
Suplementación con Creatina e Influencia de la Edad en el Metabolismo Muscular durante el Ejercicio
Sinclair A Smith1, Scott J Montain1, Ralph P Matott2, Gary P Zientara3, Ferenc A Jolesz3 y Roger A Fielding1
1Department of Health, Human Performance, and Recreation, Baylor University.
2United States Army Research Institute of Environmental Medicine, Kansas Street, Building 42, Natick, MA 01760, Estados Unidos.
Artículo publicado en el journal PubliCE, Volumen 0 del año 1998.
Publicado 18 de febrero de 2004
Resumen
Palabras clave: envejecimiento, monohidrato de creatina, músculo esquelético, espectroscopia de resonancia magnética
INTRODUCCION
Los estudios han reportado reducciones relacionadas a la edad en el tamaño de los músculos esqueléticos, diámetro de las fibras tipo II, actividad enzimática mitocondrial, metabolismo de los fosfatos de alta energía, las cuales están asociadas con la declinación en la fuerza muscular y la capacidad de resistencia que ocurre con el envejecimiento (5, 7, 8, 21, 22, 34). Sin embargo, los procesos subyacentes, a través de los cuales, ocurren estos cambios no están bien entendidos. Los niveles reducidos de fosfocreatina en reposo (PCr) reportados en la vejez pueden ser en parte responsables de estas declinaciones (22, 29). Ha sido demostrada que la declinación en la disponibilidad de la PCr es un potencial factor contribuyente a la fatiga muscular durante el ejercicio de intensidad moderada a máxima (6, 16). Además, ha sido reportado que las tasas de resíntesis de PCr después del ejercicio declinan con la edad hasta ≈8 % cada 10 años después de los 30 años de edad (24). Generalmente se piensa que la resíntesis de PCr esta regulada por la creatín quinasa unida a la membrana de las mitocondrias y que la tasa inicial de recuperación de PCr es proporcional a la tasa de consumo de oxígeno mitocondrial (18, 20, 21, 26, 31, 33, 34).
Ha sido reportado que la suplementación dietaria con creatina (20-30 g/día durante 4-6 días) incrementa la concentración de creatina muscular en tanto como 50% y mejora el rendimiento muscular durante series de ejercicio intermitente de alta intensidad (2-4, 10, 12, 13, 15). El efecto mejorador del rendimiento de la creatina resulta a partir de un incremento de la disponibilidad muscular de cretina que mantiene la inicialmente rápida resíntesis de PCr durante la recuperación e incrementa la disponibilidad de PCr durante las siguientes series de ejercicio (3, 12, 15). Ningún estudio ha investigado los efectos de la creatina en ancianos, los cuales debido a déficit intrínsecos en el metabolismo muscular, se pueden beneficiar de la suplementación con creatina.
El propósito de este estudio fue determinar los efectos de la suplementación con creatina y la edad sobre el metabolismo muscular de PCr y el rendimiento, usando espectroscopia de resonancia magnética de 31P (MRS). La 31P-MRS provee mediciones frecuentes, en serie y no invasivas de los componentes del fósforo intramuscular y mejora enormemente la resolución por sobre la técnica de biopsia muscular (27), el método más frecuente de evaluación de los efectos de la suplementación con creatina sobre el metabolismo muscular hasta la fecha. Nosotros hipotetizamos que las personas de mediana edad tendrían menores concentraciones de PCr muscular en reposo y menores tasas de resíntesis de PCr que los adultos jóvenes con hábitos de actividad y dieta similares y que la suplementación con creatina tendría mayores efectos en el metabolismo de la PCr muscular en las personas de mediana edad.
METODOS
Sujetos
Sujetos de mediana edad (>50 años) y jóvenes (<40 años) fueron reclutados para la participación en el estudio. El grupo más joven consistió de 4 hombres y 1 mujer y el grupo de mediana edad de 3 hombres y 1 mujer. Las características físicas de los sujetos son presentadas en la Tabla 1. Todos los sujetos estaban libres de enfermedades crónicas y de medicaciones regulares, como fue determinado por un cuestionario de historia clínica. El estudio fue aprobado por el Comité de Revisión Institucional apropiado, y todos los sujetos dieron su consentimiento voluntario antes de la participación.
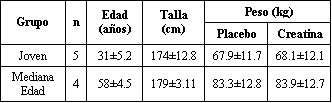
Tabla 1.
Características físicas de los sujetos. Los valores son medias±DS; n, número de
sujetos.
El nivel de actividad física habitual no fue diferente entre los grupos jóvenes y de mediana edad, como fue indicado por el Cuestionario de los Alumnos de Hardvard (Harvard Alumni Questionnaire) (30). Además, debido a un potencial efecto de la dieta sobre la creatina muscular, se le pregunto a los sujetos acerca de sus hábitos dietarios habituales. No había vegetarianos en el estudio, y todos los sujetos reportaron un consumo de al menos 5 comidas con carne a la semana.
Suplementación con Creatina
Fueron realizadas dos pruebas de ejercicio a ciego: una prueba placebo seguida por una prueba con creatina 7 días después. Las pruebas no fueron aleatorias, ya que los niveles de creatina de los músculos esqueléticos pueden permanecer elevados arriba de los niveles basales por 4-5 semanas después de interrumpir la suplementación (17). 5 días antes de cada prueba, los sujetos comenzaron a consumir 0.3 g.kg-1.día-1 de ya sea, placebo (azúcar granulada) o 3 g. kg-1.día-1 de monohidrato de creatina (Phosphagen, Experimental and Applied Sciences, Pacific Grove, CA) combinada con 3 g. kg-1.día-1 de una mezcla de bebida en polvo saborizada. La mezcla fue disuelta en agua y consumida cuatro veces al día.
