
Fatiga y Cambios de Atp, Fosfato de Creatina y Lactato durante La Carrera Corta de 400 M
Fatigue and Changes of ATP, Creatine Phosphate, and Lactate during The 400-M Sprint
J. Hirvonen 2, A. Nummela 1, H. Rusko 1 y S. Rehunen 3
Artículo publicado en el journal Revista de Entrenamiento Deportivo, Volumen 0 del año 2013.
Publicado 26 de marzo de 2013
Resumen
Palabras clave: lactato sanguíneo, lactato muscular, fosfocreatina, sprint
Abstract
Fatigue during the 400 m sprint was studied by measuring muscle ATP, creatine phosphate (CP), lactate (M-La), and blood lactate (B-La) in six male runners before and after four experimental sprints (100, 200, 300, and 400 m). During the first 100 m, muscle CP decreased from 15.8 ± 1.7 to 8.3 ± 0.3 mmol/kg while M-La increased to 3.6 ± 0.4 mmol/kg. After 200 m the CP had decreased to 6.5 ± 0.5 mmol/kg and M-La had increased to 8.3 ± 1.1 mmol/kg. At the end of the 400 meters, ATP and CP concentrations had decreased by 27% and 89%, respectively, and M-La had increased to 17.3 ± 0.9 mmol/kg. It was concluded that after 200 m the speed of running decreased, although CP was not depleted and lactate concentration was not at maximum level. Complete fatigue occurred when CP stores were depleted and B-La and M-La attained an individual maximum.
Keywords: blood lactate, muscle lactate, phosphocreatine, sprinting
INTRODUCCION
Las reservas de ATP en los músculos sin entrenamiento y entrenados son muy limitadas, 4-7 mmol/kg de peso húmedo (Hirvonen y otros, 1987; Rehunen y otros, 1982). La resíntesis del ATP a partir del fosfato de creatina (CP) comienza inmediatamente después de la iniciación de las contracciones musculares. El CP muscular (15-25 mmol/kg de peso húmedo) disminuye exponencialmente en los sprints máximos que duran 5, 11 y 30 segundos un 53%, 66% y 64%, respectivamente, mientras que la concentración de ATP sólo disminuye significativamente después de 11 y 30 segundos de sprint (Cheetham y otros, 1986; Hirvonen y otros, 1987; Rehunen y otros, 1982). Cuando se agotan las reservas de CP, la resíntesis de ATP se reduce notoriamente (Hultman y otros, 1981) dado que el ritmo de resíntesis del ATP a partir de fosfatos de elevada energía es mucho mayor que a partir de la glicólisis (Maréchal, 1981). Se ha sugerido que después de 5-7 segundos de sprint máximo, las menores reservas de CP puede que no cubran durante mucho tiempo la gran necesidad de ATP y decaiga por tanto la velocidad de carrera (Hirvonen y otros, 1987).
La glicólisis comienza dentro de los 5 segundos después de la iniciación del máximo ejercicio y contribuye a la provisión de energía durante el máximo ejercicio dinámico (Boobis y otros, 1983; Hultman y Sjöholm, 1983). El ritmo de la glicólisis aumenta con respecto a la decreciente concentración de CP y a la decreciente razón ATP/ADP (Chasiotis y otros, 1987). El ATP generado a partir de la glicólisis es máximo en los ejercicios exhaustivos a corto plazo que duran 40-50 seg cuando la concentración de lactato en los músculos activos alcanza el nivel de 25-30 mmol/kg (Hermansen y Vaage, 1977). La máxima concentración de lactato sanguíneo mencionada después del sprint de 400 m ha sido de 25,0 mmol/kg (Kindermann y Keul, 1977).
El propósito del presente estudio era investigar el desarrollo de la fatiga en los velocistas durante el sprint de 400 m analizando las concentraciones de ATP, fosfato de creatina y lactato en los músculos del muslo, y la concentración de lactato sanguíneo antes y después de los sprints más cortos que eran recorridos según los tiempos parciales del sprint inicial de 400 m.
MÉTODOS
Seis velocistas y corredores de medio fondo varones, de 25 años, se ofrecieron para el estudio y dieron su consentimiento informado después de conocer completamente los riesgos y beneficios de las mediciones. Todos los sujetos se entrenaron regularmente todo el año y sus marcas en los 400 m variaban entre 47,5 y 50,5 seg. Las mediciones se realizaron al final de la temporada competitiva en una pista cubierta de 200 m durante 2 días.
