
Grupo Sobre Entrenamiento. Grupo de Recursos sobre Ciencias del Ejercicio.
Resumen
¿QUE ES UNA AYUDA ERGOGENICA?
En primer lugar sería importante considerar a que se hace referencia cuando se habla de ayuda ergogénica. Una ayuda ergogénica es cualquier técnica de entrenamiento, dispositivo mecánico, práctica nutricioinal, método farmacológico o técnica psicológica que pueda incrementar la capacidad de rendimiento durante el ejercicio y/ incrementar las adaptaciones al entrenamiento [Williams, 1999; Leutholtz, 2001]. Esto incluye ayudas que pueden ayudar a preparar a un individuo para el ejercicio, mejorar la eficiencia en el ejercicio, y/o mejorar los procesos de recuperación luego del ejercicio [Kreider, 2004]. Las ayudas ergogénicas también pueden permitir que un individuo tolere en mayor grado entrenamientos de alta intensidad promoviendo una más rápida recuperación o ayudando a mantener la salud del atleta durante el entrenamiento. Aunque esta definición parece bastante simple, existe un considerable debate respecto del valor ergogénico de diversos suplementos nutricionales [Kreider, 2004]. Algunos especialistas en nutrición deportiva solo consideran un suplemento ergogénico si los estudios científicos muestran que el suplemento en cuestión incrementa significativamente el rendimiento deportivo (e.g., ayuda a correr más rápido, a levantar más peso y/p a realizar más trabajo durante un ejercicio dado). Por otra parte, algunos consideran que si un suplemento ayuda a un atleta a prepararse para realizar ejercicios o lo ayuda a recuperarse luego del ejercicio, entonces tiene el potencial de mejorar las adaptaciones para el entrenamiento y por lo tanto debería ser considerado ergogénico. A pesar de esto, es necesario tener una visión más amplia respecto del valor ergogénico de los suplementos. Si bien en general lo importante es determinar los efectos de un suplemento respecto de la mejora del rendimiento en una única serie de ejercicios, también es importante tener en cuenta que uno de los objetivos del entrenamiento es ayudar a las personas a tolerar el entrenamiento en un mayor grado [Kreider, 2004]. Las personas que tienen una mayor tolerancia al entrenamiento con frecuencia experimentan mayores ganancias a través del tiempo. Consecuentemente, la utilización de las prácticas nutricionales puede ayudar a preparar a los sujetos para realizar ejercicios y/o para acelerar los procesos de recuperación luego del ejercicio deberían ser consideradas ayudas ergogénicas. En la Tabla 1, se presenta una lista con los suplementos nutricionales más utilizados.
Los factores principales que afectan la capacidad de rendimiento durante el ejercicio incluyen el potencial genético del individuo, la calidad del entrenamiento y la efectividad del mismo [Leutholtz and Kreider, 2001]. Más allá de estos factores, la nutrición desempeña un rol crítico respecto de la optimización de la capacidad de rendimiento. Si un atleta no entrena lo suficiente o tiene una dieta inadecuada, su rendimiento se verá disminuido [Kreider, 1998]. Por otra Parte si el atleta entrena demasiado sin una dieta que cubra sus necesidades, entonces se instala la posibilidad de que se produzca sobreentrenamiento [Leutholtz and Kreider, 2001]. La idea de esta revisión es dar un pantallazo acerca de los suplementos nutricionales más comúnmente utilizados por los deportistas en tres áreas específicas de rendimiento: entrenamiento de la fuerza y de la potencia (incluido el incremento de la masa muscular), ejercicios intermitentes de alta intensidad y ejercicios aeróbicos.
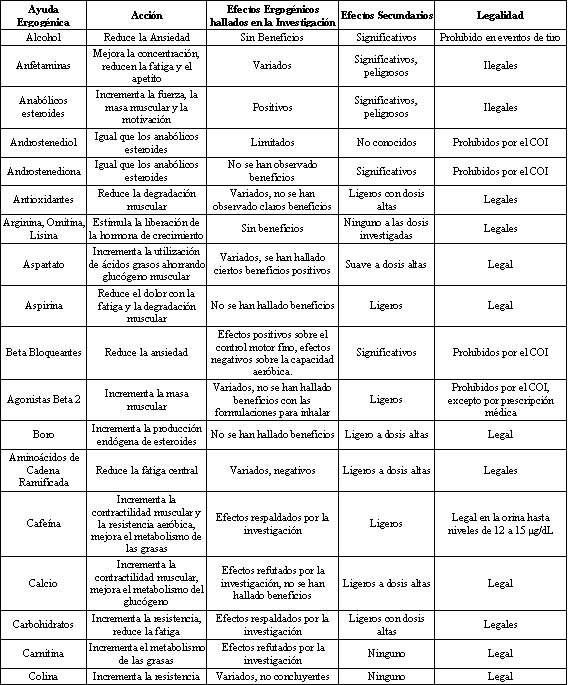
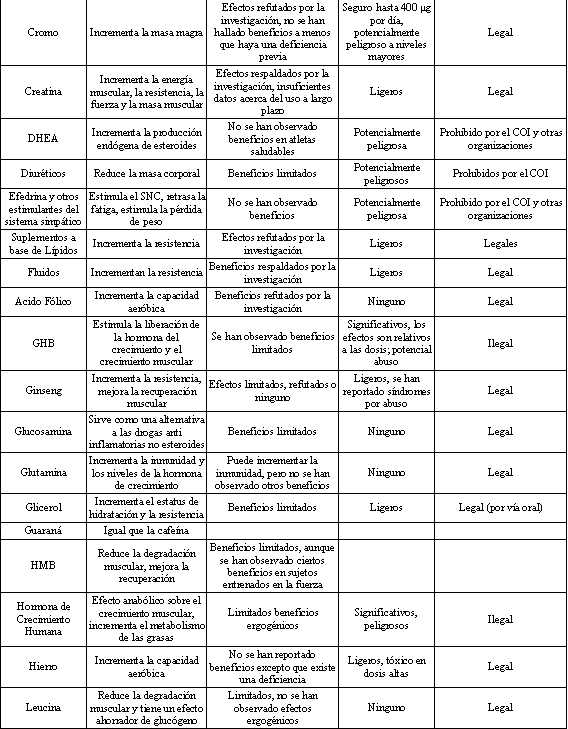
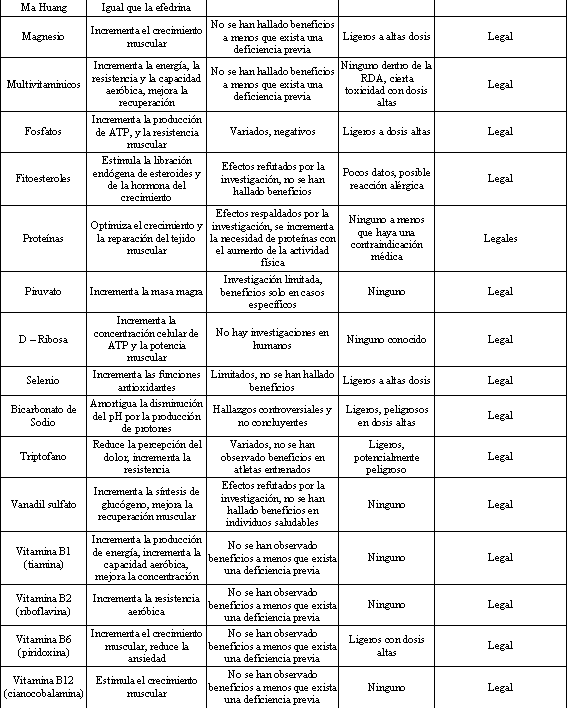

Tabla 1. Breve resumen de algunos suplementos dietarios comúnmente
utilizados. COI=Comité Olímpico Internacional. Modificado de Ahrendt DM.
“Ergogenic Aids: Counseling The Athlete” Am. Fam. Physician 63: 5 (2001).
REQUERIMIENTOS ENERGETICOS
El primer principio nutricional para optimizar el rendimiento de los atletas es asegurar que estos consumen las suficientes calorías como para cubrir las demandas energéticas impuestas por el ejercicio para mantener el balance energético [ACSM, 2000; Leutholtz and Kreider, 2001]. La ingesta calórica diaria en individuos desentrenados comúnmente se encuentra en el rango de las 1900-300 kcal/día (i.e., 27-43 kcal/kg/día para una persona de 70 kg) [Williams, 1999; Berning, 1998; ACSM, 2000; Food and Nutrition Board, 1989], particularmente, en 1989 la RDA “Recomended Dietary Allowances” [Nacional Research Council, 1989], establecía que los requerimientos energéticos para mujeres y hombres ligera a moderadamente activos de entre 19 y 50 años de edad era de 2200 a 2900 kcal por día, respectivamente. El balance energético es esencial para el mantenimiento de la masa de tejido magro, las funciones inmunes y reproductivas y para el óptimo rendimiento deportivo. El balance energético se define como el estado en el cual la ingesta energética (la suma de energía proveniente de los alimentos, fluidos y suplementos ingeridos) iguala el gasto energético (la suma de energía gastada por el metabolismo energético, el efecto termogénico de los alimentos, y cualquier actividad física voluntaria). La inadecuada ingesta en relación al gasto energético compromete el rendimiento y los beneficios asociados con el entrenamiento. Con una limitada ingesta energética, la masa grasa y la masas magra serán utilizadas por el cuerpo como fuentes energéticas [ACSM, 2000]. De esta manera la pérdida de masa muscular resultará en la pérdida de fuerza y resistencia y además la ingesta crónicamente baja de energía con frecuencia resulta en una pobre ingesta de nutrientes, particularmente de micronutrientes.
El gasto energético es influenciado por factores hereditarios, la edad, el sexo, el tamaño corporal, la masa libre de grasa, y la intensidad, frecuencia y duración del ejercicio. Es decir cuanto mayor es la duración y la intensidad de los ejercicios realizados por los deportistas, mayor es el gasto energético. Para los deportistas, se recomienda que se evalúe el tipo de ejercicio realizado, la intensidad, frecuencia y duración del mismo para de esta manera adicionar este incremento a la energía necesaria para las actividades normales diarias [Montoye, 1996, Manore, 2000; Hawley and Burke, 1998]. El gasto energético estimado para diferentes atletas se encuentra en el rango de las 3500 kcal/día para individuos que entrenar 30-60 min/día hasta 12000 kcal/día para ciclistas que compiten en el Tour de France.
SUPLEMENTOS UTILIZADOS PARA INCREMENTAR LA FUERZA Y LA POTENCIA: INCREMENTO Y REPARACIÓN DE TEJIDOS
Uno de los objetivos principales del entrenamiento de la fuerza es promover ganancias en la masa muscular. El entrenamiento crónico de sobrecargo promueve ganancias en la masa libre de grasa que están en el rango de los 0 a 1 kg por mes [Wilmore, 1974; Forbes, 1991, Kraemer, 1994]. Sin embargo, las ganancias en la masa libre de grasa durante el entrenamiento de sobrecarga varía considerablemente entre los individuos [Wilmore, 1974; Forbes, 1991]. La utilización de suplementos dietarios parece ser particularmente predominante entre los atletas de deportes de fuerza y potencia, e incluso algunas encuestas muestran que la mayoría de los atletas de estos deportes utilizan uno o más suplementos [Maughan, 2003]. Una variedad de suplementos se venden como agentes “anabólicos” o “anticatabólicos”, con efectos directos o indirectos sobre las vías de síntesis de proteínas musculares. Los mecanismos de acción propuestos incluyen, un efecto de acción de masas sobre las vías de síntesis de proteínas incrementando la disponibilidad de aminoácidos, estimulando la liberación de diversas hormonas, potenciando la acción hormonal, incrementando el volumen celular, o actuando como agentes de adaptación (i.e., promoviendo las adaptaciones al entrenamiento) [Maughan, 2003]. Los suplementos a la venta en esta amplia categoría incluyen aminoácidos, boro, cromo, colostrum, creatina. Hodroximetilbutirato (HMB), ornitina, alfacetoglutarato, proteínas, Tríbulus terrestris, vanadio y zinc.
Si bien es claro que los agentes farmacológicos, tales como los anabólicos androgénicos esteroides y la hormona del crecimiento pueden promover el crecimiento muscular [Forbes, 1985, 1992; Kuipers, 1991; Crist, 1988, 1991] existen respecto de esto ciertas preocupaciones éticas y legales [ASCM, 1987]. Consecuentemente los atletas han dependido en mayor grado de las estrategias y los suplementos nutricionales diseñados para promover el crecimiento muscular [Kreider, 1999]. Aunque existe evidencia que sugiere que la manipulación dietaria y/o la suplementación de algunos nutrientes puede incrementar las ganancias en la masa libre de grasa durante el entrenamiento de sobrecarga, mucho de los nutrientes comercializados tiene poco respaldo científico respecto de su efectividad.
Proteínas y Aminoácidos
La utilización de dietas altas en proteínas tiene una larga historia en la nutrición deportiva. Existe bastante evidencia acerca de que los requerimientos de proteínas se ven incrementados cuando se incrementa la intensidad del entrenamiento y con frecuencia se recomienda que la ingesta de proteínas en atletas de fuerza sea 50-100% mayor que la de los sujetos sedentarios (Tarnoposky, 2001]. Es una creencia común entre los atletas dedicados a deportes de fuerza que para promover el crecimiento muscular, la dieta debe ser suplementada con grandes cantidades de proteínas [Kreider, 1999]. Si bien existen estudios [Lemmon, 1992; Tarnopolsky, 1992] que indican que los atletas involucrados en entrenamientos intensos de la fuerza pueden requerir mayores cantidades de proteínas que los sujetos desentrenados (i.e., 1.3 a 2 vs 0.8 a 1 g/kg/día), la mayoría de los atletas que consumen los alimentos suficientes como para mantener el balance energético alcanzan estas recomendaciones [Lemmon, 1992, Tarnopolsky, 1992; Kreider, 1993]. Como se mencionó anteriormente, los atletas con frecuencia insisten en la necesidad de consumir grandes cantidades de proteínas para incrementar la masa muscular, pero la literatura científica no respalda esta suposición. Esta aparente inconsistencia puede ser explicada por el Modelo de Demanda Adaptativa Metabólica de Millward, el cual propone que el cuerpo se adapta a ingestas altas o bajas, y que este ajuste a los cambios en la ingesta se produce solo muy lentamente [Millward, 2001]. Con altas ingestas de proteínas, se produce una regulación ascendente de la degradación proteica y la oxidación de aminoácidos [Maughan, 2003]. El atleta que consume una dieta con grandes cantidades de proteínas y reduce agudamente su ingesta proteica experimentará una pérdida de tejido magro hasta que se alcance un nuevo equilibrio [Millward, 2001]. Además, el incremento de la ingesta de proteínas por encima de lo necesario para mantener el balance nitrogenado no parece promover el aumento de tejido magro [Lemmon, 1992, Tarnopolsky, 1992]. Por ejemplo, Tarnopolsky et al [1992] investigaron los efectos de la ingesta dietaria de proteínas sobre la fuerza y la composición corporal en sujetos desentrenados utilizando un diseño doble ciego cruzado aleatorio. Seis sujetos sedentarios y 7 atletas entrenados en la fuerza ingirieron 0.86, 1.4 y 2.4 g/kg/día de proteínas durante 13 días separados por un período de lavado de 8 días. Los atletas entrenados en la fuerza tuvieron mayores requerimientos diarios de proteínas (1.4 g/kg/día). Sin embargo, el incremento en la ingesta proteica no afecto los cambios en la masa libre de grasa en ninguno de los dos grupos. Estos hallazgos sugieren que los atletas entrenados en la fuerza pueden requerir entre 1.7 a 1.8 g/kg/día de proteínas para asegurar el balance nitrogenado, pero la ingesta de proteínas por encima de estos niveles no promueve el crecimiento muscular.
Similarmente Lemmon et al [1992] investigaron los efectos de la suplementación con proteínas sobre la composición corporal y las alteraciones en la fuerza en un grupo de físico culturistas novatos. Utilizando un diseño con repetidas mediciones transversales, 12 físico culturistas varones ingirieron 1.35 o 2.62 g/kg/día de proteínas con una dieta de 3500 kcal/día. Los participantes entrenaron durante 4 semanas, luego de lo cual llevaron a cabo un período de lavado de 7 días, y repitieron el experimento ingiriendo la cantidad de proteínas que antes había ingerido el otro grupo. Los autores no hallaron diferencias entre los grupos respecto de las ganancias en la masa corporal, masa muscular (determinada por densidad, excreción de Creatina, y tomografía axial computada) o en la fuerza.
Si los suplementos de proteínas son necesarios, es un tema diferente, y ha sido discutido ampliamente por Tipton y Wolfe [1994]. Es claramente posible alcanzar ingestas proteicas muy elevadas seleccionando los alimentos apropiados, pero también es cierto que muchos alimentos altos en proteínas tienen un alto contenido de grasas. Los suplementos de proteínas le ofrecen a los atletas la posibilidad de alcanzar la ingesta proteica deseada sin un incremento poco deseable en la ingesta de grasas y sin mayores cambios en sus hábitos alimenticios.