Ejercicio
Ambos grupos realizaron un ejercicio de extensión unilateral de rodilla mientras estaban acostados dentro de un sistema de resonancia magnética 1.5 T (RM) (General Electric SIGNA, General Electric Medical Systems, Milwaukee, WI) para todo el cuerpo. El aparato de ejercicio proporcionaba resistencia concéntrica por medio de un brazo de palanca y un sistema de polea integrado con una rueda y una correa de resistencia (Figura 1). Una cuerda elástica retornaba el brazo de palanca a la posición inicial después de la extensión de rodilla. Las extensiones de rodillas fueron realizadas desde ≈ 110 hasta ≈ 145° de la extensión de rodilla a 37 contracciones/min, establecidas por un metrónomo audible. La producción de potencia durante el ejercicio fue determinada midiendo la tensión y el desplazamiento aplicado a una polea en línea y por medio de la estimación de la masa de la pierna (36).
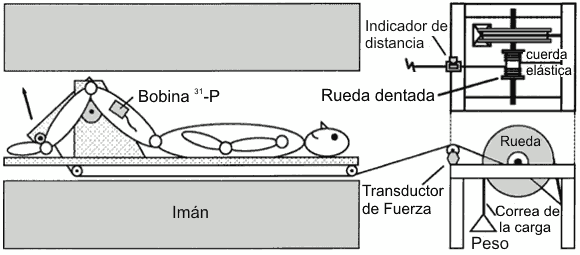
Figura 1. Diagrama
esquemático del aparato para hacer ejercicio.
Durante las pruebas experimentales, fueron realizadas tres series de ejercicio unilateral, separadas por 3 min de recuperación. Las series 1 y 2 fueron de 2 min de duración cada una, y la serie 3 fue continuada hasta el agotamiento. El agotamiento fue definido como el tiempo en donde la tasa y/o rango de movimiento no pudieran ser mantenidos por el sujeto después de haber sido dado aliento verbal por los investigadores.
Antes de las pruebas experimentales, fueron realizadas de 2 a 3 sesiones de práctica para familiarizar a los sujetos con los procedimientos experimentales y para determinar la intensidad de ejercicio apropiada. Así fue determinada la máxima carga de la rueda con la cual el sujeto fue capaz de realizar series de ejercicios de 2 min. Para alcanzar el agotamiento en 1-2 min, la carga fue ligeramente incrementada en la serie 3. La carga para cada serie fue constante durante las pruebas placebo como con creatina. Ambas piernas fueron evaluadas en cada condición experimental.
31P-MRS
El espectro del 31P fue recolectado durante el ejercicio a través de una bobina de superficie de 11 cm de transmisión y recepción de radio frecuencia dual 1H/31P (USAsia, Columbus, OH) colocada en los músculos cuadriceps. Los datos del 31P fueron adquiridos usando una excitación de pulso de 25.85 MHz (amplitud del pulso 600 μs), tiempo de repetición= 1000 ms, amplitud del espectro 2000 Hz, y 1024 puntos atenuación por inducción libre (FID). Antes del ejercicio, una imagen de RM protónica fue adquirida axialmente usando la bobina de superficie de 1H/31P para verificar la colocación de la bobina y la participación de los grupos musculares. Fue realizado un procedimiento de gradiente lineal para reducir la inhomegeneidad dentro del volumen sensible. Las ganancias de transmisión y recepción de la bobina de superficie para 31P-MRS fueron establecidas una vez para maximizar la señal de la PCr adquirida a partir del músculo y fue mantenida constante a través de todo el estudio. Fueron promediadas 10 señales FID, produciendo un espectro cada 10 s. Se tomo la precaución de asegurar que el ejercicio empezara y terminara al inicio de un ciclo FID. El sistema de RM fue calibrado para usar standars conocidos en cada día de evaluación.
El procesamiento FID consistió de la apodización de líneas de ensanchamiento de 10-Hz, puntos de 0 a 4096 y transformada de Fourier, seguido de un ajuste de fase de orden cero y primero. Las concentraciones relativas de Pi, PCr y β-ATP fueron determinadas a partir de las áreas pico del espectro. Los índices PCr/ β-ATP en reposo, Pi/ β-ATP en reposo y β-ATP/ β-ATP en reposo (donde el β-ATP en reposo fue el área media de los seis picos iniciales de β-ATP en reposo recolectados antes del ejercicio) fueron convertidos a milimoles por kilogramo de peso húmedo asumiendo que el área del β-ATP en reposo era equivalente a 5.5 mmol/kg de peso húmedo (33). La cantidad relativa de hidrólisis de PCr durante el ejercicio fue representada como el cambio en la PCr desde el reposo hasta el valor de PCr más bajo alcanzado durante cada serie de ejercicio. Finalmente, el pH fue calculado usando el cambio químico entre la frecuencia de Pi y PCr (33).
A partir de una curva monoexponencial desarrollada para los datos de recuperación de PCr (18, 20, 26, 34) fueron determinadas la tasa de resíntesis de PCr y una constante de tiempo (Tc) que indicaba la mitad de tiempo de recuperación. Fue usada la siguiente ecuación: y= a[1- exp (bx)] + c, donde y representa el valor de PCr a cualquier tiempo x dado, a es el cambio en la PCr durante la recuperación, b es la tasa constante (1/b= Tc), y c es el valor inicial de PCr al inicio de la recuperación. Los valores para la tasa inicial de resíntesis de PCr fueron determinados a partir de la pendiente de los 10 s iniciales de la curva monoexponenxial desarrollada (20, 34). La tasa inicial de resíntesis de PCr y Tc PCr fueron usadas para describir la cinética de la PCr, y no para predecir la capacidad oxidativa muscular como en estudios previos (1, 20, 34).