Durante el primer día los sujetos corrieron 400 m y 100 m y durante el segundo día corrieron 300 m y 200 m. Un sujeto sólo participó en el estudio durante el primer día, en el que corrió 400 m y 300 m. Antes de cada carrera los sujetos seguían un calentamiento minuncioso durante casi 1 hora y se realizaba un descanso de 5 horas entre las carreras cada día. Los sujetos corrieron los 400 m con el máximo esfuerzo y las demás distancias según los tiempos parciales de los 400 m iniciales. Los tiempos finales de cada carrera y los tiempos parciales de los 400 m fueron determinados con células fotoeléctricas y un cronómetro electrónico.
Las muestras de sangre venosa para determinar la concentración de lactato sanguíneo se tomaron de la vena cubital 5 min después del calentamiento e inmediatamente más tarde 3, 6, 8, 12, y 20 min (100 y 200 m) o 30 min (300 y 400 m) después de cada sprint para encontrar la máxima concentración de lactato sanguíneo. La sangre fue recogida en tubos que contenían EDTA (1,4 mg/ml) para congelarse inmediatamente y almacenarse a -75 ºC hasta que se analizara.
Las biopsias musculares se realizaron con anestesia local (lidocaína) de la piel, a través de una pequeña incisión (2 mm) con una aguja para la biopsia (TruCut®, Travenol Laboratories Inc., IL) desde la porción lateral del cuádriceps femoral, 5 min después del calentamiento e inmediatamente después (a los 3-5 seg) de cada sprint y 8 min después del sprint de 400 m.
Extractos de ácido perclórico se usaron para determinar las concentraciones de ATP, fosfato de creatina, lactato musculares y de lactato sanguíneo. Los parámetros se determinaron fluorométricamente con un Transcon 102 FN (fluoronefelómetro, Elomit Ltd., Helsinki , Finlandia) mediante los métodos enzimáticos vinculados con el nucleótico de adenina nicotinamida, descritos por Lowry y Passonneau (1972) y modificados por Näveri y otros (1978). Las concentraciones musculares de ATP, CP y lactato se calcularon en mmol/kg de peso húmedo del músculo.
Los substratos y enzimas se adquirieron de Boehringer Mannheim GmbH (Munich, Alemania) y the Sigma Chemical Company (St. Luis, MO) y el dithiothreitol (DTT) procedía de Calbiochem (Lucerna, Suiza). Todos los demás reactivos eran de grado analítico y eran de E. Merck (Darmstadt, Alemania) o de BDH Chemicals Ltd. (Poole, G.B.). Se usaron métodos convencionales para calcular las medias, errores típicos (SE) y coeficientes de correlación entre las diferentes variables. La significación estadística se puso a prueba empleando el ANOVA y el método de Scheffé (método S) junto con la prueba t para muestras emparejadas. El método de mínimos cuadrados se usaba para la curva parabólica de frecuencias acumuladas.
RESULTADOS
Los sujetos corrieron los 400 m en 51,9 ± 0,7 seg y las distancias más cortas (100, 200 y 300 m) según los tiempos parciales del sprint inicial de 400 m. La velocidad promedio de carrera disminuyó significativamente durante el sprint de 400 m (p<0.001) (Tabla 1). La velocidad comenzó a disminuir después de 200 m de sprint (p<0,05) y durante los últimos 100 m era un 15,5% (p<0,05) menor que durante los segundos 100 m. En reposo después del calentamiento, la concentración de ATP era de 4,9 ± 0,4 mmol/kg y la concentración de CP era de 15,8 ± 1,7 mmol/Kg. El ATP muscular (p<0,05) así como el CP muscular (p<0,001) disminuyó significativamente durante el sprint de 400 m (Figura 1). La máxima disminución en el CP (47,5%, p<0,001) se producía durante los primeros 100 m. Después de 200 m el CP había disminuido hasta 6,5 ± 0,5 mmol/kg, y hasta 1,7 ± 0,4 mmol/kg después de los 400 m.
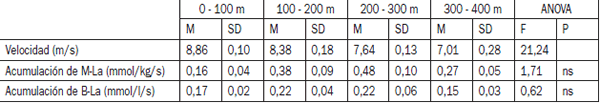
Tabla 1. Velocidad de carrera y acumulación de lactato sanguíneo y muscular durante los 100 m en el sprint de 400 m. M-La: lactado muscular; B-La: lactato sanguíneo.
