
Ciencia y Ciclismo: Conocimientos Actuales y Lineamientos Futuros para la Investigación
Greg Atkinson1, Richard Davidson2, Asker Jeukendrup3 y Louis Passfield4
1School of Sport and Exercise Sciences, Loughborough University, Reino Unido.
2International Sports Consultancy, Brisbane, QLD, Australia.
3School of Sport and Exercise Sciences, The University of Birmingham, Birghimgham, Reino Unido.
4School of Applied Sciences, The University of Glamorgan, Pontypridd, Reino Unido.
Artículo publicado en el journal PubliCE, Volumen 0 del año 2003.
Publicado 19 de mayo de 2004
Resumen
Palabras clave: aerodinámica, eficiencia, ergometría, nutrición, ritmo de carrera
INTRODUCCION
El ejercicio en cicloergómetro es una forma común de ejercicio para las investigaciones generales en las ciencias del deporte y el ejercicio. El movimiento relativamente restringido el tren superior y los brazos durante el ciclismo facilita la tarea de los investigadores para hacer mediciones invasivas, por ejemplo, usando un catéter (MacLaren et al., 1999), mientras el ciclista puede ejercitarse por largos períodos de tiempo. Además, el ciclismo está generalmente menos asociado con lesiones que la carrera. De este modo, los estudios de entrenamiento pueden ser más fáciles de implementar y pueden ser menos afectados por el abandono de los participantes. Los ciclistas de competición se benefician de tales investigaciones de modo que los resultados son generalmente relevantes a su modo de ejercicio elegido.
No obstante, para que los resultados de las investigaciones sean directamente relevantes a los ciclistas competitivos, las investigaciones necesitan integrar las influencias de varios factores, los cuales son resumidos en el modelo presentado en la Figura 1. Primero, el investigador debería considerar las características fisiológicas de los atletas que son específicos para el evento (e.g. composición de fibras, máximo consumo de oxígeno). Estas características inherentes influencian la potencia que puede ser generada en una carrera de ciclismo. Además de la capacidad fisiológica inherente, las estrategias de entrenamiento, nutrición y ritmo de carrera tienen una fuerte influencia sobre la producción de potencia total de la carrera (Figura 1). La posición del ciclista en la bicicleta, predominantemente gobernada por el desplazamiento vertical y horizontal del asiento, también puede influenciar la producción de potencia.
Hasta aquí, han sido mencionados los múltiples factores que influencian la potencia de ciclismo, pero también hay varios factores que influencian la relación entre la potencia y la velocidad de pedaleo (Figura 1). El ciclista que, a través de una capacidad superior, tiene el potencial para generar la mayor potencia durante una carrera, puede no necesariamente ganar la carrera. El ciclismo es un deporte de alta velocidad, y las carreras pueden ser llevadas a cabo en todos los terrenos y ambientes. De este modo, las fuerzas físicas resistivas presentes en el ambiente de carrera (e.g. subidas o vientos) influencian enormemente la relación entre la potencia de pedaleo y la velocidad total de carrera. La presencia de diferentes fuerzas resistivas en una carrera puede influenciar la elección del diseño de la bicicleta o sus componentes. La Figura 1 también muestra que la posición del ciclista y la estrategia de ritmo de carrera no afectan solamente la producción de potencia fisiológica. La posición del ciclista también puede afectar la extensión a la cual la resistencia del aire influencia la relación potencia-velocidad. La estrategia de ritmo de carrera también puede influenciar la magnitud del efecto del viento y la pendiente sobre la relación potencia-velocidad.
Como un ejemplo específico de la naturaleza diversa del ciclismo competitivo, durante la temporada de carrera de 1996, Chris Boardman compitió en el Tour de Francia, una carrera de 3 semanas con etapas de distancias (10 a 250 km) y terrenos variados, y poco tiempo después participó en la persecución individual de 4 km en los Campeonatos Mundiales. Tales severas intensidades y duraciones de ejercicio en los meses antes de la persecución de 4 km parecerían ir en contra del corriente pensamiento de periodización del entrenamiento para el logro de un rendimiento óptimo. No obstante, Chris Boardman ganó el Campeonato Mundial en el evento de 4 km. Boardman compitió en el Campeonato Mundial con una posición aerodinámica nueva que habría disminuido la resistencia del aire, pero pudo haber tenido una influencia paradójica sobre la posición del corredor para la producción de potencia óptima (Gnehm et al., 1997). Tales observaciones empíricas sirven para ilustrar la importancia de considerar las muchas, y algunas veces competitivas influencias sobre el rendimiento en ciclismo.
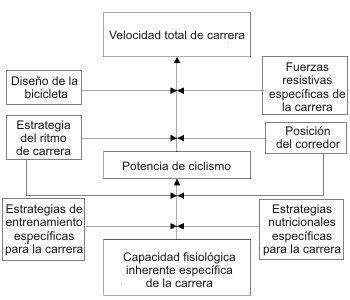
Figura 1. Diagrama que muestra los varios factores que influencian la
producción de potencia y la velocidad en el ciclismo, en línea con los tópicos
cubiertos en la presente revisión.
Varias excelentes revisiones recientemente publicadas sobre la ciencia del ciclismo han considerado la naturaleza discordante de los eventos de ciclismo (e.g. Burke, 2000; Neuman, 2000). En estas recientes revisiones fue enfatizada la especificidad para el evento de las estrategias de entrenamiento. Por lo tanto, los factores de entrenamiento no son considerados en la presente revisión. Es solo relativamente reciente que los investigadores han comenzado a considerar completamente todos los factores presentados en la Figura 1, y las interrelaciones entre estos factores, que caracterizan al ciclismo de competición. Por ejemplo, los test en ergómetro que reflejan la naturaleza intermitente de las carreras de ciclismo en ruta y que simulan las fuerzas resistivas variables que un corredor podría experimentar durante una prueba por tiempo han sido utilizados solo en los últimos 10 años (Palmer et al., 1996b; Atkinson y Brunskill, 2000).
El objetivo de este trabajo es proveer una revisión de las investigaciones recientes sobre ciclismo que han intentado “cerrar el circuito” entre el laboratorio y el ambiente de las carreras. Un enfoque “holístico” es adoptado en un intento de discutir las investigaciones actuales y futuras pertenecientes a cada factor teniendo en mente las interrelaciones entre otros factores. El trabajo esta estructurado teniendo en base a la Figura 1. Cada uno de estos factores que influencian la velocidad de ciclismo van a ser discutidos, excepto, como fue mencionado arriba, las estrategias de entrenamiento. Cada una de estas discusiones van a terminar con sugerencias para las investigaciones relevantes en el futuro.
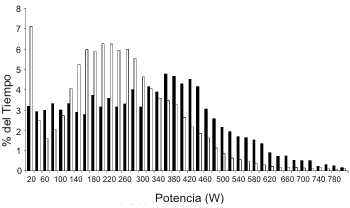
Figura 2. Distribución de la producción de potencia durante rendimientos
individuales en carreras de ruta, uno de una carrera de ruta profesional de 6 h
(■) y uno de una carrera de ruta amateur de 3,5 h (□). Datos no publicados
obtenidos a partir de uno de los autores (R.D.).
FACTORES FISIOLOGICOS
A pesar de que hicimos hincapié en la naturaleza diversa de los eventos de ciclismo, el ciclismo es predominantemente un deporte de resistencia. Ha sido demostrado que aún una prueba de ciclismo que dure menos de 5 min está altamente correlacionada con la potencia aeróbica máxima (Passfield y Doust, 2000). Las excepciones serían los eventos de velocidad de 200 m y 1 km. La optimización del rendimiento para estos últimos eventos es mencionada en una sección posterior de la revisión. En esta sección sobre factores fisiológicos, vamos a tratar principalmente con los aspectos fisiológicos del metabolismo anaeróbico que afectan el rendimiento de ciclismo. Para una revisión más detallada de los factores de integración que afectan el rendimiento de resistencia general, ver Coyle (1995) y Hawley y Stepto (2001).
La mayoría de la información nueva que revisamos viene de estudios que implicaron ciclistas profesionales europeos. Aunque tal información es de gran interés, debe ser tenida precaución a la hora de la extrapolación de esta información a todos los patrones competitivos del ciclismo. El ciclista profesional típico cubrirá 25000 a 35000 km en un año, con más de 90 días de competición y la participación en al menos uno de los principales tours de 3 semanas (Giro d’ Italia, Tour de Francia, Vuelta a España). En contraste, el ciclista amateur cubrirá normalmente no más de 25000 km en un año, teniendo menos de 50 días competitivos, y con competiciones de normalmente no más de dos días consecutivos. La Figura 2 ilustra el tiempo relativo gastado a diferentes producciones de potencia en carreras de ruta típicas profesionales y amateurs que duran 6 y 3,5 h, respectivamente. Claramente, hay una diferencia en los perfiles de potencia relativa, y de este modo, las posibles demandas fisiológicas y los determinantes del éxito también serían diferentes.
A pesar de posibles diferencias entre los patrones de competición, los factores fisiológicos clave que están relacionados al éxito en el ciclismo son la potencia aeróbica máxima (VO2 máx.), el tipo de fibras musculares, la eficiencia de pedaleo y el umbral del lactato. Las investigaciones en las cuales ha sido sugerido que estos factores son importantes van a ser discutidas ahora en detalle.
Potencia Aeróbica Máxima
Como es el caso de otros atletas basados en la resistencia, una observación consistente es que los ciclistas de clase mundial tienen un gran VO2 máx. Un rango absoluto de 5.0-6.4 l.min-1 o rango relativo de 70-85 ml.kg-1.min-1 es típico de los ciclistas profesionales (Coyle et al., 1991; Lucia et al., 1998, 2001a; Mujika y Padilla, 2001). Los valores relativos altos (>80 ml.kg-1.min-1) tienden a ser observados para escaladores especialistas que tienen una masa corporal relativamente baja (<65 kg) (Mujika y Padilla, 2001).
La mayoría de las investigaciones no han encontrado diferencias en el VO2 máx. entre ciclistas altamente entrenados y profesionales de élite (Lucia et al., 1998). La mayor diferencia entre la élite y la sub-élite es la potencia producida en el VO2 máx. De este modo, es más común para los investigadores referirse a la potencia aeróbica de los ciclistas en términos de potencia máxima a partir de un protocolo incremental o en rampa conduciendo al agotamiento en aproximadamente 10-60 min. Tales producciones de potencia máxima varían de manera característica desde 350 a 525 W (5.5 a 7.6 W.kg-1) para los ciclistas top (Lucia et al., 2001b; Mujika y Padilla, 2001). Ha sido demostrado que la potencia máxima obtenida a partir de este tipo de test es un buen predictor del tiempo para completar 20 km (r=-0.91; Hawley y Noakes, 1992) y una potencia promedio sobre 16 km (r=-0.99; Balmer et al., 2000).
Desafortunadamente, la potencia máxima es influenciada tanto por el protocolo de evaluación, como por el ergómetro elegido (Mujika y Padilla, 2001). Aunque ninguna investigación ha hecho una comparación directa de los protocolos comúnmente adoptados usando los mismos participantes, las comparaciones de los valores obtenidos a partir de diferentes protocolos con individuos tomados de la misma población (ciclistas profesionales) sugiere que los protocolos incrementales con etapas más largas tienden a dar valores máximos más bajos (400-450 W, 6.0-6.5 W.kg-1) en comparación a los protocolos de incrementos y rampas más cortas (450-500 W, 6.5-7.5 W.kg-1) (Lucia et al., 2001a).
Intensidad “Umbral” de Ejercicio
Cuando se revisa la literatura acerca de este tópico, una complicación es la variedad de términos usados para describir una intensidad de ejercicio medida a un cierto umbral fisiológico. En al literatura han sido citados el umbral del lactato (Billat, 1996), umbral anaeróbico (Wasserman y McIlroy, 1964), umbral anaeróbico individual (Stegman et al., 1981), primer umbral ventilatorio, segundo umbral ventilatorio (Lucia et al., 2002b) y punto de compensación ventilatoria (Lucia et al., 2002b). El uso de definiciones ligeramente diferentes para cada término crea una confusión adicional. Varios términos comunes son, sin embargo, evidentes. El rendimiento superior requiere claramente un alto umbral, sin embargo, como fue definido y discutido en detalle arriba, la intensidad de ejercicio en el tan llamado umbral es virtualmente siempre un mejor predictor del rendimiento que el VO2 máx.
El rango típico para la intensidad umbral de ejercicio, abarcando todas las definiciones, para ciclistas hombres entrenados es 78-90% del VO2 máx. (Coyle et al., 1991; Wilber et al., 1997; Lucia et al., 1998). De manera predictiva se puede afirmar que el límite superior de los valores para el umbral anaeróbico viene de ciclistas profesionales: segundo umbral ventilatorio= 92.1 % del VO2 máx., 87 % de la potencia máxima o 91.6 % de la frecuencia cardiaca máxima (Lucia et al., 2001b).
Usando la frecuencia cardiaca como un indicador de la intensidad de ejercicio, Fernandez-García et al. (2000) estimaron que la cantidad de tiempo que los ciclistas profesionales pasan arriba del 90 % del VO2 máx. en cualquier etapa individual de una vuelta importante es de aproximadamente 20 min, sin tener en cuenta el tipo de etapa (llana o montañosa). Esta observación condujo a los mismos a sugerir que este tiempo representa alguna limitación en la capacidad (Fernandez-García et al., 2000). En contraste, Lucia et al. (1999a) sugirieron que la cantidad de tiempo arriba del segundo umbral ventilatorio (≈88 % del VO2 máx.) depende del tipo de etapa, siendo mayor para las pruebas por tiempo individuales seguidas por las etapas de montaña y las etapas llanas. Ellos también sugirieron que la capacidad para rendir por extensos períodos de tiempo a esta intensidad de ejercicio es necesaria para tener éxito con estos patrones de competición (Lucia et al., 1999b). En la actualidad ha habido muy pocos estudios acerca de la capacidad de sujetos altamente entrenados o ciclistas profesionales para ejercitarse sostenidamente en o ligeramente arriba de una intensidad correspondiente al segundo umbral ventilatorio. Coyle et al. (1988) mostraron que un grupo de ciclistas con un alto umbral láctico se ejercitaron durante un período de tiempo más largo hasta la fatiga al 88 % del VO2 máx. que un grupo con un umbral láctico más bajo, pero este estudio no incluyó ciclistas profesionales.