Análisis Estadísticos
Fueron usados test de análisis de varianza para analizar la PCr, Pi, ATP, pH, hidrólisis de PCr, tasa inicial de resíntesis de PCr, Tc, producción de potencia, y tiempo hasta el agotamiento con el tratamiento (creatina vs. placebo), edad (jóvenes vs. mediana edad), pierna (derecha vs. izquierda) y series de ejercicio o recuperación como factores. Fue usado un test post hoc Newman-Keuls para determinar las diferencias medias entre y dentro de los factores. Un test t apareado fue usado para determinar las diferencias medias en el peso corporal entre las pruebas con placebo y creatina para los grupos jóvenes y de mediana edad. La concentración inicial de PCr, hidrólisis de PCr y tiempo hasta el agotamiento para la serie 3 de ejercicio fueron correlacionados usando correlaciones momento producto de Pearson. El nivel de significancia fue establecido a una p<0.05, y los resultados son presentados como media±DS.
RESULTADOS
Edad
Durante la prueba placebo, la PCr en reposo y durante la recuperación, fue menor en el grupo de mediana edad en comparación con el grupo joven (reposo 35.0±5.2 vs. 39.5±5.1 mmol/kg, p<0.05) como es ilustrado en la Figura 2. La hidrólisis de la PCr (Δ PCr, Tabla 2) durante las series de ejercicio tendió a ser menor en el grupo de mediana edad (p=0.06), mientras que la producción de potencia no fue diferente entre los grupos (Tabla 2). Como fue indicado por las pruebas de práctica, la carga del ejercicio fue incrementada en la serie 3 para asegurar que el agotamiento ocurriera en el min 1-2 (Tabla 2). Un sujeto joven fue excluido de todos los análisis de hidrólisis de PCr, tasa inicial de resíntesis de PCr y tiempo hasta el agotamiento debido a dificultades experimentales durante el ejercicio en la prueba placebo. El grupo joven para estas mediciones consistió de 3 hombres y 1 mujer, de 30±5 años de edad.
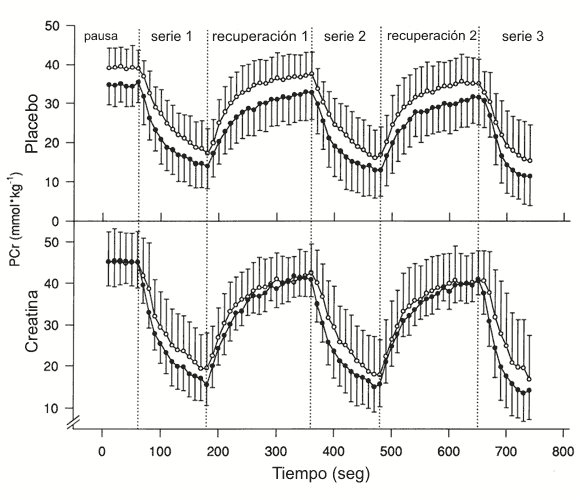
Figura 2. Resultados
de la fosfocreatina (PCr; mmol/kg de peso húmedo) vs. tiempo (s) en reposo y a
través de todas las series de ejercicio y recuperación para las pruebas de
creatina y placebo. ○ Sujetos jóvenes, ● sujetos de mediana edad. Los valores
son presentados como media±DS.
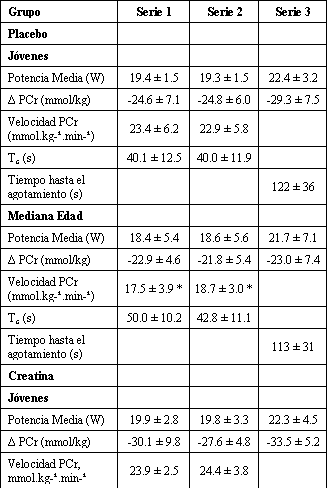
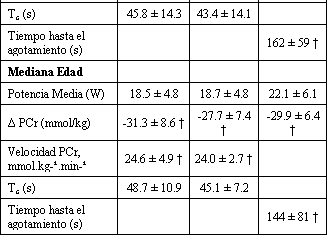
Tabla 2. Resultados
metabólicos de la PCr y rendimiento muscular. Los valores son presentados como
media±DS. Resultados de la producción de potencia media de los sujetos jóvenes y
de mediana edad durante el ejercicio (W), hidrólisis (Δ PCr, mmol/kg de peso
húmedo), velocidad inicial de resíntesis de PCr (Velocidad PCr, mmol.kg de peso
húmedo-1.min-1), constante de tiempo de resíntesis de PCr
(Tc, s) y tiempo hasta el agotamiento (s) para las series de ejercicio y
recuperación 1, 2, y 3 de las pruebas placebo y creatina. El Δ PCr para la serie
de ejercicio 3 representa la hidrólisis de PCr que ocurrió a partir del inicio
de la serie 3 hasta el agotamiento o después de la finalización de 2 min de
ejercicio, de acuerdo a cual ocurrió primero. * Diferencias significativas con
respecto al grupo joven (p<0.05). † Diferencias significativas con respecto a la
prueba placebo (p<0.05).
Después del ejercicio la tasa inicial de resíntesis de PCr fue menor en el grupo de mediana edad en comparación con el grupo joven (18±3.5 vs. 23.2±6.0 mmol.kg-1.min-1, p<0.05) como es ilustrado en la Figura 3. La Figura 4 muestra la curva monoexponencial desarrollada para los datos de PCr desde el primer período de recuperación para un sujeto representativo joven y de mediana edad. El coeficiente de determinación medio (r2) para todos los datos de la curva monoexponencial fue de 0.97±0.03 (p<0.01). La Tc obtenida a partir de la ecuación monoexponencial fue similar en el grupo joven y de mediana edad (Figura 3). Además, el Pi (Figura 5) y el pH (Tabla 3) no fueron diferentes entre los grupos durante el reposo, el ejercicio o la recuperación.