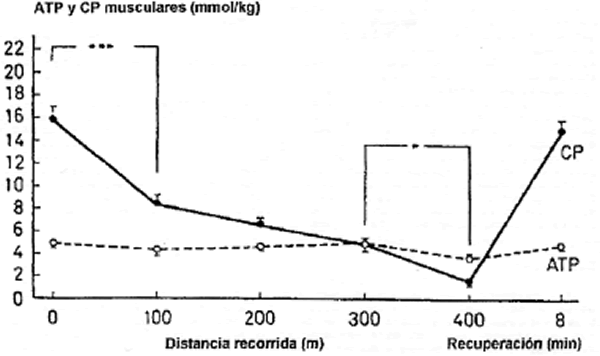
Figura 1. Cambios en la concentración de ATP y de CP para las diferentes distancias recorridas. Los valores se expresan como medias ± SE (*p<0,05; **p<0,01; ***p<0,001).
El máximo lactato sanguíneo se observaba después de los 3 min de recuperación que siguen a los sprints de 100 m y 200 m, y después de los 6 min de recuperación que siguen a los sprints de 300 m y 400 m. La correlación entre el lactato muscular y el máximo lactato sanguíneo era de 0,88 (p<0,001) (Figura 2). Las diferencias entre todas las distancias también eran estadísticamente significativas.
Después de los primeros 100 m aumentaron el lactato muscular y el sanguíneo hasta 3,6 ± 0,4 mmol/kg y 3,4 ± 0,4 mmol/l, y después de los 200 m éstos aumentaron hasta 8,3 ± 0,8 mmol/kg y 6,3 ± 0,7 mmol/l, respectivamente. Después de los 400 m, el lactato muscular era de 17,3 ± 0,8 mmol/kg, el lactato sanguíneo era de 10,8 ± 0,7 mmol/l y el máximo lactato sanguíneo 14,9 ± 0,3 mmol/l. Cuando se calculó a partir de los datos agrupados usando la curva de frecuencias acumuladas, el grado de acumulación del lactato muscular alcanzó el máximo nivel después de unos 35 seg de sprint (menos de 300 m) ; el grado de acumulación del lactato sanguíneo alcanzó el máximo nivel después de unos 27 seg de sprint (más de 200 m), después de los cuales la acumulación de lactato comenzó a disminuir.
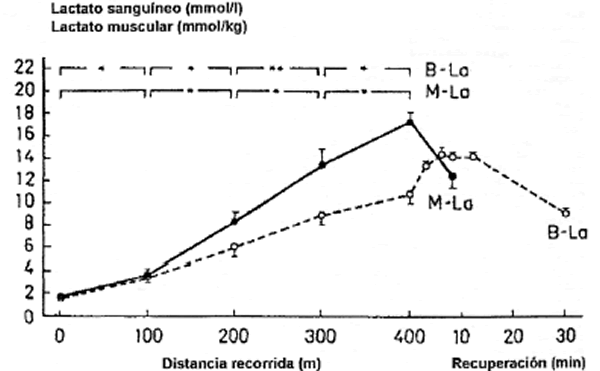
Figura 2. Cambios en el lactato sanguíneo (B-La) y el lactato muscular (M-La) para las diferentes distancias recorridas y durante la recuperación después del sprint de 400 m. Los valores se expresan como medias ± SE (*p <0,01; **p<0,001).
DISCUSIÓN
A consecuencia de las curvas cerradas de la pista cubierta, la ausencia de competición, y el momento del estudio (aproximadamente 1 mes después de la temporada competitiva), los sujetos no fueron capaces de correr los 400 m cerca de sus marcas personales en este estudio (la diferencia fue de 3,2 ± 0,4 seg). La velocidad de carrera empezó a decaer después de los 200 m, demostrando una curva de velocidad bastante usual en el sprint de 400 m. Los sujetos fueron capaces de correr las distancias más cortas cerca de los tiempos parciales en los 400 m. En consecuencia, pueden utilizarse los resultados de los análisis musculares y sanguíneos para describir el desarrollo de la fatiga y la dinámica de la energía generada en los atletas durante el sprint de 400 m.