Lucia et al. (1999b) y otros (e.g., Coyle et al., 1991) han mostrado que los ciclistas de ruta de alto nivel pueden sostener intensidades arriba del 90 % del VO2 máx. por más de 1 h. A estas intensidades, sería anticipado que hay también una contribución anaeróbica al gasto energético. En base a frecuencias cardiacas registradas, la contribución total del ejercicio anaeróbico en una vuelta importante es relativamente bajo (≤ 13 %) (7 %, Lucia et al., 1999b; 13 % Fernanez-García et al., 2000). No obstante, la importancia con respecto al resultado principal podría ser considerable ya que las mayores contribuciones anaeróbicas vienen de etapas de vueltas que normalmente determinan el resultado (pruebas por tiempo individuales, y etapas de montaña) (Lucia et al., 1999b).
Cinética del Consumo de Oxígeno
La Figura 2 demuestra la distribución variable de la potencia relativa de ciclismo en las carreras de ruta. Actualmente, no ha habido ninguna investigación acerca de la cinética del consumo de oxígeno para ciclistas profesionales en el ejercicio realizado en el laboratorio o durante la realización de ciclismo de ruta simulado. Teóricamente, sería beneficioso tener una rápida respuesta en el consumo de oxígeno ante el ejercicio, ya que esto reduciría la contribución de las vías anaeróbicas para suministrar la energía para los cambios rápidos en la intensidad de ejercicio. Uno de los problemas con la investigación actual acerca del la cinética del VO2 es la tendencia a usar ciclistas no entrenados y ejercicios en condiciones de laboratorio, empezando desde el reposo (e.g. Ozyener et al., 2002), los cuales no replican las circunstancias de las carreras de ruta, donde los períodos intermitentes de actividad de alta intensidad están superpuesto a largos períodos de trabajo de intensidad baja a moderada. Actualmente hay un pobre entendimiento acerca de que pasa con la cinética del VO2 después de varias horas de ciclismo durante el cual la intensidad de ejercicio varía.
Eficiencia
Una de las importantes capacidades funcionales del ciclista, que Coyle (1995) sugirió que está fuertemente relacionada al rendimiento de resistencia, es la eficiencia mecánica total [(trabajo realizado/energía consumida) x 100]. Ha sido reportado que la eficiencia mecánica total durante el ciclismo está entre 18 y 26 % en ciclistas de competición hombres bien entrenados (VO2 máx.= 69 ml. kg-1.min-1; Coyle et al., 1992), entre 23.6 y 25,6 % en ciclistas de competición amateurs (VO2 máx. = 4.9 L.min-1; Lucia et al., 2002a), y entre 24.4 y 28.8 % en ciclistas profesionales (VO2 máx.= 5.0 L.min-1; Lucia et al., 2002a). No hay diferencia en la eficiencia metabólica total entre los ciclistas profesionales y los amateurs a cargas de trabajo bajas (Lucia et al., 2002a). En estudios previos, fue sugerido que tanto la eficiencia mecánica total como el delta de eficiencia [(Δ del trabajo realizado/Δ del gasto energético) x 100] están relacionados a la proporción de fibras Tipo I del músculo (Coyle et al., 1992). Más recientemente, Marsch et al. (2000) demostraron que no hay diferencias en el delta de eficiencia para diferentes cadencias que iban de 50 a 100 rev.min-1 para corredores, personas no ciclistas menos entrenadas o ciclistas entrenados. Sin embargo, los autores comentaron que, aunque no era estadísticamente diferente, los ciclistas entrenados exhibieron un delta de eficiencia que fue ligeramente mayor (≈ 1-2 %) a través del rango de cadencias. La producción de potencia más alta usada para estas comparaciones fue 200 W, la cual sería considerada como una intensidad de ejercicio relativamente baja para un ciclista profesional. Es posible que hubieran sido notadas mayores diferencias para las intensidades cercanas a las intensidades de competición. Horowitz et al. (1994) mostraron que, cuando un grupo de ciclistas de competición fue dividido de acuerdo a la proporción de fibras Tipo I (alta= 73 %, normal= 48%), el subgrupo que tubo mayor proporción de fibras Tipo I produjo potencia significativamente más alta para un VO2 dado durante una evaluación de 1 h, indicando una mayor eficiencia.
Los resultados de una reciente investigación, la cual implicó un protocolo en rampa, sugirieron que los corredores profesionales tienen una mayor eficiencia mecánica total a intensidades arriba del umbral del lactato que los corredores entrenados amateur, con un incremento de la diferencia con mayores intensidades de ejercicio (Lucia et al., 2002a). No obstante, la validez de la eficiencia mecánica inusualmente alta (28.8 %) reportada por Lucia et al. (2002a) y en un estudio posterior realizado también por Lucia et al. (2003c) ha sido recientemente cuestionada (Jeukendrup y Martin, 2003). Como indicación de un posible beneficio de un incremento de la eficiencia, Jeukendrup y Martin (2001) calcularon que incrementando la eficiencia total en un 1% para un ciclista de 70 kg que puede mantener una producción de potencia de 400 W por 1 h resultaría en una mejora de 48 s para una prueba por tiempo sobre 40 km.
Un aspecto de la eficiencia que permanece relativamente no investigado es el cambio de la eficiencia a medida que el ejercicio progresa. Los resultados de un estudio demostraron que las reducciones en la eficiencia que resultaron de ≈ 60 min de ejercicio de intensidad constante (60 % del VO2 máx.) están relacionadas a la reducción en la producción de potencia durante una serie de alta intensidad (5 min) realizada antes y después del ejercicio de intensidad constantes (Passfield y Doust, 2000). De este modo, el rendimiento de ciclismo podría ser determinado tanto por la alta eficiencia total y los cambios en la eficiencia como resultado del ejercicio continuo. Actualmente está poco claro si los ciclistas profesionales exhiben una reducción aún más atenuada en la eficiencia de pedaleo a medida que el ejercicio progresa en comparación con los ciclistas entrenados, y también hasta que extensión los cambios en la eficiencia son afectados por el ejercicio de intensidad variable más que por el ejercicio de intensidad constante.
Capilarización Muscular
La importancia de la capilarización muscular para el rendimiento de resistencia ha sido destacada en varias revisiones (Coyle, 1995; Hawley y Stepto, 2001). No obstante, son muy limitados los estudios sobre la capilarización muscular y el rendimiento en ciclismo, y en la actualidad no ha habido investigaciones realizadas en ciclistas profesionales. Coyle et al. (1988) reportaron una fuerte correlación (r= 0.74) entre la densidad capilar y el tiempo hasta la fatiga a ≈ 88 % del VO2 máx. en un grupo de ciclistas bien entrenados. En un estudio posterior, Coyle et al. (1991) compararon un grupo de ciclistas nacionales de élite con corredores regionales y mostraron que los corredores élite tenían una densidad capilar muscular un 23 % mayor. La densidad capilar estuvo significativamente correlacionada con la tasa de trabajo absoluta promedio para un ejercicio de 1 h. La importancia de la densidad capilar está claramente ilustrada por el hecho de que, en ambos de los estudios arriba citados, la densidad capilar fue una variable clave en las ecuaciones de regresión para predecir el rendimiento.
Tipos de Fibras
A partir de la información disponible, parecería que, para ser un buen ciclista de resistencia, es necesario tener o alcanzar una muy alta proporción de fibras musculares Tipo I (Coyle et al., 1991; Bishop et al., 2000; Lucia et al., 2002b) o, a la inversa, una baja proporción de fibras Tipo IIx (Lucia et al., 2002b). Otra evidencia de la importancia de las fibras Tipo I, y de la optimización de su uso, son las cadencias típicamente usadas por los ciclistas profesionales. Estas cadencias relativamente altas (> 90 rev.min-1) no tienden a minimizar el consumo de oxígeno, sino que minimizan la fuerza por golpe de pedal (Lucia et al., 2001b). Esta disminución de fuerza por golpe de pedal mediante cadencias más altas parecería ser un mecanismo por medio del cual el reclutamiento de fibras Tipo II es minimizado (Ahlquist et al., 1992).
Hay buena evidencia que sugiere que la cadencia preferida de los ciclistas entrenados tiende a aumentar con el incremento de la carga de trabajo (Marsch y Martin, 1997). Por ejemplo, cuando son requeridas cargas de trabajo altas (≈ 400 W o ≈ 90 % del VO2 máx.) por un considerable período en carreras por tiempo, los ciclistas profesionales adoptan cadencias altas (≈ 93 rev.min-1) para un rendimiento óptimo (Lucia et al., 2001b). Cuando las condiciones ambientales son muy constantes y las producciones de potencia son altas y uniformes (> 600 W), por ejemplo, en una persecución por equipos en la pista, las cadencias preferidas son extremadamente altas (133-139 rev. min-1) (Broker et al., 1999).
Dada la información antes citada, es extraño que las cadencias usadas por los ciclistas para las subidas sean considerablemente bajas (≈ 60-80 rev.min-1) (Davison et al., 2000; Lucia et al., 2001b). Presumiblemente la potencia requerida es también alta en las subidas. Una examinación más minuciosa de los ciclistas de mayor nivel en las etapas de montaña sugiere que ellos adoptan cadencias relativamente altas en vez de un índice de desarrollo alto (Lucia et al., 2001b). Esta observación ha sido también recientemente enfatizada por el notable rendimiento de un corredor (Lance Armstrong) en 2001 en la subida de Alpe d’Huez, cuando alcanzó la velocidad promedio de 22 km.h-1 (se estima que para lograr esto se requiere de una producción de potencia de ≈ 500 W), el corredor mantuvo un promedio de 100 rev. min-1 durante los 40 min de duración de la subida (Lucia, 2001b). No obstante, vale la pena señalar que el segundo corredor en arribar al la parte superior de Alpe d’Huez en 2001 (Jan Ulrich) usó un desarrollo mucho mayor y tuvo una cadencia correspondiente menor (≈ 70 rev. min-1), similar a aquella medida para otros corredores profesionales en subida (≈ 71 rev. min-1, Lucia et al., 2001b), pero todavía rápida para vencer a todos, menos un corredor. Si las cadencias más altas mejoran el rendimiento en ciclismo, los posibles mecanismos podrían incluir la reducción de la fuerza por golpe de pedal para ya sea, minimizar el reclutamiento de fibras musculares Tipo II o para optimizar el uso de las más eficientes y resistentes a la fatiga fibras Tipo I, o para mejorar el flujo sanguíneo a la masa muscular activa (Takaishi et al., 2002). Ha sido también demostrado que cadencias de pedaleo más altas resultan en un bombeo más efectivo de los músculos esqueléticos, lo cual incrementa el flujo sanguíneo y el retorno venoso (Gotshall et al., 1996). Son necesarios trabajos futuros para identificar las razones de las diferencias interindividuales en la selección de la cadencia, y si entrenar a un ciclista para usar diferentes cadencias realmente incrementa el rendimiento.
Las consecuencias fisiológicas de un mayor número de fibras Tipo I altamente entrenadas es la capacidad para mantener altas producciones de potencia por períodos largos, así como una alta eficiencia mecánica total. Complementar estas características sería tener un alto nivel de capilarización para abastecer de suficiente oxígeno y sustratos para mantener las grandes producciones de potencia. Ya que muchos de los recientes datos sobre rendimiento de ciclismo han sido obtenidos de ciclistas profesionales, es difícil dilucidar la extensión a la cual las adaptaciones a estas cargas fisiológicas contribuyen a los atributos medidos en estos atletas.
Debería ser destacado que una alta proporción de fibras musculares Tipo I puede estar asociado con el éxito en las vueltas más importantes y los eventos por tiempo, pero que muchos corredores pueden tener más victorias individuales que el ganador general de las vueltas. El ganador es considerado el corredor más rápido en general, no el más consistente llegador (aunque usualmente también hay un “maillot de ganador para este corredor”), el cual es convencionalmente el corredor con la mejor capacidad para esprintar. El objetivo del corredor “sprinter” o velocista es completar la mayor parte de la carrera con el menor costo energético posible, y luego, usualmente con la ayuda de los compañeros de equipo, producir una aceleración de muy alta intensidad al final de la carrera. Para lograr estas llegadas de muy alta intensidad (> 1500 W para los últimos 200 m), podría ser que la distribución de fibras de estos corredores sea diferente a la de aquellos corredores que terminan muy arriba en la clasificación general. No obstante, no ha habido ninguna investigación para caracterizar completamente a estos corredores sprinters en particular. Los estudios que han implicado entrenamiento de la fuerza explosivo (Bastiaans et al., 2001) o entrenamiento intervalado (Stepto et al., 1999) para mejorar el rendimiento general de los ciclistas entrenados no produjeron consistentemente resultados favorables. Es también interesante señalar que ninguno de los recientes ganadores generales de ninguna de las vueltas más importantes sería considerado como un sprinter de alto nivel. Para confundir más aún la estimación de la posible contribución de tipo de fibras/anaeróbica, es común para los corredores de alto nivel en las carreras por etapas más importantes ganar o al menos estar posicionado muy arriba en las pruebas por tiempo prólogos, las cuales tienen en general menos de 10 km, y requerirán indudablemente cierta contribución anaeróbica (< 10 %) para un alto rendimiento.
Demanda Ventilatoria
Varios estudios recientes han demostrado que los ciclistas profesionales mantienen producciones de potencia y porcentajes del VO2 máx. muy altos, particularmente durante las pruebas por tiempo (Lucia et al., 2001a). Obviamente, para mantener tan altas cargas de trabajo, hay una considerable demanda ventilatoria. La ventilación promedio de los ciclistas entrenados durante una prueba por tiempo bajo techo de 40 km ha sido medida en 111±13 l.min-1 (Smith et al., 2001), y hubiera sido lógico concluir que para duraciones más cortas (<40 km, pero >5 km), en carreras de más intensidad, la demanda ventilatoria sería aún mayor. Romer et al. (2002) demostraron recientemente que después de una prueba por tiempo, los músculos inspiratorios exhiben una fatiga considerable con una tendencia a una mayor fatiga para las pruebas de menor duración y mayor intensidad (20 vs. 40 km). Estos investigadores también reportaron que 6 semanas de entrenamiento específico para los músculos inspiratorios pudieron reducir significativamente esta fatiga, resultando en mejores rendimientos en las pruebas por tiempo de 20 y 40 km.