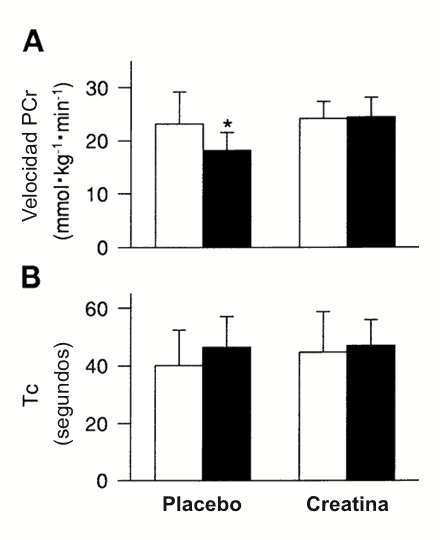
Figura 3. Tasa
inicial de resíntesis de PCr promedio de las series de recuperación 1 y 2 (A;
mmol.kg de peso húmedo-1.min-1) y constante de tiempo de
PCr (B; Tc; s) son representados para las pruebas placebo y creatina. Barras
blancas, sujetos jóvenes; barras oscuras, sujetos de mediana edad. Los valores
son presentados como media±DS. * Diferencias significativas con respecto a la
prueba de creatina del grupo de mediana edad (p<0.05) y diferencias
significativas a partir de la prueba placebo del grupo joven (p<0.05).
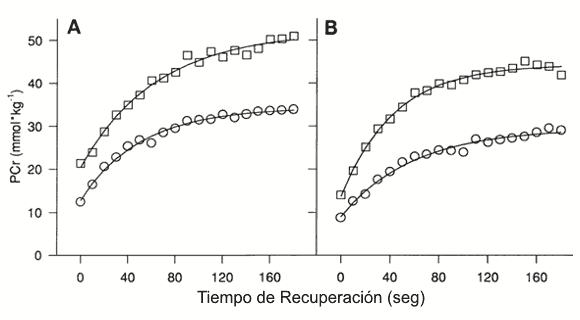
Figura 4. Resultados
de PCr de los individuos (mmol/kg de peso húmedo) vs. tiempo (s) durante la
recuperación número 1 para un sujeto representativo joven (A) y de mediana edad
(B). ○ prueba placebo; □ prueba creatina.
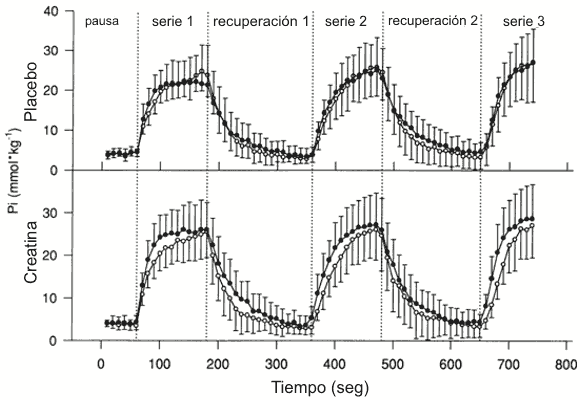
Figura 5. Resultados
de Pi (mmol/kg de peso húmedo) vs. tiempo en reposo y a través de todas las
series de ejercicio y recuperación para las pruebas de creatina y placebo. ○
Sujetos jóvenes, ● sujetos de mediana edad. Los valores son presentados como
medias±DS.
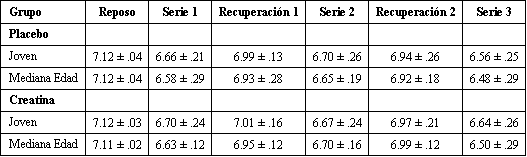
Tabla 3. Resultados
del pH muscular. Los valores son presentados como media±DS y representan el pH
al final de las series de ejercicio 1 y 2 y al final de los períodos de
recuperación 1 y 2 para los sujetos jóvenes y de mediana edad. Los valores de pH
para la serie 3 representan el pH al final de la serie 3 o después de la
finalización de 2 min de ejercicio, de acuerdo a lo que haya ocurrido primero.
Suplementación con Creatina
Después de la suplementación con creatina, la PCr en reposo y de la recuperación se incrementaron en el grupo joven en un 15 % (en reposo 45.7±7.5 mmol/kg, p<0.05) y en el grupo de mediana edad en un 30 % (en reposo 45.7±5.5 mmol/kg, p<0.05), eliminando la diferencia en la PCr de reposo entre los grupos joven y de mediana edad (Figura 2). La hidrólisis de la PCr durante el ejercicio se incremento en el grupo de mediana edad (p<0.05) después de la suplementación con creatina y tendió a incrementarse en el grupo joven (p=0.1, Tabla 2). Hubo una correlación positiva entre la concentración inicial de PCr y la hidrólisis de la PCr para ambos grupos durante la serie de ejercicio número 3 (r=0.72, p<0.05). La producción de potencia permaneció constante a través de las pruebas placebo y creatina para ambos grupos (Tabla 2).
La suplementación con creatina incrementó el tiempo medio hasta el agotamiento durante la serie de ejercicio 3 en un 30 % en los grupos joven y de mediana edad combinados (118±34 vs. 154±70 s, p<0.05, Tabla 2). No hubo interacciones o diferencias significativas en el tiempo hasta el agotamiento con respecto a la edad. Todos los sujetos fueron alentados por los investigadores durante el ejercicio y pareció que realizaron un esfuerzo máximo durante la serie de tiempo hasta el agotamiento. El tiempo hasta el agotamiento no estuvo significativamente correlacionado con la concentración inicial de PCr (r=0.26, p=0.14) o la hidrólisis de PCr (r=0.34, p=0.06) durante la serie de ejercicio número 3.