Respecto de los aminoácidos, existen datos de estudios cínicos en los cuales participaron individuos con diferentes grados de estrés (e.g., traumas, lesiones por quemaduras o cirugía) que mostraron que la suplementación con aminoácidos puede reducir la extensión de la degradación muscular, pero este estado catabólico es poco relevante para los atletas saludables que intentan incrementar la masa muscular [Maughan, 2003]. Tipton et al [1999] evaluaron la hipótesis de que la ingesta oral de aminoácidos podría incrementar la síntesis neta de proteínas musculares luego de una sesión aguda de entrenamiento de sobrecarga en sujetos desentrenados. Tres hombres y tres mujeres realizaron ejercicios de alta intensidad para el entrenamiento de la fuerza (17 series totales de prensa de piernas, sentadillas, extensiones y flexiones de rodillas, 8-10 repeticiones por serie al 75% de 1RM) en tres ocasiones llevadas a cabo en orden aleatorio. Comenzando ~ 45 min después del ejercicio, los sujetos ingirieron 1 litro de una bebida a base de placebo (endulzante artificial), 40 g de aminoácidos, o 40 g de aminoácidos esenciales a una tasa de 100 ml cada 18-20 min. Las síntesis mixta de proteínas musculares, la degradación proteica y el balance proteico neto se calcularon en base a una infusión constante de L-[ring 2H5]fenilalanina. Los autores afirmaron que la ingesta de ambas mezclas de aminoácidos resultó en una “hiperaminoacidemia significativa” en comparación con la solución placebo, pero desafortunadamente solo reportaron las concentraciones intramusculares y totales en sangre de fenilalanina, leucina y lisina. De hecho, las dos pruebas en las cuales los sujetos ingirieron aminoácidos resultaron en un incremento significativo en los tres aminoácidos medidos en la sangre arterial; sin embargo, solo la mezcla de aminoácidos esenciales resultó en un incremento significativo de las concentraciones intramusculares. La diferencia probablemente se debió al hecho de que las concentraciones de fenilalanina, leucina y lisina en la solución de aminoácidos esenciales era ~2 veces mayor en comparación con la solución de varios aminoácidos. No se hallaron diferencias significativas entre las tres condiciones respecto de la síntesis o de la degradación proteica luego del ejercicio; sin embargo, el balance proteico neto fue significativamente mayor durante los tratamientos con aminoácidos en comparación con el tratamiento con placebo. Tipton et al. [1999] concluyeron que la suplementación con aminoácidos por vía oral era tan efectiva como la infusión de aminoácidos para estimular el anabolismo proteico luego de la realización de ejercicios de sobrecarga. Además, la cantidad absoluta de aminoácidos parece ser más importante que la combinación específica de los mismos [Tipton et al 1999].
En el caso de los aminoácidos de cadena ramificada se ha reportado que la suplementación reduce la degradación proteica inducida por el ejercicio y/o el flujo de enzimas musculares [Bloomstrand, 1991; Combes and McNaughton, 1995; Kreider, 1992], posiblemente promoviendo un perfil hormonal anticatabólico [Carli, 1992; Chandler, 1994]. Teóricamente, la suplementación con aminoácidos de cadena ramificada ayuda a minimizar la degradación proteica y por lo tanto derive en mayores ganancias de masa libre de grasa [Kreider, 1993, 1998; Mourier, 1997].
Varios investigadores han demostrado que el incremento en la disponibilidad de los aminoácidos a partir de aminoácidos exógenos eleva la síntesis proteica y resulta en un balance proteico muscular neto positivo [Gelfand, 1986; Bennet, 1989; Svanberg, 1996; Biolo, 1997;]. Tomando en conjunto los resultados de estos estudios, parece evidente que hay una relación entre la tasa de síntesis de proteínas musculares y la disponibilidad intracelular de aminoácidos (i.e., la presencia de aminoácidos transportados hacia el compartimento intracelular desde el espacio extracelular y los aminoácidos derivados de la degradación de proteínas musculares). Los aminoácidos individuales que se afirma promueven el crecimiento muscular incluyen la glutamina, los previamente mencionados aminoácidos de cadena ramificada, la leucina, lisina, arginina y la ornitina.
La glutamina es un ingrediente común hallado actualmente en la mayoría de los suplementos denominados “ganadores de peso”. Se ha sugerido que la glutamina promueve el crecimiento muscular y la reducción en la inmunodepresión inducida por ele ejercicio. Estas afirmaciones están basadas en estudios llevados a cabo con animales y humanos que han investigado los efectos de la suplementación con glutamina sobre la síntesis proteica [Rennie, 1994, 1996], el volumen celular [Low, 1996] y en la síntesis de glucógeno [Varnier, 1995]. Los estudios también sugieren que el ejercicio intenso puede reducir los niveles de glutamina [Parry-Billings, 1992; Kargotich, 1996] y que esta reducción puede contribuir a la inmunodepresión inducida por el ejercicio en atletas sobreentrenados [Parry-Billings, 1992; Kargotich, 1996; Kreider, 1998]. Aparentemente la glutamina es un importante nutriente metabólico que afecta la síntesis proteica [Rennie, 1994, 1996] posiblemente incrementando el volumen celular y la presión osmótica [Low, 1996]. Además la disponibilidad de glutamina afecta directamente la función linfocítica [Parry-Billings, 1992; Kargotich, 1996; Kreider, 1998]. Consecuentemente, existe cierta evidencia para respaldar la suplementación con glutamina para promover el crecimiento muscular y/o prevenir infecciones del tracto respiratorio en los atletas. Sin embargo, aun no se han realizado estudios a largo plazo que han investigado los efectos de la suplementación con glutamina sobre la síntesis proteica, la composición corporal y la incidencia de infecciones del tracto respiratorio durante el entrenamiento de sobrecarga. Por último, se podría mencionar que si bien altas dosis de arginina, orinitina y lisina pueden resultar en aumentos en los niveles circulantes de la hormona de crecimiento y de las concentraciones de insulina, estos no han mostrado provocar cambios en la masa magra o en la función muscular (Maughan, 2003)
Leucina y Calcio β-hidroxi-β-metilbutirato
La lecina y los metabolitos de la leucina como el α-cetoisocaproato han mostrado inhibir la degradación proteica, particularmente durante períodos de proteolisis incrementada [Fair, 1992; Nissen, 1996]. Se ha sugerido que los efectos anticatabólicos de la leucina y del α-cetoisocaproato son regulados por un metabolito de la leucina, el β-hidroxi-β-metilbutirato (β-HMB). Los estudios llevados a cabo con animales indican que el HBM es sintetizado a partir de α-cetoisocaproato casi enteramente como un subproducto del metabolismo de la leucina y que aproximadamente el 5% de la leucina oxidada se convierte en HMB [Nissen, 1994; Van Koevering, 1994]. Aunque se desconoce el mecanismo de acción del HMB, se ha hipoteitzado que actúa reduciendo la proteolisis muscular o incrementando la integridad celular proveyendo sustratos para la síntesis de colesterol [Nissen and Sharp, 2003]. En sujetos previamente desentrenados, el HMB puede incrementar la masa magra y la fuerza más que el entrenamiento de sobrecarga por si solo [Nissen, 1996; Gallagher, 2000; Panton, 2000]. Además, Nissen et al [1996] han reportado que la ingesta de HMB reduce la excreción urinaria de 3-metilhistidina, un hallazgo que es consistente con la reducción de la proteolisis muscular. Asimismo se ha reportado que la infusión intravenosa de leucina reduce la degradación proteica en humanos sugiriendo que la leucina puede actuar como un regulador del metabolismo [Nair, 1992]. En 1997, Nissen et al, reportaron ganancias significativas en la masa libre de grasa y en la fuerza en hombres y mujeres desentrenados que iniciaban el entrenamiento de sobrecarga cuando se les administró calcio β-HMB con dosis de 1.5 o 3 g/día durante 3 a 4 semanas. Estas ganancias en la masa libre de grasa fueron aproximadamente 0.4 a 0.7 kg mayores que en los grupos placebo. Estos investigadores también reportaron que la suplementación con 3 g/día con calcio β-HMB ingerida conjuntamente con proteínas y carbohidratos en polvo incrementó significativamente la masa libre de grasa (~ 2.7 kg en las primeras 3 a 4 semanas) durante 7 semanas de entrenamiento de sobrecarga en jugadores de fútbol americano de nivel universitario en comparación con los participantes que ingirieron una cantidad isocalórica de jugo de naranja. Vukovich et al [1997] reportaron que 8 semanas de suplementación con calcio β-HMB (3 g/día) incrementó significativamente la masa libre de grasa (- 0.58% vs 1.5%), redujo la masa grasa (0.27 vs -2.2%), y promovió mayores ganancias de fuerza en 1RM tanto en el tren superior como inferior en un grupo de hombres y mujeres ancianos que iniciaban el entrenamiento.
Sin embargo, en individuos entrenados y en atletas de elite pertenecientes a deportes de potencia la ingesta de HMB parece no provocar incrementos en la masa magra o en la fuerza [Kreider, 1999b; Slater, 2001; Ransone, 2003] o en la capacidad anaeróbica [O’Connor and Crowe, 2003]. Asimismo el β-HMB tampoco parece afectar los marcadores del estatus catabólico o la integridad de la membrana celular en individuos bien entrenados [Kreider, 1999; Paddon-Jones, 2001; Slater, 2001].
En un estudio llevado a cabo por Kreider et al [1997] 52 jugadores de fútbol americano de nivel universitario que realizaban un entrenamiento de sobrecarga/agilidad (~ 8 horas semanales) fueron suplementados, de manera doble ciego y aleatoria, con placebo a base de carbohidratos y electrolitos, placebo con calcio β-HMB, placebo con monohidrato de creatina (15.5 g/día) o placebo con HMB (3 g/día) y monofosfato de creatina (1.5 g/día) durante 28 días. En este estudio se obtuvieron muestras sanguíneas en ayunas, se determinó la composición corporal mediante absorciometría dual de rayos X (DEXA), el máximo esfuerzo isotónico en los ejercicios de press de banca, sentadillas y cargadas de potencia, y se realizó un test de sprint máximo en cicloergómetro (12 × 6 sprints con 30 segundos de recuperación). Las ganancias en la masa libre de grasa en el grupo placebo/β-HMB (1.4±0.3 kg) no fueron estadísticamente significativas de las observadas en el grupo placebo (1.3 ± 0.3 kg) y las ganancias en la masa libre de grasa en el grupo placebo/creatina (2.4±0.4 kg) y en el grupo placebo/creatina/β-HMB (2.6±0.5 kg) fueron significativamente mayores que las de los otros dos grupos. Sin embargo no hubo evidencia de que la suplementación con calcio β-HMB redujera el flujo enzimatico en el hígado y en los músculos. Además, la suplementación con calcio β-HMB tuvo efectos mínimos sobre el rendimiento en los esprints y en el volumen levantado en los ejercicios de pesas. Estos hallazgos sugieren que la suplementación con calcio β-HMB durante el entrenamiento de alta intensidad tiene un valor limitado.
En un reciente meta-análisis de estudios llevados a cabo tanto con sujetos entrenados como desentrenados, Nissen y Sharp [2003] reporaron que la suplementación con β-HMB incrementó la masa magra en un 0.28% por semana y la fuerza en un 1.40% por semana en comparación con el entrenamiento de la fuerza por si solo. En conjunto estos hallazgos sugieren que el β-HMB puede tener cierto valor para los atletas que comienzan con un programa para el entrenamiento de la fuerza. Consecuentemente se podría decir, respecto de la suplementación con β-HMB en atletas entrenados en la fuerza, que:
- No es claro si el β-HMB promueve el crecimiento muscular y/o ganancias en la fuerza en atletas bien entrenados
- Existe evidencia de que la suplementación con creatina y/o suplementos que contienen creatina pueden ser más efectivos; y
- Si se desea probar esta estrategia nutricional se recomienda ingerir el β-HMB conjuntamente con carbohidratos y proteínas en polvo fortificado con vitaminas y minerales o en adición a suplementos de creatina o que contengan creatina.
Creatina
La suplementación dietaria con creatina y las formulaciones nutricionales que contienen creatina se han vuelto una estrategia nutricional muy popular empleada por los atletas de deportes de fuerza y potencia para promover ganancias en la fuerza y en la masa libre de grasa [Kreider, 1995]. Las principales razones para la suplementación con creatina (20 a 25 g/día durante 4 a 7 días y luego 2 a 25 g/día) son que se han reportado incrementos en la masa corporal total [Varnier, 1995; Parry-Billings, 1992; Kargotich, 1996; Kreider, 1992, 1993a, 1993b, 1995, 1996,1998; Bloomstrand, 1991; Coombes, 1995; Mourier, 1997; Greenhaf, 1994; Lemmon, 1995, Becque, 1997; Earnest, 1995; Ziegenfuss, 1997], la masa libre de grasa [Becque, 1997; Earnest, 1995; Kreider, 1996, 1997a, 1997b, 1998; Ziegenfuss, 1997; Stout, 1997; Kirksey, 1997; Vandenburghe, 1997], en la capacidad de realizar esfuerzos de esprint únicos y/o repetidos [Earnest, 1995; Kreider, 1998; Ziegenfuss, 1997; Stout, 1997; Kirksey, 1997, Casey, 1996; Birch, 1994; Greenhaff, 1993; Grindstaff, 1997; Prevost, 1997; Balsom, 19995; Dawson, 1995; Harris, 1993] en la fuerza y/o la potencia [Becque, 1997; Earnest, 1995; Stout, 1997; Vandenburghe, 1997; Casey, 1996; Birch, 1994; Volek, 1997] y en el trabajo realizado durante series de contracciones musculares máximas [Becque, 1997; Earnest, 1995, Kreider, 1996, 1998; kirksey, 1997; Greenhaff, 1993; Balsom, 1995; Volek, 1997]. Se ha sugerido que el incremento en la masa corporal se debe a que la creatina incrementa la retención de agua [Balsom, 1995], la síntesis proteica [Becque, 1997; Dawson, 1995; Harris, 1993, Balsom, 1994; Bessman, 1988; Inwall, 1976] y/o a una incrementada calidad del entrenamiento lo cual deriva en mayores ganancias en la fuerza y en la masa libre de grasa [Earnest, 1995, Kreider, 1996, 1997, 1998; Ziegenfuss, 1997, Stout, 1997; Kirksey, 1997; Vandenburghe, 1997]. Las ganancias en la masa corporal y en la masa libre de grasa luego de la suplementación con creatina o con formulas nutricionales que contienen creatina son característicamente de entre 0.8 y 3 kg mayores que las observadas en los sujetos de control, dependiendo de la duración y la cantidad de la suplementación [Kreider, 1999].
Por ejemplo, Earnest et al [1995] reportaron que la suplementación con creatina en un período de 28 días (20 g por día) durante el entrenamiento de la fuerza, incrementó significativamente la masa muscular en 1.7 kg (p<0.05) y que las ganancias de la masa libre de grasa (determinadas hidrostáticamente) dieron cuenta de 1.5 kg del total de la masa corporal ganada (p = 0.054). Asimismo, Vandenburghe et al [1997] reportaron que las mujeres que consumieron creatina (20 g/día durante 4 días seguido de un período de 66 días en donde consumieron 5 g/día) durante el entrenamiento de la fuerza experimentaron ganancias significativas en la masa libre de grasa en comparación con el grupo placebo. Estas ganancias fueron mantenidas en un subsiguiente período de desentrenamiento de 70 días durante el cual se mantuvo la suplementación (5 g/día).
Los estudios llevados a cabo por Kreider et al. [1997a; 1997b; 1998] en los cuales se investigaron los efectos de suplementos comercialmente disponibles de creatina mostraron que la ingesta de 15 a 25 g/día de creatina durante 4 a 5 semanas de entrenamiento de la fuerza produjo ganancias en la masa libre de grasa de 1.1 a 2.3 kg más (1 a 2 veces) a la observada en el grupo que ingirió cantidades isocalóricas de carbohidratos, carbohidratos y proteínas, o incluso un suplemento de carbohidratos/proteínas que contenía cromo (en forma de picolinato de cromo) y boro.
Varios estudios [Kreider, 1998b; Ziegenfuss, 1998] han demostrado incrementos significativos en el agua corporal total con la suplementación con Cr y han especulado que los cambios en el agua corporal pueden estimular la síntesis de proteínas derivando en la hipertrofia muscular [Haussinger, 1993]. Se podría especular que la creatina arrastra el agua por ósmosis hacia el compartimento intracelular potenciando de esta manera la síntesis de proteínas y de glucógeno [Haussinger, 1993].
El incremento en la fuerza muscular y en el rendimiento en levantamientos de pesas luego de la suplementación con creatina concurrente con el entrenamiento de la fuerza puede resultar de diversos mecanismos, incluyendo mayores ganancias en la masa magra corporal [Becque, 1997; Earnest, 1995; Kreider, 1996, 1997a, 1997b, 1998; Ziegenfuss, 1997; Stout, 1997; Kirksey, 1997; Vandenburghe, 1997], un efecto sobre el metabolismo proteico [Parise, 2001] incrementos en la expresión de mRNA de cadenas pesadas de miosina y de proteinas [Willoughby, 2001], una alteración en la expresión de la los factores de transcripción miogénica [Hespel, 2001]; un incremento en la actividad mitótica de las células satélite [Dangot, 2000], incrementos en la síntesis proteica secundario al incrementado inflamación celular [Bemben, 2001; Haussinger, 1993] o simplemente un incremento en la intensidad de los trabajos individuales resultante de una mejor equiparación entre la demanda y el suministro de ATP durante el ejercicio [Casey, 1996].
Debido a que la suplementación con creatina a corto plazo resulta en incrementos tanto en la fuerza en 1RM como en el rendimiento en el levantamiento de pesas [Volek, 1999], parte del efecto ergogénico de la creatina observado en los estudios de suplementación y entrenamiento de sobrecarga puede deberse a un efecto agudo de la creatina y también a la capacidad de los sujetos de entrenar con mayores cargas [Rawson and Volek, 2003]. Las contribuciones relativas de estos mecanismos siguen siendo poco claras. Arriero et al [2001] compararon las ganancias en la fuerza en 1RM luego de 4 semanas de suplementación con creatina conjuntamente y sin realizar entrenamientos de sobrecarga. La fuerza en 1RM en los ejercicios de press de banca y prensa de piernas se incrementó un 8 y un 16% respectivamente, en el grupo que solo consumió creatina y un 18 y un 42% en el grupo que realizó entrenamiento. Los resultados de este estudio sugieren que aproximadamente el 40% del incremento en la fuerza observado a lo largo de las 4 semanas de entrenamiento y suplementación con creatina se debió a los efectos agudos de la creatina sobre la producción de fuerza, con el restante 60% debido a algún otro mecanismo, presumiblemente debido a la capacidad de entrenar con mayores cargas. La mejora del sistema creatina/fosfocreatina (i.e., incremento en las reservas musculares de creatina y fosfocreatina luego de la suplementación) permite que los atletas realicen mas repeticiones por serie en un ejercicio dado (ver Tabla 2) y puede permitir una más rápida recuperación entre las series debido a la aceleración de la resíntesis de fosfocreatina [Greenhaff, 1994]. Los datos de un estudio llevado a cabo por Volek et al [1997] en el cual los sujetos que ingirieron creatina experimentaron un incremento en el volumen de levantamiento en el ejercicio de press de banca en comparación con los sujetos que ingirieron placebo, respaldan la hipótesis previamente mencionada.