No han sido reportadas mediciones del rendimiento de los músculos inspiratorios para ciclistas profesionales. De este modo, en la actualidad, no está claro si estos atletas tienen músculos inspiratorios más desarrollados como resultado de su alta carga de entrenamiento o si ellos, también, se podrían beneficiar como resultado de un entrenamiento específico para los músculos inspiratorios. Sin embargo, en un estudio fue sugerido que los ciclistas profesionales exhiben diferentes patrones respiratorios durante el ejercicio incremental que los ciclistas entrenados amateur de élite (Lucia et al., 1999b). No hubo ninguna diferencia en la ventilación máxima entre los dos grupos, pero los corredores amateurs tuvieron ventilaciones minuto significativamente más altas (VE) a 300, 350, y 400 W y mayores frecuencias respiratorias en la mayoría de las intensidades moderadas a máximas. En contraste, los corredores profesionales parecían adoptar tiempos espiratorios prolongados y no hubo ninguna evidencia de una variación taquipneica a altas intensidades de ejercicio, y el incremento en la ventilación ocurrió debido a un incremento en el volumen corriente y en la frecuencia respiratoria. No obstante, nosotros reconocemos que algunas de las diferencias antes citadas podrían ser explicadas por las diferentes intensidades relativas de ejercicio, particularmente en relación a la compensación respiratoria de la acidosis metabólica.
Lineamientos Futuros
Desde un punto de vista de las ciencias del deporte, los ciclistas profesionales representan una oportunidad para investigar la adaptación que ocurre como respuesta a la sobrecarga masiva del ejercicio de resistencia. Esta sobrecarga es probablemente mayor que para cualquier otro atleta. Desafortunadamente, hasta el reciente trabajo de Lucia, Padilla y colaboradores, el ciclismo de ruta de élite era un área que estaba relativamente bajo investigación y muchas preguntas continúan sin respuesta. Incluso este trabajo más reciente se ha focalizado sobre las respuestas fisiológicas a las intensidades de ejercicio que han sido meramente estimadas a partir de la frecuencia cardiaca. La adaptabilidad de los instrumentos de medición de potencia sobre la bicicleta proporciona la oportunidad para la realización de más estudios acerca de las respuestas fisiológicas al ciclismo competitivo, mientras se conoce la potencia exacta en las carreras.
Los ciclistas profesionales han sido entendiblemente muy reacios a donar tejido muscular para propósitos de investigación. De este modo, virtualmente nada es conocido acerca de las características morfológicas y biomecánicas de sus músculos. Ha habido también poca investigación acerca de la correlación entre los cambios en el rendimiento o la eficiencia mecánica dentro de un ciclo de entrenamiento anual y los cambios en las respuestas cardiacas, enzimáticas y de los músculos esqueléticos. Los científicos del deporte han estado investigando las respuestas fisiológicas a diferentes cadencias de pedaleo por algún tiempo, aunque todavía no entendemos completamente la interacción entre la cadencia, carga de trabajo, eficiencia y ciclismo en subida. La interacción de la cinética del VO2 y la intensidad variable de ejercicio experimentada durante una carrera de ciclismo de ruta es otra área que requiere investigación.
FACTORES FISICOS
En la sección previa, desarrollamos las características fisiológicas que influencian la producción de potencia durante una carrera. Esta producción de potencia es requerida para vencer una compleja interacción de fuerzas resistivas presentes en el ambiente de competición. Consecuentemente, la potencia de carrera puede no necesariamente estar relacionada a la velocidad de pedaleo en una manera simple y directa. Dependiendo de la naturaleza del evento, estas fuerzas resistivas es probable que incluyan a la resistencia del aire, gravedad, resistencia de rodamiento, inercia y perdidas por fricción a partir de la cadena y rodamientos. La fuerza resistiva más importante en las velocidades de carrera normales en terrenos llanos viene principalmente de la resistencia del aire, con la resistencia de rodamiento y las pérdidas friccionales explicando probablemente menos del 10 % del gasto energético total (Kyle, 1996). La inercia del corredor y la bicicleta se vuelven significativamente importantes durante la aceleración y la deceleración, mientras que el efecto de la gravedad se vuelve un factor principal en el terreno con cuestas empinadas.
Resistencia del Aire
A velocidades de pedaleo arriba de 15 km.h-1 en un terreno nivelado, la resistencia del aire se convierte en la fuerza resistiva principal (Kyle, 1986). Esta resistencia aerodinámica se vuelve significativa debido a que la resistencia del aire se incrementa como aproximadamente una función cuadrática con respecto a la velocidad. La producción de potencia, en teoría, se espera que se incremente como una función cúbica en función de la velocidad, debido a que la potencia total de ciclismo es el producto de la resistencia del aire y la velocidad. No obstante, Bassett et al. (1999) observaron que el exponente promedio para la producción de potencia versus la velocidad de ciclismo es 2.6 para el ciclismo en pista techada. La razón de que el exponente es menor que 3 es probablemente debido a la combinación de todas las fuerzas resistivas, las cuales actúan con funciones lineales, cuadráticas y cúbicas. Por consiguiente, un modelo útil de ciclismo debería incorporar estos términos diferentes y no usar una función de potencia para hacer el modelo potencia-velocidad.
La resistencia aerodinámica de un ciclista es comúnmente representada por su coeficiente de resistencia (Cd). Este complejo coeficiente puede ser esencialmente resumido como la integración de la resistencia de presión, relacionada a la forma y al área frontal, y la resistencia de fricción, afectada por la uniformidad de superficie de un ciclista y su bicicleta. A las velocidades de carrera típicas, la presión de resistencia juega un rol mucho más significativo para determinar el coeficiente de resistencia de un ciclista (Kyle, 1996). Es la predominancia de la presión de resistencia lo que le permite al corredor beneficiarse de colocarse en la estela o “chuparse” (drafting) detrás de otro corredor. Por medio del drafting, el corredor se beneficia de pedalear dentro de un vórtice de presión creado por el ciclista líder. Los ciclistas que lo siguen pueden de este modo experimentar una reducción de 30 % en la potencia requerida en comparación con el corredor líder (Broker et al., 1999). Broker et al. (1999) encontraron que hay un ahorro subsiguiente de 6 % en la producción de potencia en el tercer lugar en comparación con el segundo lugar en un grupo, por las mismas razones. Alternando el trabajo adicional asociado con el liderazgo, los corredores en un equipo o grupo son subsiguientemente capaces de mantener velocidades significativamente mayores que aquellas de un ciclista solo. A partir de los datos de Broker et al. (1999) puede ser calculado que 4 corredores que se alternan la posición de punta pueden incrementar la potencia promedio generada en aproximadamente 25 % en comparación con aquella de un único corredor. Sin embargo esta magnitud de incremento en la potencia no va a estar completamente reflejada en los tiempos de carrera, debido a la desproporcionadamente alta producción de potencia requerida para las velocidades más altas.
En años recientes, los ciclistas de competición han adoptado posiciones de una aerodinámica incremental para ayudar a minimizar su resistencia e incrementar el rendimiento. Bassett et al. (1999) han calculado que el 60 % del incremento en la distancia cubierta en el record de la hora para el ciclismo de pista a través de los últimos 30 años es debido a la mejora de la aerodinámica más que al incremento de la aptitud física. La importancia de la aerodinámica es ejemplificada por un análisis del mejor record de la hora, de Chris Boardman de 56.375 km, establecido en 1996. Bassett et al. (1999) estimaron que Boardman promedió 20 W menos que Tony Rominger, el recordman anterior. No obstante, Boardman era 6 kg más pesado que Rominger y rodó más de 1 km más. Tal hallazgo es probable que sea explicado en gran parte por la posición extremadamente aerodinámica de “Superman” que adoptó Boardman, como hizo para las Olimpíadas, para su exitosa carrera de record. Esta posición fue subsiguientemente prohibida por el cuerpo gobernante del ciclismo mundial, incrementando efectivamente la producción de potencia media requerida para cualquier futuro intento exitoso de record en un 5 % estimativo. Gnehm et al. (1997) encontraron que pedalear con las manos colocadas en la parte curva del manubrio estaba asociado con un mayor costo metabólico que cuando se pedalea en una posición menos aerodinámica (con las manos colocadas en la parte superior del manubrio). No obstante, estos autores mantuvieron que las ventajas aerodinámicas asociadas con las posiciones con las manos colocadas en la parte baja del manubrio pesan más que cualquier efecto perjudicial sobre la capacidad de mantener la producción de potencia mientras se adopta estas posiciones.
Debería ser recordado que la resistencia aerodinámica es una función del movimiento relativo de aire, no simplemente de la velocidad (en el suelo). Por consiguiente, la producción de potencia en el ciclismo es determinada tanto por la velocidad horizontal como por la velocidad del aire. Por ejemplo, pedalear a 30 km.h-1 contra un viento de 10 km.h-1 requiere una potencia equivalente a pedalear a 40 km.h-1 en aire quieto. La Figura 3 representa los efectos de la velocidad del viento (viento de frente y de cola) y la inclinación (ascenso y descenso) sobre la velocidad de pedaleo. Esta relación está muy cerca de ser lineal (Martin et al., 1998). Los cambios en la densidad del aire también van a influenciar la resistencia aerodinámica. Los principales determinantes de la densidad del aire son la temperatura, la presión barométrica y en una menor extensión, la humedad. Los cambios en estas variables van a tener un efecto proporcional en la densidad del aire y consecuentemente, la potencia requerida para mantener una velocidad dada. De esta manera, una combinación de un incremento en la temperatura de 5 ° C y una disminución en la presión barométrica de 15 mmHg va a reducir la densidad del aire y la resistencia aerodinámica en aproximadamente 4 %.
Resistencia de Rodamiento
La resistencia al rodamiento contribuye significativamente al costo energético total en el ciclismo a bajas velocidades (< 15 km.h-1). Bajo condiciones comunes de carrera en ruta o pista, se piensa que la resistencia de rodamiento tiende a no ser prácticamente significativa, ya que otros factores se vuelven mucho más importantes. Este hecho puede ser observado cuando son examinados modelos de velocidad de ciclismo. Aunque el término de resistencia de rodamiento en estos modelos ha sido relacionado a la velocidad, la contribución a la producción de potencia total es pequeña (Martin et al., 1998; Bassett et al., 1999). Las variables más importantes que influencian la resistencia de rodamiento son la superficie en la que se pedalea, el diámetro de las ruedas, construcción y presión, y carga total sobre la rueda (Kyle, 1996). De estas variables, la presión de las ruedas es sobre las que más han influenciado los corredores. Sorprendentemente, Ryschon y Spray-Gundersen (1993) no encontraron un efecto discernible en el costo de oxígeno para hacer ciclismo a través de un rango de de presiones de las ruedas típicamente usado por los ciclistas (6-10 bar). No obstante, hay cierta posibilidad de que los cambios en la resistencia de rodamiento influencien el rendimiento a altas velocidades, ya que es generalmente estimado que la resistencia de rodamiento explica ≈10 % de la producción total de potencia cuando se pedalea a 40 km. h-1 (Kyle, 1986).
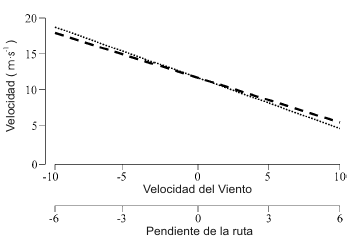
Figura 3. Influencia de la velocidad del viento y la dirección (línea con
rayas) y pendiente de la ruta (línea con puntos) sobre la velocidad para un
ciclista hipotético con una producción de potencia constante de 255 W, un área
de resistencia de 0.264 m2 y una masa de 71.9 kg. La influencia de estos dos
factores resistivos sobre la velocidad está muy cerca de ser lineal (r2 >0.99) a
través de un rango de velocidades del viento y pendientes presentes y para los
datos hipotéticos que fueron introducidos dentro del modelo matemático. Una
velocidad del viento negativa corresponde al viento de cola, un gradiente
negativo corresponde a un descenso. La Figura fue desarrollada a partir de los
datos recolectados bajo condiciones reportadas por Martin et al. (1998).
Pérdidas por Fricción
Se piensa que las pérdidas por fricción durante el ciclismo son relativamente pequeñas, pero variables (Kyle, 1988). Whitt y Wilson (1974) sugirieron que una cadena nueva y limpia pude producir pérdidas por fricción de solo 1.5 %. Kyle y Calloso (1986) examinaron las pérdidas por la circulación de la cadena por medio de la comparación de la entrada y salida de potencia a una tasa de pedaleo de 72 rev. min-1. Estos autores encontraron que el porcentaje de pérdida energética se incrementó con la producción de potencia desde 1.9% a 100 W hasta 3.9% a 300 W. De manera contraria, Martin et al. (1998) realizaron un modelo matemático de las pérdidas por la circulación de la cadena y obtuvieron un fijo de 2.4%. Martin y colaboradores basaron su modelo en hallar una diferencia consistente de esta magnitud entre dos ergómetros, uno de los cuales midió la producción de potencia antes de la circulación de la cadena y el otro después de la misma. Sin embargo, este método puedo no ser válido, ya que Jones y Passfield (1998) sugirieron que ambos de los ergómetros arriba mencionados ya incluían las pérdidas por circulación de la cadena en el cálculo de la producción de potencia. Es probable que las pérdidas por fricción en los rodamientos (e.g. en las masas de las ruedas) sean triviales. Por ejemplo, Martin et al. (1998) sugirieron que las pérdidas en los rodamientos de las ruedas alcanzaron cerca de 1.4 W (0.6 %) cuando se pedaleaba a 30 km.h-1.
