
Ácido Láctico y Ejercicio (Parte II)
Biosystem. Servicio Educativo.
Artículo publicado en el journal PubliCE, Volumen 0 del año 1997.
Publicado 10 de marzo de 2003
INTRODUCCIÓN
En la parte final de nuestro primer articulo sobre Ácido Láctico y Ejercicio, nos referimos a la importancia del conocimiento del metabolismo del ácido láctico, sustancia que a la luz de numerosas investigaciones de los últimos años, revela ser no sólo un metabolito de cuya producción se genera una importante liberación de energía anaeróbica (decisiva para aquellas pruebas o deportes donde la potencia anaeróbica lactácida es prevalente), sino también un potente sustrato para su reconversión a piruvato y ulterior oxidación mitocondrial, o alternativamente, un importante reservorio metabólico para producir glucosa sanguínea (neoglucogénesis) o glucógeno muscular y hepático (neoglucogénesis, proceso de reconstrucción de moléculas de glucógeno a partir de sustratos tales como el ácido láctico, pirúvico, glicerol, o aminoácidos tales como alanina y glutamina, proceso que debe ser diferenciado de la glucogenogénesis, que es la reconstrucción de moléculas de glucógeno a partir de glucosa).
Habiendo ya descrito los aspectos fisiológicos más importantes de la producción del ácido láctico, los mecanismos metabólicos y coenzimáticos involucrados, la reversibilidad de la reacción lactato-piruvato y la relación entre ácido láctico intracelular y sanguíneo, en el segundo artículo de esta serie, es nuestro objetivo profundizar aspectos (antes sólo mencionados), sobre los mecanismos de producción-remoción de lactato y su relación con los estímulos de entrenamiento, los caminos metabólicos del lactato durante el proceso de recuperación y la interacción del lactato y otros precursores en la reposición de los depósitos de glucosa y glucógeno que fueron depletados (es decir, consumidos, vaciados).
PRODUCCIÓN vs. REMOCIÓN DE ÁCIDO LÁCTICO DURANTE EL EJERCICIO (“LACTATE TURNOVER”)
Numerosas investigaciones con moléculas de ácido láctico “marcados” con radioisótopos (trazadores radioactivos) que son inyectados en el torrente sanguíneo para luego determinar su concentración en la sangre, después de su metabolización en órganos o territorios en particular, han probado que el ácido láctico es un metabolito muy activo durante el ejercicio realizado a diferentes grados de intensidad, tanto como en el período de reposo o descanso. Durante el reposo y el ejercicio de nivel muy moderado, el ácido láctico es producido, y a la vez removido, por la reversibilidad de la reacción, a velocidades similares. El balance entre producción y remoción es lo que puede darse en llamar “equilibrio reversible del lactato” (conocido en la literatura inglesa como “Lactate Turnover”).
Debido a que el lactato es un metabolito que se reconvierte rápidamente tendiendo a balancear su producción y su remoción, su concentración en los líquidos biológicos no cambia demasiado. Esto no sólo sucede dentro de las células, sino también en el torrente sanguíneo, en el cual el lactato volcado por las células es removido por otros tejidos, ávidos del mismo, para reconvertirlo a piruvato y oxidarlo en el Ciclo de Krebs.
Los mencionados estudios con trazadores han demostrado que, para un nivel dado de lactato sanguíneo (lactacidemia) el “turnover” (producción-remoción) de ácido láctico durante el ejercicio, es varias veces mayor que en reposo. Por lo tanto, si se observa que la lactacidemia permanece a valores cercanos a los de reposo durante el ejercicio suave, sería erróneo concluir que la causa radica en una falta de su producción (Figura 1 a).
A una intensidad de ejercicio ligeramente más elevada, la lactacidemia aumenta por encima de los valores de reposo, pero si la intensidad es mantenida (ya sea en forma continua o intercalada, con pausas muy breves), la lactacidemia se estabiliza en un nivel superior al de la Figura 1 a.
