las proteínas o carbohidratos o por la presencia de grasa en el torrente sanguíneo a partir de comer grasas.
Las hormonas primarias que activan la HSL son los catecolaminas: la adrenalina y noradernalina. La adrenalina es liberada de la corteza suprarrenal, recorriendo a través del torrente sanguíneo para afectar numerosos tejidos en el cuerpo. Esto significa que el flujo sanguíneo hacia las células grasas tiene un impacto en cuánto o cómo poca adrenalina alcanzará las células grasas. La noradrenalina es liberada desde las terminales nerviosas que actúan en forma recíproca directamente con las células.
Más técnicamente, la insulina y las catecolaminas afectan los niveles de AMP cíclico (cAMP) en la célula grasa, lo cual es lo que realmente determina cómo se activa la HSL. Cuando los niveles del cAMP son bajos, la actividad de la HSL también es baja, por lo que la degradación de grasas es baja. Cuando los niveles del cAMP son altos, la actividad de la HSL es alta y aumenta la degradación de las grasas.
La insulina desciende
los niveles de cAMP y las catecolaminas, en general, aumentan los niveles de
cAMP. Cuanto más alto es el nivel de cAMP, más activa es la HSL y más grasa
corporal es degradada y liberada desde las células grasas. Debe aclararse que,
desde un punto de vista de la pérdida de grasa, nosotros necesitamos niveles
altos de cAMP, que lo dan esfuerzos que impliquen una intensidad considerable.
Esta temática estará presente en la Webinar que realizaremos próximamente.
Bibliografía
Tremblay A, Simoneau J, Bouchard C. Impact of exercise intensity on body fatness and skeletal muscle metabolism. Metabolism 43:814–818, 1994.
Astrand I, Astrand PO, Christensen EH, Hedman R. Myohemoglobin as an oxygen-store in man. Acta Physiol Scand 48:454–460,1960.
Hansen D, Meeusen R, Mullens A, Dendale P. (2012 ). Effect of acute endurance and resistance exercise on endocrine hormones directly related to lipolysis and skeletal muscle protein synthesis in adult individuals with obesity. Sports Medicine 42(5):415-31.
M.A. Christmass, B. Dawson, Peter G. Arthur. Effect of work and recovery duration on skeletal muscle oxygenation and fuel use during sustained intermittent exercise. Eur J Appl Physiol (1999) 80: 436-447.
Vollestad NK, Blom PCS (1985) Effect of varying exercise intensity on glycogen depletion in human muscle fibres. Acta Physiol Scand 125:395±405.
Vollestad NK, Vaage O, Hermansen L (1984) Muscle glycogen depletion patterns in type 1 and subgroups of type 2 fibres during prolonged severe exercise in man. Acta Physiol Scand 122:433±441.
Jabbour G, Lemoine-Morel S, Casazza GA, Hala Y, Moussa E, Zouhal H. Catecholamine response to exercise in obese, overweight, and lean adolescent boys. Med Sci Sports Exerc. 2011 Mar;43(3):408-15.
Trapp EG, Chisholm D, Boutcher SH. Metabolic response of trained and untrained women during high-intensity intermittent cycle exercise. Am J Physiol Regul Integr Comp Physiol 293: R2370–R2375, 2007.