Resistencia debido a la Gravedad
Durante el ciclismo cuesta arriba, las velocidades frecuentemente disminuyen a una extensión tal que la resistencia aerodinámica ya no es importante. En vez de esto, la fuerza resistiva más significativa es causada por tener que levantar el peso de la propia masa del corredor y de la bicicleta por la pendiente contra la gravedad. La influencia de la pendiente de la ruta sobre la velocidad es probablemente influenciada por el viento, casi linealmente (Martin et al., 1998; Figura 3). El trabajo hecho venciendo a la gravedad cuando se pedalea cuesta arriba puede ser calculado simplemente como el cambio en la energía potencial. Así el trabajo hecho es gobernado por la masa del corredor y la bicicleta, la gravedad y la altura ganada. La potencia de pedaleo pude ser calculada simplemente considerando la tasa de este cambio en la energía potencial. Bajo condiciones en ausencia de viento, estos principios pueden ser usados para calcular la producción de potencia mientras se pedalea en contra de la pendiente, donde la pendiente y la resistencia de rodamiento son conocidas (e.g. en una cintaergómetro inclinada) si se asume que la resistencia al rodamiento y las pérdidas por fricción son despreciables.
De lo antes establecido puede ser observado que la masa del corredor y la bicicleta influencian directamente la producción de potencia cuando se enfrenta una subida. Consecuentemente, los corredores especializados en la escalada de cuestas o montañas, y sus bicicletas (dentro del reglamento) deberían ser tan livianos como fuera posible para maximizar su índice de producción de potencia/peso corporal. Una baja inercia es también una función de menos masa. Una baja inercia va a ayudar al rendimiento en condiciones de carrera que se caracterizan por aceleraciones repetidas, como las carreras criterium, o las carreras Madison y por puntos en la pista. Kyle (1996) señaló que en tales condiciones la masa de los componentes que ruedan debería ser tan pequeña como sea práctico, ya que los efectos de la inercia son más pronunciados. Para las pruebas por tiempo en terreno llano, es improbable que la reducción de la masa del corredor o la bicicleta produzca mejoras significativas en el rendimiento. Cuando se pedalea en descenso, los corredores más pesados tienen una clara ventaja por sobre los corredores más livianos, como observó Swain (1997). Esto es porque los corredores más pequeños tienden a tener una mayor resistencia del aire relativa a su masa corporal.
Lineamientos Futuros
Han sido usados una variedad de métodos para estimar los requerimientos energéticos del ciclismo de ruta y de pista bajo diferentes fuerzas resistivas. Estos métodos incluyen la medición del gasto energético humano a partir del consumo de oxígeno (Swain et al., 1987; McCole et al., 1990), técnicas de agua doblemente marcada (Westerterp et al., 1986), estimación del gasto energético a partir de la frecuencia cardiaca (Palmer et al., 1994; Padilla et al., 2000) evaluaciones de deceleración (de Groot et al., 1995) y remolque de los ciclistas (di Prampero et al., 1979; Capella et al., 1993). Usando los datos de tales estudios, Olds et al. (1993, 1995) derivaron modelos matemáticos de ciclistas de ruta y pista. Estos enfoques han proporcionado ideas útiles con respecto a las demandas del ciclismo. Sin embargo, varios de los métodos usados para identificar la magnitud de los términos en los modelos matemáticos pueden ahora ser sustituidos por el desarrollo de un dinamómetro montado en la bicicleta (SRM, Julich, Alemania). Este dinamómetro que esta en la palanca de la bicicleta permite la medición directa de la potencia tanto en condiciones de laboratorio como de campo, y ha probado ser razonablemente exacto y confiable (Jones y Passfield, 1998).
Consecuente, recientes estudios han sido capaces de describir la producción de potencia en un rango de eventos ciclísticos a partir de mediciones tomadas en el campo (Broker et al., 1999; Craig y Norton, 2001; Jeukendrup et al., 2001) y a partir de modelos matemáticos (Martin et al., 1998; Bassett et al., 1999). Estos modelos deberían permitirles a los científicos y entrenadores evaluar de manera exacta las implicaciones teóricas sobre el rendimiento de los cambios en los factores ambientales, fisiológicos, nutricionales y tecnológicos. Por ejemplo, como se describe en la sección de “estrategia de Ritmo de Carrera”, tales modelos pueden ser útiles para explorar la influencia de la estrategia del ritmo de carrera sobre el rendimiento bajo varios contextos hipotéticos (Swain, 1997).
El diseño de bicicleta usado en los estudios que han generado modelos matemáticos ha mejorado a través de los años en términos de su relevancia para la competición. Por ejemplo, Martin et al. (1998) usaron una bicicleta de competición con el corredor en una posición de prueba por tiempo. No obstante, puede ser afirmado que todos los datos derivados a partir de estos modelos son relevantes solo para las disciplinas de ruta y pista. Aunque la intensidad del ejercicio durante el ciclismo de montaña ha sido estimada a partir de la frecuencia cardiaca durante las carreras (Impellizeri et al., 2002), pocos datos parecen estar disponibles sobre las características del ciclismo de montaña, un aspecto que las futuras investigaciones deberían investigar.
ESTRATEGIA DEL RITMO DE CARRERA
Aunque la estrategia del ritmo de carrera es reconocida como importante por los entrenadores y atletas, los estudios empíricos acerca de la óptima distribución de la potencia en una carrera son relativamente raros. Robinson et al. (1958) y Foster et al. (1993) reportaron que una distribución regular de trabajo es óptimo para el ejercicio de carrera de 1.2 km y para las pruebas de ciclismo de 2 km, respectivamente. El cuidadoso examen de los más recientes trabajos de investigación acerca de la estrategia de ritmo de carrera, junto con una apreciación acerca de cómo los factores externos esbozados en la sección anterior acerca de los “Factores Fisiológicos” influencian la velocidad de pedaleo, sugiere que una estrategia más complicada que una estrategia de ritmo de carrera regular o fijo sería mejor en algunas carreras.
Es difícil arribar a una conclusión simple para la estrategia de ritmo de carrera en las carreras de ruta de “salida masiva”, debido a la complejidad del evento. Consecuentemente, el foco actual se centra sobre eventos por tiempo. Los corredores de ruta de élite generalmente tratan de evitar variar el gasto energético hasta los 50 km finales de carrera, una táctica que tiene apoyo empírico (Palmer et al., 1996b). No obstante, muchas carreras profesionales han sido ganadas por corredores que trabajan duro en la parte inicial de la carrera para establecer un primer lugar inatacable. Además, las tácticas en las carreras de ruta dependen en una innumerable variedad de factores, incluyendo las condiciones ambientales, la cantidad de trabajo de equipo que es evidente, y el número y pendiente de las subidas en la carrera (Lucia et al., 1999b). Sería extremadamente difícil presentar completamente la influencia de todos estos factores, pero no imposible dados los casos suficientes para la potencia estadística adecuada en un modelo de regresión.
Pruebas por Tiempo en Pista
Las relaciones entre el esfuerzo fisiológico, la potencia y la velocidad son relativamente fuertes en los eventos de pista, ya que hay pocos cambios en las características físicas de los ambientes de carrera (e.g. cambios en la presión barométrica, temperatura, dirección del viento). Consecuentemente, como en el pedestrismo, los tiempos parciales dentro de las carreras de pista pueden ser usados como indicadores relativamente precisos de la estrategia del ritmo de carrera. El evento Olímpico por tiempo más largo en la pista es de 4 km. Sin embargo, los records mundiales son reconocidos para eventos individuales de una mayor duración (e.g. el “record de la hora”). Los resultados de Foster et al. (1993) sugieren que una estrategia de carrera uniforme de tiempos parciales muy similares es óptima para los eventos de 4 km de duración, este consejo es formulado a partir de una respuesta fisiológica óptima al ejercicio.
Las pruebas de tiempo sobre las distancias de 1 a 4 km son complicadas por el hecho de que el tiempo requerido para llevar la bicicleta hasta alcanzar la velocidad buscada constituye una proporción relativamente grande del tiempo total de carrera y también por la contribución relativa de la energía cinética al final de una carrera. Por ejemplo, los ciclistas de élite en la persecución de 4 km completan los primeros 250 m de la carrera en un tiempo que es aproximadamente 3-4 s más lento que las otras secciones de 250 m (Wilberg y Pratt, 1988). Van Ingen Shenau et al. (1994) y de Koning et al. (1999) simularon carreras de pista de 1 y 4 km, incorporando las características de la partida detenida, y encontraron que la mejor estrategia de ritmo de carrera es un esfuerzo máximo, aún si tal táctica conduce a una caída en la potencia hacia el final de la carrera. Van Ingen Schenau et al. (1994) sostuvieron que sería importante para un ciclista de velocidad de 1 km no “dejar atrás” una estrategia de esfuerzo máximo con el resto de la carrera en mente, ya que puede ser obtenida una considerable ventaja introduciendo más energía cinética en las etapas iniciales de la carrera. En efecto, el corredor estaría liberando más energía durante la primera parte de la carrera y esta energía conduciría a menores tiempos aún si la producción de potencia cayera considerablemente en las etapas posteriores de la carrera. Van Ingen Schenau et al. (1994) citaron el reciente trabajo de Hirvonen et al. (1987) para apoyar su hipótesis acerca de que tasas máximas de liberación de energía son importantes para esprintar. Hirvonen et al. (1987) encontraron que corredores de velocidad de élite tienen una mayor tasa de catabolismo de fosfocreatina en los primeros segundos de una carrera. Recientemente, Bishop et al. (2002) encontraron que la mejora en el rendimiento asociada con una partida máxima en el kayak de velocidad puede ser mediada por una cinética del VO2 más rápida. Trabajos de investigación más específicos al ciclismo son requeridos para dilucidar el balance exacto entre la optimización biofísica y la producción energética y fisiológica de tal energía. Craig y Norton (2001) sugirieron, como hicimos nosotros en la sección sobre “Factores Físicos”, que los recientes avances en la tecnología para la medición de la producción de potencia bajo condiciones competitivas podrían ser usados para monitorear las características de tales carreras de pista y la aceptabilidad fisiológica de las altas producciones de potencia de partida.
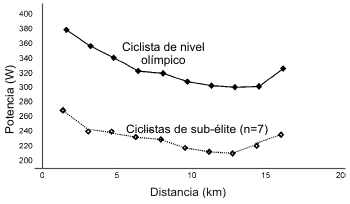
Figura 4. Perfil de potencia de un corredor olímpico de 4 km de
persecución durante una prueba simulada de 16.1 km. La potencia promedio a
través de la carrera simulada fue 327 W para el corredor (datos no publicados
obtenidos de uno de los autores, G.A.). Perfil de potencia promedio de una
muestra (n=7) de ciclistas de capacidad variable (Atkinson y Brunskill, 2000).
Todos los datos fueron recolectados usando un ergómetro (Computrainer Pro, Racer
Inc., Seattle, WA).
Pruebas por Tiempo en Ruta
Las competiciones por tiempo de ciclismo que son llevadas a cabo típicamente en rutas abiertas son mucho más largas (10-60 km en las etapas de carreras de élite) que aquellas llevadas a cabo en la pista. De este modo, la contribución de la partida detenida al tiempo total es mucho menos importante en las pruebas por tiempo en ruta. Además, un intento de maximizar el aumento de energía cinética con un esfuerzo de partida máximo conduciría a reducciones posteriores en la potencia que podrían ser desastrosas para el tiempo total de carrera. No obstante, hay evidencia acerca de que es muy difícil para un corredor no empezar una prueba por tiempo de cualquier distancia con una alta producción de potencia. En la Figura 4, presentamos el perfil de potencia de un ciclista olímpico de 4 km de persecución el cual completó una prueba por tiempo simulada de 16.1 km en un cicloergómetro (Computrainer Pro, Racer Mate Inc., Seattle, WA). En concordancia con Nikolopoulos et al. (2001), puede ser visto que la potencia inicial fue mucho más alta que para el resto de la carrera. Este perfil es característico de la mayoría de los ciclistas (e.g. la muestra estudiada usando el mismo cicloergómetro de arriba también presentada en la Figura 4) y no es necesariamente acompañado por un inminente incremento en el agotamiento percibido en comparación con una salida de menor potencia (Firth, 1998; Atkinson y Brunskill, 2000). Si los ciclistas son entrenados para mantener el esfuerzo inicial, usando una retroalimentación de la potencia, el tiempo de carrera total puede ser usualmente mejorado (Firth, 1998; Atkinson y Brunskill, 2000), sumando peso al argumento acerca de que la estrategia de ritmo de carrera óptima es aquella que varía poco con respecto a la potencia promedio a través de toda la carrera. Si no está disponible un instrumento para la medición de la potencia en la bicicleta, un buen consejo del entrenador sería elegir un desarrollo que sea ligeramente más liviano que el que sería normalmente adoptado al inicio para alentar una menor potencia en los primero minutos de una prueba por tiempo larga. La monitorización de la frecuencia cardiaca o las tasas de fatiga percibida del corredor parecen ser métodos poco confiables para detectar diferencias en la potencia de salida (Atkinson y Brunskill, 2000).
La información anterior podría sugerir que mantener la potencia relativamente constante a través de la duración de una carrera es la mejor estrategia para todas las pruebas por tiempo de ruta. Sin embargo, la identificación de una estrategia de ritmo de carrera óptima es complicada por condiciones externas variables dentro de la carrera, como la pendiente y la dirección del viento. Es extremadamente improbable que alguna prueba de ruta por tiempo sea llevada a cabo en un recorrido completamente llano sin viento, las condiciones simuladas en la mayoría de las pruebas por tiempo de laboratorio (Hickey et al., 1992; Palmer et al., 1996a). Un ciclista característico que genera una potencia constante de 255 W viajará a 6 m.s-1 en un viento en contra de 10 m.s-1 en comparación con 18 m.s-1 con un viento de cola de 10 m.s-1. A la misma tasa de trabajo la velocidad del ciclista sería de 5 m.s-1 mientras se pedalea cuesta arriba con una pendiente de 6 % en comparación con 18 m.s-1, mientras se pedalea cuesta abajo (Figura 3, Martin et al., 1998). Los efectos positivos del viento de cola y las subidas sobre la velocidad de carrera no cancelan completamente los efectos deletéreos del viento en contra y las subidas sobre la velocidad de carrera. De este modo, las carreras en circuito o ruta en terrenos montañosos o en condiciones ventosas van a ser siempre más lentas que las carreras llanas y en ambientes calmos (Kyle, 1988; White, 1994; Martin et al., 1998). Tal como la partida detenida en una carrera de pista, las secciones relativamente lentas de una carrera (viento en contra, subidas) representan la mayor proporción del tiempo total de carrera. En consecuencia, una estrategia exitosa puede ser focalizarse en mejorar la velocidad en estas secciones lentas.
