
Consistência na Análise de Sinal Emg de Superfície nos Membros Inferiores em Diferentes Contramovimentos Verticais
Consistency of Emg Surface Signal Analysis From Different Lower Limb Vertical Countermovement
Carlos Rodrigues1,2, Miguel Correia1,2, João Abrantes3, Jurandir Nadal4 y Marco Benedetti5
1Centre for Biomedical Engineering Research, INESC TEC - Technology & Science Associate Laboratory, Porto, Portugal
2Faculdade de Engenharia da Universidade do Porto, Portugal
3MovLab – Laboratório de Tecnologias e Interfaces, Universidade Lusófona de Humanidades e Tecnologias, Portugal
4Progr. de Eng. Biomédica, COPPE/UFRJ – Universidade Federal do Rio de Janeiro, Brasil
5Dep. de Eletrônica e Sistemas, Universidade Federal de Pernambuco, Brasil
Artículo publicado en el journal Gymnasium del año 2017.
Publicado 4 de junio de 2017
Resumen
Palabras clave: Consistência análise EMG
Abstract
Keywords: Consistency of EMG
INTRODUÇÃO
As limitações da análise anatómica funcional
A anatomia classifica os músculos em grupos funcionais como flexores, extensores, adutores e abdutores, de acordo com a análise das ações mecânicas postuladas, sendo estas classificações utilizadas de forma geral em diferentes domínios de aplicação. Este método baseia-se essencialmente na linha de ação muscular em relação a um eixo de rotação articular, inferindo-se uma ação articular com base na direção da linha de ação e na capacidade muscular de contração aplicada nos pontos anatómicos de fixação, Knudson (2007). Assim, apesar de, quando ativado, o músculo exercer aproximadamente a mesma força de contração em ambas as extremidades, o movimento de cada uma das extremidades depende de vários fatores biomecânicos, uma vez que a força resultante em cada uma das extremidades pode ser distinta em virtude das restantes forças musculares e extra-musculares aplicadas por tecidos conetivos na proximidade.
Uma das vias para determinação direta dos músculos que se encontram ativos num determinado movimento é a via invasiva, tendo sido detetados em estudos com animais resultados complexos e inesperados da atividade muscular. Num destes estudos realizados em perus, Roberts, Marsh, Weyand, e Taylor (1997) descobriu que o músculo flexor plantar, gastrocnémios, atuava de forma quase isométrica durante a fase de apoio na corrida em terreno nivelado, e de forma concêntrica na corrida no sentido ascendente em plano inclinado. Em virtude do caráter invasivo das medições, estas investigações têm sido realizadas principalmente em animais, embora estudos em humanos com recurso à implantação de sensores de força de fibra ótica, Komi et al. (1996), tenham também revelado comportamentos musculares complexos que vão além dos postulados para as ações musculares com base nos modelos concêntricos monoarticulares que decorrem da análise anatómica funcional, Knudson (2007).
A importância da compreensão das ações musculares
De acordo com Knudson (2007) que cita Bartlett (1999), as ações musculares durante o movimento humano são mais complexas do que as postuladas pela anatomia funcional, apontando para a necessidade de combinação de vários estudos biomecânicos quantitativos para melhorar a compreensão da função muscular no movimento humano.
Os estudos EMG têm permitido documentar tendências de ativação em grupos musculares específicos com variações consideráveis entre sujeitos, sendo a redundância muscular apontada como a principal fonte de variabilidade de acordo com Basmajian & De Luca (1985). Knudson (2007) cita vários estudos EMG que têm também vindo a demonstrar que os padrões de ativação dos músculos individuais não são representativos de todos os músculos do grupo funcional, e que mesmo os músculos individuais apresentam diferenças no recrutamento das unidades motoras de acordo com a tarefa ou ação muscular, podendo alternar a ativação entre músculos em cada grupo muscular para minimizar a fadiga. Além disso, as ativações musculares podem variar em função dos ângulos articulares e do grau de estabilização necessário, constituindo as sinergias definidas por Bernstein (1967) um mecanismo fundamental de combinação da ativação muscular, de modo a otimizar a execução de uma ação motora, podendo a distribuição da carga necessária pelos grupos musculares variar de acordo com o tipo de fibras, as propriedades contráteis, a área da secção transversal, o braço da força ou a atividade antagonista, apontando para o facto de que a ativação muscular é consideravelmente mais complexa do que era considerado. Têm ainda vindo a ser apontadas diferentes ativações das secções intramusculares que vão além da segmentação tradicional da anatomia clássica e que apontam para a importância da microanatomia, em particular no estudo dos músculos com largas regiões de fixação.
