
Acidosis Metabólica Inducida por el Ejercicio: ¿De donde vienen los Protones?
Exercise Physiology Laboratories, Exercise Science Program, Department of Phsysical Performance and Development, The University of New Mexico, Albuquerque, New Mexico 87131.
Artículo publicado en el journal PubliCE, Volumen 0 del año 2001.
Publicado 27 de enero de 2003
Resumen
Palabras clave: lactato, ácido láctico, glucólisis, ATP, hidrólisis
INTRODUCCIÓN
El método científico envuelve criterios rigurosos para la evaluación del conocimiento, pero el método no es perfecto. Los hallazgos de las investigaciones y sus interpretaciones, pueden ser llevados prematuramente al estado de verdad. Algunas de estas “verdades”, pueden convertirse en un componente principal de una base de conocimientos. Y así ser llamados principios. Consecuentemente, una continua re-evaluación de los contenidos de cualquier disciplina académica es esencial para asegurar que el conocimiento y la práctica estén basados en hechos.
En años recientes, yo he cuestionado un principio que ha sido aceptado por un amplio rango de entidades académicas, de investigación y profesionales; el mismo afirma que la concentración aumentada de protones libres dentro de un músculo esquelético en contracción, es causada por la producción incrementada de ácido láctico. Uno solo tiene que leer cualquier libro de texto de fisiología del ejercicio o de bioquímica pura, para ser informado que, cuando el “ácido pirúvico” es convertido en “ácido láctico”, el pk del “ácido láctico” resulta en una inmediata y casi completa disociación del protón del carboxilo del grupo funcional ácido. Esta interpretación resulta en la creencia lógica que el resultado neto in vivo, es la producción de iones lactato y la liberación de un protón. Una ecuación química general usada para apoyar esta explicación es la siguiente:
Ácido pirúvico + NADH + H+ ↔ ácido láctico + NAD+ → lactato- Na+ + NAD+ + H+
Esta ecuación esta típicamente desarrollada para ilustrar la amortiguación de un protón del lactato, por parte del bicarbonato, resultando en la producción no-metabólica de dióxido de carbono (Brooks et al., 2000).
lactato- H + Na+ → Na+-lactato- + H+
H+ + HCO3- ↔ H2CO3 ↔ H2O + CO2
La fisiología es entonces desplegada para proveer una asociación causa-efecto entre la producción de lactato, el desarrollo de la acidosis, los H+ libres añadidos y la estimulación de la ventilación por el CO2, y el alineamiento temporal del lactato y los umbrales ventilatorios.
Las interpretaciones fisiológicas y bioquímicas anteriores, de una acidosis dependiente del lactato durante el ejercicio, están tan inculcadas, que cientos de artículos científicos se publican cada año, refiriéndose a estas interpretaciones, directa o indirectamente. El error del principio de la “acidosis láctica” en bioquímica y en fisiología radica en que el mismo, no esta basado en hechos. La acidosis se origina en un sitio destinto de la reacción de la lactato dehidrogenasa (LDH).
La Bioquímica de la Acidosis Metabólica
Antes de comenzar con mi explicación bioquímica del desarrollo de la acidosis durante el ejercicio, debo destacar que los conceptos y explicaciones no son nuevos. El crédito es otorgado a Gevers (1977), por su publicación inicial y por su respuesta (Gevers, 1979) a las criticas (Wilkie, 1979) por sus enfoques alternativos y explicaciones acerca de la acidosis metabólica en el músculo cardíaco. Las revisiones y comentarios subsecuentes de la bioquímica de la acidosis metabólica han sostenido las opiniones de Gevers. Por ejemplo, Vaghy (1979), presento evidencia de la incorporación de protones citosólicos (iones hidrógeno libres en el citoplasma) dentro de la respiración mitocondrial en el músculo cardíaco, y el teorizó, que cualquier déficit en la respiración mitocondrial podría contribuir a la acidosis. Dennos fue co-autor de un manuscrito con Gevers, 14 años más tarde (Dennos et al., 1991), que explico la importancia de la hidrólisis del ATP, para la producción y acumulación de protones citosólicos. Similarmente, otros investigadores han cuestionado el concepto de “acidosis láctica” y han propuesto una combinación de la glucólisis y la hidrólisis del ATP, para ser las causas bioquímicas de la liberación y acumulación de protones (Busa y Nuccitelli, 1984; Hochachka y Mommsen, 1983; Noakes, 1977; Zilva, 1978).
Han pasado casi 25 años desde la primera publicación original de Gevers (1977), y no hay evidencia en libros de texto, del reconocimiento de que la producción de lactato no causa la acidosis. El ácido láctico como causa de la acidosis, la llamada “acidosis láctica”, es todavía enseñada en cursos de fisiología y bioquímica a lo largo de todo el mundo. Investigadores en prestigiosas revistas científicas internacionales todavía están usando terminología, como “ácido láctico” y “acidosis láctica” (Ej., Haberg, 1985; Juel, 1996, 1998; Katz y Sahlin, 1988; Stringer et al., 1994). Claramente, un tópico de esta importancia, en fisiología básica y aplicada, así como en medicina clínica, debe estar basado en hechos y no en una teoría no comprobada. Es largamente requerida una re-evaluación de la bioquímica de la acidosis metabólica inducida por el ejercicio.
Fundamentos de la Fisiología Ácido-Base
Antes de explicar las interpretaciones actuales y propuestas sobre la bioquímica de la acidosis metabólica, voy a clarificar la diferencia entre un ácido y una sal ácida. Un ácido es una molécula que a un ph neutro, va a liberar un protón a la solución. Dependiendo del tamaño de la molécula, el protón viene de un tipo de estructura química especial en la molécula, típicamente llamado grupo funcional. Las grandes moléculas de ácidos pueden tener más de un grupo funcional ácido, tal como los aminoácidos. Algunas moléculas ácidas son demasiado pequeñas para contener grupos funcionales ácidos, pero son todavía ácidos (Ej., ácido clorhídrico, HCl; ácido perclórico, HClO4; ácido fosfórico, H3PO4). La Fig. 1 presenta dos ejemplos de grupos funcionales ácidos dentro del metabolismo celular: los grupos carboxilo y fosforilo. Se ha teorizado, dentro del principio de la “acidosis láctica”, que el grupo carboxilo cumple el mayor rol en la acidosis metabólica celular.
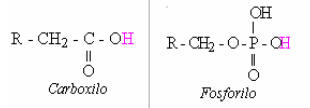
Figura 1: Una ilustración estructural de los dos grupos funcionales ácidos
principales dentro del metabolismo celular. Las estructuras están ilustradas
en su forma descargada (no ionizada). El protón liberado a la solución es
mostrado en color rosa (H).
La fuerza de un ácido esta relacionada a la tendencia de la molécula a liberar un protón en una solución, aún cuando la solución es ya ácida (ph inferior a 7). Así, los ácidos fuertes van a liberar un protón hasta que un ph relativamente bajo sea alcanzado, tiempo en el cual hay un equilibrio dinámico entre los protones que dejan y los que se vuelven a unir al grupo funcional ácido de la molécula.