Numerosos trabajos y nuestro propia experiencia de investigación, demuestran que se pueden sostener trabajos en 50 y 80 minutos de duración a una tasa de balance (“turnover” o producción-remoción) tal, que la lactacidemia oscila entre 2 y 3, y hasta 4 mM/Lt y que la curva de lactato (Figura 1 b) puede tener fluctuaciones dentro de este rango y hasta descender ligeramente ante la prolongación del esfuerzo a una velocidad sostenida. Las razones de la mantención del estado de equilibrio de lactacidemia (o "steady-state" lactácido) en un nivel por sobre el de reposo, pero relativamente bajo (menos de 4 mM/Lt) y en un esfuerzo tan prolongado, se debe a un mecanismo multifactorial en el que intervienen la potencia oxidativa mitocondrial, que oxida el piruvato proveniente de la remoción, una mayor participación de los ácidos grasos en la degradación metabólica aeróbica, y una mayor capacidad para transferir el lactato al torrente sanguíneo y transportarlo del mismo modo a otros sitios metabólicos (grupos musculares menos activos o inactivos, corazón, etc.) con activa respiración celular, que toman al lactato como combustible oxidativo. El nivel funcional de ejercicio de este tipo de entrenamiento con un balance entre 2-4 mM/Lt, ha sido por nosotros nomenclado como nivel Subaeróbico.
En una tercera situación, (Figura 1 c) ante un ejercicio continuo o intervalado de mayor intensidad (con pausas mayores), la lactacidemia alcanza un nuevo estado de equilibrio (“steady-state”) entre su producción y su remoción, que transcurre en una franja entre 4-6 mM/Lt de concentración sanguínea. Los trabajos que se toleran en ese nivel fisiológico o franja funcional (nominado Superaeróbico), varían entre los 25’ y 40’ de duración según los individuos.
Los estudios con trazadores indican que la tasa de remoción o “clearence metabólico” (expresado por el cociente “turnover” o remoción de lactato/nivel sanguíneo de lactato) se incrementa cuando aumenta la lactacidemia (es decir que ante un incremento dado del denominador, se genera un aumento mayor del numerador). Esto significa, como lo habíamos descrito en el primer artículo de esta serie, que la tasa de remoción es dependiente de la concentración de lactato, es decir que la lactacidemia necesita alcanzar determinado nivel para forzar la remoción de lactato, y estimular a las enzimas y coenzimas que catalizan el proceso de reversibilidad.
De acuerdo con este razonamiento sería lógico pensar que convendría realizar ejercicios más intensos que generen un “steady-state” o estado de equilibrio entre producción-remoción más elevado, para aumentar la intensidad del “turnover”. Ello sucede si realizamos ejercicios continuos o intervalados a una intensidad más elevada que en la Figura 1 c, alcanzando una franja o área funcional entre 6 y 9 mM/Lt de lactacidemia (Figura 1 d).
El inconveniente de este tipo de estimulación, es que es muy difícil mantener el equilibrio dinámico en este nivel funcional por más de 12’ a 18’ de trabajo (o trabajo más pausa, en el caso del entrenamiento intervalado). Hemos denominado a este tipo de entrenamiento como área funcional de Consumo Máximo de O2 (VO2 máx.), ya que numerosas investigaciones, con control bilateral de VO2 y lactato alcanzan este nivel de lactacidemia, durante esfuerzos graduados donde fue alcanzado el mayor nivel de potencia aeróbica.
Teóricamente, si comparamos ahora la beneficiosa potencialidad de los entrenamientos Subaeróbicos (Figura 1 c) y de Consumo Máximo de Oxígeno (Figura 1 d), la mayor lactacidemia de éstos últimos garantizará una mayor potencia de producción-remoción. Sin embargo, la poca duración del trabajo (12’ a 18?), contrasta con una potencia quizás menor de remoción, pero de una duración sensiblemente más prolongada en los trabajos Superaeróbicos (25’ a 40?), por lo cual estos tipos de entrenamiento, en área funcional de 4 a 6 mM/Lt son los que probablemente tienen mayor responsabilidad de aumentar el “turnover” de lactato, complementándose con entrenamientos de VO2 máximo, que como su nombre lo indica son, quizás, los más incrementan la máxima potencia aeróbica.