En una simulación, Swain (1997) usó la ecuación de movimiento de un ciclista proporcionada por Di Prampero et al. (1979) para predecir que, cuando una carrera es mantenida en un recorrido montañoso o en un recorrido con secciones iguales de viento en contra y viento de cola, los tiempos más rápidos serían registrados si el ciclista incrementa la potencia en las secciones de subida y viento en contra y compensa con una reducción de la potencia en las secciones descenso y viento a favor (el trabajo total o “neto” realizado seguiría siendo el mismo). Por ejemplo, Swain (1997) predijo que un corredor que varía la producción de potencia en un 10 % en condiciones variables de un fuerte viento (24 km. h-1) podrían ser más de un minuto más rápido a través de 40 km que un corredor que elige una estrategia de ritmo de carrera consistente a la misma producción de potencia promedio. Hasta aquí, no ha habido ningún intento general de replicar el trabajo de investigación de Swain (1997) con el modelo matemático aportado por Martin et al. (1998). Usando los datos aportados para el ciclista en el Apéndice I de Martin et al. (1998), nosotros simulamos los efectos de variar la potencia en paralelo con variaciones en el viento y la pendiente a través de una “carrera” de 50 km. Encontramos que si el ciclista produce una potencia de 400 W en una carrera de 50 km sin montañas ni viento, el tiempo para la finalización sería de 64 min 12 s. Si el mismo ciclista produce la misma potencia, pero contra un viento en contra de 10 m. s-1 por los primeros 25 km y con un viento de cola de la misma velocidad por los 25 km restantes, entonces el tiempo del corredor sería de 76 min 10 s. No obstante, si el corredor es capaz de incrementar la producción de potencia en 20 W (5 %) en la sección de viento en contra y disminuir la potencia en la sección de viento de cola de modo que la potencia promedio a través de toda la carrera se todavía 400 W, entonces el tiempo de 50 km sería de 75 min 7 s, un ahorro de 1 min en comparación con una estrategia de potencia constante.
Atkinson y Brunskill (2000) confirmaron tal hipótesis de “ahorro de tiempo” cuando examinaron la influencia de una estrategia de ritmo de carrera constante versus variable durante una prueba por tiempo de 16.1 km realizada en un ergómetro (Computrainer Pro, Racer Mate Inc., Seattle, WA) que podía simular los cambios en la dirección del viento en una carrera. El tiempo de carrera fue ligeramente menor y el agotamiento percibido total fue significativamente menor cuando la producción de potencia varió en paralelo con la fuerza resistiva debido al viento simulado en comparación con una estrategia de ritmo de carrera más constante.
Swain (1997) predijo que, en condiciones ventosas o montañosas, mientras más puede variar la potencia un corredor, más tiempo es ahorrado durante una carrera. Tal predicción dependería de la distancia recorrida en cada sección de viento en contra y, por lo tanto, de la distancia de la carrera y la naturaleza del curso de la misma. En la sección sobre “Factores Fisiológicos”, nosotros destacamos que los ciclistas de élite pueden pedalear a aproximadamente 90 % del máximo consumo de oxígeno por 1 h, y así sería imposible variar el consumo de oxígeno en más de un 15 %. Liedl et al. (1999) estimaron recientemente una variación aceptable de la potencia en una prueba por tiempo simulada en el laboratorio. La potencia media (correspondiente al 78 % del VO2 máx.) a partir de una prueba por tiempo inicial de 1 h fue usada para examinar el esfuerzo fisiológico y subjetivo de dos estrategias de ritmo de carrera: (1) ritmo constante a la tasa de trabajo media y (2) ritmo variable que consistió de períodos alternados de 5 min de pedaleo a un 5 % arriba y un 5 % debajo de la tasa de trabajo media. No fueron encontradas diferencias en el esfuerzo fisiológico entre las dos estrategias de ritmo de carrera. Atkinson y Brunskill (2000) también encontraron que los corredores fueron capaces de variar la potencia en un 5 % durante una prueba por tiempo simulada de 16.1 km. Tal como la examinación de la estrategia del ritmo de carrera en los eventos de velocidad, todavía está poco claro cuales son los cambios exactos entre la optimización fisiológica de la producción de potencia y la optimización biofísica de la velocidad de pedaleo.
Lineamientos Futuros
Ya que la magnitud relativa de la potencia de salida puede ser particularmente importante para las pruebas por tiempo, tanto en la pista como en la ruta, las futuras investigaciones deberían intentar identificar la forma más apropiada de monitorear la potencia de salida.
De manera ideal, el corredor debería ser entrenado para identificar diferentes potencias de salida y debería ser examinada la influencia de estas potencias sobre el tiempo total de carrera. Es necesaria más información acerca de cuales medidas fisiológicas o subjetivas son sensibles a los cambios sutiles en el ritmo de carrera.
En la actualidad, las ideas de Swain (1997) no han sido confirmadas usando los más recientes y posiblemente más exactos modelos matemáticos (e.g. Martin et al., 1998). Tal trabajo debería estimular varias condiciones de carrera de montaña y viento que pueden ser enfrentadas por los corredores. Las futuras investigaciones deberían establecer si la optimización de los atletas de la distribución de potencia en línea con las variaciones en el ambiente de carrera hace una diferencia al tiempo total de carrera. Un componente importante de tales investigaciones debería ser la identificación de cuanta potencia puede ser variada sin un incremento en el esfuerzo fisiológico o subjetivo total en comparación con una estrategia de potencia constante.
ESTRATEGIAS NUTRICIONALES
Las producciones de potencia observadas de manera característica durante las competiciones de ciclismo amateur y profesional (ver Figura 2) están asociadas con un gasto energético muy alto. Puede ser estimado que un ciclista, con una eficiencia y aerodinámica típicas, gasta aproximadamente 83 kJ. min-1, cuando pedalea a 40 km. h-1 (Jeukendrup et al., 2001). Los ciclistas profesionales pueden gastar hasta 120 kJ. min-1 para períodos de tiempo más largos, con la mayoría de esta energía derivada a partir de la ruptura del glucógeno muscular. Durante el Tour de Francia, los gastos energéticos pueden llegar a 36 MJ. día-1 (Saris et al., 1989). En el Tour de Francia los corredores corren un alto riesgo de sufrir una depleción de glucógeno muscular si los costos energéticos no son restituidos con la nutrición adecuada (Jeukendrup et al., 2001). Puede ser producida una cantidad significativa de calor como un derivado del trabajo muscular implicado en el ciclismo, particularmente en temperaturas correspondientes a un ambiente cálido. La transpiración es un mecanismo efectivo para remover algo de este calor del cuerpo, con el riesgo de perjudicar el rendimiento, a través de la deshidratación, dependiendo de lo adecuado de la ingesta de fluidos. Las estrategias nutricionales diseñadas para aminorar los problemas arriba mencionados son complicadas, pero pueden ser descritas en términos de tiempo, antes, durante o después de una carrera.
La Semana antes de la Carrera
Los clásicos estudios de Bergstrom et al. (1967) y Hultman (1967) demostraron que el rendimiento de resistencia disminuye cuando una dieta alta en carbohidratos es adoptada en comparación con una dieta normal mixta en la semana anterior a la competición. Parece haber una relación lineal entre la concentración de glucógeno muscular y el tiempo hasta la fatiga a una potencia constante. Aunque el protocolo original de “carga de glucógeno” incluía a la comúnmente denominada “fase de depleción”, estudios más recientes indicaron que esta fase no es necesaria en atletas entrenados (Sherman et al., 1981). En estos primeros estudios, fue investigado el tiempo hasta el agotamiento en vez del rendimiento en una prueba de tiempo. La relevancia de las mediciones de tiempo hasta el agotamiento para el rendimiento en carreras reales ha sido cuestionada, esto es, no existen carreras en las cuales los corredores tengan que pedalear durante tanto tiempo como sea posible (Jeukendrup et al., 1996). De este modo, son requeridos más estudios para evaluar las variables de rendimiento externamente válidas en vez de la capacidad de resistencia general.
Los efectos del glucógeno muscular elevado sobre el rendimiento en pruebas por tiempo (i.e., para cubrir una cierta distancia tan rápido como sea posible) han sido investigados por muchos grupos de investigación y, distinto de los efectos de la carga de carbohidratos sobre la capacidad de resistencia (tiempo hasta el agotamiento), los efectos sobre el rendimiento en las pruebas por tiempo parecen ser menos esclarecidos. Los investigadores han encontrado generalmente pequeños efectos de la carga de carbohidratos sobre el rendimiento de las pruebas por tiempo que duran 1 h o menos. En una excelente y completa revisión, Hawley et al. (1997) concluyeron que el rendimiento en las pruebas por tiempo puede ser incrementado en un 2-3 % por la carga de carbohidratos. Sin embargo, es importante señalar que en la mayoría de los estudios la concentración de glucógeno en la condición de control era bastante baja y es probable que no puedan ser observadas diferencias en el rendimiento de las pruebas por tiempo cuando la ingestión de carbohidratos es de 7 g. kg-1.día-1 o más. De manera interesante, Coyle et al. (2001) demostraron recientemente que ciclistas que se ejercitaron diariamente (2 h al 65 % del VO2 máx.) por 7 días fueron capaces de mantener y aún incrementar sus reservas de glucógeno a través de toda una semana. Cuando recibieron una dieta que contenía 11 g. kg-1.día-1 en comparación con una dieta que contenía 7 g. kg-1.día-1, sus concentraciones de glucógeno muscular estuvieron elevadas. De hecho, las concentraciones de glucógeno muscular en aquel estudio están entre las más altas reportadas alguna vez (Coyle et al., 2001). A pesar de las reservas de glucógeno más altas en las dietas altas en carbohidratos, el rendimiento (como fue medido por medio de una prueba de tiempo después de 2 h de ciclismo en estado estable) no fue afectado. De este modo, fue recientemente concluido que la ingesta de 7-10 g. kg-1.día-1 de carbohidratos debería ser suficiente para repletar completamente las reservas de carbohidratos y permitir un rendimiento óptimo (Jentjens y Jeukendrup, 2003). Para una discusión más destacada acerca del rol del glucógeno muscular para el rendimiento en ejercicio, el lector es remitido a las recientes revisiones de Hawley et al. (1997) y Jentjens y Jeukendrup (2003).
3 a 4 Horas antes de la Carrera
Ha sido demostrado que la ingestión de una comida rica en carbohidratos que contenga 140-330 g de carbohidratos, 3-4 h antes del ejercicio, incrementa el glucógeno muscular (Coyle et al., 1985) y aumenta el rendimiento del ejercicio. Después de una noche de ayuno, el glucógeno hepático puede estar sustancialmente reducido (Nilsson y Hultman, 1973), lo cual puede resultar en dificultades para mantener las concentraciones de glucosa sanguínea durante la serie de ejercicio subsiguiente. La ingestión de carbohidratos puede repletar las reservas de glucógeno hepáticas y contribuir, junto con cualquier absorción en curso de los carbohidratos ingeridos, a mantener la glucosa sanguínea.
La ingesta de carbohidratos en pocas horas antes del ejercicio tiene profundos efectos sobre el metabolismo. Generalmente, el uso de carbohidratos es estimulado y la movilización y oxidación de ácidos grasos es disminuida. Estos cambios metabólicos pueden persistir por hasta 6 h después de la ingestión de carbohidratos (Montain et al., 1991). No obstante, los cambios no parecen ser perjudiciales para con el rendimiento de ejercicio, debido a que un incremento en la disponibilidad de carbohidratos compensa la mayor utilización de los mismos.
Una Hora antes de la Carrera
La ingestión de carbohidratos en reposo resulta en un gran incremento en la glucosa e insulina plasmáticas. Sin embargo, cuando el ejercicio es iniciado 30-60 min después de la ingestión de carbohidratos, y aún si se continúa ingiriendo carbohidratos, hay una caída rápida en la glucosa plasmática, como consecuencia de la combinación de los efectos estimulantes de las altas concentraciones de insulina y de la contracción muscular, sobre la captación muscular de glucosa.
Ya que los efectos metabólicos de la ingestión de carbohidratos preejercicio son consecuencia de altas concentraciones de glucosa e insulina, ha habido interés en estrategias que minimicen los cambios en la glucosa plasmática y la insulina antes del ejercicio. Estas estrategias han incluido la ingesta de otros tipos de carbohidratos (e.g. fructosa) distintos de la glucosa, variando la carga de carbohidratos y la programación en el tiempo de la ingestión, adicionando grasa, o incluyendo ejercicio de entrada en calor en el período preejrcicio. En general, mientras estas distintas intervenciones modifican la respuesta metabólica al ejercicio, parece no haber ninguna ventaja para el rendimiento de ejercicio en el aumento de las concentraciones de glucosa e insulina preejercicio (Hargreaves et al., 1987; Okano et al., 1988; Calles-Escandon et al., 1991; Goodpaster et al., 1996). No obstante, puede haber muchos individuos que sufren más que otros de las consecuencias de una glucosa sanguínea baja (hipoglucemia). Cuando se ingiere carbohidratos durante el ejercicio prolongado, el tipo de carbohidratos preejercicio ingeridos no tiene ningún efecto sobre el metabolismo y el rendimiento (Burke et al., 1998).
Puede ser sostenido que si la ingestión de carbohidratos preejercicio es el único mecanismo por el cual un ciclista puede incrementar la disponibilidad de carbohidratos durante el ejercicio, el ciclista podría ser bien aconsejado acerca de ingerir tantos carbohidratos como sea posible, sin experimentar estrés gastrointestinal. Esta táctica compensaría el uso reducido de grasa y proporcionaría un pool de glucosa que se vuelve disponible para el uso en las etapas posteriores del ejercicio. Parece haber poca evidencia que apoye la práctica de evitar la ingesta de carbohidratos en la hora anterior al ejercicio. Los procedimientos individuales deben ser determinados en base a la experiencia personal con varios protocolos de ingestión de carbohidratos prejercicio.