Consecuentemente, para entender mejor el potencial de un ácido de liberar protones, es necesario saber a que ph alcanza la liberación de protones, el equilibrio dinámico. Este ph es definido como el logaritmo negativo de la constante de ionización, abreviada como pk’.
En equilibrio; HA ↔ H+ + A-, donde
K = productos / sustratos = ([H+] [A-]) / [HA]
pK' = -log K = log(1/K)
El pk’, que representa el ph en el cual la mitad de las moléculas ácidas están deprotonadas (ionizadas), puede ser determinado in vitro por titulación. Como se debe ser capaz de predecir, los ácidos fuertes o grupos funcionales ácidos fuertes, tiene un pk’ mucho más bajo de 7, y los ácidos débiles, tienen valores de pk’ cercanos a 7.0. En la Tabla 1 son presentados los valores de pk’ de una selección de ácidos y grupos funcionales ácidos.


Tabla 1: Valores de pk’ de ácidos específicos o de sus grupos funcionales. (a
25°C).
Después de que una molécula ácida pierde un protón, alcanza una carga iónica negativa. Para mantener la neutralidad de cargas, un catión se une ionicamente a la carga negativa, resultando esto en una sal ácida.
Debido a la abundancia intracelular y extracelular de sodio (Na+) y potasio (K+), ambos son individualmente cationes cargados, los ácidos deprotonados son predominantemente sales de sodio y potasio. Hay que notar que en la Tabla 1, el pk’ del ácido láctico es reportado en 3.86. Por lo tanto, la forma principal del “ácido láctico” en los sistemas fisiológicos es la de lactato sódico (La-Na+).
Finalmente, debe ser enfatizado que la producción de ácido no es el único origen de liberación de protones dentro de una célula. Los protones también pueden ser liberados a partir de reacciones químicas, y voy a demostrar que este origen de protones es la principal causa de acidosis en los músculos esqueléticos en contracción. Además, Stewart (1983) ha indicado claramente que el movimiento de iones cargados a través de la membrana celular muscular puede influenciar el balance ácido base celular, y este enfoque del entendimiento del balance ácido base ha sido llamado “la fuerte diferencia iónica”. Investigaciones adicionales sobre “la fuerte diferencia iónica” han demostrado que está asociada con la contribución a la acumulación de protones dentro de las células musculares en contracción, presumiblemente debido al flujo de potasio desde el músculo durante el ejercicio intenso (Lindinger y Heigenhauser, 1991). En este manuscrito, yo me focalize en la liberación y consumo de protones, y no voy a considerar la influencia adicional de la fuerte diferencia iónica sobre la cinética de los protones pre-existentes.
El Origen de los Protones durante el catabolismo en el Músculo Esquelético
En la sección que sigue, voy a explicar las reacciones citosólicas del catabolismo energético. Voy a comenzar con las reacciones del sistema energético de los fosfatos, y luego las reacciones de la glucólisis, terminando con la reacción de la LDH. Para todas las reacciones que involucran, ya sea el consumo o la liberación de protones, proporciono estructuras para ilustrar el intercambio de átomos, electrones y protones. Estas ecuaciones balanceadas atómicamente, no son dadas en los libros de texto de bioquímica o fisiología del ejercicio, lo que puede explicar, porque la bioquímica de la acidosis metabólica es tan pobremente entendida!
Sistema Energético Fosfágeno: Reacción de la Creatín Quinasa
La reacción de la creatín quinasa (CK) es de vital importancia para la contracción del músculo esquelético. Esta reacción proporciona el medio más inmediato para reponer ATP en el citosol. Tradicionalmente, la reacción ha sido interpretada para ser aplicable principalmente a las necesidades metabólicas del ejercicio intenso, la transición a las intensidades en aumento, o durante condiciones de hipoxia. Sin embargo, la fosfocreatina (CrP) probablemente ayude en la transferencia de fosfatos terminales en todo el citosol, así como también desde la mitocondria al citosol. Esta función es resumida como las reacciones de vínculo de la “lanzadera de fosfato de creatina” (Karlsson, 1971; Kent-Braun et al., 1993). La ecuación química de la reacción de la CK es la siguiente:
Fosfocreatina + ADP + H+ ↔ Creatina + ATP
In vivo, la reacción de la CK es realmente una reacción acoplada, que envuelve la ruptura de CrP y la fosforilación de ADP. Es incorrecto referirse a esta reacción in vivo como hidrólisis de la CrP. La hidrólisis de la CrP puede ocurrir in vitro, donde es requerida el agua para proveer los átomos y electrones necesarios para producir creatina, fosfato inorgánico (Pi), y un protón.
La reacción de la CK, es calificada como una reacción de equilibrio, puesto que in vivo el cambio de energía libre (∆G) se aproxima a cero. Así, cuando el producto de las moléculas del lado izquierdo de la ecuación se incrementa en forma relativa al lado derecho de la ecuación, como durante el ejercicio de intensidad incremental, la reacción se convierte en exergónica en la dirección de la regeneración del ATP. La reacción revierte su dirección durante la recuperación luego del ejercicio.
Los componentes estructurales de la reacción de la creatín quinasa están detallados en la Fig. 2. La reacción implica la transferencia de un fosfato desde la CrP al ADP para formar ATP. Durante el ejercicio, la velocidad incrementada de la reacción de la CK causa realmente una pequeña alcalinización del músculo esquelético debido al consumo de un protón durante la reacción (Karlsson, 1971; Dennis, 1991; Gevers, 1977). Con el objeto de reformar el amino terminal de la creatina, es consumido un protón de la solución durante la reacción, explicando así la alcalinización. El grupo carboxilo de la creatina (Cr) esta todavía ionizado a ph fisiológico (Tabla 1) y no contribuye a la alcalinización.
La bioquímica de la reacción de la CK indica que 1 protón es consumido por cada fosfato transferido desde la CrP al ADP, formando ATP. Así, la reacción de la CK, funciona como un pequeño “consumidor” de protones, con una capacidad inmediata durante el ejercicio, igual al número de moléculas de CrP que transfieren sus fosfatos al ADP.
Sistema Energético Fosfágeno: Reacción de la Adenilato Quinasa
En los ejercicios de intensidad incremental que no se desarrollan en condiciones de estado-estable, no solo aumenta la actividad de la reacción de la CK, también se incrementa la segunda reacción del sistema fosfágeno, la reacción de la adenilato quinasa (AK) (o mioquinasa). La ecuación química de la reacción de la AK, es la siguiente:
ADP + ADP ↔ ATP + AMP
La producción de AMP es importante. El AMP incrementa la actividad de la fosforilasa, por eso aumenta la glucogenólisis, así como estimula la actividad de la fosfofructo quinasa. El resultado de esta estimulación es una velocidad incrementada de la formación de glucosa-6-fosfato para alimentar la glucólisis y una velocidad incrementada del flujo glucolítico. Como vamos a discutir, este flujo incrementado aumenta, a través de la glucólisis, la liberación de protones y eventualmente disminuye el ph celular.