La tasa inicial de resíntesis de PCr se incrementó en el grupo de mediana edad (24.3±3.8 mmol.kg-1.min-1, p<0.05) a un nivel no diferente con respecto al grupo joven (24.2±3.2 mmol. kg-1.min-1) después de la suplementación con creatina (Figura 3). Además, la tasa de resíntesis rápida de PCr pareció continuar dentro de la recuperación, manteniendo las elevadas concentraciones de PCr en ambos grupos a través del tiempo (Figura 4). La suplementación con creatina no cambió significativamente la Tc de resíntesis de PCr en cualquiera de los grupos (Tabla 2, Figura 3).
El Pi en reposo no fue afectado por la suplementación con creatina en el grupo joven y de mediana edad (Figura 5). La producción de Pi durante el ejercicio tendió a incrementarse luego de la suplementación con creatina (Figura 5) y fue similar a los cambios observados en la hidrólisis de PCr (Tabla 2). El β-ATP (no mostrado) permaneció constante desde del reposo a través de todo el ejercicio y la recuperación, y no hubo diferencias significativas en el β-ATP entre los grupos o entre las pruebas experimentales. Además, el pH declinó durante el ejercicio (p<0.05) y no hubo diferencias significativas entre los grupos o pruebas (Tabla 3).
DISCUSION
Este estudio investigó los efectos de la edad en el metabolismo muscular durante el ejercicio por medio de la comparación de sujetos jóvenes y de mediana edad (Tabla 1). Además, fueron examinados los efectos de la suplementación oral con creatina sobre el metabolismo muscular de la PCr y el rendimiento. Para recolectar los datos metabólicos y de rendimiento, simultáneamente, el ejercicio de extensión de rodilla fue realizado dentro de un sistema de RM (Figura 1). Durante el ejercicio, la cadencia y la carga fueron mantenidas constantes, y el tiempo hasta el agotamiento fue usado como una medición de la capacidad de resistencia muscular del músculo cuadriceps. Fue empleada 31P-MRS no invasiva para medir la PCr, Pi, ATP y pH del músculo cuadriceps a través de todo el ejercicio y la recuperación durante una prueba de ejercicio, con suplementación con placebo y creatina, de mediciones repetidas. Nosotros hipotetizamos que las personas de mayor edad tendrían menores concentraciones de PCr de reposo y una menor velocidad inicial de resíntesis de PCr que las personas más jóvenes y que la suplementación oral con creatina provocaría mayores incrementos en el metabolismo muscular de la PCr en las personas de mayor edad.
Edad
Nuestros resultados confirman los reportes acerca de que las personas de mayor edad tienen una menor concentración muscular en reposo de PCr en comparación con las personas más jóvenes (22, 29) (Figura 2). La disponivilidad de PCr en reposo fue un 11 % menor en el grupo de mediana edad (p<0.05), y la cantidad de PCr hidrolizada durante el ejercicio tendió a ser menor (p=0.06, Tabla 2). Ha sido reportado que con la edad ocurre un incremento en el porcentaje de fibras tipo I dentro del músculo y esto puede explicar la concentración de PCr en reposo reducida, encontrada en sujetos ancianos debido a las característicamente menores concentraciones PCr en las fibras tipo I (7). Sin embargo, debería ser destacado que no todos los estudios que investigaron los cambios relacionados a la edad en el metabolismo muscular, reportaron reducciones en la PCr en reposo e incrementos en el porcentaje de fibras tipo I (21, 22, 32). McCully et al. (22) reportaron niveles de PCr/Pi en reposo que fueron ≈ 38% menores en sujetos de mediana edad (66.8±1.9 años) y ancianos (80.0±5.1 años) en comparación con sujetos jóvenes (24.6±4.7 años). En un estudio subsecuente, McCully et al. (21) no encontraron diferencias en los valores PCr/Pi en reposo, ninguna diferencia en la distribución de tipos de fibras en sujetos ancianos (66.0±6.0 años) vs. jóvenes (28.2±6.8 años). Dados estos hallazgos, el efecto del envejecimiento sobre la concentración de PCr en reposo y la distribución de tipos de fibras requieren más investigación.
Los resultados de este estudio también están de acuerdo con reportes acerca de que los individuos de mayor edad tienen una velocidad de resíntesis de PCr reducida después del ejercicio (8, 21-24) (Figura 3). La velocidad inicial de resíntesis de PCr representa la velocidad a la cual la creatín quinasa repleta la PCr inmediatamente después del ejercicio y puede ser afectada por la concentración de creatina muscular (1, 12, 26, 33). Los sujetos de mediana edad tenían una velocidad inicial de resíntesis de PCr menor, la cual puede estar asociada con sus concentraciones de PCr en reposo reducidas (Figura 2). A medida que la PCr es hidrolizada para resintetizar ATP durante el ejercicio, la concentración de creatina libre se incremento en el músculo. Durante la recuperación, las concentraciones de creatina muscular elevadas condujeron la reacción de la cratín quinasa hacia la producción de PCr (28). Una baja concentración muscular de PCr es indicativa de una baja disponibilidad de creatina total, lo cual puede limitar la resíntesis de PCr por medio de la creatín quinasa (3, 12, 15). Además, ha sido reportado que un incremento en la disponibilidad de creatina muscular provocado por la suplementación oral con creatina, prolonga la velocidad inicial rápida de la resíntesis de PCr muscular durante la recuperación (12).