En el presente estudio, durante los primeros 100 m la concentración del CP disminuyó rápidamente mientras que la energía glucolítica generada no fue considerablemente mejorada, como sugerían las reducidas concentraciones de lactato muscular y sanguíneo. Si los 100 m se hubieran corrido lo más rápido posible, podrían haberse observado una mayor disminución del CP y una producción del ATP considerablemente mayor a partir de la glicólisis (Hirvonen y otros, 1987). Comparando el estudio de Hirvonen y otros (1987) con el presente estudio, la principal diferencia en la energía generada entre el sprint máximo y submáximo parece estar en el grado de glicólisis, dado que el grado de acumulación del lactato sanguíneo era 3,2 veces mayor durante el máximo sprint de 100 m que durante los primeros 100 m proporcionales en el sprint de 400 m.
La comparación entre el sprint de 400 m y el de 100 m sugiere que los fosfatos de elevada energía se usan para el ATP generado mientras sea posible y que la producción de energía glucolítica mejora cuando la concentración de CP disminuye un 50-60% del nivel de reposo. Según los estudios previos, el CP también ha disminuido excesivamente durante la fase inicial de la máxima carrera (Cheetham y otros, 1986; Hirvonen y otros, 1987), lo que sugiere que durante la fase de aceleración de la carrera la mayoría del ATP se resintetiza mediante la degradación del CP.
La velocidad de carrera empezó a disminuir después del sprint de 200 m aunque había bastante energía en forma de ATP y CP en los músculos que trabajan. Esto podría explicarse con el mejor grado de resíntesis del ATP y con el cambio en la utilización de diferentes fibras musculares. Después de los 200 m el ATP generado a partir de la glicólisis mejoraba considerablemente, como sugería el máximo grado de acumulación del lactato en el músculo y en la sangre durante los terceros 100 m. Simultáneamente, el ATP generado a partir del CP era menor, como muestra la pequeña disminución en el CP entre los 200 m y los 300 m. Anteriores estudios han mostrado que la resístesis del ATP a partir de la glicólisis es suprema en los máximos ejercicios que duran 40-50 seg (Kindermann y Keul, 1977). Dado que el grado de resístesis del ATP a partir del CP es mucho mayor que a partir de la glicólisis, la menor energía generada podría explicar la reducida velocidad de carrera.
Otra explicación podría ser un cambio en la utilización de diferentes fibras musculares después del sprint de 200 m. Durante la segunda mitad de los 400 m, la relativa proporción de fibras de concentración lenta (ST) en la generada por la fuerza aumenta como resultado de la mayor fatigabilidad de las fibras de contracción rápida (FT) (Tesch, 1980). Además, Rehunen y otros (1982) han sugerido que durante el ejercicio a corto plazo las fibras FT consumen más reservas del CP que las fibras ST, y en los velocistas entrenados el CP posiblemente se resintetiza más rápidamente en las fibras musculares FT que en las ST.
La velocidad de carrera disminuyó drásticamente durante los últimos 100 m, indicando la fatiga muscular. En esta fase, el CP se agotó y el ritmo del ATP generado a partir de la glicólisis se retrasaba, como sugería el menor grado de acumulación del lactato. El ATP glucolítico generado disminuyó al final de los 400 m, probablemente porque la acidez muscular y el lactato acumulado inhibe la glicólisis (Karlsson y otros, 1974; Tesch, 1978; Trivedi y Danforth, 1966).
Mientras que la asociación de la acidosis con la fatiga está clara, una implicación adicional de que es la causa de la fatiga es mucho menos segura. El problema es que gran número de cambios bioquímicos y biofísicos se producen al mismo tiempo mientras la fatiga se está desarrollando. Dawson y otros (1978), por ejemplo, mostraron una buena correlación entre cuatro variables (CP, ADP, ΔGATP, y pHi) y la decadencia de la tensión muscular. Además, existen mecanismos para invertir o excluir la inhibición del pH del PFK muscular (Dobson y otros, 1986). En el presente estudio, el tiempo final en el sprint de 400 m no se conectó ni con la concentración de lactato muscular ni la del sanguíneo. En consecuencia, el grado de acidosis del lactato no es la única explicación para el rendimiento atlético de gran nivel en el sprint de 400 m.