En una última hipotética situación, ante un ejercicio con intensidad progresiva constante (continuo o intervalado pero a velocidad irreversiblemente creciente), es naturalmente imposible alcanzar un estado equilibrio (“steady-state”) lactacidémico, ya que, por la intensidad creciente los procesos de producción superan a los de remoción y entonces se genera una relación entre velocidad y lactato, que podemos visualizar en la Figura 1 e.
A través de esta gráfica podemos comprender con mayor claridad la definición de umbral aeróbico-anaeróbico lactácido y su diferencia con el estado de equilibrio en una tasa de producción-remoción determinada (“steady-state” del “lactate turnover”).
El umbral aeróbico-anaeróbico lactácido significa el punto de inflexión de una curva de relación entre velocidad (abcisa) y lactato (ordenada), que de un comportamiento casi lineal cambia a otro exponencial. La velocidad determinante de la forma de la curva es característica de cada sujeto.
Arribar a un estado de equilibrio o balance entre producción-remoción de lactato significa alcanzar determinados balances estables (lactacidemia constante), a intensidades determinadas, y durante un tiempo o duración más o menos prolongado (Figuras 1 a, 1 b, 1 c y 1 d). También es una característica individual el tiempo durante el cual un individuo puede sostener una intensidad de entrenamiento y un determinado balance lactacidémico. De allí la importancia de la evaluación de lactato intra y post-esfuerzo, ante diferentes volúmenes, intensidades y pausas de trabajo, para brindar un respaldo científico al concepto de especificidad de la carga de trabajo, en la búsqueda del efecto metabólico perseguido por el tipo de entrenamiento impartido.
Además, es importante destacar que durante el ejercicio sostenido al 50-70 % del VO2 máx., la tasa de producción-remoción (“lactate turnover”) se correlaciona linealmente con la curva en ascenso de VO2 máx. Durante el ejercicio sostenido aumentan, tanto la tasa absoluta como la relativa de oxidación, desde el estado de reposo hasta un ejercicio moderado al 50 % VO2 máx, la oxidación metaboliza el 30 % del ácido láctico removido, y la tasa de oxidación aumenta 3,5 veces con respecto a la tasa basal. Esto constituye un respaldo científico para la instrumentación de los trabajos regenerativos activos para acelerar los procesos de recuperación, como veremos más adelante. Veamos ahora qué sucede con la tasa de producción o aparición, y la tasa de remoción o desaparición de lactato, ante ejercicios que no mantienen un estado de equilibrio (“stady state”), es decir ante esfuerzos de intensidad progresiva. El lactato producido y removido (y por ende el balance entre producción y remoción intracelular) tienen una alta correlación lineal con el nivel de lactato arterial y venoso, y una correlación exponencial con el VO2. De allí la importancia de la determinación de lactato sanguíneo como medio de reflejar lo que está sucediendo con los procesos metabólicos aeróbicos-anaeróbicos. Esto está hipotetizado en trabajos de Boyd y cols. (1981) y Brooks y cols. (1985), donde se presume que el punto de ruptura de la curva, es decir, el umbral aeróbico-anaeróbico lactácido (Figura 1 e), está dado por un aumento de la tasa de producción o aparición, por sobre la tasa de remoción o desaparición.