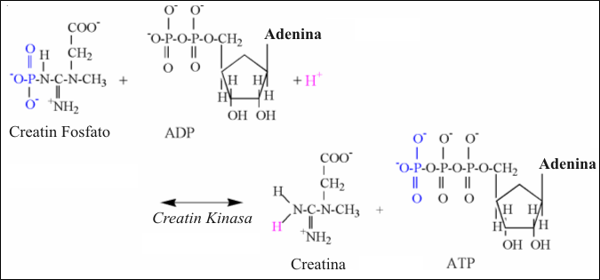
Figura 2: Ilustración estructural de la reacción de la creatín quinasa.
Sistema Energético Fosfágeno: Reacción de la AMP deaminasa
La actividad de la reacción de la adenilato quinasa es mejor detectada por los incrementos de los adenosinmonofosfatos (AMP) y los inosinmonofosfatos (IMP) en el músculo. La producción de IMP resulta de una actividad incrementada de la reacción de la AMP deaminasa, la que es activada por la acidosis y produce IMP y amonio (NH4);
AMP + H+ ↔ IMP + NH4
La reacción consume un protón debido a la formación inicial de NH3 (Fig.3). El alto pk’ del amoníaco resulta en la adición de un protón.
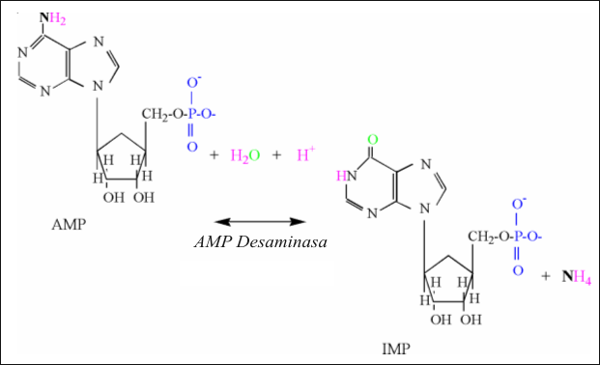
Figura 3: Ilustración estructural de la reacción de la AMP desaminasa.
La suma de la concentración incrementada de ADP, a las de AMP e IMP producidas en el músculo (∆ ADP + AMP + IMP) justifican la pequeña disminución de ATP experimentada durante el ejercicio intenso hasta la fatiga. Es importante reconocer que las reacciones de la AK y la AMP deaminasa, reflejan una inhabilidad de la respiración mitocondrial en el abastecimiento total de ATP dentro del citosol de la célula. Las investigaciones indican que estas condiciones celulares están asociadas con las mayores regeneraciones de ATP por parte del sistema fosfágeno y la glucólisis, y coinciden con un rápido incremento del lactato y acumulación de protones (disminución del ph) (Karlsson, 1971; Sahlin, 1978; Sahlin et al., 1987; Katz y Sahlin, 1988).
Sistema Energético Fosfágeno: Hidrólisis del ATP
La contracción muscular necesita de la ruptura (hidrólisis) de ATP a ADP y Pi (HPO4-2). La enzima que cataliza esta reacción es la miosín ATPasa, y la ecuación química es la siguiente:
ATP + H2O ↔ ADP + Pi + H+
La liberación de protones asociada a esta reacción resulta de la participación del agua, la que es necesaria para proveer un átomo de oxígeno para que se una al fosfato terminal del ADP y un grupo hidroxilo que se une al Pi (Fig.4). Un protón es así liberado en condiciones de ph fisiológico, ya que el pk’ del átomo de oxígeno que queda en el grupo fosfato es muy bajo para ser protonado (Tabla 1).
La hidrólisis del ATP es el principal estímulo para el catabolismo energético incrementado, durante la contracción muscular. La función principal del catabolismo energético parece ser el mantenimiento de la concentración celular de ATP. Al principio del ejercicio de intensidad moderada, el sistema fosfágeno y la regeneración glucolítica del ATP mantienen el ATP celular, hasta que la respiración mitocondrial sea adecuadamente estimulada.
Los productos de la hidrólisis del ATP pueden ser todos usados por la célula en condiciones de estado-estable. El ADP citosólico esta envuelto en la transferencia de grupos fosfatos desde el ATP mitocondrial hasta la Cr citosólica, y en la re-formación de ATP, como fue descrito en la sección de la reacción de la CK. El ADP es también directamente transportado dentro de la mitocondria como sustrato para la fosforilación oxidativa. El Pi es usado como sustrato en la glucogenólisis (reacción de la fosforilasa) y en la reacción de la gliceraldehido-3-fosfato dehidrogenasa de la glucólisis. Además, el Pi también puede ser transportado dentro de la mitocondria, donde es necesario como sustrato para la fosforilación oxidativa. Los protones de la hidrólisis del ATP, también pueden ser transportados dentro de la mitocondria a través de las lanzaderas, malato-aspartato y glicero-fosfato, o por trasporte directo a través de los transportadores de protones (Ej., el transportador proton-lactato monocarboxilato). Los protones luego ayudan en el mantenimiento del gradiente de protones entre el espacio de la membrana mitocondrial interna y la matriz.
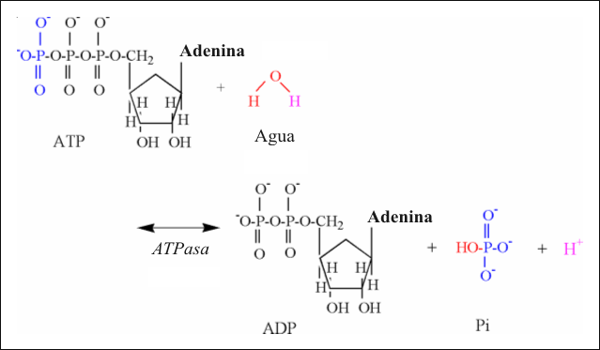
Figura 4: Ilustración estructural de la hidrólisis del ATP.
Cuando la velocidad de hidrólisis del ATP excede la velocidad a la cual la mitocondria puede remover y/o utilizar los productos de la reacción, los mismos pueden acumularse. El ADP no se acumula en un grado significativo debido a las reacciones de la adenilato quinasa y la creatín quinasa. Sin embargo, el Pi y los protones pueden acumularse, con la ganancia de protones siendo potencialmente mayor que la de Pi, debido al uso del mismo como sustrato en la Fase 2 de la glucólisis, como fue previamente explicado. Consecuentemente, la hidrólisis del ATP puede convertirse en un origen de protones significativo durante ejercicios a intensidades moderadas a intensas, y por ello contribuir al desarrollo de la acidosis.
El fosfato inorgánico libre, no es un ácido fuerte, ya que todos sus átomos de hidrógeno, menos uno se disocian a ph fisiológico, dejando HPO4-2. Interesantemente, el fosfato inorgánico puede funcionar como un amortiguador cuando el ph cae, debido a que el pk’ de uno de los grupos funcionales hidroxilo es de 6.82 (Tabla 1). El potencial amortiguador del Pi, dependiente del ph, es revelado en la espectroscopia por resonancia magnética con fósforo 31 (31P-MRS), por medio de un cambio en el espectro de frecuencia, cuando el Pi es protonado. Este cambio es usado para calcular el ph citosólico, usando una ecuación de Henderson-Hasselbach modificada (Kent-Braun et al., 1993).