As descrições da anatomia músculo-esquelética não atendem frequentemente à variação inter-indivíduos dos pontos de fixação muscular com o consequente efeito na ação muscular e o caráter interligado do corpo humano faz com que as ações musculares possam provocar efeitos consideráveis em articulações próximas ou até distantes, de acordo com Richmond (1998) e Zajac (1991) citados por Knudson (2007), que aponta como exemplo a sobreposição da ação do músculo sóleo na extensão do joelho relativamente à flexão plantar quando o pé se encontra apoiado. A multiplicidade de ações musculares pode ainda ser ilustrada pela contribuição dos músculos biarticulares e multiarticulares, distinta quando comparada com a contribuição dos músculos monoarticulares, sendo necessário atender à distribuição das ações pelas articulações em interação para compreensão das ações dos músculo multiarticulares, conforme van Ingen Schenau et al. (1995).
Os ciclos de alongamento e encurtamento musculares
As ações musculares mais estudadas são as formas isoladas correspondentes às ações isométricas, concêntricas e excêntricas. Os movimentos naturais envolvem no entanto o recurso à ação muscular correspondente ao ciclo de alongamento e encurtamento (CAE) muscular, como uma forma específica e independente de ação muscular controlada pelo sistema neuromuscular, para antecipação do impacto com vista a proteção da integridade estrutural, eficiência em ações submáximas e potenciação em ações máximas, como acontece no caminhar, na corrida e no salto, Komi et al. (2011).
Os estudos da potenciação da ação muscular por via do CAE remontam aos trabalhos ex vivo de Cavagna, Saibene, e Margaria (1965) que demonstraram, com recurso a músculos de rã, que o trabalho mecânico realizado pela contração muscular é superior quando precedido por uma ação de alongamento, no que se designa por ciclo de alongamento e encurtamento muscular, Knudson (2007). Recorrendo a medições diretas invasivas com sensores de fibra ótica e estimativas de comprimento das unidades músculo-tendinosas (UMT), Komi et al. (1996) desenvolveram uma estimativa da relação in vivo entre o momento de força e a velocidade angular, apontando para o aumento de desempenho por via do CAE muscular. Apesar do elevado esforço de investigação e da diversidade de variáveis biomecânicas analisadas, a origem e os mecanismos funcionais das vantagens mecânicas do CAE mantêm-se um assunto em aberto, sendo apontadas como principais fontes na produção de maior força muscular, além do efeito potenciador da resistência a movimentar (Zatsiorsky & Kraemer, 2006), o efeito dos movimentos precedentes no comportamento muscular (Herzog & Leonard, 2000), o reflexo miotático resultante do alongamento (Komi & Golhofer, 1997), a energia elástica armazenada durante o alongamento (Gregor, 1997) e o tempo necessário para desenvolvimento do nível de força (Bobbert, Gerritsen, Litjens & van Soest, 1996).
Estudos posteriores in vivo como o realizado por Kubo, Kanehisa, Takeshita, Kawakami, Fukashiro e Fukunaga (2001) sobre o comportamento muscular do gastrocnémios em movimentos com recurso ao CAE mostraram que a elasticidade do tendão permite a atuação das fibras musculares em condições quase isométricas no momento da inversão articular enquanto o músculo contrai, permitindo o armazenamento de energia elástica nas estruturas tendinosas para realização de maior trabalho positivo.
Sobre a questão em aberto relativa ao significado do aumento brusco e do pico acentuado de potência durante a flexão plantar do tornozelo registado na fase de pré-transferência que se segue ao duplo suporte durante o caminhar, Perry e Burnfield (2010) apontam como explicação mais utilizada a força exercida pelo gastrocnémios e pelo sóleo para propulsão do corpo para diante. Do ponto de vista fisiológico esta hipótese não era aceite em virtude da ausência de atividade EMG associada à rápida flexão plantar do tornozelo. Perry e Burnfield (2010) apontam estudo posterior com recurso a sensores portáteis de ultrassom utlizados para visualizar os fascículos musculares do gastrocnémios medial durante o caminhar, tendo permitido registar a resposta simultânea da variação individual do comprimento da unidade músculo-tendinosa (UMT) e dos fascículos do gastrocnémios medial no movimento do tornozelo durante a fase final do apoio. A dorsiflexão na fase final de apoio apresentou considerável aumento de comprimento da UMT sem variação de comprimento das fibras musculares, com intensa atividade isométrica EMG. A flexão plantar que antecede a fase de transferência apresentou uma contração rápida da UMT com uma reduzida variação de comprimento das fibras musculares sem atividade EMG, sendo o encurtamento do tendão associado à produção de força.