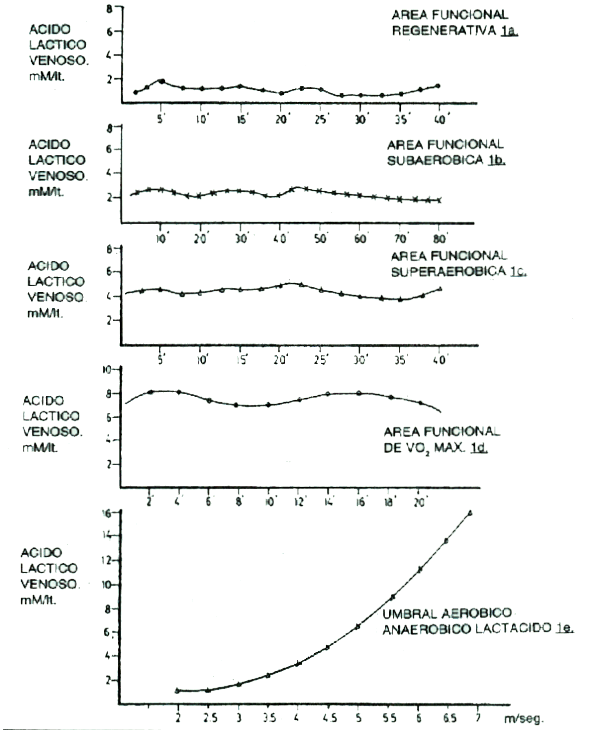
Figura 1
CAMINOS METABÓLICOS DEL ÁCIDO LÁCTICO DURANTE LOS PROCESOS DE RECUPERACIÓN
Se producen aproximadamente 100 mg/kg/hora de lactato en condiciones de reposo, estimándose que el 50 % se reconvierte a piruvato y es oxidado en el ciclo de Krebs. En la Figura 2 podemos ver porcentuales de lactato que siguen distintos caminos metabólicos, sea reconversión y oxidación mitocondrial, o utilizándose como un importante precursor neoglucogénico (sustrato para regenerar glucosa) o neoglucogenogénico (sustrato para regenerar glucógeno hepático o muscular), o bien como precursor de aminoácidos y proteínas. Esta información vital ha sido obtenida usando radioisótopos en animales, por Brooks y cols. (1973), y Brooks y Gaesser (1980), inyectando (U-14 C) lactato a ratas durante ejercicio intenso, hasta la fatiga absoluta. Durante varios momentos de la recuperación fueron medidos los niveles redioisotópicos en sangre, hígado, riñon, corazón y músculos. Las conclusiones respecto de los caminos metabólicos seguidos por el lactato se representan en la Figura 2. A su vez, se debe considerar que el comportamiento metabólico del lactato, cuando el ejercicio se detiene, dependería de las condiciones metabólicas internas. Por ejemplo, altos niveles de lactato y condiciones casi normales para otros sustratos, como glucógeno hepático y glucosa sanguínea, favorecerían la oxidación del lactato. Por el contrario, un gran vaciamiento glucogénico y/o una hipoglucemia, favorecerían tanto la neoglucogénesis como la neoglucogenogénesis, con una menor tasa de oxidación de lactato.
Por lo tanto, podemos afirmar que, durante el proceso de recuperación, más del 70 % del lactato es reconvertido a piruvato y oxidado en el Ciclo de Krebs (con producción de ATP más C2O y H2 O).
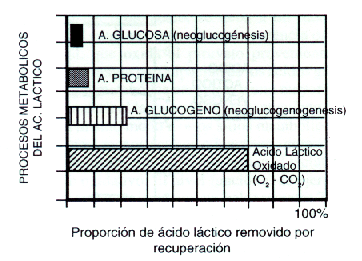
Figura 2
Este mecanismo es también utilizado en elevada proporción en los ejercicios que alcanzan estados de equilibrio (“steady state”) entre niveles de producción y remoción. Esto es por demás significativo, ya que cada molécula de lactato que se oxida es sustitutiva de la glucosa, ahorrando su consumo, ya sea proveniente del torrente sanguíneo o de la ruptura del glucógeno muscular.
El otro 30 % del lactato removido en el proceso de recuperación tiene diferentes destinos ya enunciados; en orden de prioridades:
1) Un 2-3 % se utiliza para la restitución de la glucosa sanguínea.