Sistema Energético Fosfágeno: Resumen
Durante el ejercicio de intensidad incremental, que no se realiza en estado-estable, se incrementa la actividad de la reacción de la CK. Esta reacción disminuye la CrP, consumiendo al mismo tiempo, un protón. La reacción de la CK junto con de la AK, logran mantener bien las concentraciones celulares de ATP, a pesar de la inadecuada velocidad de regeneración de ATP, por parte de la respiración mitocondrial.
Estas condiciones celulares están también asociadas con un incremento del Pi. Sin embargo, la acumulación de esta molécula, no es un resultado de la reacción de la CK como generalmente es creído dentro de las ciencias del deporte y el ejercicio, sino de los resultados de una defosforilación neta de ATP durante la contracción muscular. La concentración aumentada de Pi celular indica, de este modo, que la célula se esta retrasando en la generación de ATP, a partir de la respiración mitocondrial, ya que el Pi, no es rehusado en la glucólisis o transportado dentro de la mitocondria como sustrato para la fosforilación oxidativa.


Tabla 2: Las dos fases de la glucólisis, con la producción total de de ATP y
protones (H+).
El desarrollo de una inhabilidad por parte de la célula para abastecer todas las necesidades de ATP celulares a partir de la respiración mitocondrial, es un proceso gradual y no fácilmente detectable evaluando el ATP, debido a la efectividad de las reacciones de la CK y AK, así como una velocidad incrementada de regeneración de ATP a partir de la glucólisis. No obstante, la hidrólisis del ATP, libera un protón, y cuando esta no es equilibrada por una igual velocidad de regeneración de ATP derivado de la respiración mitocondrial, este protón puede acumularse en el citosol (Kent-Braun et al., 1993). La potencia de la liberación de protones a partir de la hidrólisis del ATP está en proporción directa con la velocidad de producción de ATP. Sin embargo, en una pequeña extensión, el protón producido a partir de la hidrólisis del ATP, es reducido por el consumo de protones, en las reacciones de la CK y AMP deaminasas. Cuando la acidosis aumenta (ph < 6.9), los grupos fosfatos (Pi, hexosa y triosa fosfato) proveen la amortiguación de los protones adicionados.
Glucólisis
Las reacciones de la glucólisis son enumeradas en la Tabla 2. Dos reacciones de la Fase 1 y una de la Fase 2, liberan protones, mientras que una reacción de la Fase 2, consume protones. Consecuentemente, cuando empieza a partir de glucosa o glucógeno, la glucólisis produce un neto de 2 protones, por el flujo de glucosa a dos piruvatos. Cuando el flujo glucolítico se incrementa también lo hace la liberación neta de protones y la necesidad de amortiguación de los mismos.
La reacción de la hexoquinasa es la primer reacción de la glucólisis que libera protones, y es ilustrada en la Fig.5. El grupo hidroxilo del carbono 6, es dividido durante esta reacción, liberando un protón. El oxígeno y el electrón restantes, aceptan el grupo fosfato transferido desde el ATP.
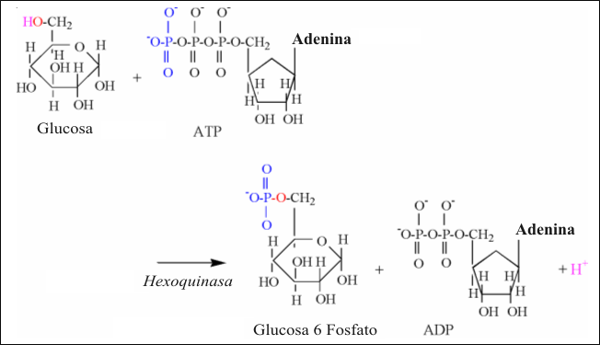
Figura 5: Ilustración estructural de la reacción de la hexoquinasa. Notar los
átomos de colores, unidos a los mecanismos de la reacción.
La segunda reacción de la glucólisis, que libera protones es catalizada por la fosfofructo quinasa (PFK) y es ilustrada en la Fig.6. Como la reacción de la hexoquinasa, el grupo hidroxilo del primer carbono es dividido, liberando un protón, seguido de la aceptación del grupo fosfato transferido desde el ATP.
La gliceraldehido-3-fosfato dehidrogenasa (G3PDH) cataliza la tercera reacción de la glucólisis que libera protones. El grupo aldehído del tercer carbono es oxidado por el NAD+, resultando en la eliminación de dos electrones y un protón. También es eliminado un protón del Pi, permitiendo a este unirse al tercer carbono, formando 1,3 bisfosfoglicerato.
La reacción de la G3PDH esta funcional y bioenergeticamente acoplada a la reacción de la fosfoglicerato quinasa (PGK) (Fig.8). Hay que notar que la reacción de la PGK produce el primer ácido intermediario de la glucólisis, el 3-fosfoglicerato. En una edición anterior de su libro, Lehninger (1993) explicó que esta reacción produce un protón por medio de la ionización del 3-fofoglicerato a ph fisiológico. Sin embargo, esta apreciación no debería ser aplicada a condiciones in-vivo, ya que Lehninger está ilustrando el mecanismo de reacción de la hidrólisis, el cual ocurre in-vitro sin el acoplamiento de la reacción para la fosforilación del ADP.
La reacción de la PGK comprende una simple transferencia de fosfato desde el primer carbono del 1,3 bisfosfoglicerato al ADP, formando ATP. Un oxígeno y un electrón permanecen en el grupo funcional carboxilo del 1,3 bisfosfoglicerato. En esta transferencia no hay ningún protón implicado y el 3-fosfoglicerato es formado desprovisto de un protón.
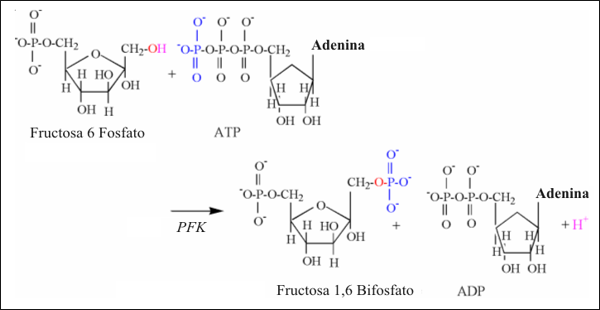
Figura 6: Ilustración estructural de la reacción de la fosfofructo quinasa.
Notar los átomos de colores, unidos a los mecanismos de la reacción.
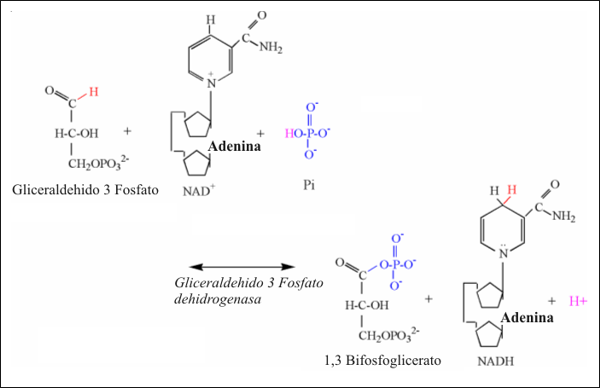
Figura 7: Ilustración estructural de la reacción de la gliceraldehido-3-fofato
dehidrogenasa.