La capacidad oxidativa mitocondrial muscular y el pH también afectan la recuperación de PCr, y de este modo, pueden influenciar las mediciones de velocidad inicial de resíntesis y de Tc (1, 26, 33). En este estudio, no hubo ninguna diferencia en el pH entre los grupos o pruebas (Tabla 3). Ha sido reportado que tanto la Tc de recuperación de PCr como la velocidad inicial de resíntesis de PCr son independientes del pH y de la intensidad de ejercicio cuando la variación en el pH es similar a aquella observada entre los grupos jóvenes y de mediana edad en este estudio (26). Aunque hubo diferencias en la velocidad inicial de resíntesis de PCr entre los grupos (Figura 3), esta parece ser la fase de recuperación menos afectada por el pH y depende más del ATP muscular y las concentraciones de creatina (1, 26, 33). De este modo, es improbable que el pH influencie significativamente las diferencias en la velocidad inicial de resíntesis de PCr observada entre los grupos jóvenes y de mediana edad.
Ha sido encontrado que la capacidad oxidativa muscular declina con la edad y puede haber limitado la velocidad inicial de resíntesis de PCr en el grupo de mediana edad, limitando el suministro de ATP requerido por la creatín quinasa para resintetizar PCr después del ejercicio (5, 7, 8, 21, 22, 34). Sin embargo, dado que el único cambio en la rutina diaria de los sujetos entre pruebas fue la ingestión de creatina, nosotros asumimos que la capacidad oxidativa muscular no fue afectada durante los 7 días entre las pruebas y que los cambios en el metabolismo de PCr después de la suplementación con creatina resultaron a partir de factores distintos que la capacidad oxidativa.
Suplementación con Creatina
Después de la suplementación oral con creatina, la concentración de PCr en reposo se incrementó en el grupo joven (p<0.05), y en una mayor medida, en el grupo de mayor edad (p<0.05), eliminando las diferencias entre los sujetos jóvenes y de mediana edad (Figura 2). La variación en la respuesta a la suplementación con creatina de los grupos jóvenes y de mediana edad es consistente con los reportes acerca de que las personas que tienen concentraciones de PCr muscular de reposo relativamente bajas tienden a tener mayores incrementos en la PCr muscular en reposo después de la suplementación con creatina (12, 15).
Además del incremento en la PCr en reposo, hubo un incremento en la hidrólisis de PCr durante el ejercicio en el grupo de mediana edad sin un cambio en la producción de potencia en el ejercicio (Tabla 2), sugiriendo que una mayor proporción de ATP suministrado a las fibras musculares que trabajaban fue derivado de la PCr después de la suplementación con creatina. La PCr parece ser la fuente energética preferida del músculo esquelético para repletar el ATP durante la actividad. Así, a medida que se incrementa la disponibilidad de PCr, la hidrólisis de PCr durante el ejercicio se incrementa, como fue indicado por la correlación positiva entre la concentración inicial de PCr y la hidrólisis de PCr durante la serie de ejercicio 3 (r=0.72, p<0.05). Otros han reportado un incremento en el uso de PCr y una reducción en el lactato muscular durante el ejercicio después de la suplementación con creatina (3). El grupo joven tendió a usar más PCr después de la suplementación con creatina (p=0.1).
La menor resíntesis inicial de PCr observada en los sujetos de mediana edad fue incrementada (p<0.05) a una velocidad similar a aquella del grupo joven después de la suplementación con creatina (Figura 3). Estos resultados apoyan el concepto presentado previamente acerca de que la disponibilidad de creatina muscular puede afectar la actividad de la creatín quinasa durante la recuperación, y consecuentemente, afectar la velocidad de resíntesis de PCr (3, 12, 15). Ha sido reportado que ocurren incrementos similares en la velocidad de resíntesis de PCr después del entrenamiento físico intenso (19), y existen diferencias similares cuando son comparadas poblaciones desentrenadas y entrenadas (14, 25, 31). Sin embargo, el incremento en la velocidad de resíntesis de PCr en estos estudios fue atribuida a las mejoras en la capacidad oxidativa mitocondrial asociada con el entrenamiento, lo cual le permite a la mitocondria suministrar ATP al sistema de la creatín quinasa a una mayor tasa. Asumiendo que la capacidad oxidativa muscular no fue afectada por la suplementación con creatina en el presente estudio, el incremento en la velocidad inicial de resíntesis de PCr observado en el grupo de mediana edad resultó más probablemente de un incremento en la disponibilidad muscular de creatina. Esto resultados sugieren que la disponibilidad de creatina muscular puede influenciar significativamente la velocidad inicial de resíntesis de PCr después del ejercicio en personas sanas de mediana edad, independientemente de la capacidad oxidativa y el pH.
La Tc monoexponencial es una función de la cantidad de PCr hidrolizada y la velocidad de resíntesis de PCr (18, 20, 26, 34). Debido al incremento combinado en la hidrólisis de PCr y la velocidad de resíntesis (p<0.05) en el grupo de mediana edad después de la suplementación con creatina, la Tc permaneció relativamente constante (Tabla 2). Similarmente, la falta de una diferencia significativa entre la Tc media de los sujetos jóvenes y de mediana edad durante la prueba placebo (Figura 3) puede ser explicada por la menor hidrólisis de PCr (p=0.06) y la menor velocidad de resíntesis de PCr (p<0.05) observada en el grupo de mediana edad (Tabla 2). Estos resultados sugieren que la Tc sola no puede describir adecuadamente las diferencias en el metabolismo energético muscular que pueden existir entre los grupos experimentales y las condiciones.
La velocidad inicial rápida de resíntesis de PCr en los grupos jóvenes y mediana edad parece continuar durante un mayor tiempo, después de la suplementación con creatina (Figura 4). Esto mantiene un nivel de PCr elevado durante la recuperación (Figura 2), a pesar de la hidrólisis incrementada de PCr durante el ejercicio observada después de la suplementación con creatina en el grupo de mediana edad (Tabla 2). La alta velocidad prolongada de resíntesis de PCr observada en este estudio concuerda con los resultados reportados por Greenhaff et al. (12) que muestran una velocidad inicial de resíntesis sostenida en sujetos que respondieron a la suplementación con creatina durante el segundo minuto de recuperación a partir del ejercicio. Han sido propuestos un incremento en la disponibilidad y utilización de PCr durante el ejercicio y un incremento en la capacidad de resíntesis de PCr durante la recuperación como los medios a través de los cuales la suplementación con creatina mejora el rendimiento muscular durante series de ejercicio intermitente (3, 12, 15).