Otros Suplementos
Existen otros elementos que también son promocionados como agentes anabólicos. El cromo desempeña un rol en la sensibilidad a la insulina y la insulina es una potente hormona anabólica. El ejercicio puede incrementar la excreción urinaria de cromo [Maughan, 2003]. Existen datos limitados respecto de los efectos de los suplementos de cromo en atletas, y los estudios publicados respecto de la suplementación con cromo muestran incrementos, reducciones o no muestran cambios en la masa corporal. Sin embargo, los estudios mejores controlados no muestran efecto alguno sobre la masa muscular o sobre la fuerza [Clarkson and Rawson, 1999; Nissen and Sharp, 2003]. Además, recientemente se ha expresado que el cromo, ingerido como picolinato de cromo, puede no ser enteramente seguro, habiéndose reportado diversos efectos secundarios [Vukovich, 2001].
El vanadio también se promociona como agente de estimulación de la insulina, pero existen limitados datos en animales para respaldar esta afirmación [Maughan, 2003]. Sin embargo, los datos en humanos no muestran efectos de la suplementación con vanadio sobre la composición corporal y la fuerza en atletas entrenados en al fuerza [Fawcett, 1996].
Existe cierto debate acerca de si el boro es un elemente esencial en la nutrición humana, sin embargo, se afirma que la suplementación con boro incrementa los niveles circulantes de testosterona, con el prospecto de una acción anabólica. Este resultado, sin embargo, fue obtenido en mujeres post menopáusicas que estaban ingiriendo dietas bajas en boro [Nielson, 1987], y no existen estudios que muestren que la suplementación con boro resulte en hipertrofia muscular en sujetos con una alimentación normal y con una función endocrina normal [Maughan, 2003]. La administración de suplementos de boro a un grupo de físicoculturistas varones no provocó efecto alguno sobre la testosterona circulante (libre o total) ni en el tamaño muscular o la fuerza [Ferrando and Green, 1993]. Los pocos estudios que se han llevado a cabo para examinar los efectos de la suplementación con boro sobre las alteraciones en la fuerza y la composición corporal, y los resultados de estos estudios indican que la suplementación con boro (2.5 mg/día) parece no tener impacto sobre la masa muscular o la fuerza [Ferrando and Green, 1993, 1994].
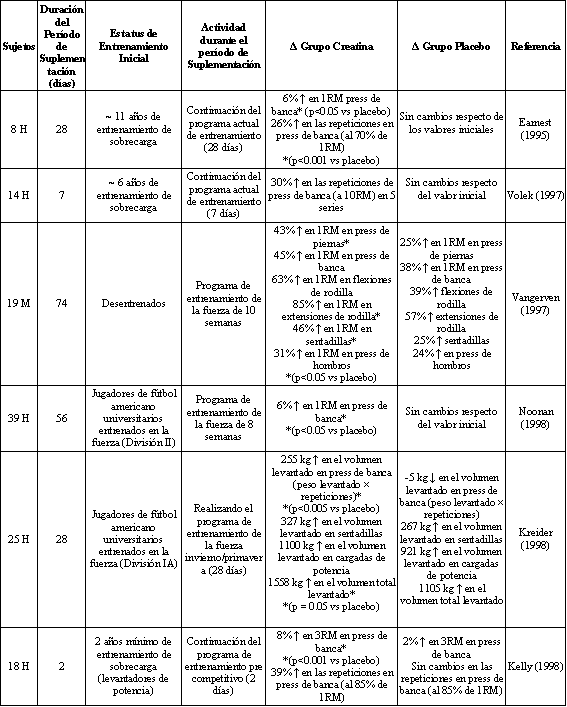
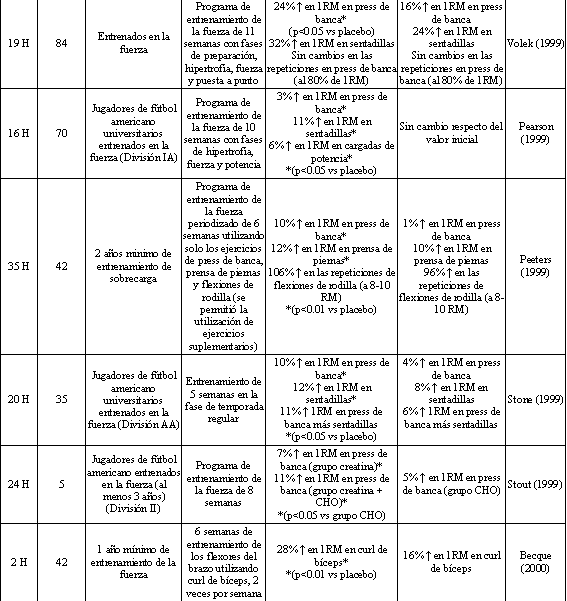
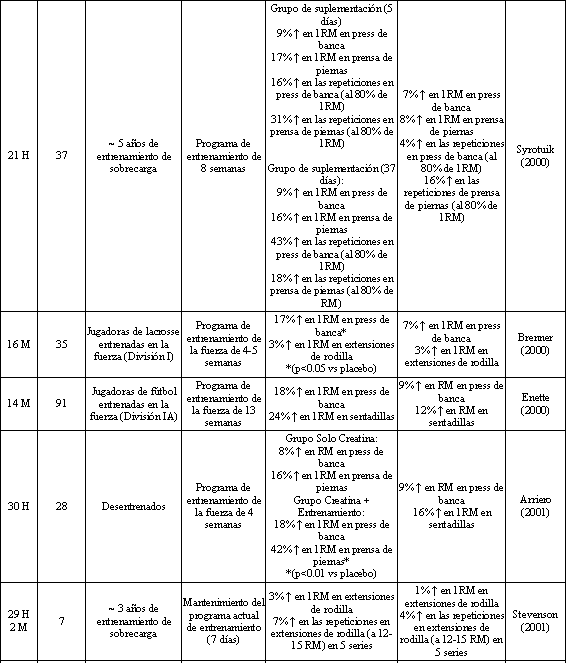
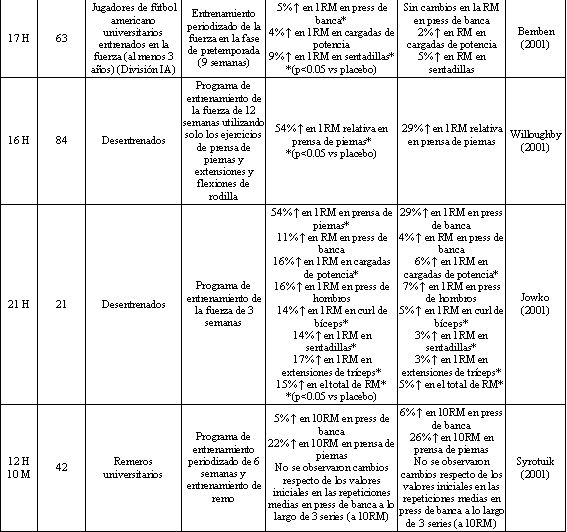

Tabla 2. Fuerza y rendimiento muscular luego de la suplementación con
creatina o placebo concurrente con un programa para el entrenamiento de la
fuerza. *Los cambios porcentuales presentados son significativamente diferentes
de los valores iniciales, y se han señalado las diferencias significativas entre
los grupos. H=Hombres, M=Mujeres.
Sulfa-polisacáridos (Inhibidores de la Miostatina)
La miostatina o factor de crecimiento 8 (GDF-8) es un factor de crecimiento que ha mostrado servir como determinante genético del limite superior del tamaño muscular y del crecimiento [Gonzalez-Cadavid, 1998]. Investigaciones recientes han indicado que la eliminación y/o la inhibición de la expresión genética de la miostatina en ratones [McPherron, 1997a] y en ganado [McPherron, 1997b; Grobet, 1997; Kambadur, 1997] promueve marcados incrementos en la masa muscular durante el crecimiento temprano. Se ha teorizado que en humanos la inhibición de la expresión genética de la miostatina podría ser una forma de evitar o enlentecer la degradación muscular en varias enfermedades, acelerar la recuperación de músculos lesionados, y/o para pomover incrementos en la masa y en la fuerza muscular de los atletas [Ivey, 2000]. Si bien estas posibilidades teóricas pueden parecer prometedoras, las investigaciones respecto del rol de la inhibición de la miostatina sobre el crecimiento y la reparación muscular se encuentra en las primeras etapas. En un estudio llevado a cabo por Ivey et al [2000] se observó que las mujeres atletas con un alelo poco común de la miostatina (un subtipo genético que puede ser más resistente a la miostatina) experimentaron mayores ganancias en la masa muscular durante el entrenamiento y menos perdida de masa muscular durante el desentrenamiento. Dicho patrón no ha sido observado en hombres con diversas historias de entrenamiento y diferentes masas musculares. Recientemente ciertas empresas dedicadas a la fabricación de suplementos nutricionales han comercializado sulfa-polisacáridos (derivados de la alga marina denominada Cytoseira canariensis) como una vía para ligar parcialmente la miostatina a proteínas séricas. Aunque la teoría es interesante y se están llevando a cabo estudios para examinarla, no existen actualmente datos que respalden el uso de los sulfa-polisacáridos como un suplemento efectivo para el incremento de la masa muscular.
Zinc/Aspartato Magnesio (ZMA)
Las formulaciones de ZMA se han vuelto recientemente populares como un suplemento que promueve el anabolismo nocturno. La teoría se basa en los estudios que sugieren que las deficiencias de zinc y magnesio pueden reducir la producción de testosterona y de factores de crecimiento tipo insulínico (IGF-1). Se ha teorizado que la suplementación con ZMA incrementa los niveles de testosterona e IGF-1 derivando en una mejor recuperación, un mayor anabolismo y en mayores ganancias de fuerza durante el entrenamiento. En respaldo a esta teoría, Brilla y Conte [2000] reportaron que la administración de una formula a base de zinc y magnesio incrementó los niveles de testosterona e IGF-1 (dos hormonas anabólicas) derivando en mayores ganancias de fuerza en jugadores de fútbol americano.
SUPLEMENTOS PARA LA PERDIDA DE PESO Y DE GRASA
Carnitina
La absorción de los ácidos grasos hacia la célula y la translocación a través de la membrana mitocondríal son pasos claves en la oxidación de las grasa. La carnitina se combina con la acetil-coenzima A (acetil-CoA) en el citoplasma celular y permite que los ácidos grasos ingresen a la mitocondría. El primer paso es catalizado por la carnitina palmitoil transferasa 1 (CPT 1) y el transporte transmembrana es facilitado por la acetilcarnitina transferasa. Dentro de la mitocondria, la acción de la carnitina palmitoil transferasa 2 (CPT2) regenera la carnitina libre y el acetil-CoA es librado hacia la beta-oxidación. Dentro de la mitocondria la carnitina también funciona como regulador de la concentración de acetil-CoA y de la concentración de CoA libre. La CoA libre esta involucrada en la reacción de la piruvato deshidrogenasa y también en el proceso de β-oxidación, por lo cual desempeña un papel clave en la integración de la oxidación de grasas y carbohidratos. Se ha propuesto que un incremento en la disponibilidad de carnitina dentro de la mitocondria podría permitir que la célula mantenga una alta concentración de CoA libre, con un efecto estimulador sobre el metabolismo oxidativo [Maughan, 2003]. Debido al papel que desempeña la carnitina en la oxidación de las grasas y los carbohidratos, se ha propuesto que la suplementación con carnitina podría incrementar el rendimiento durante el ejercicio. En base a esta lógica, la carnitina se vende ampliamente como un suplemento para atletas de deportes de resistencia. Sin embargo, no hay buena evidencia que indique que halla una deficiencia de carnitina en la población general o entre los atletas. La carnitina está presente en las carnes rojas y en productos lácteos, por lo cual se piensa que los sujetos que siguen una dieta vegetariana podrían estar en riesgo de sufrir deficiencia de carnitina. Sin embargo la carnitina puede ser sintetizada a partir de lisina y metionina en el hígado y los riñones. Barnett et al [1994] y Vukovich et al [1994] han reportado que la suplementación a corto plazo con carnitina (4-6 g/día durante 7-14 días) no tiene efectos sobre la concentración muscular de carnitina o sobre las respuestas metabólicas durante el ejercicio. Incluso cuando se estimuló la movilización de ácidos grasos mediante comidas altas en grasas o mediante la administración de heparina, no se observaron efectos de la suplementación con carnitina sobre la oxidación de las grasas [Vukovich, 1994].
En contraste a estos hallazgos negativos, existen algunos reportes publicados que sugieren que la suplementación con creatina incrementa la contribución de los ácidos grasos al metabolismo oxidativo y por ende promueven la utilización de las reservas corporales de grasas [Maughan, 2003]. En una revisión de la literatura, Spriet [1997] identificó ocho estudios que examinaron los efectos de la suplementación sobre las respuestas metabólicas a ejercicios de resistencia, y halló que tres de estos estudios reportaron un incremento en la oxidación de las grasas. Por lo tanto, si bien existen ciertos datos que indican que la suplementación con carnitina puede ser beneficiosa para algunas poblaciones, la mayoría de los estudios bien controlados indican que la suplementación con carnitina no afecta el contenido muscular de carnitina, el metabolismo de las grasas y/o la pérdida de peso corporal en sujetos entrenados o en sujetos con sobrepeso [Brass, 2000]. Por ejemplo, Villani et al. [2000] reportaron que la suplementación con carnitina (2 g/día durante 8 semanas) no afecto la pérdida de peso, la composición corporal o los marcadores del metabolismo graso en mujeres con sobrepeso.
Efedrina, Cafeína y Salicina
Los suplementos termogénicos son diseñados para estimular el metabolismo incrementando así el gasto energético y la pérdida de peso. Estos característicamente contienen “ECA” alcaloides de la efedra (e.g., Ma Huang, 1R, 2S norepinefrina HCl, Sida Crdifolia), cafeína (e.g., guaraná, Bissey Nut, Kola) y aspirina/salicina (e.g., Extracto de Corteza de Sauce). Se han llevado a cabo numerosas investigaciones para evaluar la seguridad y la eficacia de los suplementos tipo ECA. Estos estudios muestran que la utilización de fuentes sintéticas o vegetales de efedrina y cafeína (EC) promueven la pérdida de hasta 2 libras mensuales de peso mientras a la vez que la dieta (con o sin ejercicio) y la suplementación EC es generalmente bien tolerada por individuos saludables [Boozer, 2001, 2002a, 2002b; Molnar, 1993, 2000; Greenway, 1999a, 1999b, 2000, 2001; Bent, 2003].
Por ejemplo, Boozer et al. [2001a] reportaron que 8 semanas de suplementación con efedrina (72 mg/día) y cafeína (240 mg/día) promovió una pérdida de 9 libras en la masa corporal y una pérdida del 2.1% en la grasa corporal con efectos secundarios menores. Molnar et al [2000] reportaron que niños con sobrepeso tratados durante 20 semanas con efedrina y cafeína experimentaron una pérdida del 14.4% en la masa corporal y una reducción del 6.6% en la grasa corporal sin experimentar efectos secundarios. Interesantemente, Greenway et al [1999a] reportaron que la suplementación con EC era la vía con menor costo/beneficio para reducir el peso corporal, el riesgo de enfermedades cardíacas y disminuir el colesterol LDL en comparación con otras drogas para la reducción del peso corporal (fenfluramina con mazindol o fentermina). Por último, Boozer et al [2002a] reportaron que 6 meses de suplementación con EC promovió la pérdida de peso sin provocar efectos negativos clínicamente significativos en adultos saludables. A pesar de estos hallazgos, en Estados Unidos, la Administración de Alimentos y Drogas (FDA, “Food and Drug Administration”) ha prohibido la venta de suplementos que contienen efedra. Esto se basó en reportes de eventos adversos que sugieren un vínculo entre la ingesta de efedra y diversas complicaciones médicas (e.g., alta presión sanguínea, elevación de la frecuencia cardíaca, arritmias, muerte súbita, etc.) [Fleming, 2000, Bent, 2003].
Calcio
La investigación ha indicado que el calcio modula la 1,25-dihidroxivitamina D cuyo rol es el de regular los niveles intracelulares de calcio en las células del tejido adiposo [Zemel, 2002, 2003a, 2003b]. Se ha demostrado que el calcio dietario suprime el metabolismo de las grasas y la ganancia de peso durante períodos de altas ingestas calóricas [Zemel, 2000, 2002]. Además, el incremento en la ingesta de calcio ha mostrado incrementar el metabolismo graso y preservar la termogénesis durante la restricción calórica [Zemel, 2000, 2002]. En respaldo a esta teoría, Davies et al [2000] reportaron que el calcio dietario correlacionaba negativamente con el peso y que la suplementación con calcio (1.000 mg/día) explicaba la pérdida de peso de 8 kg en un período de semanas. Asimismo, en otro estudio llevado a cabo por Zemel et al. [2002b] se reportó que la suplementación con calcio (800 mg/día) o la alta ingesta de calcio en la dieta (1200 – 1300 mg/día) durante un programa para la pérdida de peso corporal de 24 semanas, promovió pérdidas de peso (26-70%) y de masa grasa determinada por DEXA (absorciometría dual de rayos X) (38-64%) significativamente mayores en comparación con los sujetos que tuvieron una ingesta de calcio baja (400-500 mg/día). Estos hallazgos sugieren una fuerte relación entre la ingesta de calcio y la pérdida de grasa.
Fosfatos
El rol del fosfato de sodio y del fosfato de calcio sobre el metabolismo energético y el rendimiento en el ejercicio ha sido estudiado por décadas [Kreider, 1999]. Estos estudios han revelado que la suplementación con fosfato de sodio parece poseer propiedades ergogénicas, particularmente en eventos de resistencia [Kreider, 1990, 1992]. Kaciuba-Uscilko et al [1993] reportaron que la suplementación con fosfatos durante un programa para la pérdida de peso de 4 semanas incrementó la tasa metabólica basal y el índice de intercambio respiratorio (sugiriendo una mayor utilización de carbohidratos y un mayor gasto calórico) durante la realización de ciclismo submáximo. Además, Nazar et al. [1996] reportaron que la suplementación con fosfatos durante un programa para la pérdida de peso de 8 semanas de duración incrementó la tasa metabólica basal en un 12-19% y evitó la declinación normal en la actividad de las hormonas tiroideas. Consecuentemente, es posible que los fosfatos sirvan como un potencial nutriente termogénico en suplementos que no contienen efedrina. De todas maneras es necesaria mayor investigación para evaluar esta hipótesis.