Notar los átomos de colores, unidos a los mecanismos de la reacción.
Este mismo grupo carboxilo permanece deprotonado en los otros intermediarios de la glucólisis. Estos importantes hechos bioquímicos significan que nunca hubo una liberación de protones por el grupo carboxilo del 3-fosfoglicarato o alguno de los intermediarios glucolíticos siguientes. Así, no hay ningún protón asociado con el grupo carboxilo cuando es producido el lactato. La única conclusión que se puede hacer a partir de este hecho bioquímico es que es imposible para la producción de lactato, o para alguno de los intermediarios carboxílicos “corriente abajo” a partir del 3-fofoglicerato, causar la liberación de un protón y la acidosis subsiguiente. Este hecho por si solo, elimina el concepto de una “acidosis láctica”.
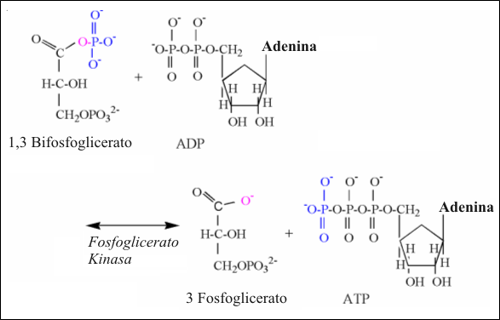
Figura 8: Ilustración estructural de la reacción de la fosfoglicerato quinasa.
Notar los átomos de colores, unidos a los mecanismos de la reacción.
La reacción de la piruvato quinasa consume un protón, y es ilustrada en la Fig. 9. El grupo fosfato unido al segundo carbono del fosfo-enol-piruvato es trasferido al ADP, formando ATP. El estado químico preferido del piruvato es en la forma de cetona (segundo carbono con una unión doble con el oxígeno), un protón de la solución es requerido para unirse al tercer carbono, para completar la estructura química.
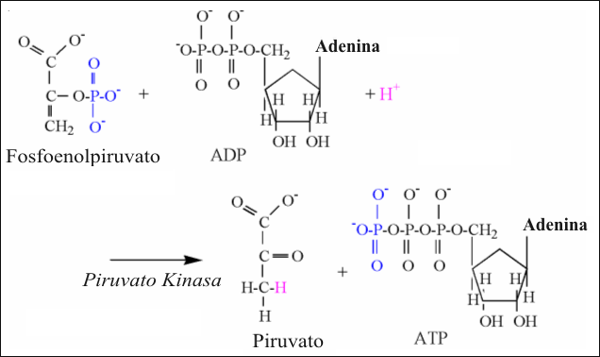
Figura 9: Ilustración estructural de la reacción de la piruvato quinasa. Notar
los átomos de colores, unidos a los mecanismos de la reacción.
La Reacción de la Lactato Dehidrogenasa
Una vez que es producido el piruvato, puede ser transportado dentro de la mitocondria y oxidado por medio de la compleja reacción de la piruvato dehidrogenasa, o puede ser reducido a lactato en el citosol por la LDH. La ecuación química de la reacción de la LDH es la siguiente:
Piruvato + NADH + H+ ↔ lactato + NAD+
La reducción del piruvato implica la adición de 2 electrones y protones del NADH, y 1 protón del citosol (Fig.10). El segundo carbono del piruvato es reducido por medio de la adición de un electrón y un protón del NADH, formando una unión covalente con el hidrógeno. Otro electrón del NADH y un protón, son usados para formar el grupo hidroxilo. Como fue previamente explicado, no hay ningún protón asociado con el grupo carboxilo, y de este modo ninguna liberación de protones e ionización. Así, la reacción de la LDH consume un protón, y por ello funciona como un consumidor de los protones producidos en el catabolismo y en la hidrólisis del ATP.
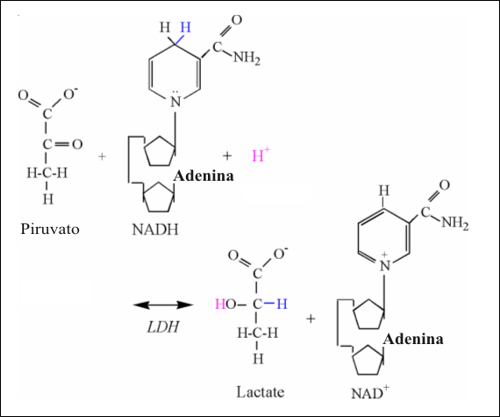
Figura 10: Ilustración estructural de la reacción de la lactato dehidrogenasa.
Notar los átomos de colores, unidos a los mecanismos de la reacción.
Balance de la Producción y Consumo de Protones en la Contracción Muscular
Basándose en la bioquímica metabólica presentada, puede ser hecha una estimación del balance de la producción y consumo de protones (incluyendo la amortiguación). Sin embargo, tal estimación solo representa el intercambio de protones como resultado de las reacciones del sistema fosfágeno y glucolítico, modificados por la oxidación de carbohidratos en la respiración motocondrial.
Un intercambio adicional de protones ocurre durante la oxidación de aminoácidos y las reacciones asociadas de aminación y deaminación. Además, el balance ácido base tisular y sanguíneo es aún más complicado durante la cetosis. No obstante, durante el ejercicio intenso de corta duración hasta la fatiga, es acertado concluir que los sistemas fosfágeno y glucolítico representan la mayor parte del intercambio de protones.
Cuando se considera la producción de protones en el citosol durante la contracción muscular, el origen se encuentra en la glucólisis y en la hidrólisis del ATP. Para el consumo de protones, los contribuyentes son las reacciones de la CK, AMP deaminasa, respiración mitocondrial, Pi y los amortiguadores intracelulares adicionales, y el flujo de protones desde la célula (Tabla 3). Se debe tener en cuenta que a medida que el ph disminuye, el Pi (libre y las hexosas y triosas fosfato) se convierte en un fuerte amortiguador de protones debido a la proporción incrementada de moléculas que consumen un protón de la solución (formando H2PO3-). Por simplicidad, no he basado los cálculos en las contribuciones fraccionales del intercambio de protones. Esta decisión esta basada en investigaciones y revisiones pasadas acerca de este tópico, que revelaron que el magnesio está unido a todos los adenilatos y por ello disminuye los valores de pk hasta niveles ácidos no fisiológicos (Karlsson, 1971). Además, la amortiguación de protones por parte del Pi es mínima, y la representación fraccional de este componente causa un cambio mínimo al intercambio total de protones.
Los datos de la Tabla 3 se refieren al ejercicio intenso hasta la fatiga, y están derivados a partir de las investigaciones de Spriet et al. (1987, 1987) y Melbo et al. (1993). He usado los datos de Melbo et al. basados en 3 minutos de cicloergómetro al 120 % del VO2 máx., para calcular un balance entre la liberación y el consumo de protones. Ya que la contribución glucolítica a la producción de ATP, no fue calculada por Medro, yo estime la misma basándome en un 60 % de contribución glucolítica a la producción total de ATP, como fue recomendado por Spriet (1990).