Además de la depleción de carbohidratos, la deshidratación es uno de los principales factores que contribuyen a la fatiga en los ambientes calurosos, con pérdidas tan pequeñas como el equivalente al 1-2 % de la masa corporal (600-1500 ml), asociadas con un rendimiento de ejercicio perjudicado (Walsh et al., 1994). Así, mientras se preparan para carreras de un día, los ciclistas deberían ingerir suficientes cantidades de fluidos para asegurar que ellos esten bien hidratados antes del ejercicio. Esto es mejor monitoreado por mediciones de la masa corporal. La ingesta de grandes cantidades de agua puede resultar en una hiperhidratación antes del ejercicio, pero es también probable que incremente la diuresis (fluido excretado) y el estrés gastrointestinal (Maughan y Nadel, 2000). Ha habido cierto interés en adicionar glicerol a las bebidas de hidratación para mejorar la retención y mantener una relativa hiperhidratación antes y durante el ejercicio. Los estudios reportados en la literatura han producido resultados conflictivos (Latzka et al., 1998; Hitchins et al., 1999), y está la posibilidad acerca de que el glicerol puede promover la deshidratación intracelular con consecuencias potencialmente negativas tales como dolor de cabeza. Es quizás prematuro en esta etapa recomendar glicerol para lograr una hiperhidratación.
Durante la Carrera
Durante el ciclismo prolongado, la energía debe ser proporcionada en forma de carbohidratos y las pérdidas de fluidos necesitan ser repletadas para prevenir la deshidratación. Anteriores investigaciones demostraron que los ciclistas pueden pedalear un 20-25 % más en carreras hasta el agotamiento al 65-70 del VO2 máx. (Coyle et al., 1986). Trabajos más recientes con protocolos externamente válidos han mostrado que el rendimiento en una prueba simulada de 40 km mejoró en más de 1 min con la ingestión de carbohidratos en comparación con un placebo con agua (Jeukendrup et al., 1997). Consecuentemente, generalmente se cree que la ingestión de carbohidratos durante el ejercicio más largo que aproximadamente 45 min puede mejorar el rendimiento de ejercicio. Durante el ejercicio de 90 min o más largo, cualquier mejora en el rendimiento debido a intervenciones dietarias es probable que sea el resultado de un mejor mantenimiento de la concentración de la glucosa sanguínea y tasas más altas de utilización de carbohidratos. Durante el ejercicio de alta intensidad, tal como las pruebas por tiempo de 40 km, los mecanismos están menos claros, ya que solo pequeñas cantidades de los carbohidratos ingeridos se van a volver disponibles para los músculos que trabajan. Ha sido sugerido que la alimentación durante el ejercicio puede tener efectos centrales (Jeukendrup et al., 1997).
Los carbohidratos ingeridos durante el ejercicio van a reducir el catabolismo de glucógeno hepático (Jeukendrup et al., 1999a, b). Generalmente se piensa que es beneficiosa una mayor contribución de los carbohidratos ingeridos al gasto energético, en parte debido a que tal contribución va a reducir el catabolismo de las fuentes endógenas de carbohidratos (i.e. glucógeno hepático). Ha sido demostrado que ciertos tipos de carbohidratos son oxidados más rápidamente que otros (Jeukendrup y Jentjens, 2000). La glucosa, maltodextrinas, sucrosa, maltosa y féculas solubles pueden ser oxidadas a tasas relativamente altas, mientras que la fructosa y la galactosa son oxidadas a tasas más lentas.
Mientras más carbohidratos se ingieran, más van a ser oxidados. Sin embargo, cuando los carbohidratos son ingeridos a una tasa de aproximadamente 1.0-1.2 g. min-1, la ingesta de carbohidratos adicionales no va a resultar en un incremento en la utilización de los mismos (Jeukendrup y Jentjens, 2000). Aún la ingestión de cantidades muy grandes de carbohidratos no va a incrementar el uso de aproximadamente 1 g. min-1.
La mayoría de los ciclistas no toman suficientes fluidos en las carreras. Las mediciones durante varias carreras de ciclistas profesionales (Tour de Andalucia, Tour del Mediterráneo, Tour de Suiza, Tour de Francia) han demostrado que los corredores pierden 2.1-4.5 kg de masa corporal después de una etapa típica. Parte de esta pérdida en la masa corporal (100-300 g) puede ser carbohidratos y grasas usadas durante el ejercicio, pero la mayor parte va a ser pérdida de fluidos. En algunas partes de la carrera, es muy difícil beber, simplemente debido a que se necesita tener las manos en el manubrio o porque hay muy poco tiempo para hacerlo. Esto es especialmente así es las partes de ascenso y descenso abrupto y cuando los corredores están atacando. Durante el ciclismo de montaña, beber puede ser aún más difícil en las partes técnicas. Otro problema es el hecho que las botellas para beber no están siempre fácilmente disponibles y un máximo de cerca de 1 litro puede ser llevado en la bicicleta.
Después de la Carrera
Después del ejercicio, y particularmente en las carreras por etapas, es importante restituir las pérdidas de fluidos y el glucógeno muscular. Dependiendo de la temperatura ambiental, la restitución de fluidos puede ser más importante, menos importante o igual de importante que la repleción del glucógeno muscular. En la mayoría de los casos, los corredores pretenderán alcanzar ambos objetivos al mismo tiempo.
La efectividad de la rehidratación postejercicio es principalmente determinada por el volumen y composición de los fluidos consumidos. El agua no es la bebida de rehidratación ideal postejercicio cuando es necesario un reestablecimiento rápido y completo del balance corporal de fluidos (Maughan y Nadel, 2000). La ingestión de agua sola en el período postejercicio resulta en una rápida caída en la concentración del sodio plasmático y una caída en la osmolalidad del plasma.
Estos cambios tienen el efecto de reducir la estimulación a beber (sed) e incrementar la salida de orina, y ambos factores van a perjudicar el proceso de rehidratación. El volumen plasmático es más rápidamente y completamente reestablecido en el período postejercicio si algo de cloruro de sodio (77 mmol. l-1) es adicionado al consumo de agua (Nose et al., 1998). Esta concentración es similar al límite superior de la concentración de sodio encontrada en el sudor, pero es consistentemente más alta que la concentración de sodio de la mayoría de las bebidas deportivas comercialmente disponibles, las cuales usualmente contienen 10-25 mmol. l-1.
La completa rehidratación después del ejercicio puede solo ser alcanzada si el sodio perdido en la transpiración es restituido, así como el agua. Para alcanzar una euhidratación, la ingesta de sodio debe ser superior a la pérdida del mismo. La ingestión de una bebida que contenga sodio no solo va a promover una rápida absorción de fluidos en el intestino delgado, sino que permitirá que la concentración de sodio plasmático se mantenga elevada durante el período de rehidratación y ayuda a mantener la sed mientras que retrasa la estimulación de la producción de orina. Se piensa que la inclusión de potasio en la bebida consumida después del ejercicio mejora la restitución de agua intracelular y así promueve la rehidratación, pero actualmente hay poca evidencia experimental para apoyar esto. La bebida de rehidratación debería tener también carbohidratos (glucosa o polímeros de glucosa), debido a que la presencia de glucosa va también a estimular la absorción de fluidos en el intestino y mejorar el sabor de la bebida. Después del ejercicio, la captación de glucosa dentro del músculo para la resíntesis de glucógeno debería también promover la rehidratación intracelular.
Hasta hace poco tiempo, era generalmente recomendado que los atletas deberían consumir un volumen de fluidos equivalente a la pérdida de fluidos ocurrida durante el ejercicio para rehidratarse adecuadamente en el período de recuperación postejercicio. Fue recomendado que los atletas consumieran aproximadamente 1 litro de fluido por cada kilogramo perdido durante la sesión de ejercicio. Actualmente está claro que esta cantidad de fluido es insuficiente, debido a que la misma no tiene en cuenta las pérdidas por la orina que ocurren después del consumo de bebida, a través de un período de horas. Los datos existentes indican que la ingestión de 150 % o más de la pérdida de masa corporal (i.e 1.5 litros de fluido consumidos durante la recuperación por cada kilogramo de masa corporal perdido durante el ejercicio) puede ser requerido para alcanzar la hidratación normal dentro de las 6 h después del ejercicio (Maughan y Shirreffs, 1996).
Los ciclistas deberían ser alentados a consumir comida sólida así como fluidos dentro de una serie de ejercicio, a menos que la ingestión de comida tenga probabilidad de resultar en molestias gastrointestinales. En un estudio, el mismo volumen de fluidos consumido como una combinación de comida más agua en comparación con una bebida deportiva resultó en la producción de un menor volumen de orina y por lo tanto en una mayor retención de fluidos (Maughan y Shirreffs, 1996). La mayor eficacia del tratamiento de comida más agua fue probablemente una consecuencia de su mayor contenido total de sodio y potasio. Bajo condiciones de ejercicio donde las pérdidas de sudor son grandes, la cantidad total de cloruro y sodio perdidos va a ser alta. Por ejemplo, la pérdida de 10 litros de sudor, con una concentración de 50 mmol.l-1, alcanza a una pérdida de aproximadamente 29 g de cloruro de sodio. Claramente, la ingesta de comida va a ser importante en la restitución de estas pérdidas de sal, ya que como fue antes mencionado las bebidas deportivas más disponibles comercialmente contienen solo cerca de 10-25 mmol.l-1.
La Figura 5 ilustra las máximas tasa de síntesis de glucógeno muscular reportadas como respuesta a diferentes tasas de ingesta de carbohidratos en las primeras horas postejercicio. Puede ser observado que la máxima tasa de resíntesis de glucógeno muscular es alcanzada a una ingesta de carbohidratos de entre 1.2 y 1.4 g. min-1 (75-90 g de carbohidratos por hora). Una ingesta de más de 90 g por hora no provee ningún beneficio adicional en términos de almacenamiento de glucógeno muscular y puede solo incrementar el riesgo de disconfort gastrointestinal. El tiempo en el que se realiza la ingesta de carbohidratos es también importante cuando es requerida una recuperación rápida. La completa repleción del glucógeno puede ser lograda dentro de 24 h siempre que la cantidad y el tiempo en el que se realiza la ingesta sean correctos (Jentjens y Jeukendrup, 2003). El más rápido reestablecimiento de las reservas de glucógeno ocurre en los primeros 60-90 min después del ejercicio. En un estudio, fue encontrado que la tasa de almacenamiento de glucógeno fue casi el doble de rápida si los suplementos de carbohidratos eran proporcionados después del ejercicio en vez de varias horas después del mismo (Ivy et al., 1988).
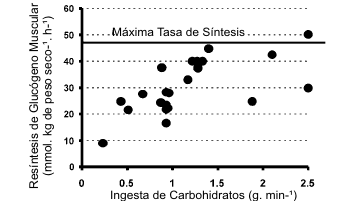
Figura 5. Máximas tasas de síntesis de glucógeno después de la ingestión
de diferentes cantidades de carbohidratos en las primeras horas después del
ejercicio.
La más altas tasas de síntesis de glucógeno muscular han sido reportadas en estudios en los cuales los carbohidratos fueron proporcionados en intervalos regulares (cada 15-30 min) (Jentjens y Jeukendrup, 2003). Esto es más probablemente debido al mantenimiento de altas concentraciones de glucosa sanguínea e insulina. Así, comer comidas pequeñas frecuentemente parece tener un beneficio extra sobre comer menos veces más comida. Adicionalmente, comer menos comida reduce probablemente el riesgo de disconfort gastrointestinal.
La mayoría de las formas de carbohidratos van a proporcionar cantidades similares de almacenamiento de glucosa o glucógeno. Cuando es ingerida fructosa, las tasas de síntesis de glucógeno son relativamente bajas debido a que la fructosa debe ser convertida a glucosa en el hígado antes de que pueda ser usada para la síntesis de glucógeno en el músculo. La mayoría de los tipos de carbohidratos, sin embargo, van a resultar en similares tasas de resíntesis de glucógeno muscular (glucosa, sucrosa, polímeros de glucosa).
Parece que no hay diferencias en la síntesis de glucógeno muscular cuando los carbohidratos son consumidos en forma líquida o sólida en las primeras horas después del ejercicio (Keizer et al., 1987). Sin embargo, en el estudio de Keizer et al., la cantidad total de carbohidratos ingeridos fue pequeña y las tasas de resíntesis de glucógeno fueron subóptimas. Las formas líquidas de carbohidratos o las comidas con carbohidratos con un alto contenido de fluido son frecuentemente recomendadas a los atletas debido a que las mismas son fáciles de digerir y dan menos saciedad, y de este modo, no tienden a afectar el apetito de la persona de manera adversa. Los suplementos de carbohidratos líquidos también proveen una fuente de fluido que puede ser beneficiosa para una rápida rehidratación.
La insulina promueve las síntesis del glucógeno muscular después del ejercicio y cualquier medio por el cual las concentraciones de insulina puedan ser elevadas podría ayudar a la resíntesis de glucógeno después del ejercicio. Algunos aminoácidos y proteínas van a causar la liberación de insulina desde el páncreas y varios estudios han mostrado que la co-ingestión de proteínas y/o aminoácidos con los carbohidratos casi dobla la respuesta de la insulina (Zawadski et al., 1992; Van Loon et al., 2000; Jentjens et al., 2001). La síntesis de glucógeno muscular fue también incrementada en estos estudios en un 40-100 % (Zawakzki et al., 1992; Van Loon et al., 2000). Sin embargo, un estudio reciente (Jentjens et al., 2001) ha mostrado que cuando la ingesta total de carbohidratos es muy alta (1.2 g. kg-1 de masa corporal por hora), la presencia de una mezcla de proteínas-aminoácidos no aumenta la tasa de síntesis de glucógeno muscular a pesar de que ocurre una respuesta de la insulina mucho más alta. De este modo, no hay ninguna necesidad de ingerir proteínas o aminoácidos con los carbohidratos para maximizar la síntesis de glucógeno cuando la ingesta total de carbohidratos es suficiente.