En conclusión, los resultados de este estudio demuestran que durante la fase de aceleración del sprint de 400 m, la mayoría del ATP se resintetiza mediante la ruptura del CP y el papel de la glicólisis es pequeño. Entre los 100 m y los 200 m, se reduce la contribución del fosfato de elevada energía y aumenta el papel de la glicólisis. Después de los 200 m, se observa la fatiga, como muestra la menor velocidad de carrera, aunque el CP no se agota y la concentración de lactato no está en el máximo nivel. Al final del sprint de 400 m, se agotan las reservas de CP y la concentración de lactato alcanza un máximo individual.
Referencias
1. Boobis, L.H.; Williams, C. y Wootton, A. (1983). Human muscle metabolism during brief maximal exercise.. J. Physiol. 338: 21 -22.
2. Chasiotis, D.; Bergström, M., y Hultman, E. (1987). ATP utilization and force during intermittent and continuous muscle contractions.. J. Appl. Physiol. 63(1): 167-174.
3. Cheetham, M.E.; Boobis, L.H.; Brooks, S. y Williams, C. (1986). Human metabolism during sprint running.. J. Appl. Physiol. 61(1): 54-60.
4. Dawson, M.J.; Gadian, D.G. y Wilkie, D.R. (1978). Muscular fatigue investigated by phosphorus nuclear magnetic resonance.. Na¬ture 274: 861-866.
5. Dobson, G.P.; Yamamoto, E., y Hochachka, P.W (1986). Phospho¬fructokinase control in muscle: Nature and reversal of pH-dependent ATP inhibition. . Am. J. Physiol. 250: 71-76.
6. Hermansen, L. y Vaage, O. (1977). disappearance and glycogen synthesis in human muscle after maximal exercise. . Am. J. Physiol. 233: 422-429.
7. Hirvonen, J.; Rehunen, S.; Rusko, H., y Härkönen, M. (1987). Breakdown of high-energy phosphate compounds and lacta¬te accumulation during short supramaximal exercise. . Eur. J. Appl. Physiol. 56: 253-259.
8. Hultman, E., y Sjöholm, H. (1983). metabolism and contraction force of skeletal muscle in situ during electrical stimu¬lation. . J. Physiol. 345: 525-532.
9. Hultman, E.; Sjöholm, H.; Sahlin, K., y Ëdström, L. (1981). Hultman, E.; Sjöholm, H.; Sahlin, K., y Ëdström, L. (1981). Glyco¬lytic and oxidative energy metabolism and contraction charac¬teristics of intact human muscle. . In: R. Porter and J. Whelan (eds.), Human Muscle Fatigue: Physiological Mechanismus, pp. 19-40. London: Pitman Medical.
10. Karlsson, J.; Hultén, B., y Sjödin, B. (1974). Substrate activation and product inhibition of LDH activity in human skeletal mus¬cle. . Acta Physiol. Scand. 92: 21-26.
11. Kindermann, W., y Keul, J. (1977). ). Lactate acidosis with different forms of sports activities. . Can. J. Appl. Sports Sci. 2: 177-182.
12. Lowry, O.H., y Passonneau, J.V. (1972). A Flexible System of Enzy¬matic Analysis. . New York: Academic Press.
13. Maréchal, G. (1981). Immediate energy sources in muscle contraction. . In: E. Jokl (ed.). Medicine and Sport, pp. 1-9. Basel: Karger.
14. Näveri, H.; Rehunen, S.; Kuoppasalmi, K.; Tulikoura, I., y Härkö¬nen, M. (1978). Muscle metabolism during and after strenuous intermittent running. . Scand. J. Lab. Invest. 38: 329-336.
15. Rahunen, S.; Näveri, H.; Kuoppasalmi, K., y Härkönen, M. ( (1982). High-energy phosphate compounds during exercise in human slow-twitch and fast-twitch muscle fibres. . Scand. J. Clin. Lab. Invest. 42(6): 499-506.
16. Tesch, P.A. (1978). Muscle fatigue in man and lactate concentra¬tion. . In: E. Asmussen y K. Jørgensen (eds.), Biomechanics IV-A, pp. 68-72. Baltimore: University Park Press.
17. Tesch, P.A. (1980). Fatigue pattern in subtypes of human skeletal muscle fibers. . Int. J. Sports Med. 1(2): 78-81
18. Trivedi, B. y Danforth, W.H. (1966). Effect of pH on the kinetics of frog muscle phosphofructokinase. . J. Biol. Chem. 241: 4110- 4114.