Cuando es registrada la suma de todos los componentes liberadores y consumidores de protones, quedan ~145 mmol H+/kg/3 min para ser manipulados por los amortiguadores y el transporte extracelular desde la célula. Esto me parece apropiado, ya que las investigaciones han producido valores de amortiguación muscular de entre 40 y 80 mmol H+/L/pH. Ya que esta es una capacidad que se adapta con el entrenamiento, es razonable usar un valor alto de 80 mmol H+/L/pH, que se aproxima a 59 mmol H+/L/pH. Es difícil convertir esto en una capacidad, pero un descenso del ph muscular de 7 hasta 6.4, equivale a 35.4 mmol H+/kg. Consecuentemente, el flujo de protones desde los músculos debe aproximarse a 110 mmol H+/kg/3 min, o a 37 mmol H+/kg/min; un valor que es distribuido entre la remoción pasiva de protones, la amortiguación por el bicarbonato, y los transportadores de protones (Na+, HCO3-, lactato). Desafortunadamente, no son numerosas las investigaciones acerca del flujo de protones desde el músculo humano de tipo de fibras heterogéneas, y es difícil, sino imposible, determinar la validez de los 37 mmol H+/kg/min estimados, que representan el flujo de protones (Brooks, 2000; Juel, 1996, 1998).


Tabla 3. Reacciones predominantes que contribuyen a la liberación y consumo de
protones en el músculo esquelético humano en contracción.
Ya que los protones libres que quedan en solución dentro de una célula no alcanzan una gran concentración (Ej., una disminución del ph desde 7 a 6.4, resulta en una acumulación de protones de 0.3 mmol/L, o aproximadamente 0.22 mmol/kg), el valor de 30 a 40 mmol H+/kg/min para el flujo de protones, es alto. No obstante, los cálculos de balance de protones anteriormente mencionados son más realistas que si la acidosis fuera dependiente de la producción de lactato.
Asumiendo que el 23 % del lactato muscular total es removido durante 3 minutos de ejercicio intenso hasta la fatiga, son producidos aproximadamente 40 mmol/kg lactato/3 min. Cuando se tiene en cuenta las reacciones del metabolismo, que consumen protones adicionales (ignorando que la LDH consume un protón!), la liberación neta de protones a partir del lactato durante 3 minutos de ejercicio extenuante, alcanzaría solo los 17 mmol/kg (40–20–3 = 17 mmol/kg). Basándose en la capacidad amortiguadora muscular (35.4 mmol H+/kg), y las estimaciones anteriores del flujo de protones desde el músculo (110 mmol H+/kg/3 min), este valor para la liberación de protones relacionada al lactato es solo un 15 % del total de protones considerados por la amortiguación y el flujo. Claramente, el concepto de “acidosis láctica” no es apoyado por la bioquímica, o por los datos de producción de lactato, y amortiguación y flujo de protones durante el ejercicio intenso hasta la fatiga.
Resumen del Intercambio Citosólico de Protones
El balance de las reacciones de liberación y consumo de protones del catabolismo en el músculo esquelético, necesita ser presentado de una manera que exprese la actividad simultánea de todas las reacciones pertinentes. Yo he tratado de hacer esto para todas las reacciones citosólicas en la Fig.11, y este resumen ilustrativo del contenido principal de este manuscrito necesita ser aplicado a la capacidad y potencia de liberación y consumo de protones, resumidas en la Tabla 3.
Durante el ejercicio de baja intensidad, el flujo glucolítico es bajo, el sustrato principal del catabolismo energético son los ácidos grasos, y consecuentemente, los requerimientos musculares de ATP, son largamente abastecidos por la respiración mitocondrial. Con un incremento de la intensidad del ejercicio, aumentan el flujo sanguíneo y la captación de glucosa dentro del músculo. Además, el calcio y el Pi libre aumentan poco, por ello se incrementa la actividad de la fosforilasa. La captación de glucosa adicional y la velocidad incrementada de la glucogenólisis incrementan el flujo glucolítico, aún todavía a intensidades de estado-estable, hay también un incremento en la liberación de protones. Sin embargo, los protones son consumidos por la producción de lactato y por su transporte dentro de la mitocondria para ser usados en la fosforilación oxidativa.
A medida que la intensidad del ejercicio aumenta, hasta exceder el punto de umbral de mantención de la demanda de ATP citosólico por parte de la respiración mitocondrial, hay un incremento transitorio del ADP, que causa una velocidad aumentada de la reacción de la CK. Además, el Pi comienza a acumularse, proveyendo un sustrato adicional de la glucogenólisis y la glucólisis, favoreciendo el flujo de sustratos incrementado a través de la glucólisis. Estos eventos llevan a un rápido incremento en la liberación de protones debido a una dependencia incrementada sobre la glucólisis para sostener las concentraciones celulares de ATP. Consecuentemente, la causa principal de una liberación incrementada de protones es la mayor velocidad del flujo glucolótico, más la dependencia incrementada sobre la producción glucolítica del ATP. El flujo de sustratos incrementado a través de la glucólisis, acompañado por las disminuciones en el redox citosólico (NAD+/NADH) resulta en una velocidad de producción de lactato incrementada (Sahlin et al., 1987).
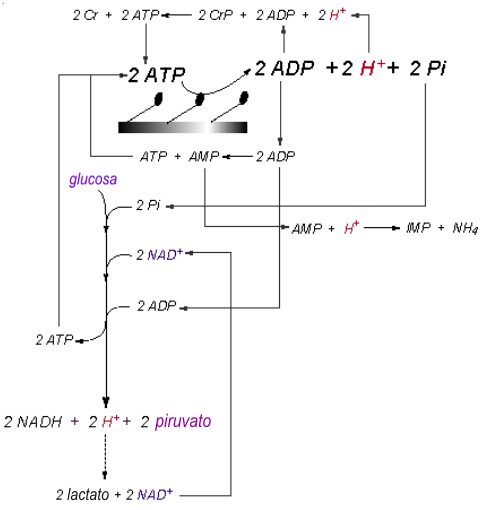
Figura 11: Resumen de los eventos bioquímicos que contribuyen a la acidosis
metabólica. Los protones, de color rojo, son producidos en la glucólisis y en
la hidrólisis del ATP. Cuando la velocidad de producción de ATP por parte de
la respiración mitocondrial cae debajo de la velocidad de demanda del ATP en
el citosol, se pueden acumular ADP, Pi, y AMP. Bajo estas condiciones, y a
pesar de que los protones consumidos por las reacciones de la CK y AMP
deaminasa, los protones liberados en la hidrólisis del ATP y en la glucólisis
(el exceso no consumido por la producción de lactato) contribuya con el
desarrollo de la acidosis celular.