Lineamientos Futuros
Dado el predominio de carreras por etapas en el ciclismo y la frecuencia general de las carreras descritas en la sección sobre “Factores Fisiológicos”, las futuras investigaciones sobre nutrición podrían enfatizar la influencia de la dieta sobre la recuperación luego del ejercicio. Deberían ser exploradas nuevas estrategias para mejorar la resíntesis de glucógeno y para reducir el catabolismo de proteínas después de series repetidas de ejercicio.
Aunque la ingesta de proteínas no siempre tiene un efecto sobre la síntesis de glucógeno muscular, hay cierta evidencia acerca de que la ingesta de aminoácidos esenciales en las horas después del ejercicio puede incrementar la síntesis de proteínas (Rasmussen et al., 2000). La síntesis proteica incrementada después del ejercicio podría ayudar a reparar el daño muscular y a la síntesis de varias enzimas y de las mitocondrias. Hasta la fecha, los trabajos de investigación en el área anteriormente mencionada han sido limitados, debido a que las técnicas para estudiar el metabolismo proteico son complicadas y muy invasivas. Además, deberían ser hechos refinamientos a través de los resultados de investigaciones empíricas para optimizar el aporte de fluidos y carbohidratos a partir de las bebidas deportivas, especialmente bajo diferentes condiciones de esfuerzo en el calor sobre la termorregulación.
CONCLUSION
En esta revisión hemos intentado hacer un enfoque “holístico” para analizar el rendimiento en ciclismo. Los factores que influencian la potencia y la velocidad de pedaleo han sido considerados con una apreciación en tanto la naturaleza de diferentes disciplinas de ciclismo y las interrelaciones entre los diferentes factores mediadores.
Las características de los ciclistas que son comunes entre la mayoría de las disciplinas de ciclismo incluyen una alta producción de potencia aeróbica máxima y una capacidad para trabajar a producciones de potencia relativamente altas por extensos períodos de tiempo. La relación entre estas características y los factores fisiológicos inherentes tales como la capilarización muscular y el tipo de fibras musculares es complicada por las diferencias interindividuales en la selección de la cadencia en diferentes condiciones de carrera. Las demandas fisiológicas de muchas disciplinas ciclísticas tienen que ser todavía completamente investigadas.
Recientes avances en los modelos matemáticos acerca de las fuerzas resistivas sobre la bicicleta están empezando a ayudar a revelar las interrelaciones entre estas fuerzas resistivas. Los datos de estos modelos pueden ser suplementados con información derivada del uso de galgas de extensión montadas en las bicicletas, las cuales pueden medir potencia directamente en las carreras. Las carreras con bicicleta de montaña constituyen una disciplina descuidada en términos de caracterización de las producciones de potencia en condiciones de carrera y del modelamiento de los efectos de componentes de la bicicleta muy diferentes y las fuerzas resistivas asociadas.
La estrategia del ritmo de carrera durante el ciclismo depende de la magnitud relativa de determinadas fuerzas resistivas durante una carrera. Una distribución constante de la producción de potencia, con una potencia de salida “no máxima”, es mejor para pruebas por tiempo largas (> 4 km) mantenidas en condiciones invariables de viento y pendiente. Para carreras de pista más cortas, se aconseja una salida más potente para “ahorrar” tiempo durante la fase que contribuye más al tiempo total de carrera y para optimizar la contribución de energía cinética al tiempo total de carrera. La estrategia de ritmo de carrera óptima para las pruebas por tiempo en ruta puede necesitar tener en cuenta las condiciones variables de pendiente y viento, pero es necesaria más investigación para dilucidar completamente en que medida una producción de potencia variable puede ser tolerada por un corredor y cuanto tiempo puede podría ser ahorrado mediante tal estrategia de ritmo de carrera en una carrera real o una simulada con los últimos modelos matemáticos que estén disponibles.
La dieta es un aspecto multifactorial y muchos investigadores han tratado de examinar los aspectos de la nutrición en el ciclismo (e.g. organización en el tiempo, cantidad, composición) de manera aislada. Solo recientemente los investigadores intentaron analizar las interrelaciones entre los factores dietarios (e.g. la comunicación entre los efectos de la dieta precarrera e intracarrera sobre el rendimiento). El ambiente cálido es un factor mediador en la elección de la dieta, ya que puede haber intereses en la competición de reestablecimiento de la pérdida de fluidos y del glucógeno repletado durante y después de una carrera.
Los futuros lineamientos para la investigación deberían centrarse en cerrar más “circuitos” entre los diferentes factores que influencian el rendimiento en el ciclismo. Los científicos que investigan en las ciencias del ciclismo deberían tener una buena apreciación de las limitaciones de sus trabajos en términos de para que corredores, para que ambientes de carrera y para que disciplina competitiva del ciclismo son relevantes sus resultados.
Referencias
1. Ahlquist, L.E., Bassett, D.R., Su.t, R., Nagle, F.J. and Thomas, D.P (1992). The effect of pedaling frequency on glycogen depletion rates in type I and type II quadriceps muscle .bers during submaximal cycling exercise. European Journal of Applied Physiology, 65, 360364
2. Atkinson, G. and Brunskill, A (2000). Pacing strategies during a cycling time trial with simulated headwinds and tailwind. Ergonomics, 43, 14491460
3. Balmer, J., Davison, R.C. and Bird, S.R (2000). Peak power predicts performance power during an outdoor 16.1-km cycling time trial. Medicine and Science in Sports and Exercise, 32, 14851490
4. Bassett, J.R., Kyle, C.R., Pass.eld, L., Broker, J.P. and Burke, E.R (1999). Comparing cycling world hour records, 19671996: modelling with empirical data. Medicine and Science in Sports and Exercise, 31, 16651676
5. Bastiaans, J.J., van Diemen, A.B., Veneberg, T. and Jeukendrup, A.E (2001). The effects of replacing a portion of endurance training by explosive strength training on performance in trained cyclists. European Journal of Applied Physiology, 86, 7984
6. Bergstrom, J., Hermansen, L., Hultman, E. and Saltin, B (1967). Diet, muscle glycogen and physical performance. Physiologica Scandinavica, 71, 140150
7. Billat, V.L (1996). Use of blood lactate measurements for prediction of exercise performance and for control of training. Sports Medicine, 22, 157175
8. Bishop, D., Jenkins, D.G., McEniery, M. and Carey, M.F (2000). Relationship between plasma lactate parameters and muscle characteristics in female cyclists. Medicine and Science in Sports and Exercise, 32, 10881093
9. Bishop, D., Bonetti, D. and Dawson, B (2002). The in.uence of pacing strategy on VO2 and supramaximal kayak performance. Medicine and Science in Sports and Exercise, 34, 10411047
10. Broker, J.P., Kyle, C.R. and Burke, E.R (1999). Racing cyclist power requirements in the 4000-m individual and team pursuits. Medicine and Science in Sports and Exercise, 31, 16771685
11. Burke, E.R (2000). Physiology of cycling. In Exercise and Sport Science (edited by W.E. Garrett and D.T. Kirkendall), pp. 759770. Philadelphia, PA: Lippincott Williams & Wilkins
12. Burke, L.M., Claassen, A., Hawley, J.A. and Noakes, T.D (1998). Carbohydrate intake during prolonged cycling minimizes effect of glycemic index of preexercise meal. Journal of Applied Physiology, 85, 22202226
13. Calles-Escandon, J., Devlin, J.T., Whitcomb, W. and Horton, E.S (1991). Pre-exercise feeding does not affect endurance cycle exercise but attenuates post-exercise starvation-like response. Medicine and Science in Sports and Exercise, 23, 818824
14. Capelli, C., Rosa, G., Butti, F., Ferretti, G., Veicsteinas, A. and di Prampero, P.E (1993). Energy cost and ef.ciency of riding aerodynamic bicycles. European Journal of Applied Physiology and Occupational Physiology, 67, 144149
15. Coyle, E.F (1995). Integration of the physiological factors determining endurance performance ability. In Perspectives in Exercise Science and Sports Medicine (edited by C.V. Gisol. and D.R. Lamb), pp. 2563. Indianapolis, IN: Benchmark Press
16. Coyle, E.F., Coggan, A.R., Hemmert, M.K., Lowe, R.C. and Walters, T.J (1985). Substrate usage during prolonged exercise following a preexercise meal. Journal of Applied Physiology, 59, 429433
17. Coyle, E.F., Coggan, A.R., Hemmert, M.K. and Ivy, J.L (1986). Muscle glycogen utilization during prolonged strenuous exercise when fed carbohydrate. Journal of Applied Physiology, 61, 165172
18. Coyle, E.F., Coggan, A.R., Hopper, M.K. and Walters, T.J (1988). Determinants of endurance in well-trained cyclists. Journal of Applied Physiology, 64, 26222630
19. Coyle, E.F., Feltner, M.E., Kautz, S.A., Hamilton, M.T., Montain, S.J., Baylor, A.M., Abraham, L.D. and Petrek, G.W (1991). Physiological and biomechanical factors associated with elite endurance cycling performance. Medicine and Science in Sports and Exercise, 23, 93107
20. Coyle, E.F., Sidossis, L.S., Horowitz, J.F. and Beltz, J.D (1992). Cycling ef.ciency is related to the percentage of type I muscle bers. Medicine and Science in Sports and Exercise, 24, 782788
21. Coyle, E.F., Jeukendrup, A.E., Oseto, M.C., Hodgkinson, B.J. and Zderic, T.W (2001). Low-fat diet alters intramuscular substrates and reduces lipolysis and fat oxidation during exercise. American Journal of Physiology Endocrinology and Metabolism, 280, E391E398
22. Craig, N.P. and Norton, K.I (2001). Characteristics of track cycling. Sports Medicine, 31, 457468
23. Davison, R.C., Swan, D., Coleman, D. and Bird, S (2000). Correlates of simulated hill climb cycling performance. Journal of Sports Sciences, 18, 105110
24. De Groot, G., Sargeant, A. and Geysel, J (1995). Air friction and rolling resistance during cycling. Medicine and Science in Sports and Exercise, 27, 10901095
25. De Koning, J.J., Bobbert, M.F. and Foster, C (1999). Determination of optimal pacing strategy in track cycling with an energy low model. Journal of Science and Medicine in Sport, 2, 266277
26. Di Prampero, P.E., Cortili, G., Mognoni, P. and Saibene, F (1979). Equation of motion of a cyclist. Journal of Applied Physiology, 47, 201206
27. Fernandez-Garcia, B., Perez-Landaluce, J., Rodriguez-Alonso, M. and Terrados, N (2000). Intensity of exercise during road race pro-cycling competition. Medicine and Science in Sports and Exercise, 32, 10021006
28. Firth, M (1998). From high-tech to low tech: another look at time trial pacing strategy. Coaching News, 3, 710
29. Foster, C., Snyder, A.C., Thompson, N.N., Green, M.A. and Foley, M (1993). Effect of pacing strategy on cycle time trial performance. Medicine and Science in Sports and Exercise, 25, 383388
30. Gnehm, P., Rechenbach, S., Altpeter, E., Widmer, H. and Hoppeler, H (1997). Influence of different racing positions on metabolic cost in elite cyclists. Medicine and Science in Sports and Exercise, 29, 818823
31. Goodpaster, B.H., Costill, D.L., Fink, W.J., Trappe, T.A., Jozsi, A.C., Starling, R.D. and Trappe, S.W (1996). The effects of pre-exercise starch ingestion on endurance performance. International Journal of Sports Medicine, 17, 366372
32. Gotshall, R.W., Bauer, T.A. and Fahrner, S.L (1996). Cycling cadence alters exercise hemodynamics. International Journal of Sports Medicine, 17, 1721
33. Hargreaves, M., Costill, D.L., Fink, W.J., King, D.S. and Fielding, R.A (1987). Effect of pre-exercise carbohydrate feedings on endurance cycling performance. Medicine and Science in Sports and Exercise, 19, 3336
34. Hawley, J.A. and Noakes, T.D (1992). Peak power output predicts maximal oxygen uptake and performance time in trained cyclists. European Journal of Applied Physiology, 65, 7983
35. Hawley, J.A. and Stepto, N.K (2001). Adaptations to training in endurance cyclists: implications for performance. Sports Medicine, 31, 511520
36. Hawley, J.A., Schabort, E.J., Noakes, T.D. and Dennis, S.C (1997). Carbohydrate loading and exercise performance: an update. Sports Medicine, 24, 7381
37. Hickey, M.S., Costill, D.L., McConell, G.K., Widrick, J.J. and Tanaka, H (1992). Day to day variation in time trial cycling performance. International Journal of Sports Medicine, 13, 467470
38. Hirvonen, J., Rekunen, S., Rusko, H. and Harkonen, M (1987). Breakdown of high-energy phosphate compounds and lactate accumulation during short supramaximal exercise. European Journal of Applied Physiology, 56, 253 259
39. Hitchins, S., Martin, D.T., Burke, L., Yates, K., Fallon, K., Hahn, A. and Dobson, G.P (1999). Glycerol hyperhydration improves cycle time trial performance in hot humid conditions. European Journal of Applied Physiology, 80, 494501
40. Horowitz, J.F., Sidossis, L.S. and Coyle, E.F (1994). High ef.ciency of type I muscle bers improves performance. International Journal of Sports Medicine, 15, 152157
41. Hultman, E (1967). Physiological role of muscle glycogen in man, with special reference to exercise. Circulation Research, 10, I-99I-114
42. Impellizzeri, F., Sassi, A., Rodriquez-Alonso, M., Mognoni, P. and Marcora, S (2002). Exercise intensity during offroad cycling competitions. Medicine and Science in Sports and Exercise, 34, 18081813