Komi et al. (2011) refere que a intensidade de pré-ativação do músculo gastrocnémios na corrida depende diretamente da carga esperada no impacto, sendo a unidade músculo-tendinosa (UMT) pré-ativada, alongada durante o impacto e esta fase excêntrica retardada, seguida de imediato pela fase concêntrica de encurtamento. Sobre a definição e a identificação do recurso ao CAE muscular, Komi et al. (2011) afirma que a definição do CAE deve ser encarada de forma mais crítica e que a identificação precisa das fases excêntrica e concêntrica da ação muscular requer a utilização de análise cinemática, dinâmica, EMG e ultrassom (US). Recorrendo a US de elevada frequência, até 200 Hz, este estudo procura aprofundar a definição de CAE, discriminando ao nível da UMT o comportamento dos fascículos musculares (FM) relativamente ao comportamento do tendão (T), uma vez que os fascículos musculares são controlados quer pelas forças externas de alongamento quer pela ativação interna neuromuscular, tornando-se necessário diferenciar as variações de comprimento dos fascículos e dos tendões em vários músculos com exercícios envolvendo CAE. Os resultados obtidos apresentaram comportamento muscular consistente e diferenças intermusculares caraterísticas, colocando-se a hipótese de que além da potenciação do CAE pelo reflexo de alongamento, as variações individuais do comprimento dos fascículos e do tendão possam depender na sua ação conjunta, do tipo de movimento, da sua intensidade e dos músculos envolvidos, levantando-se a questão sobre qual o motivo pelo qual o comprimento dos fascículos varia de forma distinta em diferentes movimentos e da necessidade de avaliação da atividade EMG e US.
Blazevich (2011) afirma que, em várias ações envolvendo o CAE, os músculos são ativados para aumentar a sua rigidez, sendo o alongamento e a energia armazenada dominados pelo tendão que é responsável pela maior parte da variação do comprimento conjunto da unidade músculo-tendinosa.
Blazevich (2011) cita o estudo de Kurokawa et al. (2003) sobre o salto de impulsão vertical para afirmar que o músculo e o tendão realizam a contração e alongamento em momentos distintos para várias ações envolvendo o CAE. O estudo de Kurokawa et al. (2003) permitiu verificar que, no salto com contramovimento, a UMT formada pelo tendão de Aquiles e o músculo gastrocnémios se mantém com comprimento aproximadamente constante até ao momento em que contrai rapidamente durante a flexão plantar do tornozelo na sequência de movimentos do salto. Apesar disso, o tendão sofre inicialmente o alongamento por ação da ativação do músculo, contraindo em seguida rapidamente, enquanto o músculo contrai lentamente durante o movimento mantendo-se em seguida quase-isométrico. Desta forma a fase de maior velocidade de impulsão durante o salto é associada à contração rápida do tendão de Aquiles com uma contração mínima do músculo gastrocnémios.
Os contramovimentos verticais na avaliação dos CAE
Atendendo ao objetivo de produção do máximo nível de força no mais curto intervalo de tempo, mantendo em seguida o maior nível de força durante o maior período de tempo, a realização de um contramovimento no sentido contrário ao movimento pretendido baseia-se enquanto ação natural na utilização do peso corporal como forma de aumento da carga para produção de uma maior resistência e consequentemente da produção do maior nível de força muscular.
Os efeitos do aumento da resistência externa no aumento da capacidade de produção de força muscular encontram-se bem documentados em Zatsiorsky e Kraemer (2006) e enquadram-se no conjunto de fatores extrínsecos que, em complemento dos fatores intrínsecos correspondentes entre outros ao tempo disponível para produção de força, velocidade, direção do movimento e posição corporal, apontam para a necessidade do estudo in vivo da capacidade de produção de força em condições e movimentos naturais, com vista a avaliação da interdependência destes fatores na capacidade de produção de força.