Extracto de Te Verde
El té verde es uno de los suplementos herbales más interesantes y se ha sugerido recientemente que afecta la pérdida de peso. El té verde contiene altas cantidades de cafeína y catechin polifenoles. La investigación sugiere que el catechin polifenol posee propiedades antioxidantes [Nakagawa, 1999]. Además se ha propuesto que el té verde incrementa el gasto energético estimulando la termogénesis del el tejido adiposo marrón. En respaldo a esta teoría, Dulloo et al [1999, 2000] reportaron que la suplementación con té verde en combinación con cafeína incrementó significativamente el gasto energético de 24 horas y la utilización de grasas en humanos. El efecto termogénico de la suplementación con té verde fue mucho mayor que el observado con una cantidad equivalente de cafeína, lo que sugiere un efecto sinérgico.
Piruvato de Calcio
La razón principal del uso del piruvato de calcio se basa en los estudios de principios de los 90’ en los cuales se reportó que la suplementación con piruvato de calcio (16-25 g/día con o sin fosfato de dihidroxiacetona [DHAP]) promovía la pérdida de grasa en pacientes con sobrepeso u obesos que seguían un programa para la perdida de peso corporal [Stanko, 1992, 1994, 1996]. Aunque el mecanismo responsable de estos hallazgos no es claro, los investigadores especulan que puede estar relacionado con la supresión del apetito y/o alteración del metabolismo de las grasas y los carbohidratos. Si bien existen datos que indican que la suplementación con piruvato de calcio en altas dosis (6-16 g/día) puede incrementar la pérdida de grasas, no existe evidencia que indique que la ingesta de las dosis característicamente halladas en los suplementos de piruvato de calcio (0.5 a 2 g/día) tenga algún efecto sobre la composición corporal [Kalman, 1999; Stone, 1999].
Fosfatidil Colina (Lecitina)
La colina es considerada un nutriente esencial necesario para la preservación de las membranas celulares y para facilitar el movimiento de las grasas dentro y fuera de las células. También es un componente del neurotransmisor acetilcolina y es necesario para el normal funcionamiento del cerebro, particularmente en los infantes. Por esta razón, se ha reportado que la fosfatidil colina es un suplemento potencialmente efectivo para promover la pérdida de grasas y para mejorar la función neuromuscular. Ciertos datos de estudios llevados a cabo con animales respaldan el valor potencial de la fosfatidil colina como suplemento para la pérdida de peso [Rama Rao, 2001]. También existe cierto interés por determinar el potencial ergogénico de la suplementación con fosfatidil colina durante ejercicios de resistencia [Buchman, 1999, 2000]. Sin embargo, actualmente no se conoce si la suplementación con fosfatidil colina afecta la composición corporal en humanos.
SUPLEMENTOS QUE PROMUEVEN EL SUMINISTRO ENERGETICO
Una forma de fatiga inducida por el ejercicio es la que ocurre cuando la tasa de hidrólisis de ATP en los músculos activos supera la tasa a la cual el ATP puede ser resintetizado. De esto se desprende que el comienzo de la fatiga puede retrasarse y que el rendimiento puede mejorarse si es posible mantener una alta tasa de resíntesis de ATP. Los suplementos que se afirman mejoran el rendimiento incrementando el suministro energético y retrasando la fatiga incluyen el bicarbonato, la cafeína, la creatina, el hierro, el magnesio y la ribosa.
Bicarbonato
Durante los ejercicios que provocan la instalación de la fatiga dentro de los pocos minutos, la glucólisis anaeróbica hace la mayor contribución desde el punto de vista del metabolismo energético. Aunque la glucólisis permite que se puedan alcanzar mayores tasas de resíntesis de ATP por parte del metabolismo aeróbico, la capacidad de este sistema es limitada, y la fatiga es inevitable cuando se requiere una alta tasa de glucólisis anaeróbica. La acidosis metabólica que acompaña a la glucólisis ha sido implicada en el proceso de fatiga, tanto por la inhibición de las enzimas clave de la glucólisis, como interfiriendo con el transporte y la unión del calcio o por el efecto directo sobre la interacción de la actina y la miosina. Varios estudios han tratado de determinar los efectos de la alcalosis metabólica (comúnmente inducida por la ingesta de bicarbonato o citrato de sodio) sobre el rendimiento en ejercicios de alta intensidad y corta duración, pero los resultados no son para nada consistentes o concluyentes [Maughan, 1999; McNaughton, 2000]. En un estudio diseñado para estimular la competición deportiva, corredores de distancia entrenados pero no de elite (mejor marca en los 800 mts, aproximadamente 2 min 5 seg) realizaron una carrera simulada de 800 metros. En la condición de alcalosis, los corredores cubrieron la distancia casi 3 s más rápido que los sujetos de los grupos placebo y control [Wilkes, 1983]. En un estudio llevado a cabo por Bird et al [1995] se observaron mejoras similares (3-4 s) en una distancia de 1500 m. Aunque estos efectos sobre el rendimiento parecen pequeños, son de considerable significación para el deportista, para quién una mejora de incluso una fracción de segundo en estos eventos es un logro importante.
Las razones de los resultados inconsistentes en la literatura no son claras pero pueden deberse en parte a las variaciones en la intensidad y en la duración de los test de ejercicio utilizados, la naturaleza de la tarea deportiva, las dosis de bicarbonato de sodio administradas y el tiempo de retraso entre la administración del bicarbonato y el comienzo del test de ejercicio [Maughan, 2003]. El rendimiento ha sido monitoreado con duraciones de ejercicio que están en el rango de los pocos segundos a más de 1 hora, y durante ejercicios continuos, progresivos e intermitentes y también durante contracciones isométricas sostenidas. Por lo tanto se observa que no hay un patrón claro de duración de ejercicio entre los estudios que observaron un efecto positivo y los estudios que no observaron efecto alguno. La carga de bicarbonato (e.g., 0.3 g/kg 2 veces por día durante 5 días) ha mostrado ser una forma efectiva para amortiguar la acidez durante ejercicios de alta intensidad de entre 1-3 minutos de duración [McNaughton, 1999; Applegate, 1999; Kronfeld, 1994; Kraemer, 1995]. Esto podría incrementar la capacidad de ejercicio en eventos tales como los 400 o los 800 m de carrera o los 1000 y 200 m en natación [Matson, 1993]. Horswill et al [1988] examinaron los efectos de la ingesta de 0.1-0.2 g de bicarbonato por kg de peso corporal sobre el rendimiento en una prueba de esprint en cicloergómetro de 2 min de duración. Estos investigadores no hallaron mejoras en el rendimiento aun cuando se observó una elevación en la concentración sanguínea de bicarbonato. McKenzie et al [1986] reportaron que una dosis de 0.3 g/kg de peso corporal no era más efectiva que una dosis de 1.5 g/kg de peso corporal.
No obstante esto, existen varios problemas potenciales asociados con la utilización de altas dosis de bicarbonato. Con frecuencia se reportan vómitos y diarrea como resultado de la ingesta de incluso dosis pequeñas de bicarbonato, limitando de esta manera cualquier mejora en el rendimiento entre aquellos individuos susceptibles a sufrir problemas gastrointestinales.
La administración de citrato de sodio, que también resulta en la alcalinización del fluido extracelular, ha mostrado provocar incrementos en el pico de potencia y en el trabajo total en un test de ejercicio de 60 segundos, pero sin provocar síntomas gastrointestinales adversos [McNaughton, 2000].
Hay varios mecanismos propuestos acerca de cómo la ingesta de citrato, mejora la fisiología ácido-base sanguínea. Por ejemplo, los mecanismos propuestos para el incremento del ph sanguíneo y el bicarbonato, luego de la ingesta de citrato, incluyen:
- Amortiguación celular y sanguínea de protones [McNaughton, 1990].
- Consumo de protones durante la oxidación del citrato en el ciclo de los ácidos tricarboxílicos [Tiryaki, 1995].
- Producción de bicarbonato como un producto del metabolismo [Parry-Billings, 1986; Tiryaki, 1995].
- La carga negativa del citrato aumenta el gradiente de carga entre la sangre y las células, causando una disminución de los protones y un incremento del bicarbonato [Kowalchuk, 1989].
- Potenciación de la inhibición de la fosfofructo quinasa por parte del ATP, lo que causa una disminución de la velocidad de la glucólisis y de la producción de protones [Kowalchuk, 1989].
Creatina
La creatina ha sido utilizada por muchos atletas exitosos, particularmente por los atletas de pista y campo, pero también ha sido utilizada en muchos otros deportes. Hay muchas excelentes revisiones acerca de los efectos de la suplementación con creatina, pero la imagen cambia rápidamente a medida que emerge nueva información en esta área.
Las mayores concentraciones de creatina se hallan en los músculos esqueléticos, y aproximadamente dos tercios del total se encuentra como fosfato de creatina. El fosfato de creatina es capaz de regenerar rápidamente el ATP dentro del citoplasma celular, pero esta disponible solo en pequeñas cantidades [Maughan, 2003]. Muchos autores que han revisado la literatura publicada han concluido que la evidencia disponible respalda los beneficios de la suplementación con creatina sobre el rendimiento en ejercicios de alta intensidad y corta duración [Greenhaff, 2000]. De tres meta-análisis publicados, dos han concluido que la suplementación con creatina tiene efectos positivos sobre las ganancias de fuerza, potencia y masa magra [Branch, 2003; Nissen and Sharp, 2003] mientras que el tercero [Misic and Kelley, 2002] concluyó que no existe efecto alguno. Las razones de estas discrepancias no son completamente claras. Los efectos de la suplementación con creatina son observados en las últimas etapas de esfuerzos múltiples de corta duración con limitada recuperación, pero a veces es posible observar mejoras en esprints únicos de menos de 30 s de duración. Existe poca información respecto de los efectos de la suplementación con creatina sobre el rendimiento en ejercicios más prolongados, pero hay pocas razones para esperar un efecto positivo [Maughan, 2003]. Las investigaciones que han estudiado los beneficios potenciales de la carga con Cr sobre el rendimiento de resistencia son relativamente escasas. En hombres entrenados [Balsom, 1993], se ha observado una reducción en el rendimiento de resistencia, lo cual fue atribuido al peso ganado (+ 0.9kg). En este estudio, los sujetos completaron una prueba de 6 km al aire libre y una carrera en cinta ergométrica hasta el agotamiento con una carga equivalente al 120 % del consumo máximo de oxígeno (VO2 máx.), lo cual fue repetido luego de la administración de Cr (6 días de carga con 5 g/día) o de la administración de placebo. No se observaron diferencias en el pico de VO2 entre los grupos Cr y placebo respecto de la prueba inicial, y los sujetos corrieron a una velocidad significativamente más lenta (p<0.05) luego de la administración de Cr (23.79±0.85 minutos vs. 23.36±0.82 minutos) en comparación con la administración de placebo (23.76±0.80 minutos vs. 23.92±0.79 minutos). En un estudio más reciente (8), 5 días de suplementación con Cr (6 g/día) no alteraron las funciones cardiovasculares, el VO2 o la frecuencia cardíaca durante una prueba de resistencia de 30 minutos seguida de ejercicios intervalados. Consecuentemente, es probable que la carga con Cr no solo no altere el rendimiento de resistencia, sino que potencialmente reduzca el rendimiento debido a la ganancia de peso. A pesar de que los resultados refutan cualquier efecto ergogénico de la carga con Cr sobre el rendimiento en carreras de fondo, es posible que la suplementación pueda incrementar la resistencia y/o la cantidad de entrenamiento fraccionado completado por los corredores lo cual puede indirectamente incrementar el rendimiento.
El mecanismo por el cual la suplementación con creatina incrementa el rendimiento no es muy claro aunque si es claro que este efecto está relacionado con un incremento en las reservas musculares de fosfato de creatina. La tasa de resíntesis de fosfato de creatina luego de la realización de ejercicios de alta intensidad es incrementada con la suplementación con creatina en dosis altas [Greenhaff, 1994]. Esto permite una mayor recuperación luego de la realización de esprints y también permite que se realice mas trabajo durante cada esfuerzo subsiguiente de alta intensidad (ver discusión de suplementación con creatina y entrenamiento de la fuerza).
Ribosa
La ribosa es un carbohidrato de 3 carbonos que esta involucrado en la síntesis del trifosfato de adenosina (ATP) en el músculo. Los estudios clínicos han mostrado que la suplementación con ribosa puede incrementar la capacidad de ejercicio en pacientes cardíacos [Tullson, 1991; Gross, 1991; Wagner, 1991; Pauly, 2000]. Por esta razón se ha sugerido que la ribosa puede ser una ayuda ergogénica para los deportistas. Aunque se requiere de mayor investigación en este aspecto, la mayoría de los estudios no muestran beneficios ergogénicos de la suplementación con ribosa sobre la capacidad de ejercicio en poblaciones de sujetos saludables desentrenados o entrenados [Op ‘t Eijnde, 2001; Berardi, 2003; Kreider, 2003].
Cafeína
Los humanos tienen una muy larga historia de consumo de cafeína (5, 8-10), es la droga más comúnmente consumida en el mundo y el riesgo de salud es mínimo. Las concentraciones fisiológicas de cafeína son normalmente menores a 70 μmol/L; las concentraciones plasmáticas de 20 a 50 μmol/L son comunes. Sin embargo, las concentraciones empleadas en la mayoría de las investigaciones in vitro varían entre 500 y 5000 μmol/L. La significancia fisiológica de tales estudios no esta clara. Mientras que han sido identificados varios modos de acción de la cafeína, el único que es importante, dentro del rango de concentración fisiológico de la cafeína, es la inhibición de los receptores de adenosina. La cafeína es muy similar en estructura a la adenosina y pude unirse a los receptores de la membrana de la célula para la adenosina, así bloqueando su interacción. Los receptores de adenosina son encontrados en la mayoría de los tejidos, incluyendo al cerebro, corazón, músculo liso, adipositos y músculo esquelético (aunque la naturaleza de estos receptores en el músculo esquelético es pobremente entendida). La naturaleza ubicua y los variados tipos de receptores de adenosina facilitan que la cafeína afecte simultáneamente una variedad de tejidos, resultando en un amplio rango de respuestas que frecuentemente interactúan entre si.
Los efectos ergogénicos de la cafeína están bien documentados. Desde los estudios iniciales de Rivers y Webber [1907], numerosos reportes han mostrado que la ingesta de cafeína puede incrementar el rendmietno durante el ejercicio. Los trabajos en el laboratorio de Costill [Costill, 1978; Ivy, 1977: Essig, 1980] en la década del 70’ renovaron el interés por la cafeína, y los trabajos de Graham. y Spriet [1991] y de Spiet [1992] a principios de los 90’ demostraron más allá de cualquier duda que la ingesta de cafeína mejoraba substancialmente la resistencia. Casi 20 artículos de revisión respecto de los efectos ergogénicos de la cafeína en diversos modos de ejercicio han sido publicados en los últimos 15 años [Graham, 1994, 1997, 2001a, 2001b; Nehlig, 1994; Spriet, 1995, 1999, 2000, 2002; Wilcox, 1990; Tarnopolsky, 2000; Magkos, 2004]. La observación más consistente es que la cafeína incrementa el tiempo hasta el agotamiento durante series de ejercicio submáximo de entre 30-60 min de duración. La velocidad y la producción de potencia durante dichas actividades también pueden incrementar. La resistencia aeróbica en eventos de menor duración (5-25 min) parece verse mejorada o no afectada por la ingesta de cafeína. En series de ejercicio más intensos y más cortos los efectos positivos parecen menos frecuentes, y lo mismo ocurre con ejercicios progresivos. Por último, los pocos datos disponibles sugieren que la cafeína puede incrementar ciertos aspectos de la función neuromuscular en humanos [Magkos, 2004]
La creciente evidencia de un efecto positivo de la cafeína sobre el rendimiento en ausencia de un efecto ahorrador de glucógeno, y de los efectos sobre el ejercicio de alta intensidad en donde la disponibilidad de glucógeno no es un factor limitante, ha estimulado la investigación de mecanismos alternativos de acción. Existe evidencia de que la cafeína actúa de diversas formas sobre los músculos esqueléticos. Puede afectar la actividad de la Na+ /K+ ATPasa y la localización del sitio intracelular de unión del calcio, y puede provocar una elevación en los niveles intracelulares de AMP cíclico como resultado de la inhibición de la acción de la fosfodiesterasa, y puede tener un efecto directo sobre diversas enzimas, incluyendo la glucógeno fosforilasa [Spriet, 1997; Graham, 2001]. Sin embargo no es claro si todos estos efectos pueden producirse con dosis moderadas de cafeína [Maughan, 2003].
REQUERIMIENTOS DE LIQUIDOS Y NUTRIENTES DURANTE Y DESPUES DEL EJERCICIO
La actividad física resulta en un incremento en la necesidades de agua para equilibrar las pérdidas de sudor para el intercambio evaporativo de sudor [Sawka, 1996]. Los estudios acerca del intercambio de agua corporal han demostrado que grandes volúmenes diarios (50 km de ciclismo) o semanales (100 km de carrera) de actividad física en ambientes templados incrementan el flujo de agua en 1.2 a 1.4 L/día, que se deben principalmente al volumen de sudor perdido y a su reemplazo [Leiper, 1996, 2001]. Estas mismas actividades llevadas a cabo en un ambiente caluroso podrían exacerbar la producción de sudor.