43. Ivy, J.L., Katz, A.L., Cutler, C.L., Sherman, W.M. and Coyle, E.F (1988). Muscle glycogen synthesis after exercise: effect of time of carbohydrate ingestion. Journal of Applied Physiology, 64, 14801485
44. Jentjens, R.L. and Jeukendrup, A.E (2003). eterminants of post-exercise glycogen synthesis during short-term recovery. Sports Medicine, 33, 117144
45. Jentjens, R.L., van Loon, L.J., Mann, C.H., Wagenmakers, A.J. and Jeukendrup, A.E (2001). Addition of protein and amino acids to carbohydrates does not enhance postexercise muscle glycogen synthesis. Journal of Applied Physiology, 91, 839846
46. Jeukendrup, A.E. and Jentjens, R.L.P.G (2000). Eficacy of carbohydrate feedings during prolonged exercise: current thoughts, guidelines and directions for future research. Sports Medicine, 29, 407424
47. Jeukendrup, A.E. and Martin, J (2001). Improving cycling performance how should we spend our time and money. Sports Medicine, 31, 559569
48. Jeukendrup, A.E. and Martin, D.T (2003). Are world-class cyclists really more eficient?. Medicine and Science in Sports and Exercise, 35, 12381239
49. Jeukendrup, A.E., Saris, W.H.M., Brouns, F. and Kester, A.D.M (1996). A new validated endurance performance test. Medicine and Science in Sports and Exercise, 28, 266 270
50. Jeukendrup, A., Brouns, F., Wagenmakers, A.J. and Saris, W.H (1997). Carbohydrate-electrolyte feedings improve 1 h time trial cycling performance. International Journal of Sports Medicine, 18, 125129
51. Jeukendrup, A.E., Wagenmakers, A.J., Stegen, J.H., Gijsen, A.P., Brouns, F. and Saris, W.H (1999). arbohydrate ingestion can completely suppress endogenous glucose production during exercis. American Journal of Physiology, 276, E672E683
52. Jeukendrup, A.E., Raben, A., Gijsen, A., Stegen, J.H., Brouns, F., Saris, W.H. and Wagenmakers, A.J (1999). Glucose kinetics during prolonged exercise in highly trained human subjects: effect of glucose ingestion. Journal of Physiology, 515, 579589
53. Jeukendrup, A.E., Craig, N.P. and Hawley, J.A (2001). The bioenergetics of world class cycling. Journal of Science and Medicine in Sport, 3, 414433
54. Jones, S.M. and Pass.eld, L (1998). The dynamic calibration of bicycle power measuring cranks. In The Engineering of Sport (edited by S.J. Haake), pp. 265274. Oxford: Blackwell Science
55. Keizer, H.A., Kuipers, H., van Kranenburg, G. and Geurten, (1987). Influence of liquid and solid meals on glycogen resynthesis, plasma fuel hormone response, and maximal physical working capacity. International Journal of Sports Medicine, 8, 9910
56. Kyle, C.R (1986). Mechanical factors affecting the speed of a bicycle. In Science of Cycling (edited by E.R. Burke), pp. 123136. Champaign, IL: Human Kinetics
57. Kyle, C.R (1988). The mechanics and aerodynamics of cycling. In Medical and Scienti.c Aspects of Cycling (edited by E.R. Burke and M.M. Newsom), pp. 235251. Champaign, IL: Human Kinetics
58. Kyle, C.R (1996). Selecting cycling equipment. In High- Tech Cycling (edited by E.R. Burke), pp. 143. Champaign, IL: Human Kinetics
59. Kyle, C.R. and Caiozzo, V.J (1986). Experiments in human ergometry as applied to the design of human powered vehicles. International Journal of Sport Biomechanics, 2, 619
60. Latzka, W.A., Sawka, N., Montain, S.J., Skrinar, G.S., Fielding, R.A., Matott, R.P. and Pandolf, K.B (1998). Hyperhydration: tolerance and cardiovascular effects during uncompensable exercise-heat stress. Journal of Applied Physiology, 84, 18581864
61. Liedl, M.A., Swain, D.P, Branch, J.D., Bryant, T.L., Cory, L.M. and Leete, D.S (1999). Physiological effects of constant vs variable power during endurance cycling. Medicine and Science in Sports and Exercise, 31, 14721477
62. Lucia, A., Pardo, J., Durantez, A., Hoyos, J. and Chicharro, J.L (1998). Physiological differences between professional and elite road cyclists. International Journal of Sports Medicine, 19, 342348
63. Lucia, A., Carvajal, A., Calderon, F.J., Alfonso, A. and Chicharro, J.L (1999). Breathing pattern in highly competitive cyclists during incremental exercise. European Journal of Applied Physiology, 79, 512521
64. Lucia, A., Hoyos, J., Carvajal, A. and Chicharro, J.L (1999). Heart rate responses to professional road cycling: The Tour de France. International Journal of Sports Medicine, 20, 167172
65. Lucia, A., Hoyos, J. and Chicharro, J.L (2001). Physiology of professional road cycling. Sports Medicine, 31, 325337
66. Lucia, A., Hoyos, J. and Chicharro, J.L (2001). Preferred pedalling cadence in professional cycling. Medicine and Science in Sports and Exercise, 33, 13611366
67. Lucia, A., Hoyos, J., Santalla, A., Perez, M. and Chicharro, J.L (2002). Kinetics of _ VO2 in professional cyclists. Medicine and Science in Sports and Exercise, 34, 320325
68. Lucia, A., Rivero, J.L., Perez, M., Serrano, A.L., Calbet, J.A., Santalla, A. and Chicharro, J.L (2002). Determinants of VO2 kinetics at high power outputs during a ramp exercise protocol. Medicine and Science in Sports and Exercise, 34, 326331
69. Lucia, A., Hoyos, J., Perez, M., Santalla, A. and Chicharro, J.L (2002). Inverse relationship between _ VO2max and economy/ef.ciency in world-class cyclists. Medicine and Science in Sports and Exercise, 34, 20792084
70. MacLaren, D.P.M., Reilly, T., Campbell, I.T. and Hopkins, C (1999). Hormonal and metabolic responses to maintained hyperglycaemia during prolonged exercise. Journal of Applied Physiology, 87, 12413
71. Marsh, A.P. and Martin, P.E (1997). Effect of cycling experience, aerobic power, and power output on preferred and most economical cycling cadences. Medicine and Science in Sports and Exercise, 29, 12251232
72. Marsh, A.P., Martin, P.E. and Foley, K.O (2000). Effect of cadence, cycling experience, and aerobic power on delta eficiency during cycling. Medicine and Science in Sports and Exercise, 32, 16301634
73. Martin, J.C., Milliken, D.L., Cobb, J.E., McFadden, K.L. and Coggan, A.R (1998). Validation of a mathematical model for road cycling power. Journal of Applied Biomechanics, 14, 276291
74. Maughan, R.J. and Nadel, E.R (2000). Temperature regulation and .uid electrolyte balance. n IOC Encyclopaedia of Sports Medicine: Nutrition in Sport (edited by R.J. Maughan), pp. 562573. Oxford: Blackwell Science
75. Maughan, R.J. and Shirreffs, S.M (1996). Biochemistry of Exercise IX. Champaign, IL: Human Kinetics
76. McCole, S.D., Claney, K., Conte, J.C., Anderson, R. and Hagberg, J.M (1990). Energy expenditure during bicycling. Journal of Applied Physiology, 68, 748753
77. Montain, S.J., Hopper, M.K., Coggan, A.R. and Coyle, E.F (1991). Exercise metabolism at different time intervals after a meal. Journal of Applied Physiology, 70, 882888
78. Mujika, I. and Padilla, S (2001). Physiological and performance characteristics of male professional road cyclists. Sports Medicine, 31, 479487
79. Neuman, G (2000). Cycling. In Endurance in Sport (edited by R.J. Shephard and P.O. A¢ª strand), pp. 857871
80. Oxford: Blackwell. Nikolopoulos, V., Arkinstall, M.J. and Hawley, J.A (2001). Pacing strategy in simulated cycle time-trials is based on perceived rather than actual distance. Journal of Science and Medicine in Sport, 4, 212219
81. Nilsson, L.H. and Hultman, E (1973). Liver glycogen in man: the effects of total starvation or a carbohydrate-poor diet followed by carbohydrate feeding. Scandinavian Journal of Clinical Laboratory Investigations, 32, 325330
82. Nose, H., Gack, G.W., Shi, X. and Nadel, E.R (1998). Role of osmolality and plasma volume during rehydration in humans. Journal of Applied Physiology, 65, 325331
83. Okano, G., Takeda, H., Morita, I., Katoh, M., Mu, Z. and Miyake, S (1988). Effect of pre-exercise fructose ingestion on endurance performance in fed man. Medicine and Science in Sports and Exercise, 20, 105109
84. Olds, T.S., Norton, K.I. and Craig, N.P (1993). Mathematical model of cycling performance. Journal of Applied Physiology, 75, 730737
85. Olds, T.S., Norton, K.I., Lowe, E.L., Olive, S., Reay, F. and Ly, S (1995). Modeling road-cycling performance. Journal of Applied Physiology, 78, 15961611
86. Ozyener, F., Rossitier, H.B., Ward, S.A. and Whipp, B.J (2002). Influence of exercise intensity on the on- and offtransient kinetics of pulmonary uptake in humans. Journal of Physiology, 533, 891902
87. Padilla, S., Mujika, I., Orbananos, J. and Angulo, F (2000). Exercise intensity during competition time trials in professional road cycling. Medicine and Science in Sports and Exercise, 32, 850856
88. Palmer, G.S., Hawley, J.A., Dennis, S.C. and Noakes, T.D (1994). Heart rate responses during a 4-d cycle stage race. Medicine and Science in Sports and Exercise, 26, 12781283
89. Palmer, G.S., Dennis, S.C., Noakes, T.D. and Hawley, J.A (1996). Assessment of the reproducibility of performance testing on an air-braked cycle ergometer. International Journal of Sports Medicine, 17, 293298
90. Palmer, G.S., Noakes, T.D. and Hawley, J.A (1996). Effect of steady state versus stochastic exercise on subsequent cycling performance. Medicine and Science in Sports and Exercise, 25, 684687
91. Pass.eld, L. and Doust, J.H (2000). Changes in cycling ef.ciency and performance after endurance exercise. Medicine and Science in Sports and Exercise, 32, 19351941
92. Rasmussen, B.B., Tipton, K.D., Miller, S.L., Wolf, S.E. and Wolfe, R.R (2000). An oral essential amino acidcarbohydrate supplement enhances muscle protein anabolism after resistance exercise. Journal of Applied Physiology, 88, 38639
93. Robinson, S., Robinson, D.L., Mountjoy, R.J. and Bullard, R.W (1958). Influence of fatigue on the eficiency of men during exhausting runs. Journal of Applied Physiology, 12,197201
94. Romer, L.M., McConnell, A.K. and Jones, D.A (2002). Inspiratory muscle fatigue in trained cyclists: effects of inspiratory muscle training. Medicine and Science in Sports and Exercise, 34, 785792
95. Ryschon, T.W. and Stray-Gundersen, J (1993). The effect of tyre pressure on the economy of cycling. Ergonomics, 36, 661666
96. Saris, W.H.M., van Erp-Baart, M.A., Brouns, F., Westerterp, K.R. and ten Hoor, F (1989). Study on food intake and energy expenditure during extreme sustained exercise: the Tour de France. International Journal of Sports Medicine, 10, S26S31
97. Sherman, W.M., Costill, D.L., Fink, W.J. and Miller, J.M (1981). The effect of exercise and diet manipulation on muscle glycogen and its subsequent utilization during performance. International Journal of Sports Medicine, 2, 114118
98. Smith, M.F., Davison, R.C., Balmer, J. and Bird, S.R (2001). Reliability of mean power recorded during indoor and outdoor self-paced 40 km cycling time-trials. International Journal of Sports Medicine, 22, 270274
99. Stegmann, H., Kindermann, W. and Schnabel, A (1981). Lactate kinetics and individual lactate threshold. International Journal of Sports Medicine, 2, 160165
100. Stepto, N.K., Hawley, J.A., Dennis, S.C. and Hopkins, W.G (1999). Effects of different interval-training programs on cycling time-trial performance. Medicine and Science in Sports and Exercise, 31, 736741
101. Swain, D.P (1997). A model for optimizing cycling performance by varying power on hills and in wind. Medicine and Science in Sports and Exercise, 29, 11041108
102. Swain, D., Coast, J.R., Clifford, P.S., Milliken, M.C. and Gundersen, J.S (1987). Influence of body size on oxygen consumption during bicycling. Journal of Applied Physiology, 62, 668672
103. Takaishi, T., Sugiura, T., Katayama, K., Sato, Y., Shima, N., Yamamoto, T. and Moritani, T (2002). Changes in blood volume and oxygenation level in a working muscle during a crank cycle. Medicine and Science in Sports and Exercise, 34, 520528
104. Van Ingen Schenau, G.J., Dekoning, J.J. and DeGroot, G (1994). Optimisation of sprinting performance in running, cycling and speed skating. Sports Medicine, 17, 259275
105. Van Loon, L.J., Saris, W.H., Kruijshoop, M. and Wagenmakers, A.J (2000). Maximizing postexercise muscle glycogen synthesis: carbohydrate supplementation and the application of amino acid or protein hydrolysate mixtures. American Journal of Clinical Nutrition, 72, 106111
106. Walsh, R.M., Noakes, T.D., Hawley, J.A. and Dennis, S.C (1994). Impaired high-intensity cycling performance time at low levels of dehydration. International Journal of Sports Medicine, 15, 39239
107. Wasserman, K. and McIlroy, M.B (1964). Detecting the threshold of anaerobic metabolism in cardiac patients during exercise. American Journal of Cardiology, 14, 844852
108. Westerterp, K.R., Saris, W.H.M., Van Es, M. and Ten Hoor, F (1986). se of doubly labelled water technique in humans during sustained exercise. Journal of Applied Physiology, 61, 21622167
109. White, A.P (1994). Factors affecting speed in humanpowered vehicles. ournal of Sports Sciences, 12, 419424
110. Whitt, F.R. and Wilson, D.G (1974). Bicycling Science. Cambridge, MA: MIT Press
111. Wilber, R.L., Zawadzki, K.M., Kearney, J.T., Shannon, M.P. and Disalvo, D (1997). Physiological proles of elite off-road and road cyclists. Medicine and Science in Sports and Exercise, 29, 10901094
112. Wilberg, R.B. and Pratt, J (1988). A survey of the race pro.les of cyclists in the pursuit and kilo track events. Canadian Journal of Sports Sciences, 13, 208213
113. Zawadzki, K.M., Yaspelkis III, B.B. and Ivy, J.L (1992). Carbohydrateprotein complex increases the rate of muscle glycogen storage after exercise. Journal of Applied Physiology, 72, 18541859