2) Un 20 % es precursor neoglucogénico (en músculo y corazón en primera
instancia, y luego en el hígado).
3) 5-7 % se emplea para la restitución de aminoácidos y proteínas.
TASA DE REMOCIÓN EN LA RECUPERACIÓN ACTIVA Y PASIVA
Numerosas investigaciones han demostrado que la curva de desaparición del lactato sanguíneo, es sensiblemente diferente y mucho más rápida con trabajo regenerativo activo, como lo muestra la Figura 3. Mucho se ha polemizado acerca de la intensidad (en % de VO2 máx.), o la velocidad (en % de la velocidad máxima de competición), a las que deben realizarse los trabajos de recuperación activos.
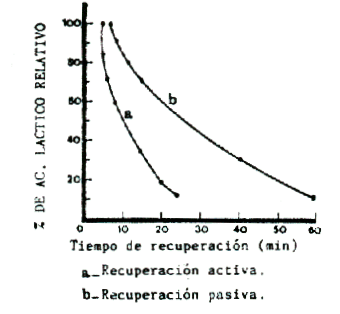
Figura 3
Considerando los deportes de base como el atletismo y la natación, podemos sinterizar las conclusiones de numerosas investigaciones, diciendo que en carrera se alcanzan los más elevados niveles de oxidación y remoción a una intensidad entre el 30 y 45 % VO2 máx., equivalente a velocidades entre el 35 y el 50 % de la velocidad máxima (Figura 4).
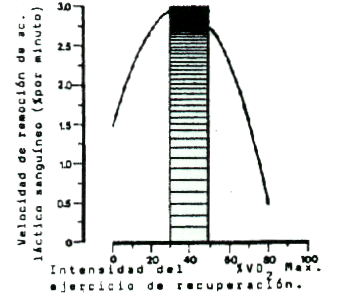
Figura 4
En natación (Figura 5), la más elevada tasa de remoción de lactato se obtiene a intensidades que oscilan en el 55 y el 70 % del VO2 máx., o a velocidades entre el 60 y el 75 % de la máxima velocidad competitiva.
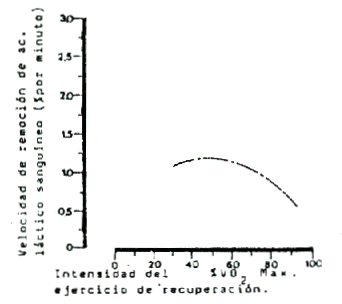
Figura 5
El papel del trabajo regenerativo activo es de capital importancia en los procesos de recuperación activos. El respaldo científico de esta aseveración radica en que la actividad física a ritmo suave, es activadora del proceso de reconversión de lactato residual y de su posterior oxidación intramitocondrial. Con respecto a este proceso, podemos considerar dos situaciones:
a) El transporte de lactato dentro del músculo activo (“Lactate Shuttle”
dentro del músculo).
b) El transporte de lactato hacia la sangre y de allí a otros grupos
musculares y regiones celulares de alto potencial oxidativo (“Lactate Shuttle”
intravascular).
Con respecto a la situación a), como lo sugieren las diferencias arteriovenosas en mediciones de lactato con trazadores radioisotópicos, aproximadamente sólo la mitad del lactato formado en el músculo ejercitado es liberado a la circulación venosa. Aproximadamente la otra mitad, junto con una significativa cantidad de lactato removido de la circulación arterial, es consumido dentro del músculo y aparece como C2O en la sangre venosa. Basados en resultados de Baldwin y cols. (1977, 1978), puede observarse que el lactato producido preponderantemente en fibras glicolíticas anaeróbicas (II b), es difundido y oxidado dentro del propio músculo a fibras lentas o aeróbicas (Tipo I), o también a fibras rápidas semioxidativas (II a) )Figura 6).