Por lo tanto, es de esperar que las pérdidas de agua, y con ello las necesidades de agua, variarán considerablemente entre las personas moderadamente activas. La magnitud de la pérdida de sudor durante el ejercicio en ambientes calurosos depende principalmente de la intensidad del ejercicio y de la duración [Sawka, 1996]. El calor producido por el metabolismo es equilibrado por la pérdida de calor tanto seca como evaporativa (sudoración), pero las muy altas tasas metabólicas asociadas con ambientes calurosos hacen que los requerimientos biofísicos para el enfriamiento evaporativo sean mayores [Nielsen, 1938], lo que deriva en mayores pérdidas de sudor y consecuentemente mayores requerimientos de agua. Por lo tanto es de esperarse que los atletas requieran de una cantidad relativamente mayor de fluidos para cubrir las mayores pérdidas de agua por evaporación y de esta manera mantener el balance hídrico [Sawka, 2005]. Pero, nuevamente hay que señalar que las tasas de sudoración varían ampliamente entre los diferentes deportes e incluso entre las diferentes posiciones de los jugadores [Sawka, 2005]. Incluso bajo condiciones estandarizadas en poblaciones deportivas más o menos homogéneas, la variabilidad entre los sujetos en la tasa de sudoración es significativa [Maughan, 2004].
Las pérdidas de sudor con el ejercicio agudo comúnmente caen dentro del rango de 1 a 2 L/h para un equipo [Burke, 1997] y para deportes individuales de resistencia [Rehrer, 1996]. Aunque dichas tasas raramente pueden sostenerse por más de algunas horas debido al nivel de esfuerzo requerido, dichas pérdidas pueden afectar marcadamente los requerimientos de agua. Por ejemplo, el fútbol es un deporte de resistencia que implica la realización de actividad física durante 90 minutos de intensidades variadas [Maughan, 1994]. La distancia total cubierta por un jugador durante un partido es ~ 8-13 km, y la tasa media de gasto energético es de ~67 kJ/min (16 kcal/min) correspondiente a un consumo de oxígeno de aproximadamente el 75% [Reilly, 1990]. Ekblom [1986] ha reportado un a pérdida de peso de 1.0 a 2.5 kg durante partidos jugados en climas templados, siendo estas pérdidas mayores en partidos de nivel internacional y menores en partidos regulares. Leatt [1986] reportó una pérdida de peso de 1.0 kg (1.4% del peso corporal), en un estudio donde los jugadores consumieron 1 litro de fluido durante el partido, indicando un total de pérdida de sudor cercana a los 2 litros. También fueron reportadas por Bangsbo [1994], grandes pérdidas de sudor (3.5 L), en algunos individuos. Está claro que tales pérdidas podrían desmejorar seriamente el rendimiento físico y cognitivo, por lo cual el reemplazo de fluidos es imperante, si se quiere mantener el rendimiento.
De lo expuesto hasta ahora es claro que el rendimiento durante el ejercicio es óptimo cuando los atletas mantienen el equilibrio hídrico durante el ejercicio; a la inversa, el rendimiento durante el ejercicio se reduce con la deshidratación progresiva [Barr, 1991; Montain and Coyle, 1992; Walsh, 1994; Below, 1995; McConnell, 1997]. En concordancia, los deportistas deberían intentar mantenerse bien hidratados antes y durante el ejercicio, y como veremos más adelante después del ejercicio. La posición del Colegio Americano de Medicina del Deporte acerca del Ejercicio y el Reemplazo de Fluidos [1996] y la posición de la Asociación Nacional de Entrenadores Deportivos (NATA) sobre el Reemplazo de Fluidos para Deportistas [2000] proveen una revisión extensa de las investigaciones y recomendaciones para el mantenimiento de la hidratación durante el ejercicio. La siguiente información resume los puntos clave y provee recomendaciones para la realización de ejercicios en condiciones ambientales especiales.
Recomendaciones para la Ingesta de Fluidos y Electrolitos antes del Ejercicio
Los atletas deberían estar bien hidratados cuando comienzan el ejercicio. Además de beber cantidades generosas de fluidos en las 24 h previas a la sesión de ejercicio, el ACSM y la NATA recomiendan beber de 400 a 600 mL de fluidos 2-3 horas antes del ejercicio [ACSM, 1996; Casa, 2000]. Dicha práctica debería optimizar el estátus de hidratación permitiendo suficiente tiempo para que cualquier exceso de fluido sea excretado como orina antes del comienzo del ejercicio.
El tema de la hiperhidratación pre ejercicio ha sido investigado en la última década. En individuos saludables, los riñones excretan cualquier exceso de agua, y por lo tanto la ingesta en exceso de fluidos antes del ejercicio es generalmente inefectiva para inducir una hiperhidratación pre ejercicio [Shirreffs, 2004]. También se ha investigado la ingesta de soluciones salinas o con glicerol como posibles medios para minimizar la diuresis característica cuando un individuo euhidratado ingiere un exceso de agua. Se ha observado una hiperhidratación temporaria cuando se ingieren bebidas con altas concentraciones (4100 mmol/L) de sodio [Fortney et al., 1984], pero existen problemas con relacionados con el sabor y con la provocación de nauseas.
El glicerol ha mostrado ser un efectivo agente para la hiperhidratación. Varios estudios han sugerido que la ingesta de 1.0-1.5 g de glicerol/kg de masa corporal, conjuntamente con grandes volúmenes de agua, pueden incrementar significativamente la retención de agua e incrementar el tiempo de ciclismo hasta el agotamiento [Riedesel, 1987; Lyons, 1990; Montner, 1996; Hitchins, 1999]. Sin embargo, otros investigadores [Latzka, 1997, 1998] no han observado diferencias en los parámetros termorreguladores o de rendimiento. Además, existen numerosos reportes acerca de los efectos secundarios de la ingesta de glicerol [Latzka and Sawka, 2000] van en contra de este método de hiperhidratación. De hecho, en una reciente revisión acerca de la hiperhidratación con glicerol se llegó a la conclusión que “si se mantiene la euhidratación durante el ejercicio en el calor entonces la hiperhidratación (pre ejercicio) parece no tener una ventaja significativa [Latzka and Sawka, 2000]. Sin embargo hay que enfatizar que “la práctica de beber en las horas previas al ejercicio es efectiva para asegurar la euhidratación antes del ejercicio”.
Hidratación durante el Ejercicio
Los atletas deberían intentar beber suficiente fluido como para mantener el balance hídrico, ya que incluso una deshidratación parcial puede comprometer el rendimiento. El comienzo de la fatiga durante ejercicios submáximos prolongados de alta intensidad está asociada con (a) la reducción, y hasta la depleción, del glucógeno muscular [Bergstrom et al 1967], (b) la reducción de la concentración de glucosa en sangre [Coyle et al 1986] y (c) la deshidratación [Sawka and Pandolf 1990]. La ingesta de fluidos con carbohidratos y electrolitos durante el ejercicio prolongado puede evitar la deshidratación y atenuar los efectos de la pérdida de fluidos sobre la función cardiovascular y el rendimiento durante el ejercicio [Montain and Coyle, 1992], y retrasar el comienzo de la fatiga [Coyle, 1983; Coggan and Coyle 1989]. La mejora en la capacidad de resistencia puede ser una consecuencia de un efecto ahorrador de glucógeno [Tzintzas et al 1996].
Si no se puede mantener el balance hídrico, se debería ingerir la máxima cantidad de liquido que pueda ser tolerado [ADA/DC/ACSM, 2000]. La hidratación óptima puede ser facilitada consumiendo entre 150 a 350 mL de fluidos a intervalos de 15 a 20 min, comenzando cuando comienza el ejercicio [ACSM, 1996]. Las bebidas que contienen carbohidratos en concentraciones de 4% a 8% son recomendadas para eventos de ejercicio intensos que duran más de 1 hora [ACSM, 1996]. Estas bebidas también son adecuadas para la hidratación en eventos que duren menos de 1 hora, aunque el agua común es apropiada para estas condiciones.Varios estudios han reportado que la ingesta de soluciones a base de carbohidratos y electrolitos durante ejercicios intermitentes prolongados puede retrasar el agotamiento. Sin embargo, pocos estudios se han preocupado en los efectos de la ingesta de CHO y electrolitos durante los períodos de reposo y recuperación. En estudios de laboratorio llevados a cabo por Tabata et al (1984) y Coggan y Coyle [1989] se pudo posponer el agotamiento luego de un protocolo específico de ejercicio por medio de la suplementación oral con una solución de glucosa. Estudios de campo, Kirkendall et al [1988] y Leatt y Jacobs [1989] reportaron que los efectos beneficiosos de la ingesta de CHO y electrolitos sobre el rendimiento durante el ejercicio se producen por medio de un efecto ahorrador de glucógeno muscular. Por ejemplo, Bangsbo [1994] consideró que hacia el final de un partido de fútbol, la mayoría de los jugadores tendrían una depleción de las reservas musculares de glucógeno y que la ingesta de CHO y electrolitos durante el entretiempo podría producir el ahorro del glucógeno muscular.
Parece no haber necesidad fisiológica de reemplazar electrolitos durante una única sesión de ejercicios de duración moderada (e.g., menos de 3 a 4 horas), particularmente si el sodio estuvo presente en la comida previa. Sin embargo, la inclusión de sodio en cantidades de 0.5 a 0.7 g/L es recomendada durante la realización de ejercicios de duración mayor a 1 h. Debería señalarse que esta cantidad de sodio excede la cantidad característicamente disponible en bebidas comerciales. La inclusión de sodio además puede ayudar a prevenir la hiponatremia en personas susceptibles [ACSM, 1996; Vrigens, 1999]. Aunque la mayoría de los atletas que beben mas fluido del que pierden por el sudor excretan el exceso por el sudor, en algunas personas el fluido es retenido [Speedy, 1999]. Si el fluido contiene sodio, se puede ayudar a prevenir la dilución de los niveles de sodio sérico, y por lo tanto reducir el riesgo de hiponatremia. Limitar la ingesta de fluidos para que no exceda la tasa de sudoración también puede ayudar a reducir el riesgo de hiponatremia.
Rehidratación Post-Ejercicio
Los principales factores que influencian la rehidratación post ejercicio son el volumen y la composición del fluido consumido. El volumen consumido estará influenciado por varios factores, incluyendo el sabor de la bebida y sus efectos sobre el mecanismo de la sed [Shirreffs, 2004]. La ingesta de alimentos sólidos, y la composición de los alimentos, también puede ser importante, pero existen muchas circunstancia en las cuales se evita el consumo de alimentos sólidos entre sesiones de ejercicio o inmediatamente después del ejercicio [Shirreffs, 2004].
Se debería consumir hasta el 150% del peso perdido durante una sesión de ejercicio puede ser necesario para cubrir las pérdidas por sudoración más la producción de orina [Shirreffs, 1996]. La inclusión de sodio en o con los fluidos consumidos post ejercicio reduce la diuresis que se produce cuando se consume agua común. El sodio ayuda al proceso de rehidratación manteniendo la osmolalidad del plasma y por lo tanto el deseo de beber. Debido a que la mayoría de las bebidas deportivas no contienen suficiente sodio como para optimizar el reemplazo de fluidos post ejercicio, los atletas pueden rehidratarse consumiendo conjuntamente una comida que contenga sodio [Maughan, 1996].
Contenido de las Bebidas
Sodio
El agua común no es la bebida ideal para la rehidratación post ejercicio cuando es necesario una rápida reposición del balance hídrico y cuando lo único que se ingiere es líquidos. Algunos estudios en esta área [Costill and Sparks, 1973; Nielsen, 1986] establecieron que luego de la ingesta de grandes volúmenes de bebidas libres de electrolitos se produce un gran flujo de orina, lo cual no permite mantener el balance hídrico por más de un período muy corto de tiempo. Estos investigadores también establecieron que el volumen plasmático era mejor mantenido cuando los electrolitos estaban presentes en el fluido ingerido, un efecto atribuido a la presencia de sodio en las bebidas. Los primeros estudios en investigar los mecanismos de rehidratación post ejercicio mostraron que la ingesta de grandes volúmenes de agua común luego del ejercicio, en sujetos deshidratados, resultaba en una rápida caída de la osmolalidad del plasma y de la concentración de sodio [Nose et al., 1988a], derivando en una abrupta y marcada diuresis causada por un rápido retorno a los niveles de control de la actividad de la renina y de la aldosterona [Nose et al., 1988b].
En una investigación sistemática de la relación entre las pérdidas de sodio corporales totales y la efectividad de las bebidas deportivas con diferentes concentraciones de sodio para restaurar el balance hídrico, Shirreffs y Maughan [1998] mostraron que, dado que se consume un adecuado volumen, la euhidratación se alcanza cuando la ingesta de sodio es mayor que la pérdida, aunque, como se discutiera previamente, no todos los estudios han reportado resultados similares. Se ha demostrado que la concentración de sodio en las bebidas deportivas es más importante que su contenido osmótico para incrementar el volumen plasmático luego de la deshidratación [Greenleaf, 1998]. El sodio además estimula la absorción de glucosa en el intestino delgado por medio del transporte activo de glucosa y sodio, lo cual crea un gradiente osmótico que actúa para promover la absorción neta de agua [Shirreffs, 2004].
Potasio
El potasio es el ion principal del fluido intracelular. Por lo tanto el potasio puede ser importante para alcanzar la rehidratación aumentando la retención de agua en el espacio intracelular [Shirreffs, 2004]. Yawata [1990], sugirió que en el espacio extracelular, la restauración de la concentración de sodio es más importante que la restauración del volumen, pero que la restauración del volumen tiene prioridad en el fluido intracelular. Además el papel del potasio en la restauración del volumen intracelular es más modesto que el papel del sodio en la restauración del volumen extracelular. Este tema fue investigado en hombres deshidratados en ~ 2% de su masa corporal quienes ingirieron una bebida a base de glucosa (90 mmol/L), una bebida que contenía sodio (NaCl 60 mmol/L), una bebida que contenía potasio (KCl 25 mmol/L) o una bebida que contenía los tres componentes [Maughan, 1994]. Todas las bebidas fueron consumidas en un volumen equivalente a la pérdida de masa corporal, sin embargo un menor volumen de orina fue excretado luego de la rehidratación cuando cada bebida que contenía electrolitos fue ingerida (aproximadamente 250-300 mL) en comparación con la bebida libre de electrolitos (un volumen promedio de 577 mL). Por lo tanto no hubo diferencias en la fracción ingerida de fluidos retenida 6 horas después de haber consumido las bebidas que contenían electrolitos. Esto es debido a que el volumen de bebida consumido fue equivalente al volumen de sudor perdido. Se observó una reducción estimada del 4.4% en el volumen plasmático con la deshidratación, pero la tasa de recuperación fue más lenta cuando se consumió la bebida con KCl. Por lo tanto, el potasio puede ser importante para mejorar la rehidratación aumentando la rehidratación intracelular, per se requieren más investigaciones para proveer evidencias concluyentes [Shirreffs, 2004].
Otros Electrolitos
La importancia de incluir magnesio en las bebidas deportivas ha sido sujeto de mucha discusión. El magnesio se pierde en el sudor y muchos creen que esto provoca la reducción de las concentraciones plasmáticas de magnesio, lo cual se ha asociado con los calambres musculares [Shirreffs, 2004]. Aun cuando puede haber una reducción en las concentraciones plasmáticas de magnesio durante el ejercicio, es más probable que se deba a la redistribución compartimental de fluidos más que la pérdida por el sudor. Por lo tanto, no existe una buena razón aparente para incluir magnesio en las bebidas deportivas para la rehidratación y la recuperación.
El sodio es el electrolito más importante en términos de recuperación luego del ejercicio. Sin su reemplazo, la retención de agua obstaculizada [Shirreffs, 2004]. Aunque existe evidencia contundente para la inclusión de sodio, este no es el caso para el potasio. No existe evidencia para la inclusión de cualquier otro electrolito [Shirreffs, 2004]
Carbohidratos
Los CHO se encuentran disponibles como monómeros, dímeros y oligómeros. Los dos monómeros más fácilmente disponibles, la glucosa y la fructosa, tienen la ventaja de ser sustratos directos para el transporte, no requieren digestión [Shi, 1995]. La concentración de CHO en la solución es uno de los factores que regulan la absorción de fluidos. Muchos estudios sobre soluciones de glucosa y electrolitos revelan que la concentración luminal de glucosa de 50 a 90 mmol, en ratas [Wapnir, 1985; Cunha Ferreira, 1987; Farthing, 1990; Hunt, 1991,] y de 80 a 200 mmol en seres humanos [Fordtran, 1975], eleva al máximo la tasa de absorción de líquidos. Una observación reciente de Gisolfi et al [1992] indica que la absorción de agua no fue reducida significativamente por el aumento de la concentración de CHO hasta un 8% (67 o 233 mmol), con soluciones que contenían maltodextrinas o sucrosa; pero una solución al 8% de glucosa (444 mmol) causó secreción de agua. En un estudio llevado a cabo por Shi et al. [1995], se observó que las soluciones al 8% de sucrosa no produjeron una absorción de agua significativamente menor que las soluciones al 6% pero si lo hicieron las que tenían un 8% de glucosa y maltodextrinas. En este mismo estudio la administración de una solución hipertónica (8% de glucosa) redujo el volumen plasmático, sugiriendo que un 8% de glucosa produciría una secreción de agua en el segmento mixto del intestino (duodeno proximal). En general las soluciones al 8% de CHO y electrolitos tienden a tener tasas de absorción más lentas de agua que las soluciones al 6%.
Además, una bebida ingerida durante un ejercicio prolongado debería brindar los CHO para reponer las pérdidas asociadas con el aumento del metabolismo [Shi, 1995]. La observación de que la ingesta de carbohidratos durante el ejercicio está asociada con el incremento del rendimiento físico, durante ejercicio prolongados, ha sido interpretada como indicativo de que los carbohidratos exógenos pueden servir como combustible suplementario en el momento en que las reservas de glucógeno muscular se vuelven limitadas [Coyle, 1986]. En un estudio llevado a cabo por Shi et al [1995] se observó que las bebidas que contenían sucrosa y glucosa mostraron una tendencia a producir la mayor absorción de agua y sal pero estimularon una tendencia a producir solo una modesta cantidad de absorción de CHO. En este mismo estudio las soluciones con fructosa y glucosa libres provocaron la mayor absorción de CHO y una absorción moderada de agua, pero la más baja de Na+ Los resultados de un estudio realizado por Murray et al [1987] indicaron que el consumo de agua (placebo) y de bebidas que contienen entre el 6-8% de CHO provocaron respuestas sensoriales y fisiológicas similares, pero solo la bebida al 6% de CHO mejoró el rendimiento físico. En la Tabla 11 se muestran los lineamientos generales de requerimientos de líquidos y CHO para actividades de diferentes duraciones [Gisolfi, 1992].