Encontrando-se o CAE presente numa diversidade de movimentos como o caminhar, a corrida e o salto, de acordo com Knudson (2007), Komi et al. (2011) e Blazevich (2011), a opção pela sua avaliação em saltos de máxima impulsão vertical, com e sem contramovimento curto e longo, deve-se ao elevado grau de normalização no controlo das condições dos ensaios, para isolamento do fenómeno em estudo face a outras variáveis, da maior expressão no contraste da presença do CAE nos diferentes ensaios, da naturalidade do movimento, disponibilidade dos equipamentos necessários para avaliação e da maturidade e estabilidade dos protocolos utilizados na avaliação.
O desenvolvimento e a utilização de protocolos de saltos de impulsão vertical com e sem contramovimento para avaliação da energia mecânica produzida remontam aos trabalhos de Marey e Demeny (1885), Sargent (1921), Abalakov (1938), Margaria, Aghemo e Rovelli (1966), Cavagna et al. (1971) e Asmussen e Bonde-Petersen (1974) na forma mais natural, permitindo a oscilação dos braços e posteriormente adaptados de modo a manter as mãos na anca para avaliação exclusiva da capacidade de produção de força pelos membros inferiores, Komi e Bosco (1978), Bobbert et al. (1986), Aragón-Vargas e Gross (1997), mantendo-se estes procedimentos utilizados, nomeadamente por Bobbert (2011) e Komi et al. (2011), em virtude do seu caráter padrão, reprodutibilidade e simplicidade.
Se por um lado a questão do máximo desempenho, expresso pela altura máxima durante a fase de voo do salto vertical, se pode explicar pelo desenvolvimento durante a fase de impulsão, do maior impulso linear obtido através do maior nível de força de impulsão durante a maior duração temporal, da maior potência mecânica obtida através do maior nível de força aplicada à máxima velocidade, ou do maior trabalho mecânico realizado pela aplicação do maior nível de força durante o maior deslocamento, a realidade é que a capacidade de força aplicada diminui em geral com o tempo de aplicação, com o aumento da velocidade e do deslocamento durante o qual é aplicada, Zatsiorsky e Kraemer (2006). Além da questão da intensidade das grandezas força, velocidade e deslocamento, a fase temporal relativa entre estas grandezas apresenta também um papel determinante no desempenho obtido.
Os saltos utilizados para avaliação dos CAE’s associados a cada tipo de contra-movimento são o Squat Jump (SJ) sem contramovimento, o Counter Movement Jump (CMJ) com contramovimento longo e o Drop Jump (DJ) com contra-movimento curto. O SJ consiste na realização de uma impulsão vertical cuja característica principal é a ausência de contramovimento e consequentemente a ausência de CAE. A posição de partida corresponde ao agachamento num ângulo aproximado entre a coxa e a perna de 90°. Mantendo em seguida essa posição solicita-se aos sujeitos a realização exclusiva de um movimento vertical máximo ascendente. O CMJ e o DJ caracterizam-se pela presença de um contra-movimento e consequentemente de um CAE. No CMJ a posição de partida corresponde à posição ortostática realizando um contramovimento seguido de um movimento ascendente e solicitando-se ao sujeito que procure inverter o sentido do movimento para um ângulo entre a coxa e a perna próximo dos 90°. No Drop Jump o contramovimento é imposto pela queda livre, pedindo-se ao sujeito que se coloque sobre um plano elevado e que dê um passo em frente iniciando um movimento de queda livre, a que se segue o contacto com o solo e o contramovimento em resultado do qual inicia um movimento ascendente.
Durante a execução destes saltos procura-se reduzir substancialmente o deslocamento angular da metade superior do corpo, principalmente dos membros superiores. Pede-se por isso aos sujeitos que coloquem o corpo próximo da vertical, mantendo as mãos na região pélvica e procurando manter este alinhamento durante a execução do salto. Esta restrição tem por objetivo concentrar o estudo nas ações realizadas pelos membros inferiores, aumentando o controlo e reduzindo as variáveis experimentais determinantes.
Músculos selecionados dos membros inferiores
Weineck (2008) refere que a marcha, a corrida e os saltos solicitam aproximadamente os mesmos músculos, residindo as diferenças principalmente na distribuição do trabalho entre os diversos grupos musculares, sendo que a locomoção bípede requer uma musculatura forte na região dorsal da perna já que estes músculos têm que atuar vencendo todo o peso do corpo. Assim, a musculatura humana apresenta um predomínio dos músculos flexores cuja função consiste em estender o pé relativamente aos músculos extensores. Desta forma, enquanto os músculos extensores do pé são constituídos por um único grupo muscular na face anterior da perna, os músculos flexores encontram-se agrupados em duas camadas, uma mais superficial composta pelo grupo tríceps sural que inclui os gastrocnémios medial / lateral e o sóleo, além de uma camada mais profunda.