El tiempo hasta el agotamiento en la serie 3 de la prueba de ejercicio se incremento después de la suplementación con creatina en los grupos jóvenes y de mediana edad combinados (Tabla 2), indicando que la suplementación con creatina mejora la resistencia a la fatiga. Esto es consistente con los resultados de varios otros estudios de creatina-rendimiento que implicaron ejercicio intermitente de alta intensidad (2-4, 10, 13). Pocos estudios han reportado que la suplementación con creatina no afecto significativamente el rendimiento del ejercicio intermitente (9, 11). Es necesario un mayor tamaño de muestra para determinar si hay una diferencia en la magnitud del cambio en el tiempo hasta el agotamiento entre personas jóvenes y de mediana edad. En pruebas de control realizadas por dos sujetos jóvenes, no hubo diferencias en el tiempo de ejercicio hasta el agotamiento, 99±27 y 101±37 s, respectivamente, o mediciones de 31P-MRS entre las pruebas, sugiriendo que el tiempo hasta el agotamiento fue una medición confiable del rendimiento muscular.
Conclusión
Ha sido reportado en estudios previos (8, 21-24, 29) que la concentración de PCr en reposo y la velocidad de resíntesis de PCr están reducidas en las personas ancianas. Nosotros hipotetizamos que la suplementación con creatina incrementaría la disponibilidad de PCr muscular y la velocidad de resíntesis en personas de mediana edad más que en personas jóvenes con una actividad y hábitos alimenticios similares. Los resultados de este estudio indicaron que las personas de mediana edad tuvieron mayores incrementos en la disponibilidad de PCr, hidrólisis de PCr durante el ejercicio, y velocidad inicial de resíntesis de PCr, después de la suplementación con creatina, en comparación con las personas más jóvenes, y que la suplementación con creatina mejoró la capacidad de resistencia muscular en ambos grupos combinados. Nuestros resultados no indicaron que la suplementación con creatina mejoró la resistencia muscular en una mayor magnitud en los individuos de mayor edad vs. los más jóvenes.
Agradecimientos
Esta investigación fue apoyada en parte por una Beca de investigación Dudley Allen Sargent de la Sargeant Collage of Health y Rehabilitation Sciences, Boston University. R. A. Fieldieng es un miembro nacional Brookdale en la Boston University. Los enfoques, opiniones y/o hallazgos que contiene este reporte pertenecen a los autores y no deberían ser interpretados como una posición, política o decisión oficial de Departamento del Ejército
Notas al Pie de Página
Dirección para Correspondencia
R. A. Fielding, Boston Univ., Sargent College of Health and Rehabilitation Sciences, 4th floor, 635 Commonwealth Ave., Boston, MA 02215 (correo electrónico: fielding@bu.edu).
Referencias
1. Arnold, D. L., P. M. Matthews, and G. K. Radda (1984). Metabolic recovery after exercise and the assessment of mitochondrial function in vivo in human skeletal muscle of 31P-NMR. Magn. Reson. Med. 1: 307-315
2. Balsom, P. D., B. Ekblom, K. Soderlund, B. Sjodin, and B. Ekblom (1993). Creatine supplementation and dynamic high-intensity intermittent exercise. Scand. J. Med. Sci. Sports 3: 143-149
3. Balsom, P. D., K. Soderlund, B. Sjodin, and B. Ekblom (1995). Skeletal muscle metabolism during short duration high-intensity exercise: influence of creatine supplementation. Acta Physiol. Scand. 154: 303-310
4. Birch, R., D. Noble, and P. L. Greenhaff (1994). The influence of dietary creatine supplementation on performance during repeated bouts of maximal isokinetic cycling in man. Eur. J. Appl. Physiol. 69: 268-270
5. Brooks, S. V., and J. A. Faukner (1994). Skeletal muscle weakness in old age: underlying mechanisms. Med. Sci. Sports Exerc. 26: 432-439
6. Cady, E. B., D. A. Jones, J. Lynn, and D. J. Newham (1989). Changes in force and intracellular metabolites during fatigue of human skeletal muscle. J. Physiol. (Lond.) 418: 311-325
7. Coggan, A. R., R. J. Spina, D. S. King, M. A. Rogers, M. Brown, P. M. Nemeth, and J. O. Holloszy (1992). Histochemical and enzymatic comparison of the gastrocnemius muscle of young and elderly men and women. J. Gerontol. B Psychol. Sci. Soc. Sci. 47: B71-B76
8. Conley, K. E., M. E. Cress, S. A. Jubrias, P. C. Esselman, and I. R. Odderson (1995). From muscle properties to human performance, using magnetic resonance. J. Gerontol. A Biol. Sci. Med. Sci. 50A: 35-40
9. Cooke, W. H., P. W. Grandjean, and W. S. Barnes (1995). Effect of oral creatine supplementation on power output and fatigue during bicycle ergometry. J. Appl. Physiol. 78: 670-673
10. Earnest, C. P., P. G. Snell, R. Rodriguez, A. L. Almada, and T. L. Mitchell (1995). The effect of creatine monohydrate ingestion on anaerobic power indices, muscle strength and body composition. Acta Physiol. Scand. 153: 207-209