Tabla 3. Requerimientos de fluidos para actividades de diferentes
intensidades y duraciones. [Gisolfi, 1992].
Aminoácidos
El efecto de la incorporación de aminoácidos en las soluciones de reposición de fluidos es un tema controversial [Cunha Ferreira, 1987, Nalin, 1970]. Por ejemplo, agregar glicina a una solución, tanto no tuvo efectos beneficiosos [Cunha Ferreira, 1987], como redujo la tasa neta de absorción de Na+ y agua [Sandhu, 1989], o fue tan eficiente como la glucosa para promover la absorción de agua y sodio [Patra, 1989].
CONCLUSIONES
La utilización de suplementos es una práctica muy difundida en el deporte, incluso cuando la mayoría de los suplementos utilizados son probablemente inefectivos. Los atletas que consumen suplementos deberían hacerlo solo luego de realizar un cuidadoso análisis de los costos y beneficios. Aunque estos suplementos son en su mayoría benignos, esto no siempre es así. Por lo tanto los deportistas (y la población general) deben ser cautos a la hora del uso indiscriminado de los suplementos dietarios. La utilización de suplementos puede desempeñar un papel fundamental cuando la ingesta de alimentos se ve restringida, o como un remedio a corto plazo (i.e., cuando existe alguna deficiencia). La utilización de algunos o varios suplementos no sustituye una buena alimentación.
REFERENCIAS
1. American College of Sports Medicine and Dietitians of Canada (2000). Nutrition and Athletic Performance: Position Stand. Med. Sci. Sports Exerc. 32(12): 2130-2145
2. American College of Sports Medicine (1987). Position Stand on the use of anabolic-androgenic steroids in sport. Med. Sci. Sports Exerc. 19:534-539
3. Arciero PJ, NS Hannibal 3rd, BC Nindl, CL Gentile, J Hamed, and MD Vukovich (2001). Comparison of creatine ingestión and resistance training on energy expenditure and limb blood flow. Metabolism. 50:14291434
4. Barnett C, Costill DL, Vukovich MD, Cole KJ, Goodpaster BH, Trappe SW and Fink WJ (1994). Effect of L-carnitine supplementation on muscle and blood carnitine content and lactate accumulation during highintensity sprint cycling. Int. J. Sport Nutr. 4:280288
5. Becque B, Lochmann J, Melrose D (1997). Effect of creatine supplementation during strength training on 1RM and body composition. Med. Sci. Sports Exerc. 29:S146
6. Becque MD, JD Lochmann, and DR Melrose (2000). Effects of oral creatine supplementation on muscular strength and body composition. Med. Sci. Sports Exerc. 32:654658
7. Bemben MG, DA Bemben, DD Loftiss, and AW Knehans (2001). Creatine supplementation during resistance training in college football athletes. Med. Sci. Sports Exerc. 33:16671673
8. Bent S, Tiedt T, Odden M, Shlipak M (2003). The relative safety of ephedra compared with other herbal products. Ann Intern Med. Sci. Sports Exerc
9. Berning JR (1998). Energy Intake, diet and muscle wasting. In: Kreider RB, Fry AC, OToole ML eds., Overtraining in Sport. Human Kinetics Publishers, Champaign, IL pp 275-288
10. Bessman S, Savabi F (1988). The role of phosphocreatine energy shuttle in exercise and muscle hipertrophy. In: Taylor A, Gollnick P, Green H (eds). International Series on Sport Sciences: Biochemistry of Exercise VII. Champaign (IL): Human Kinetics 167-178
11. Birch R, Noble D, Greenhaff P (1994). The influence of dietary creatine supplementation on performance during repeated bouts of maximal isokinetic cycling in man. Eur. J. App. Physiol. 69:268-270
12. Bloomstrand E, Hassmen P, Ekblom B, et al (1991). Administration of branch-chain amino acids during sustained exercise effects on performance and on plasma concentration of some amino acids. Eur. J. App. Physiol. 63:83-88
13. Boozer C, Daly P, Homel P, et al (2002). Herbal ephedra/caffeine for weight loss: a 6-month randomized safety and efficacy trial. Int J Obesity. 26:593-604
14. Boozer C, Nasser J, SB H, Wang V, Chen G, Solomon J (2001). An herbal supplement containing Ma Huang-Guarana for weight loss: a randomized, double-blind trial. Int J Obes Relat Metab Disord 25:316-24
15. Boozer CN, Daly PA, Homel P, et al (2002). Herbal ephedra/caffeine for weight loss: a 6-month randomized safety and efficacy trial. nt J Obes Relat Metab Disord; 26(5):593-604
16. Boozer CN, Nasser JA, Heymsfield SB, Wang V, Chen G, Solomon JL (2001). An herbal supplement containing Ma Huang-Guarana for weight loss: a randomized, double-blind trial. Int J Obes Relat Metab Disord; 25(3):316-24
17. Brass EP (2000). Supplemental carnitine and exercise. Am J Clin Nutr 72(2 Suppl):618S-23S
18. Brenner M, J Walberg Rankin, and D Sebolt (2000). The effect of creatine supplementation during resistance training in women. J. Strength Cond. Res. 14:207213
19. Brilla LR, Conte V (2000). Effects of a novel zing-magnesium formulation on hormones and strength.. J Exerc Physiol Online 3(4)
20. Buchman AL, Awal M, Jenden D, Roch M, Kang SH (2000). The effect of lecithin supplementation on plasma choline concentrations during a maratón. J Am Coll Nutr 19(6):768-70
21. Buchman AL, Jenden D, Roch M (1999). Plasma free, phospholipid-bound and urinary free choline all decrease during a maratón run and may be associated with impaired performance. J Am Coll Nutr 18(6):598-601
22. Carli G, Bonifazi M, Lodi L, et al (1992). Changes in exercise-induced hormone response to branched chain amino acid administration . Eur. J. App. Physiol Ocupp. Physiol. 64:272-277
23. Casey A, Constantin-Teodosiu D, Howell D, et al (1996). Creatine ingestión favorably affects performance and muscle metabolism during maximal exercise in humans. Am. J. Physiol 271:E31-E37
24. Chandler RM, Byrne HK, Patterson JG, et al (1994). Dietary supplements affect the anabolic hormones after weight training exercise. J. App. Physiol. 76:839-845
25. Clarkson PM and Rawson ES (1999). Nutritional supplements to increase muscle mass. Critical Reviews in Food Science and Nutrition, 39, 317328
26. Coombes J, McNaughton L (1995). The effects of branched chain amino acid supplementation on indicators of muscle damage after prolonged strenuous exercise. Med. Sci. Sports Exerc. 27:S149
27. Crist DM, Peake GT, Egan PA, et al (1988). Body composition response to exogenous GH during training in highly conditioned adults. J. App. Physiol. 65: 579-584
28. Crist DM, Peake GT, Loftfield RB, et al (1991). Supplemental growth hormone alters body composition, muscle protein metabolism and serum lipids in fit adults: characterization of dose-dependent and response-recovery effects. Mech Ageing Dev. 58:191-205
29. Dangott B, E Schultz, and PE Mozdziak (2000). Dietary creatine monohydrate supplementation increases satellite cell mitotic activity during compensatory hypertrophy. Int. J. Sports Med. 21:1316
30. Davies KM, Heaney RP, Recker RR, et al (2000). Calcium intake and body weight. J Clin Endocrinol Metab 85(12):4635-8
31. Dawson B, Cutler M, Moody A, et al (1995). Effects of oral creatine loading on single and repeated maximal shor sprints. Aust. J. Sci. Med. Sport 27:56-61
32. Dulloo AG, Duret C, Rohrer D, et al (1999). Efficacy of a green tea extract rich in catechin polyphenols and caffeine in increasing 24-h energy expenditure and fat oxidation in humans. Am J Clin Nutr 70(6):1040-5
33. Dulloo AG, Seydoux J, Girardier L, Chantre P, Vandermander J (2000). Green tea and thermogenesis: interactions between catechinpolyphenols, caffeine and sympathetic activity. Int J Obes Relat Metab Disord 24(2):252-8
34. Earnest CP, PG Snell, R Rodriguez, AL Almada, And TL Mitchell (1995). The effect of creatine monohydrate ingestion onanaerobic power indices, muscular strength and body composition. Acta Physiol. Scand. 153:207209
35. Enette Larson-Meyer D, GR Hunter, CA Trowbridge, JC Turk, JM Ernest, SL Torman, and PA Harbin (2000). The effect of creatine supplementation on muscle strength and body composition during off-season training in female soccer players. J. Strength Cond. Res. 14:434442
36. Fawcett JP, Farquhar SJ, Walker RJ, Thou T, Lowe G and Goulding A (1996). The effect or oral vanadyl sulphate on body composition and performance in weighttraining athletes. Int J. Sport Nutr., 6, 382390
37. Ferrando AA and Green NR (1993). The effect of boron supplementation on lean body mass, plasma testosterona levels and strength in male body builders. Int. J. Sports Med.3:140149
38. Fleming GA (2000). The FDA, regulation, and the risk of stroke. N Engl J Med 343(25):1886-7
39. Food and Nutrition Board (1989). National Research Council: Recommended Dietary Allowances, revised. National Academy of Sciences. Washington DC
40. Forbes GB (1991). Exercise and body compositiion. J. App. Physiol. 70: 994-997
41. Forbes GB (1992). Sequence of changes in body composition induced by testosterone and reversal of changes after drug is stoped. JAMA, 267:397-399
42. Forbes GB (1985). The effect of anabolic steroids on lean body mass: the dose response curve. Metabolism 34:571-573
43. Gallagher PM, Carrithers JA, Godard MP, Schulze KE, Trappe SW (2000). Beta-hydroxy-betamethylbutyrate ingestion. Part I: Effects on strength and fat free mass. Med. Sci. Sports Exerc. 32:21092115
44. Gonzalez-Cadavid NF, Taylor WE, Yarasheski K, et al (1998). Organization of the human myostatin gene and expression in healthy men and HIV-infected men with muscle wasting. Proc Natl Acad Sci USA; 95(25):14938-43
45. Green NR, Ferrando AA (1994). Plasma boron and the effects of boron supplementation in males. Environ Health Perspect 102 Suppl 7:73-7
46. Greenway F, Herber D, Raum W, Morales S (1999). Double-blind, randomized, placebo-controlled clinical trials with nonprescription medications for the treatment of obesity. Obes Res 7(4):370-8
47. Greenway FL, Raum WJ, DeLany JP (2000). The effect of an herbal dietary supplement containing ephedrine and caffeine on oxygen consumption in humans. J Altern Complement Med 6(6):553-5
48. Greenway FL, Ryan DH, Bray GA, Rood JC, Tucker EW, Smith SR (1999). Pharmaceutical cost savings of treating obesity with weight loss medications. Obes Res 7(6):523-31
49. Greenway FL (2001). The safety and efficacy of pharmaceutical and herbal caffeine and ephedrine use as a weight loss agent . Obes Rev; 2(3):199-211
50. Grindstaff P, Kreider R, Bishop R, et al (1997). Effects of creatine supplementation on repetitive srpint performance and body composition in competitive swimers. Int. J. Sport. Nutr. 7:330-346
51. Grobet L, Martin LJ, Poncelet D, et al (1997). A deletion in the bovine myostatin gene causes the double-muscled phenotype in cattle. Nat Genet; 17(1):71-4
52. Harris R, Viru M, Greenhaff P, et al (1993). The effect of oral creatine supplementation on running performance during maximal short term exercise in man. J. Physiol. 467:74P
53. Haussinger D, E Roth, F Lang, and W Gerok (1993). Cellular hydration state: An important determinant of protein catabolism in health and disease. Lancet. 341:13301332
54. Haussinger D, ER Roth, F Lang, and W Gerok (1993). Cellular Hidration state: an important determinant of protein catabolism in health and disease. Lancet 341: 1330-1332
55. Hawley J, Burke L (1998). Peak Performance: Training and Nutritional Strategies for Sports. St Leonards, NWS: Allen & Unwin Publishers
56. Inwall J (1940). Creatine and the control of muscle-specific protein synthesis in cardiac skeletal muscle. Circ. Res. 38: 115-1123
57. Ivey FM, Roth SM, Ferrell RE, et al (2000). Effects of age, gender, and myostatin genotype on the hypertrophic response to heavy resistance strength training. J Gerontol A Biol Sci Med Sci. 55 (11):M641-8
58. Jowko E, P Ostaszewski, M Jank, J Sacharuk, A Zieniewicz, J Wilczak, and S Nissen (2001). Creatine and beta-hydroxybeta- methylbutyrate (HMB) additively increase lean body mass and muscle strength during a weight-training program. Nutrition. 17:558566
59. Kreider RB (1995). Effects of creatine loading on muscular strength and body composition. Nat. Strength Cond. Assoc. J. Oct: 72-73
60. Kaciuba-Uscilko H, Nazar K, Chwalbinska-Moneta J, et al (1993). Effect of phosphate supplementation on metabolic and neuroendocrine responses to exercise and oral glucose load in obese women during weight reduction. J Physiol Pharmacol 44(4):425-40
61. Kalman D, Colker CM, Wilets I, Roufs JB, Antonio J (1999). The effects of pyruvate supplementation on body composition in overweight individuals. Nutrition 15(5):337-40
62. Kambadur R, Sharma M, Smith TP, Bass JJ (1997). Mutations in myostatin (GDF8) in double-muscled Belgian Blue and Piedmontese cattle. Genome Res; 7(9):910-6
63. Kargotich S, Rowbottom DG, Keast D, et al (1996). Plasma glutamine changes after high intensity exercise in elite male swimmers. Med. Sci. Sports Exerc 28:S133
64. Kelly VG, and DG Jenkins (1998). Effect of oral creatine supplementation on near-maximal strength and repeated sets of highintensity bench press exercise. J. Strength Cond. Res. 12:109115
65. Kirksey K, Warren B, Stone M, et al (1997). The effects of six weeks of creatine monohidrate supplementation in male and female track athletes. Med. Sci. Sports Exerc.29:S145
66. Kraemer WJ (1994). General adaptations to resistance training and endurance training. In: Beachle T, ed. Essentials of strength training and conditioning. Champaing (IL): Human Kinetics, 127-150
67. Kreider RB, Almada AL, Antonio J, Broeder C, Earnest C, Greenwood M, Incledon T, Kalman DS, Kleiner SM, Leutholtz B, Lowery LM, Mendel R, Scout JR, Willoughby DS, Ziegenfuss TN (2004). Issn Exercise & Sport Nutrition Review: Research & Recommendations. Sports Nutrition Review Journal. 1 (1):1-44
68. Kreider RB, Ferreira M, Wilson M, Almada AL (1999). Effects of calcium beta-hydroxy-beta-methylbutyrate (HMB) supplementation during resistance-training on markers of catabolism, body composition and strength. Int. J. Sports Med., 20: 503509
69. Kreider RB, Ferreira M, Wilson M, et al (1997). Effects of calcium β-HMB supplementation with or without creatine during training on body composition alterations. FASEB J. 11:A374
70. Kreider RB, Ferreira M, Wilson M, et al (1997). Effects of creatine supplementation with and without glucosa on body composition in trained and untrained men and women. J. Strength Cond. Res. 11:283
71. Kreider RB, Klesges R, Harmon K, et al (1996). Effects of ingesting supplements designed to promote lean tissue accretion on body composition during resistance exercise. Int. J. Sport Nutr. 6:234-246
72. Kreider RB, M Ferreira, M Wilson, P Grindstaff, S Plisk, J Reinardy, E Cantler, and AL Almada (1998). Effects of creatine supplementation on body composition, strength, and sprint performance. Med. Sci. Sports Exerc. 30:7382
73. Kreider RB, Miller GW, Mitchell M, et al (1992). Effects of amino acid supplementation on ultraendurance triathlon performance. Proceedings of the I World Congress on Sport Nutrition; Jun 16-18: Barcelona. Barcelona: Enero, 1992: 488-536
74. Kreider RB, Miller GW, Schenck D, et al (1992). Effects of phosphate loading on metabolic and myocardial responses to maximal and endurance exercise. Int J Sport Nutr 2(1):20-47
75. Kreider RB, Miller GW, Williams MH, Somma CT, Nasser TA (1990). Effects of phosphate loading on oxygen uptake, ventilatory anaerobic threshold, and run performance. Med Sci Sports Exerc 22(2):250-6
76. Kreider RB, Miriel V, Bertun E et al (1993). Effects of amino acid and carnitine supplementation on markers of protein catabolism and body composition during 25 weeks of swim training. Southeast American College of Sports Medicine Conference Abstracts; 1993 Jan 29. American College of Sports Medicine, 20:45
77. Kreider RB, Miriel V, Bertun E (1993). Amino acid supplementation and exercise performance: proposed ergogenic value. Sports Med 16:190-209
78. Kreider RB (1998). Central fatigue hypothesis and overtraining. In: Kreider RB, Fry AC, OToole ML (eds). Overtraining in Sport. Champaign (IL): Human Kinetics, 3099-331