Considerando ahora la situación b), el trabajo regenerativo activo logra que el lactato circulante en una fracción de 1 minuto, alcance el corazón y sea enviado por la circulación arterial a territorios musculares, algunos inactivos o poco activos durante el proceso de ejercicio previo, que están ávidos de ese lactato para oxidarlo en sus comportamientos celulares de alta capacidad de respiración oxidativa.
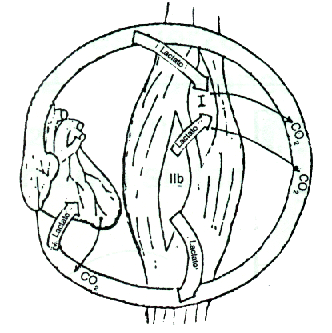
Figura 6
Es obvio concluir que este proceso será beneficioso por el aumento del volumen minuto y de la redistribución del flujo sanguíneo, variables incrementadas con el trabajo activo y disminuidas durante la recuperación pasiva. En pocas palabras, a la luz de esta actualizada información científica, debemos considerar al lactato como un metabolito que vehiculiza la liberación de energía adicional, importante para deportes de prestación anaeróbica masiva o aeróbico-anaeróbicos alternados, y que gran parte de los procesos de entrenamiento modernos están dirigidos a lograr su producción racional y su utilización durante el esfuerzo, y/o durante el proceso de recuperación, como una gran fuente de combustible para los procesos aeróbicos (en sustitución de la glucólisis) o como un potente metabolito neoglucogenogénico.
Referencias
1. Balwin K M, Hooker A M, y Herrick R E (1978). Lactate oxidative capacity in different types of muscle. Biochem. Biophys. Res Commun. 83: 151-157
2. Balwin K M, Campbell P J, y Cooke D A (1977). Glycogen, lactate, and alanine charges in muscle fiber type during graded exercise. J Appl Physiol, 43: 288-291
3. Barr D P, y Himwich H E (1923). Studies in the physiology of muscular exercise. II Comparison or arterial and venous blood following vigorous exercise. J Biol Chem, 55: 523-533
4. Belcastro A N, y Bonen A (1975). Lactic Acid Renoval rates during controlled and uncontrolled recovery exercise. J Appl Physiol, 39 (6), 932-935
5. Boyd M E, Albright E B, Foster D W, y Mc Garry J D (1981). In vitro reversal of the fasting state of liver metabolism in the rat. J Clin Invest, 88: 142-152
6. Brooks, G A (1986). The lactate shuttle during exercise and recovery. Med Sci Sport Exerc; 18: 360-368
7. Brooks G A, Brauner K E, y Cassens R G (1973). Glycogen synthesis and metabolism of lactic acid after exercise. Am J Physiol, 224: 1162-1166
8. Brooks, G A, Fahey T (1985). Exercise Physiology. Human Bioenergetics and its Applications. Capítulos 5 (pp. 67-95) y 10 (pp. 189-219). Macmillan Publ. Co. New York
9. Brooks, G A y Gasser G A (1980). End points of lactate and glucose metabolism after exhausting exercise. J Appl Physiol, 49: 1057-1069
10. Brooks, G A, Stainley W C, Gertz E W, Wisneski J A, Morris D L, y Neese R A (1985). Systemic lactate kinetics during graded exercise in man (Abstract). Med Sci Sport Exerc, 17: 206
11. Cazorla G, Oufurt C, Cervetti J P, y Montpetit R R (1983). The influence of Active Recovery on Blood Lactate Disappearence after Supramaximal Swimming. Hollander A P, Huijing P A and Degroot G (Eds.), Human Kinetics Publ., pp. 244-250
12. Gollnick, P D, Bayly, W M, Hodson, D R (1986). Exercise intensity, training, diet, and lactate concentration in muscle and blood. Med Sci Sport Exerc; 18: 334-340
13. Katz, J (1986). The application of isotopes to the study of lactate metabolism. Med Sci Sport Exerc; 18: 352-359