La producción de lactato es beneficiosa para regenerar NAD+, así como también para consumir un protón. No obstante, la capacidad de la reacción de la LDH para mantener el redox citosólico y retardar una acidosis peor, depende de la máxima velocidad de flujo de protones desde la célula. Afortunadamente, el transportador de lactato co-trasporta también a un protón. También existen trasportadores de lactato adicionales (Na+ y HCO3-). Así, la producción de lactato tiene una tercera ventaja: ayuda al flujo de protones desde el músculo. A pesar de estos beneficios, el transportador de protones-lactato es limitado por la velocidad, y a medida que el lactato se acumula en el citosol, la bioenergética de la reacción de la LDH se convierte en menos favorable, y la velocidad de producción de lactato disminuye. Durante el ejercicio continuo a baja intensidad, la velocidad de producción de piruvato y lactato, es también disminuida debido a una reducción de la velocidad de la glucogenólisis y la glucólisis, lo cual ocurre en un período de tiempo tan temprano como 30 segundos a 3 minutos, en el ejercicio intenso. La acumulación de piruvato en el citosol de la célula, y la acumulación de unidades de acetil en la mitocondria, reflejan una actividad glucolítica que no termina en la producción de lactato o en la oxidación completa de los carbonos de la glucosa. Así, se acumulan protones adicionales, y la acidosis es demorada.
Aplicación de la Bioquímica de la Acidosis a la Fisiología del Ejercicio
Claramente, no hay ninguna evidencia bioquímica, que haya comprobado que la producción de lactato libera un protón y causa acidosis. Tampoco hay evidencia alguna de que la producción de lactato se incremente en iguales cantidades que el número de protones liberados dentro y desde el músculo esquelético. Consecuentemente, debería enseñarse que la causa de la acidosis es un resultado de las intensidades del ejercicio, el cual no es realizado en estado estable.
Tales condiciones resultan en un incremento posterior de la velocidad de la glucólisis, y en una dependencia incrementada sobre la producción de ATP citosólico, debido a un desequilibrio entre el índice de demanda de ATP (contracción muscular) y el aporte por parte de la respiración mitocondrial.
Estas condiciones celulares tienen grandes implicaciones sobre como entendemos la fisiología del ejercicio. Por ejemplo, la producción de lactato retarda, no empeora la acidosis. Una mayor capacidad para producir y remover el lactato de la célula, demoraría el comienzo de la acidosis. Esto significa que durante el ejercicio intenso, una alta producción de lactato es beneficiosa para el atleta, especialmente cuando es acompañada por una alta capacidad de transporte de lactato y protones desde la célula, capacidades que se sabe, se incrementa con ambos, entrenamiento de resistencia y de fuerza y velocidad (Juel, 1998).
El alineamiento temporal entre las condiciones celulares que conducen a la acidosis y la producción de lactato, no cambian por esta explicación bioquímica de la acidosis. El lactato es obviamente un buen marcador indirecto de una alteración en el metabolismo celular que causa acidosis y de un entorno metabólico celular en un estado no estable. Sin embargo, la producción de lactato no causa la acidosis.
Otra aplicación de la bioquímica de la acidosis metabólica, esta relacionada al reclutamiento de unidades motoras rápidas. A medida que la intensidad del ejercicio se incrementa, las unidades motoras tipo IIa y IIb son reclutadas progresivamente. Ya que las fibras musculares de estas unidades motoras tienen una densidad mitocondrial más baja que las fibras tipo I, son más dependientes de la glucólisis y de la producción citosólica de ATP. Ya que estas dos características se combinan para incrementar la velocidad neta de liberación de protones por el metabolismo, durante el ejercicio de intensidades asociadas a un reclutamiento incrementado de unidades motoras Tipo II, se origina una considerable producción de protones. Las fibras tipo II contribuyen a la acidosis, no porque producen más lactato, sino debido a que tienen menos mitocondrias para ayudar a la regeneración de ATP, y a la captación de protones. La bioquímica de la acidosis, también tiene implicaciones clínicas. Obviamente, tratar de prevenir la acidosis inhibiendo la producción de lactato va a empeorar, no a prevenir la acidosis. Las mejores maneras de prevenir o retardar la acidosis es, disminuir la dependencia sobre la glucólisis, aumentar la contribución de la respiración mitocondrial a la producción de ATP, y aumentar la capacidad de amortiguación de protones y de remoción de lactato-protones.
Las estrategias anteriores son típicas del entrenamiento de resistencia, y las siguientes son aplicables al entrenamiento de fuerza y potencia. Sin embargo, las estrategias clínicas se relacionarían a incrementar la oxidación de lípidos, aumentando los ácidos grasos libres plasmáticos, o estimulando la función mitocondrial.
¿Porque Todavía se Piensa que el Lactato causa la Acidosis?
A pesar de las realidades bioquímicas que he presentado, el hecho es que la mayoría de los académicos e investigadores dentro del campo puro y aplicado de la fisiología y bioquímica todavía piensan que la producción de lactato causa la acidosis.
Una gran explicación de este hecho es que las secciones de bioquímica de los libros de texto, no presentan ecuaciones químicas balanceadas para los protones y el agua. Así, tuve que aplicar mis conocimientos de química orgánica para derivar los diagramas de las reacciones químicas presentadas en este artículo. La mayoría de los Doctores (Ph.D’s) y Médicos, simplemente no han sido educados correctamente acerca de la bioquímica del metabolismo energético en el músculo esquelético. Irónicamente, aún los principales libros de texto de bioquímica, no dedican un capítulo para explicar la bioquímica de la acidosis metabólica. Esta garantizada una cobertura específica de este importante tópico. Los libros de texto de fisiología del ejercicio son peores aún en su tratamiento de la bioquímica de la acidosis. La acidosis es atribuida a la producción de “ácido láctico”, el cual cuando se ioniza a ph celular, y libera un protón a la solución. Esta explicación es hecha sin ningún apoyo de investigaciones o de la bioquímica. Yo he demostrado claramente los defectos de esta explicación. Hasta que los libros de texto detallen las realidades de la bioquímica de la acidosis, el mito de la acidosis láctica va a continuar.
Comentario del Editor
La Fig.11 de Rob es un elegante resumen del flujo de los componentes involucrados en el consumo de energía y en la producción anaeróbica de la misma durante el ejercicio. Si seguimos el destino de una molécula de glucosa por este camino, terminamos con dos lactatos. En el camino, usamos dos moléculas de ADP y generamos dos ATP, y eso es todo. No hay una producción neta de H+. Pero el ATP y el ADP no cambian, porque la contracción muscular rompe dos ADP, dos Pi, y por supuesto dos H+. Entonces terminamos con dos lactatos y dos H+, por cada molécula de glucosa. Pasamos de glucosa a ácido láctico, pero el lactato y el ácido vienen de lugares diferentes: el H+ de la ruptura de ATP, y el lactato de la ruptura de glucosa. Este parece ser el enfoque de Rob, y parece perfectamente razonable. Pero también es perfectamente razonable argumentar que el H+ viene de la glucosa, de una u otra forma. El H+ producido en la hidrólisis del ATP viene del agua, seguro, pero la sustancia es conservada. En algún lugar y de alguna forma, en la ruptura de la glucosa un H+ es transferido al agua. Finalmente, una molécula de glucosa termina como dos moléculas de lactato y dos H+. No tengo una objeción real contra la “acidosis láctica”.
Respuesta del Autor
Will me contacto primero a raíz de tópico después de que leyó uno de mis resúmenes en la reunión anual del American College of Sport Medicine en el 2001. Este artículo despertó nuestra interacción.