79. Kreider RB (1999). Dietary supplements and the promotion of muscle growth with resistance training . Sports Med 27(2): 97-110
80. Kreinder RB (1998). Creatine supplementation análisis of ergogenic value, medical safety, and concerns. J. Exerc. Physiol. (serial online). 1:1-12
81. Kuipers HJ, Winnen AG, Hartgens F, et al (1991). Influence of anabolic steroids on body composition, blood pressure, lipid profile and liver functions in body builders. Int. J. Sport Med. 12:413-418
82. Lemon PW, Tarnopolsky MA, McDougall JD et al (1992). Protein requirements and muscle mass/strength changes during intensive training in novice body builders. J. App. Physiol 73: 767-775
83. Leutholtz B, Kreider RB (2001). Exercise and Sport Nutrition. In: Wilson T, Temple N, eds. Nutritional Health. Totowa, NJ:Humana Press, Inc.;207 39
84. Leutholtz B, Kreider RB (2001). Optimizing Nutrition for Exercise and Sport. Nutritional Health: Strategies for Disease Prevention. Wilson T and Temple NJ Eds., Humana Press Inc., Totowa, NJ
85. Low SY, Taylor PM, Rennie MJ (1996). Responses of glutamine transport in cultured rat skeletal muscle to osmotically induced changes in cell volume. J. Physiol (Lond) 492(3):877-885
86. Manore Mm, Thompson Jl (2000). Sport Nutrition for Health and Performance. Champaign, Ill: Human Kinetics
87. McPherron AC, Lawler AM, Lee SJ (1997). Regulation of skeletal muscle mass in mice by a new TGF-beta superfamily member. Nature; 387(6628):83-90
88. McPherron AC, Lee SJ (1997). Double muscling in cattle due to mutations in the myostatin gene. Proc Natl Acad Sci USA; 94(23):12457-61
89. Millward DJ (2001). Protein and amino acid requirements of adults: current controversias. Canadian Journal of Applied Physiology, 26, S130S140
90. Molnar D, Torok K, Erhardt E, Jeges S (2000). Safety and efficacy of treatment with an ephedrine/caffeine mixture. The first doubleblind placebo-controlled pilot study in adolescents. Int J Obes Relat Metab Disord 24(12):1573-8
91. Molnar D (1993). Effects of ephedrine and aminophylline on resting energy expenditure in obese adolescents. Int J Obes Relat Metab Disord 17 Suppl 1:S49-52
92. Montoye Hj, Kemper Hcg, Saris Whm, Washburn Ra (1996). Measuring Physical Activity and Energy Expenditure. Champaign, IL: Human Kinetics Publication
93. Mourier A, Bigard AX, de Kerviler E (1997). Combined effects of caloric restriction and branched chain amino acid supplementation on body composition and exercise performance in elite wrestlers. Int. J. Sports Med. 18:47-55
94. Nair KS, Schwartz RG, Welle S (1992). Leucine as a regulator of whole body and skeletal muscle protein metabolism in humans. Am. J. Physiol. 263 (5 Pt I): E928-E934
95. Nakagawa K, Ninomiya M, Okubo T, et al (1999). Tea catechin supplementation increases antioxidant capacity and prevents phospholipid hydroperoxidation in plasma of humans. J Agric Food Chem 47(10):3967-73
96. National Research Council (1989). Recommended Dietary Allowances.10th ed. Washington, DC: National Academy Press
97. Nazar K, Kaciuba-Uscilko H, Szczepanik J, et al (1996). Phosphate supplementation prevents a decrease of triiodothyronine and increases resting metabolic rate during low energy diet. J Physiol Pharmacol 47(2):373-83
98. Nielson FH, Hunt CD, Mullen LM and Hunt JR (1987). Effect of dietary boron on mineral, estrogen and testosterone metabolism in post-menopausal women. Federation of the American Societies for Experimental Biology Journal, 1, 394397
99. Nissen S, Faidley TD, Zimmerman DR, et al (1994). Colostral milk fat and pig performance are enhanced bay feeding the leucine metabolite β-hidroxi-β-methylbutyrate to sows . J. Anim. Sci. 72:2331-2337
100. Nissen S, Panton L, Fuller J, et al (1997). Effect of feeding β-hidroxy-β-methylbutyrate (HMB) on body composition and strength of women. FASEB J. 11:A150
101. Nissen S, Sharp R, Ray M, et al (1996). Effect of leucine metabolite beta-hidroxi-beta-metilbutirato on muscle metabolism during resistance-exercise training. J. App. Physiol 81:2095-2104
102. Nissen SL and Sharp RL (2003). Effect of dietary supplements on lean mass and strength gains with resistance exercise: a meta-analysis. App. Physiol, 94:651659
103. Noonan D, K Berg RW Latin, JC Wagner, and K Reimers (1998). Effects of varying dosages of oral creatine relative to fat free body mass on strength and body composition. J. Strength Cond. Res. 12:104108
104. Paddon-Jones D, Keech A and Jenkins D (2001). Shortterm beta-hydroxy-beta-methylbutyrate supplementation does not reduce symptoms of eccentric muscle damage. Int. J. Sport Nutrit. Exerc. Metab. 11:442450
105. Panton LB, Rathmacher JA, Baier S, Nissen S (2000). Nutritional supplementation of the leucine metabolite beta-hydroxy-beta-methylbutyrate (hmb) during resistance training. Nutrition 16:734739
106. Parise G, S Mihic, D Maclennan, KE Yarasheski, and MA Tarnopolsky (2001). Effects of acute creatine monohydrate supplementation on leucine kinetics and mixed-muscle protein synthesis. J. Appl. Physiol. 91:10411047
107. Parry-Billings M, Budgett R, Koutedakis K, et al (1992). Plasma amino acid concentrations in the overtraining syndrome: possible effects on the immune system. Med. Sci. Sports Exerc. 24:1353-1358
108. Pearson DR, DG Hamby, W Russel, and T Harris (1999). Longterm effects of creatine monohydrate on strength and power. J. Strength Cond. Res. 13:187192
109. Peeters BM, CD Lantz, and JL Mayhew (1999). Effect of oral creatine monohydrate and creatine phosphate supplementation on maximal strength indices, body composition, and blood pressure. J. Strength Cond. Res. 13:39
110. Prevost M, Nelson A, Morris G (1997). The effects of creatine supplementation on total work output and metabolism during high intensity intermittent exercise. Res. Q. Exerc. Sport 68:233-240
111. Rama Rao SV, Sunder GS, Reddy MR, Praharaj NK, Raju MV, Panda AK (2001). Effect of supplementary choline on the performance of broiler breeders fed on different energy sources. Br Poult Sci 42(3):362-7
112. Ransone J, Neighbors K, Lefavi R, Chromiak J (2003). The effect of beta-hydroxy beta-methylbutyrate on muscular strength and body composition in collegiate football players. J. Strength Cond. Res. 17:3439
113. Rawson ES, Volek JS (2003). Effects of Creatine Supplementation and Resistance Training on Muscle Strength and Weightlifting Performance. J. Strength Cond. Res. 17(4), 822831
114. Rennie MJ, Tadros L, Khogali S, et al (1994). Glutamine transport and its metabolic effects. J. Nutr. 124(8):1503S-1508S
115. Rennie MJ (1996). Glutamine metabolism and transport in skeletal muscle and heart and their clinical relevance. J. Nutr. 126(4): 1142S-1149S
116. Slater G, Jenkins D, Logan P, Lee H, Vukovich M, Rathmacher JA, Hahn AG (2001). Beta-hydroxybeta-methylbutyrate (HMB) supplementation does not affect changes in strength or body composition during resistance training in trained men. Int. Sport Nutrit. Exerc. Metab. 11:384396
117. Spriet LL (1997). Ergogenic aids: recent advances and retretas. In Optimizing Sports Performance (edited by D.R. Lamb and D. Murray), pp. 185238. Carmel, IN: Cooper Publishing.
118. Stanko RT, Arch JE (1996). Inhibition of regain in body weight and fat with addition of 3-carbon compounds to the diet with hyperenergetic refeeding after weight reduction. Int J Obes Relat Metab Disord 20(10):925-30
119. Stanko RT, Reynolds HR, Hoyson R, Janosky JE, Wolf R (1994). Pyruvate supplementation of a low-cholesterol, low-fat diet: effects on plasma lipid concentrations and body composition in hyperlipidemic patients. Am J Clin Nutr 59(2):423-7
120. Stanko RT, Tietze DL, Arch JE (1992). Body composition, energy utilization, and nitrogen metabolism with a severely restricted diet supplemented with dihydroxyacetone and pyruvate. Am J Clin Nutr 55(4):771-6
121. Stevenson SW, and GA Dudley (2001). Creatine loading, resistance exercise performance, and muscle mechanics. J. Strength Cond. Res. 15:413419
122. Stone MH, Sanborn K, Smith LL, et al (1999). Effects of in-season (5 weeks) creatine and pyruvate supplementation on anaerobic performance and body composition in American football players. Int J Sport Nutr 9(2):146-65
123. Stout J, Eckerson J, Noonan D, et al (1997). The effects of a supplement designed to augment creatine uptake on exercise performance and fat-free mass in football players. Med. Sci. Sports Exerc. 29:S251
124. Stout J, J Eckerson, D Noonan, and G Moore (1999). Effects of 8 weeks of creatine supplementation on exercise performance and fat-free weight in football players during training. Nutr. Res. 19:217225
125. Swinburn B, Ravussin E (1993). Energy balance or fat balance?. Am. J.Clin. Nutr. 57(suppl):766S-771S
126. Syrotuik DG, AB Game, EM Gillies, and GJ Bell (2001). Effects of creatine monohydrate supplementation during combined strength and high intensity rowing training on performance. Can. J. Appl. Physiol. 26:527542
127. Syrotuik DG, GJ Bell, R Burnham, LL Sim, RA Calvert, and IM Maclean (2000). Absolute and relative strength performance following creatine monohydrate supplementation combined with periodized resistance training. J. Strength Cond. Res. 14:182190
128. Tarnopolsky MA; Atkinson SA, McDougall JD, et al (1992). Evaluation of protein requirements for trained strenth athletes. J. App. Physiol, 73: 1986-1995
129. Tarnopolsky, M (2001). Protein and amino acid needs for training and bulking up. In Clinical Sports Nutrition, 2nd edn (edited by L. Burke and V. Deakin), Roseville, Australia: McGraw Hill. pp. 90123
130. Tipton KD, Ferrando AA, Phillips SM, Doyle D, Wolfe RR (1999). Postexercise net protein synthesis in human muscle from orally administered amino acids. Am. J. Physiol. Endocrinol. Metab. 276:E628-E634
131. Tipton KD, Wolfe RR (2004). Protein and amino acids for athletes. Journal of Sports Scienecs, 22, 6579
132. Van Koevering MT; Dolezal HG, Gill DR, et al (1994). Effects of ß-hidroxy-ß-methylbutyrate on performance and carcass quality of feedlot steers. J. Anim. Sci. 72:1927-1935
133. Vandenberghe K, M Goris, P Van Hecke, M Van Leemputte, L Vangerven, and P Hespel (1997). Long-term creatine intake is beneficial to muscle performance during resistance training. J. Appl. Physiol. 83:20552063
134. Vandenburghe K, Goris M, Van Hecke P, et al (1997). Long-term creatine intake is beneficial to muscle performance during resistance training. J. App. Physiol. 83:2055-2063
135. Varnier M, Lesse GP, Thomson J, et al (1995). Stimulatory effect of glutamine on glycogen accumulation in human skeletal muscle. Am. J. Physiol. 269(2 Pt 1):E309-E315
136. Villani RG, Gannon J, Self M, Rich PA (2000). L-Carnitine supplementation combined with aerobic training does not promote weight loss in moderately obese women. Int J Sport Nutr Exerc Metab; 10(2):199-207
137. Volek JS, ND Duncan, SA Mazzetti, RS Staron, M Utukian, AL Gomez, DR Pearson, WJ Fink, and WJ Kraemer (1999). Performance and muscle fiber adaptations to creatine upplementation and heavy resistance training. Med. Sci. Sports Exerc. 31:11471156
138. Volek JS, WJ Kraemer, JA Bush, M Boetes, T Incledon, KL Clark, JM Lynch (1997). Creatine supplementation enhances muscular performance during high-intensity resistance exercise. J. Am. Diet Assoc. 97:765770
139. Vukovich MD, Costill DL and Fink WJ (1994). Carnitine supplementation: effect on muscle carnitine and glycogen content during exercise. Med. Sci. Sports Exerc. 26:11221129
140. Vukovich MD, Stubbs NB, Bohlken RM, et al (1997). Effec of dietary ß-hydroxy-ß-methylbutyrate (HMB) on strength gains and body composition changes in older adults. FASEB J. 11:A376
141. Vukovich, M (2001). Fat reduction. n Sports Supplements (edited by J. Antonio and J.R. Stout), pp. 84110. Philadelphia, PA: Lippincott Williams & Wilkins
142. Williams MH (1999). Nutritión for Health, Fitness and Sport. WCB/McGraw-Hill, Dubuque, IA
143. Williams MH (1999). Nutrition for Health, Fitness, and Sport. Dubuque, IA: ACB/McGraw-Hill
144. Willoughby DS, and J Rosene (2001). Effects of oral creatine and resistance training on myosin heavy chain expression. Med. Sci. Sports Exerc. 33:16741681
145. Wilmore JH (1974). Alterations in strength, body composition and anthropometric measurements consequent to al 10 week weight training program. Med. Sci. Sports Exerc 6: 133-138
146. Zemel M, Thompson W, Zemel P, et al (2002). Dietary calcium and dairy products accelerate weight and fat-loss during energy restriction in obese adults. Clin Nutri 75
147. Zemel MB, Shi H, Greer B, Dirienzo D, Zemel PC (2000). Regulation of adiposity by dietary calcium. FASEB J 14(9):1132-8
148. Zemel MB (2003). Mechanisms of dairy modulation of adiposity. J Nutr 133(1):252S-6S
149. Zemel MB (2003). Role of dietary calcium and dairy products in modulating adiposity. Lipids 38(2):139-46
150. Ziegenfuss T, Lemon P, Rogers M, et al (1997). Acute creatine ingestión: Effects on muscle volume, anaerobic power, fluid volumen, and protein turnover. Med. Sci. Sports. Exerc. 29:S127
151. Ziegenfuss TN, LM Lowery, and PW Lemon (1998). Acute fluid volume changes in men during three days of creatine supplementation. J. Exerc. Physiol. (serial online) 1:1-9
152. Maughan, R.J (1999). Nutritional ergogenic aids and exercise performance. Nutrition Research Reviews, 12, 255280
153. McNaughton, L (2000). Bicarbonate and citrate. In Nutrition in Sport (edited by R.J. Maughan). pp. 393404. Oxford: Blackwell
154. Wilkes, D., Gledhill, N. and Smyth, R (1983). Effect of acute induced metabolic alkalosis on 800-m racing time. Medicine and Science in Sports and Exercise, 15, 277280
155. Bird, S.R., Wiles, J. and Robbins, J (1995). The effect of sodium bicarbonate ingestion on 1500-m racing time. Journal of Sports Sciences, 13, 399403
156. McNaughton L, Backx K, Palmer G, Strange N (1999). Effects of chronic bicarbonate ingestion on the performance of high-intensity work. Eur J Appl Physiol Occup Physiol 80(4):333-6
157. Applegate E (1999). Effective nutritional ergogenic aids. Int J Sport Nutr 9(2):229-39
158. Kronfeld DS, Ferrante PL, Grandjean D (1994). Optimal nutrition for athletic performance, with emphasis on fat adaptation in dogs and horses. J Nutr 124(12 Suppl):2745S-53S
159. Kraemer WJ, Gordon SE, Lynch JM, Pop ME, Clark KL (1995). Effects of multibuffer supplementation on acid-base balance and 2,3- diphosphoglycerate following repetitive anaerobic exercise. Int J Sport Nutr 5(4):300-14
160. Matson LG, Tran ZV (1993). Effects of sodium bicarbonate ingestion on anaerobic performance: a meta-analytic review. Int J Sport Nutr;3(1):2-28
161. Horswill, C.A., Costill, D.L., Fink, W.J., Flynn, M.G., Kirwan, J.P., Mitchell, J.B. and Houmard, J.A (1988). Infuence of sodium bicarbonate on sprint performance: relationship to dosage. Medicine and Science in Sports and Exercise, 20, 566569
162. McKenzie, D.C., Coutts, K.D., Stirling, D.R., Hoeben, H.H. and Kuzara, G (1986). Maximal work production following two levels of arti.cially induced metabolic alkalosis. Journal of Sports Sciences, 4, 3538
163. McNaughton LR (1990). Sodium citrate and anaerobic performance: implications of dosage. Eur J Appl Physiol Occup Physiol 61(5-6):392-7
164. Tiryaki GR, Atterbom HA (1995). The effects of sodium bicarbonate and sodium citrate on 600 m running time of trained females. J Sports Med Phys Fitness 35(3):194-8
165. Parry-Billings M, MacLaren DP (1986). The effect of sodium bicarbonate and sodium citrate ingestion on anaerobic power during intermittent exercise. Eur J Appl Physiol Occup Physiol 55(5):524-9
166. Kowalchuk JM, Maltais SA, Yamaji K, Hughson RL (1989). The effect of citrate loading on exercise performance, acid-base balance and metabolism. Eur J Appl Physiol Occup Physiol 58(8):858-64