11. Febbraio, M. A., T. R. Flanagan, R. J. Snow, S. Zhao, and M. F Carey (1995). Effect of creatine supplementation on intramuscular TCr, metabolism and performance during intermittent, superamaximal exercise in humans. Acta Physiol. Scand. 155: 387-395
12. Greenhaff, P. L., K. Bodin, K. Soderlund, and E. Hultman (1994). Effect of oral creatine supplementation on skeletal muscle phosphocreatine resynthesis. Am. J. Physiol. 266 (Endocrinol. Metab. 29): E725-E730
13. Greenhaff, P. L., A. Casey, A. H. Short, R. C. Harris, K. Soderlund, and E. Hultman (1993). Influence of oral creatine supplementation on muscle torque during repeated bouts of maximal voluntary exercise in man. Clin. Sci. (Colch.) 84: 565-571
14. Guthrie, B. M., S. P. Frostick, J. Goodman, D. J. Mikulis, M. J. Plyley, and K. W. Marshall (1996). Endurance-trained and untrained skeletal muscle bioenergetics observed with magnetic resonance spectroscopy. J. Appl. Physiol. 21: 251-263
15. Harris, R. C., K. Soderlund, and E. Hultman (1992). Elevation of creatine in resting and exercised muscle of normal subjects by creatine supplementation. Clin. Sci. (Colch.) 83: 367-374
16. Hultman, E., J. Bergstrom, and N. M. Anderson (1967). Breakdown and resynthesis of phosphorylcreatine and adenosine triphosphate in connection with muscular work in man. Scand. J. Clin. Lab. Invest. 19: 56-66
17. Hultman, E., K. Soderlund, J. Timmons, G (1996). Cederblad, and P. Greenhaff. Muscle creatine loading in men. J. Appl. Physiol. 81: 232-237
18. Kemp, G. J., D. J. Taylor, and G. K. Radda (1993). Control of phosphocreatine resynthesis during recovery from exercise in human skeletal muscle. NMR Biomed. 6: 66-72
19. Kent-Braun, J. A., K. K. McCully, and B. Chance (1990). Metabolic effects of training in humans: a 31P-MRS study. J. Appl. Physiol. 69: 1165-1170
20. Mahler, M (1985). First-order kinetics of muscle oxygen consumption, and an equivalent proportionality between QO2 and phosphorylcreatine level. J. Gen. Physiol. 86: 135-164
21. McCully, K. K., R. A. Fielding, W. J. Evans, J. S. Leigh, and J. D. Posner (1993). The relationship between in vivo and in vitro measurements of metabolism in young and old human calf muscle. J. Appl. Physiol. 75: 813-819
22. McCully, K. K., M. A. Forciea, L. M. Hack, E. Donlon, R. W. Wheatley, C. A. Oatis, T. Goldberg, and B. Chance (1991). Muscle metabolism in older subjects using 31P magnetic resonance spectroscopy. Can. J. Physiol. Pharmacol. 69: 576-580
23. McCully, K. K., H. Kakihira, K. Vandenborne, and J. Kent-Braun (1991). Noninvasive measurements of activity-induced changes in muscle metabolism. J. Biomech. 24: 153-161
24. McCully, K., and J. Posner (1992). Measuring exercise-induced adaptations and injury with magnetic resonance spectroscopy. Int. J. Sports Med. 13: S147-S149
25. McCully, K. K., K. Vandenborne, K. DeMeirleir, J. D. Posner, and J. S. Leigh (1992). Muscle metabolism in track athletes, using 31P magnetic resonance spectroscopy. Can. J. Physiol. Pharmacol. 70: 1353-1359
26. Meyer, R. A (1988). A linear model of muscle respiration explains monoexponential phosphocreatine changes. Am. J. Physiol. 254 (Cell Physiol. 23): C548-C553
27. Meyer, R. A., M. J. Kushmerick, and T. R. Brown (1982). Application of 31P-NMR spectroscopy to the study of striated muscle metabolism. Am. J. Physiol. 242 (Cell Physiol. 11): C1-C11
28. Meyer, R. A., H. L. Sweeney, and M. J. Kushmerick (1984). A simple analysis of the "phosphocreatine shuttle.". Am. J. Physiol. 246 (Cell Physiol. 15): C365-C377
29. Moller, P., J. Bergstrom, P. Furst, and K. Hellstrom (1980). Effect of aging on energy-rich phosphagens in human skeletal muscles. Clin. Sci. (Colch.) 58: 553-555
30. Paffenbarger, R. S., A. L. Wing, and R. T. Hyde (1978). Physical activity as an index of heart attack risk in college alumni. Am. J. Epidemiol. 108: 161-175
31. Takahashi, H., M. Inaki, K. Fujimoto, S. Katsuta, I. Anno, M. Niitsu, and Y. Itai (1995). Control of the rate of phosphocreatine resynthesis after exercise in trained and untrained human quadriceps muscles. Eur. J. Appl. Physiol. 71: 396-404
32. Taylor, D. J., M. Crowe, P. J. Bore, P. Styles, D. L. Arnold, and G. K. Radda (1984). Examination of the energetics of aging skeletal muscle using nuclear magnetic resonance. Gerontology 30: 2-7
33. Taylor, D. J., P. Styles, P. M. Mathews, D. L. Arnold, D. G. Gadian, P. Bore, and G. K. Radda (1986). Energetics of human muscle: exercise-induced ATP depletion. Magn. Reson. Med. 3: 44-54
34. Thompson, C. H., G. J. Kemp, A. L. Sanderson, and G. K. Radda (1995). Skeletal muscle mitochondrial function studied by kinetic analysis of postexercise phosphocreatine resynthesis. J. Appl. Physiol. 78: 2131-2139
35. Trappe, S. W., D. L. Costill, M. D. Vukovich, J. Jones, and T. Melham (1996). Aging among elite distance runners: a 22-yr longitudinal study. J. Appl. Physiol. 80: 285-290
36. Winter, D. A (1979). Biomechanics of Human Movement. New York: Wiley, p. 151