Desde mi enfoque, y desde el de muchos fisiólogos y bioquímicos del ejercicio, el “ácido láctico” no causa directamente la acidosis metabólica inducida por el ejercicio. Will y yo hemos estado intentando resolver el razonamiento que hay detrás de este enfoque, y si este tiene algún significado real a como nosotros, como profesionales de las ciencias del ejercicio y el deporte, interpretamos a la acidosis y a la producción de lactato inducida por el ejercicio. Los comentarios de Will son consistentes con la evidencia bioquímica.
Ciertamente, si miramos al metabolismo desde una perspectiva general, el ejercicio intenso induce a la acidosis, que coincide con una acumulación de lactato. Mi problema con el concepto de “acidosis láctica” es que es otro ejemplo de una sobresimplificación de un hecho metabólico por parte de las ciencias del deporte y del ejercicio, por sencillez y conveniencia. Me preocupa que Will todavía quiera generalizar el hecho de una asociación entre la acidosis y el lactato. En mi experiencia, las generalizaciones dentro de las ciencias del deporte y el ejercicio son reflexiones negativas acerca de nosotros mismos, y no es de extrañar que nuestro campo sea visto pobremente por muchos (no todos) los académicos e investigadores de las ciencias puras. La acidosis metabólica inducida por el ejercicio es algo mucho más complejo, como para echarle la culpa a una reacción y a un producto, y nosotros deberíamos aceptar el desafió de ser verdaderos científicos para explicar la realidad de la acidosis celular. El punto no es si este enfoque hace una gran diferencia, o si el resultado neto es o no es el lactato, acompañado de protones. Enseñemos los hechos, y luego permitámonos ver adonde nos lleva la verdad: en educación, investigación, y en aplicaciones clínicas.
Finalmente, es importante darse cuenta que a pesar de que la evaluación de Will, del balance y la producción de protones, es razonable (2 lactatos + 2 protones), el hecho permanece en que debido a que los protones no vienen de la producción del lactato, hay un potencial para un desacoplamiento en la proporción de protones a lactato. Por ejemplo, la producción de lactato va a subestimar la liberación neta de protones cuando el piruvato, que no entra en la mitocondria, es incompletamente convertido a lactato (incremento del piruvato celular). Similarmente, cuando el piruvato es transportado dentro de la mitocondria, es convertido a acetil-CoA, y si este grupo acetilo se acumula dentro de la mitocondria debido a una respiración mitocondrial insuficiente, también se acumulan protones adicionales en el citosol debido a la ausencia de producción de lactato a partir de estos carbonos y por la falta del consumo de protones que produce el lactato. También ocurre una acumulación extra de protones en el citosol a partir de la hidrólisis del ATP, que se producen a partir de la reacción de la adenilato quinasa. Por todas estas razones, la liberación de protones es mayor que la producción de lactato.
Referencias
1. Brooks, G., A (2000). Intra- and extra-cellular lactate shuttles. Med Sci Sport Exerc; 32: 790-799
2. Busa, W., B., Nuccitelli, R (1984). Metabolic regulation via intracellular pH. Am J Physiol; 246: R409 R438
3. Dennis, S., C., Gevers, W., Opie, L., H (1991). Protons in ischemia: Where do they come from; where do they go to?. J Molecular Cell Cardiol; 23: 1077-1086
4. Gevers, W (1977). Generation of protons by metabolic processes in heart cells. J Molecular Cell Cardiol; 9: 867-874
5. Gevers, W (1979). Generation of protons by metabolic processes other than glycolysis in muscle cells: a critical view. [letter to the editor]. J Molecular Cell Cardiol; 11: 328
6. Hagberg, H (1985). Intracellular pH during ischemia in skeletal muscle: relationship to membrane potential, extracellular pH, tissue lactic acid and ATP. Pflugers Archives; 404: 342-347
7. Hochachka, P., W., Mommsen, T., P (1983). Protons and anaerobiosis. Science; 219: 1391-1397
8. Juel, C (1996). Lactate/proton co-transport in skeletal muscle: regulation and importance for Ph homeostasis. Acta Physiol Scand; 156: 369-374
9. Juel, C (1998). Muscle pH regulation: role of training. Acta Physiol Scand; 162: 359-66
10. Karlsson, J (1971). Lactate and phosphagen concentrations in working muscle of man. Acta Physiol Scand Supplementum; 358: 1-72
11. Katz A., Sahlin, K (1988). Regulation of lactic acid production during exercise. J Appl Physiol; 65: 509-18
12. Kent-Braun, J., A., Miller, R., G., Weiner, M., W (1993). Phases of metabolism during progressive exercise to fatigue in human skeletal muscle. J Appl Physiol; 75: 573-80
13. Lehninger AB, Nelson DL, Cox MM (1993). Principles of biochemistry. (second edition). New York, NY: Worth Publishers
14. Lindinger, M., I., Heigenhauser, G., J (1991). The roles of ion fluxes in skeletal muscle fatigue. Can J Physiol Pharmacol; 69: 246-253
15. Medbo, J., I., Tabata, I (1993). Anaerobic energy release in working muscle during 30 s to 3 min of exhausting bicycling. J Appl Physiol; 75: 1654-1660
16. Nelson, D., L., Cox, M., M (2000). Lehninger principles of biochemistry (third edition). New York,NY: Worth Publishers
17. Noakes, T., D (1997). Challenging beliefs: ex Africa semper aliquid novi. Med Sci Sports Exerc; 29: 571-590
18. Sahlin, K (1978). Intracellular pH and energy metabolism in skeletal muscle of man. Acta Physiol Scand; 455: S7-50
19. Sahlin, K., Katz, A., Henriksson, J (1987). Redox state and lactate accumulation in human skeletal muscle during dynamic exercise. Biochem J; 245: 551-556
20. Spriet, L., L., Sunderland, K., Bergstrom, M., Hultman, E (1987). Anaerobic energy release in skeletal muscle during electrical stimulation in men. J Appl Physiol; 62: 611-615
21. Spriet, L.,L., Sunderland, K., Bergstrom, M., Hultman, E (1987). Skeletal muscle glycogenolysis, glycolysis, and pH during electrical stimulation in men. J Appl Physiol; 62: 616-621
22. Spriet, L., L (1990). Anaerobic meatabolism in human skeletal muscle during short-term, intense activity. Can J Physiol Pharmacol; 70: 157-165
23. Stewart, P., A (1983). Modern quantitative acid-base chemistry. Can J Physiol Pharmacol; 61: 1444-1461
24. Stringer, W., Wasserman, K., Casaburi, R., Porszasz, J, Maehara, K., French, W (1994). Lactic acidosis as a facilitator of oxyhemoglobin dissociation during exercise. J Appl Physiol; 76: 1462-1467
25. Stryer, L (1988). Biochemistry (third edition). New York, NY: W.H. Freeman
26. Vaghy, P., L (1979). Role of mitochondrial oxidative phosphorylation in the maintenance of intracellular pH. J Molecular Cell Cardiol; 11: 933-940
27. Wilkie, D., R (1979). Generation of protons by metabolic processes other than glycolysis in muscle cells: a critical view. J Molecular Cell Cardiol; 11: 325-330
28. Zilva, J., F (1978). The origin of the acidosis in hyperlactataemia. Annals Clin Biochem; 15: 40-43