167. Greenhaff, P.L (2000). Creatine. In Nutrition in Sport (edited by R.J. Maughan), pp. 379392. Oxford: Blackwell
168. Branch, J.D (2003). Effects of creatine supplementation on body composition and performance: a meta-analysis. Int. J. Sports Nutr. Exerc. Metab., 13, 198226
169. Misic M and Kelley GE (2002). The impact of creatine supplementation on anaerobic performance: a metaanalysis. Am. J. Med. Sports, 4, 116124
170. Balsom, P.D., S.D.R. Harridge, K. Soderlund, B. Sjodin, and B. Ekblom (1993). Creatine supplementation per se does not enhance endurance exercise performance. Acta. Physiol. Scand. 149:521523
171. Engelhardt, M., G. Neumann, A. Berbalk, and I. Reuter (1998). Creatine supplementation in endurance sports. Med. Sci. Sports Exerc. 30:11231129
172. Tullson PC, Terjung RL (1991). Adenine nucleotide synthesis in exercising and endurance-trained skeletal muscle. Am J Physiol 261(2 Pt 1):C342-7
173. Gross M, Kormann B, Zollner N (1991). Ribose administration during exercise: effects on substrates and products of energy metabolism in healthy subjects and a patient with myoadenylate deaminase deficiency. Klin Wochenschr; 69(4):151-5
174. Wagner DR, Gresser U, Kamilli I, Gross M, Zollner N (1991). Effects of oral ribose on muscle metabolism during bicycle ergometer in patients with AMP-deaminase-deficiency. Adv Exp Med Biol 383-5
175. Pliml W, von Arnim T, Stablein A, Hofmann H, Zimmer HG, Erdmann E (1992). Effects of ribose on exercise-induced ischaemia in stable coronary artery disease. Lancet 340(8818):507-10
176. Pauly DF, Pepine CJ (2000). -Ribose as a supplement for cardiac energy metabolism. J Cardiovasc Pharmacol Ther 5(4):249-58
177. Op 't Eijnde B, Van Leemputte M, Brouns F, et al (2001). No effects of oral ribose supplementation on repeated maximal exercise. 91(5):2275-81
178. Berardi JM, Ziegenfuss TN (2003). Effects of ribose supplementation on repeated sprint performance in men. J Strength Cond Res 17(1):47-52
179. Kreider RB, Melton C, Greenwood M, et al (2003). Effects of oral d-ribose supplementation on anaerobic capacity and selected metabolic markers in healthy males. Int J Sport Nutr Exerc Metab 13(1):87-96
180. Rivers WHR, Webber HN (1907). The action of caffeine on the capacity for muscular work. J Physiol; 36: 33-47
181. Costill DL, Dalsky GP, Fink WJ (1978). Effects of caffeine ingestión on metabolism and exercise performance. Med Sci Sports; 10 (3): 155-8
182. Ivy JL, Costill DL, Fink WJ, et al (1979). Influence of caffeine and carbohydrate feedings on endurance performance. Med Sci Sports; 11 (1): 6-11
183. Essig D, Costill DL, VanHandel PJ (1980). Effects of caffeine inges- tion on utilization of muscle glycogen and lipid during leg ergometer cycling. Int J Sports Med; 1 (1): 86-90
184. Spriet LL, MacLean DA, Dyck DJ, et al (1992). Caffeine ingestion and muscle metabolism during prolonged exercise in humans. Am J Physiol; 262 (6 Pt 1): E891-8
185. Graham TE, Spriet LL (1991). Performance and metabolic responses to a high caffeine dose during prolonged exercise. J Appl Physiol; 71 (6): 2292-8
186. Graham TE (2001). Caffeine and exercise: metabolism, endurance and performance. Sports Med; 31 (11): 785-807
187. Nehlig A, Debry G. (1994). Caffeine and sports activity: a review. Int J Sports Med; 15 (5): 215-2
188. Spriet LL (1995). Caffeine and performance. Int J Sport Nutr; 5 Suppl.: S84-99
189. Graham TE (2001). Caffeine, coffee and ephedrine: impact on exercise performance and metabolism. Can J Appl Physiol; 26 Suppl.: S103-19
190. Conlee RK (1991). Amphetamine, caffeine, and cocaine. In: Lamb DR, Williams MH, editors. Ergogenics: enhancement of performance in exercise and sport. Dubuque (IA): Wm. C. Brown Publishers, 285-330
191. Spriet LL, Howlett RA (2000). Caffeine. In: Maughan RJ, editor. Nutrition in sport. Oxford: Blackwell Science Ltd: 379-92
192. Wilcox AR (1990). Caffeine and endurance performance. Sports Sci Exch; 3 (1): 1-5
193. Clarkson PM (1993). Nutritional ergogenic aids: caffeine. Int J Sport Nutr; 3 (1): 103-11
194. Graham TE, Rush JW, van Soeren MH (1994). Caffeine and exercise: metabolism and performance. Can J Appl Physiol; 19 (2): 111-38
195. Graham TE, Spriet LL (1996). Caffeine and exercise performance. Sports Sci Exch 9 (1): 1-5
196. Spriet LL (1997). Ergogenic aids: recent advances and retreats. In: Lamb DR, Murray R, editors. Optimizing sport performance. Carmel (IN): Cooper Publishing Group, 185-238
197. Spriet LL, Graham TE (1999). Caffeine and exercise performance. Curr Comm ACSM; Jul: 1-3
198. Williams JH (1991). Caffeine, neuromuscular function and high-intensity exercise performance. J Sports Med Phys Fitness; 31 (3): 481-9
199. Tarnopolsky MA (1994). Caffeine and endurance performance. Sports Med; 18 (2): 109-25
200. Sinclair CJ, Geiger JD (2000). Caffeine use in sports: a pharmacological review. J Sports Med Phys Fitness; 40 (1): 71-9
201. Spriet LL (2002). Caffeine. In: Bahrke MS, Yesalis CE, editors. Performance-enhancing substances in sport and exercise. Champaign (IL): Human Kinetics, 267-78
202. Paluska SA (2003). Caffeine and exercise. Curr Sports Med Rep; 2 (4): 213-9
203. Magkos F, Kavouras SA (2004). Caffeine. In: Wolinsky I, Driskell JA, editors. Nutritional ergogenic aids. Boca Raton (FL): CRC Press, 275-323
204. Ahrendt DM (2001). Ergogenic Aids: Counseling The Athlete. Am. Fam. Physician 63:5
205. American College of Sports Medicine, American Dietetic Association, Dietitians of Canda (2000). Joint Position Statment. Med. Sci. Sports Exerc. Vol. 32, No. 12, pp. 21302145; J. Am. Diet. Assoc. Vol. 12, pp. 1543 1556; Diet of Canada Vol. 61, pp. 176-192
206. American College Of Sports Medicine (1996). Position Stand: Exercise and fluid replacement. Med. Sci. Sports Exerc. 28:i-vii
207. Bangsbo J (1994). The physiology of soccer – with special reference to intense intermittent exercise. Acta Physiologica Scandinavica 151, suppl. 619
208. Barr SI, Costill DL, Fink WJ (1991). Fluid replacement during prolonged exercise: Effects of water, saline, or no fluid. Med. Sci. Sports Exerc. 23:811 817
209. Below PR, Mora-Rodriguez R, Gonzalez-Alonso J, Coyle EF (1995). Fluid and carbohydrate ingestion independently improve performance during 1 h of intense exercise. Med. Sci. Sports Exerc. 27:200 210
210. Bergstrom J, Hultman E (1967). A study of the glycogen metabolis exercise in man. Scand J Clin Lab Invest 19: 218-228
211. Burke LM (1997). Fluid balance during team sports. J Sports Sci. 15:287295
212. Casa DJ, Armstrong LE, Hillman SK, Montain SJ, Reiff RV, Rich BSE, Roberts WO, Stone JA (2000). National Athletic Trainers Association Position Statement. Fluid replacement for athletes. J. Athletic Training. 35(2):212224
213. Coggan, A. and Coyle, E (1989). Metabolism and performance following carbohydrate ingestion late in exercise. Med. Sci. Sports Exerc. 21, 59-65
214. Coyle EF, Coggan AR, Hemmert MK, Ivy JL (1986). Muscle glycogen utilization during prolonged strenuous exercise when fed carbohydrate. J. App. Physiol. 61, 165-172
215. Ekblom B (1986). Applied physiology of soccer. Sports Medicine, 3:50-60
216. Fortney SM, Wenger CB, Bove JR, Nadel ER (1984). Effect of hyperosmolality on control of blood flow and sweating. J. App. Physiol. 57, 16881695
217. Hitchins S, Martin DT, Burke L, Yates K, Fallon K, Hahn A, Dobson GP (1999). Glycerol hyperhydra- tion improves cycle time trial performance in hot humid conditions . Eur. J. App. Physiol. Occup. Physiol. 80:494501
218. Kirkendall, D.T., Foster, C., Dean, J.A., Grogan, J. and Thompson, N.N (1998). Effects of glucose polymer supplementation on performance of soccer players. In: Science and Football. Eds: Reilly, T., Less, A., Davies, K., Murphy, W.J. London: E & FN Spon. 33-41
219. Latzka WA and Sawka MN (2000). Hyperhydration and glycerol: thermoregulatory effects during exercise in hot climates. Can. J. App. Physiol. 25:536 545
220. Latzka WA, Sawka MN, Montain SJ, Skrinar GS, Fielding RA, Matott RP, Pandolf B (1997). Hyperhydration: thermoregulatory effects during compen- sable exercise-heat stress. J. App. Physiol. 83:860866
221. Latzka WA, Sawka MN, Montain SJ, Skrinar GS, Fielding RA, Matott RP, Pandolf KB (1998). Hyperhydration: tolerance and cardiovascular effects during uncompensable exercise-heat stress. J. App. Physiol. 84:18581864
222. Leatt P (1986). The effect of glucose polymer ingestion on skeletal muscle glycogen depletion during soccer match play and its resynthesis following match". MSc Thesis, University of Toronto
223. Leiper J, Pitsiladis Y, Maughan R (2001). Comparison of water turnover rates in men undertaking prolonged cycling exercise and sedentary men. Int J Sports Med. 22:181185
224. Leiper JB, Carnie A, Maughan RJ (1996). Water turnover rates in sedentary and exercising middle aged men. Br J Sports Med. 30:2426
225. Lyons TP, Riedesel ML, Meuli LE, Chick TW (1990). Effects of glycerol-induced hyperhydration prior to exercise in the heat on sweating and core temperature. Med. Sci. Sports Exerc. 22, 477483
226. Maughan RJ, Leiper JB (1994). Requerimientos para la sustitución o reemplazo de liquidos en el fútbol. J. Sports Sci. 12:S29-S34
227. Maughan RJ, Merson SJ, Broad NP, Shirreffs SM (2004). Fluid and electrolyte intake and loss in elite soccer players during training. Int J Sport Nutr Exerc Metab. 14:333346
228. Mcconnell GK, Burge CM, Skinner SL, Hargreaves M (1997). Influence of ingested fluid volume on physiological responses during prolonged exercise. Acta Physiol. Scan. 160:149 156
229. Montain, SJ., and EF. Coyle (1992). Influence of graded dehydration on hypertermia and cardiovascular drift during exercise. J. Appl. Physiol. 73:1340-1350
230. Montner P, Stark DM, Riedesel ML, Murata G, Robergs R, Timms M, Chick TW (1996). Pre- exercise glycerol hydration improves cycling endurance time. International Journal of Sports Medicine 17, 2733
231. Nielsen M (1938). Die Regulation der Korpertemperatur bei Muskelarbeit. Skand Arch Physiol. 79:193230
232. Rehrer NJ, Burke LM (1996). Sweat losses during various sports. Aust J Nutr Diet 53:S13S16
233. Riedesel ML, Allen DY, Peake GT, Al-Qattan K (1987). Hyperhydration with glycerol solutions. J. App. Physiol. 63:22622268
234. Sawka MN, Cheuvront SN, Carter III R (2005). Human Water Needs. Nutrition Reviews 63(6): (II)S30 S39
235. Sawka MN, Pandolf KB (1990). Effects of body water loss on physiological function and exercise performance. In: Gisolfi CV, Lamb DR, eds. Perspectives in exercise science and sports medicine. Vol 3. Carmel, IN: Benchmark Press, 138
236. Sawka MN, Wenger CB, Pandolf KB (1996). Thermoregulatory responses to acute exercise-heat stress and heat acclimation. In: Fregly MJ, Blatteis CM, eds. Handbook of physiology. Section 4, environmental physiology. New York: Oxford University Press,:15785
237. Shirreffs SM, Armstrong LE, Cheuvront SN (2004). Fluid and electrolyte needs for preparation and recovery from training and competition . Journal of Sports Sciences, 22:5763
238. Tabata I, Atomi Y, Miyashita M (1984). Blood glucose concentration dependent ACTH and cortisol responses to prolonged exercise. Clinical Physiology 4:299-307
239. Tzintzas O-K, Williams C, Wilson W, Burrin, J (1996). Influence of carbohydrate supplementation early in exercise on endurance running capacity. Med. Sci. Sports Exerc. 28:1373-1379
240. Walsh RM, Noakes TD, Hawley JA, Dennis SC (1994). Impaired high-intensity cycling performance time at low levels of dehydration. Int. J. Sports Med. 15:392398
241. Vrigens DMG, Rehrer NJ (1999). Sodium-free fluid ingestion decreases plasma sodium during exercise in the heat. J Appl Physiol. 86:18471851
242. Speedy DB, Noakes TD, Rogers IR, Thompson JMD, Campbell RGD, Kuttner JA, Boswell DB, Wright S, Hamlin M (1999). Hyponatremia in ultradistance triathletes. Med. Sci. Sports Exerc. 31:809815
243. Shirreffs SM, Taylor AJ, Leiper JB, Maughan RJ (1996). Post-exercise rehydration in man: Effects of volume consumed and drink sodium content. Med. Sci. Sports Exerc. 28:1260 1271
244. Maughan RJ, Leiper JB, Shirreffs SM (1996). Restoration of fluid balance after exercise-induced dehydration: Effects of food and fluid intake. Eur. J. Appl. Physiol. 73:317325
245. Costill DL and Sparks KE (1973). Rapid fluid replacement following thermal dehydration. J. App. Physiol. 34:299303
246. Nielsen B, Sjogaard G, Ugelvig J, Knudsen B, Dohlmann B (1986). Fluid balance in exercise dehydration and rehydration with different glucoseelectrolyte drinks. Eur. J. App. Physiol. 55:318325
247. Nose H, Mack GW, Shi X, Nadel ER (1988). Role of osmolality and plasma volume during rehydration in humans. J. App. Physiol. 65:325331
248. Yawata T (1990). Effect of potassium solution on rehydration in rats: comparison with sodium solution and water. Jap. J. Physiol. 40:369381
249. Wapnir RA adn Lipshitz F (1985). Osmolality and solute concentration their relationship with an oral hydration solution effectiveness: an experimental assessment. Pediatr. Res. 19:894-898
250. Cunha Ferreira RMC, Elliot EJ, Brennan EA, Walker-Smith JA, Farthing MJG (1987). Glycine and glucose polymer in oral rehydration solution (ORS). Pediatr. Res. 22:100
251. Farthing MJG (1990). Studies of oral rehydration solution in animal models. Clin. Ther. 12(A):51-60
252. Hunt JB, Carnaby S, Farthing MJG (1991). Assessment of water and solute absorption from experimental hypotonic and established oral rehydration solutions in secreting rat intestine. Aliment Pharmacol Ther 5:273-281
253. Fordtran JS (1975). Stimulation of active and passive sodium absorption by sugars in the human jejunum. J. Clin. Invest. 55:728-737
254. Gisolfi CV, Summers RW, Schedl HP, Blejler TL (1992). Intestinal water absorption from select carbohydrate solutions in humans. J. App. Physiol. 73:2142-2150
255. Murray R (1987). The effects of consuming carbohydrate electrolyte beverages on gastric emptying and fluid absorption during and following exercise. Sport Med. 4:322-351
256. Nalin DR, Cash RA, Rahman M, Yunus MD (1970). Effect of glycine and glucose on sodium and water absorption in patients with cholera. Gut 11:768-722
257. Patra FC, Mahalanabis D, Jalan KN (1982). Stimulation of sodium and water absorption by sucrose in the rat small intestine . Acta Pediatr. Scand 71:103-107
258. Sandhu DK, Christobal FL, Brueton MJ (1989). Optimizing oral rehydration solution composition in model systems: studies in normal mammalian small intestine. Acta Pediatr. Scand. Suppl. 364:17-22
259. Shi X, Summers RW, Schedl HP, Flanagan SW, Chang R, Gisolfi CV (1995). Effects of carbohydrate type and concentration and solution osmolality on water absorption. Med. Sci. Sports Exerc. 27(12):1607-15
Un episodio agudo de ejercicio aeróbico reduce el tiempo de movimiento en una tarea de Fitts
El movimiento tiene una importancia crucial en el control motor, reflejado a través del tiempo de...
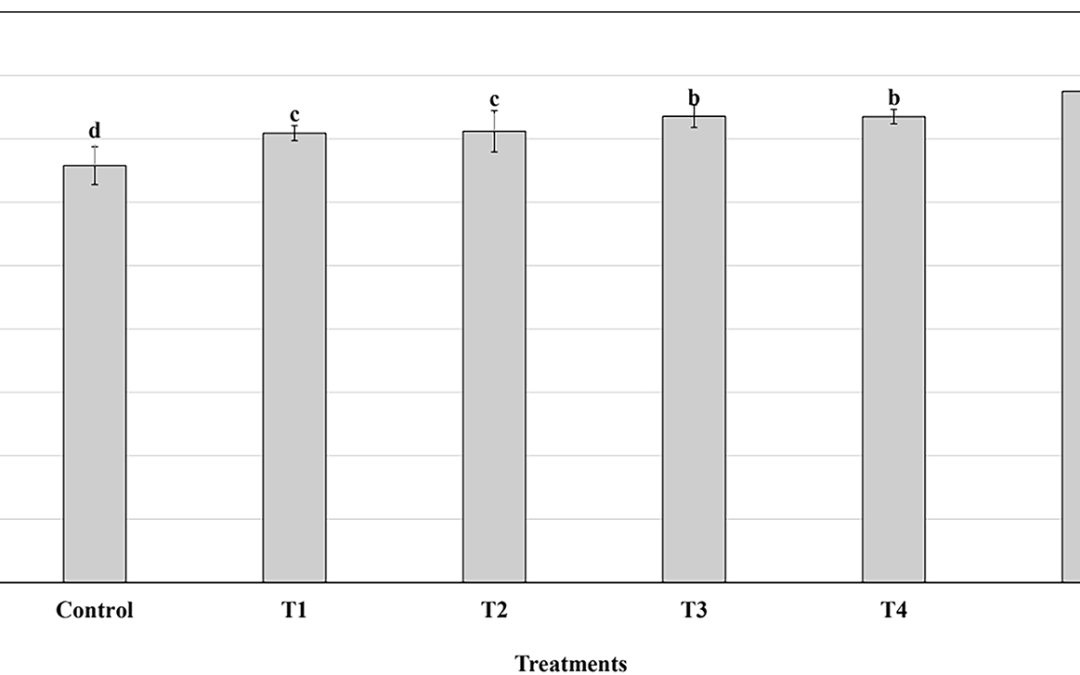
Mejora de las propiedades nutricionales y de textura del jerky reestructurado con polvo de sangre de pato para aplicaciones en kits de comida
Resumen del Estudio sobre el Uso de Polvo de Sangre de Pato en Jerky Reestructurado En un contexto...
El ejercicio aeróbico a lo largo de la vida protege contra la inflamaging y el cáncer
Ejercicio Aeróbico y su Impacto en el Envejecimiento y el Riesgo de Cáncer La investigación...
La combinación de nandrolona y entrenamiento de resistencia indujo remodelación cardíaca y estrés oxidativo a pesar de la contractilidad mejorada de los cardiomiocitos
Este artículo investiga cómo la combinación de entrenamiento de resistencia (RT) y el uso de...
El efecto del ciclismo en la función cognitiva y el bienestar en adultos mayores
El impacto del ciclismo al aire libre en la función cognitiva y bienestar de los adultos mayores...