De acordo com Hamill et al. (2009), a maior parte dos músculos cruzam apenas uma articulação, decorrendo por isso a ação dominante dos músculos monoarticulares sobre a articulação que cruzam. Os músculos biarticulares constituem um caso particular de músculos que cruzam duas articulações, dando origem a vários movimentos articulares que ocorrem frequentemente em sequências opostas, como é o caso do reto femoral (RF) que é um músculo biarticular responsável pela flexão da anca e pela extensão do joelho. Atendendo que nos saltos de impulsão vertical a extensão da anca e do joelho são utilizadas para impulsionar verticalmente, coloca-se a questão sobre se o músculo RF, enquanto flexor da anca e extensor do joelho, contribui para a extensão do joelho, opõem-se à extensão da anca ou ambos.
A ação de um músculo biarticular depende da posição do corpo e da interação muscular com os elementos externos como o solo, de acordo com Zajac (1989), citado por Hamill et al. (2009). No caso do RF, o músculo contribui principalmente para a extensão do joelho em função da posição articular da anca, Gowitzke (1984). Esta posição tem como resultado que a força produzida pelo RF se exerce mais próximo da articulação da anca, limitando assim a ação do músculo e a produção efetiva de força nesta articulação.
Ainda de acordo com Hamill et al. (2009), o grupo hamstrings atua principalmente na extensão da anca em vez da flexão do joelho em virtude do maior braço de força em relação à anca. O gastrocnémios atua principalmente na flexão plantar relativamente ao tornozelo em vez da flexão do joelho, uma vez que o seu braço de força em relação ao tornozelo é superior. Nos saltos de impulsão vertical a altura máxima obtém-se quando se efetua primeiro a extensão das articulações proximais, deslocando-se o movimento distalmente para um ponto em que ocorre a extensão da articulação do tornozelo, flexão plantar. No momento em que a articulação do tornozelo participa na sequência do movimento são necessários momentos de força e velocidades articulares muito intensas, van Soest et al. (1993), sendo o papel dos músculos biarticulares determinante. O músculo biarticular gastrocnémios cruza as articulações do joelho e do tornozelo, sendo a sua contribuição influenciada pela posição do joelho. Durante o salto a extensão da articulação do joelho permite otimizar o comprimento do gastrocnémios mantendo a sua velocidade de contração baixa, apesar da elevada velocidade de flexão plantar no tornozelo, permitindo ao gastrocnémios níveis de força superiores por via da redução da velocidade de rotação, Bobbert (1988).
A utilização de EMG de superfície na avaliação muscular
Atendendo à necessidade de compreensão das ações musculares, a análise EMG de superfície afigura-se como uma ferramenta importante na avaliação do movimento humano natural, dado o seu caráter não invasivo, fácil utilização e baixo custo. Tal como afirmado por Winter (2009), o principal motivo de interesse no processamento de sinal EMG consiste no relacionamento entre este e algum tipo de medida da função muscular, mantendo-se em aberto a questão da utilidade do sinal EMG na estimativa da tensão muscular e do potencial de interesse desta relação. Em virtude da subjetividade das comparações qualitativas torna-se cada vez mais necessário a aplicação de métricas capazes de reproduzir de forma quantitativa e objetiva a interpretação dos resultados relativos aos padrões de ativação EMG.
OBJETIVO
Atendendo à diversidade de fontes de variabilidade na atividade muscular, do elevado número de variáveis associadas à atividade muscular e do registo de comportamentos distintos dos esperados, nomeadamente durante o CAE muscular, o objetivo do presente estudo consiste na avaliação da consistência de análise e resultados obtidos por aplicação de diferentes técnicas de estudo dos sinais EMG de superfície associados a grupos musculares estratégicos em diferentes contramovimentos (CM) nos membros inferiores avaliados em saltos de máxima impulsão vertical (MIV).
MÉTODO
Amostra
Caraterização da amostra
A amostra experimental é composta por um grupo de n=6 estudantes de licenciatura em educação física e desporto, sem aptidão ou treino físico específico. Os sujeitos com idades (21,5±1,4) anos foram pesados (76,7±6,7) kg e medida a sua estatura (1,79±0,05) m, tendo sido colocados após preparação e limpeza da pele com álcool, elétrodos Skintact F55 Aqua-Wet com forma circular e 55 mm de diâmetro, longitudinalmente às fibras musculares na configuração bipolar nos músculos reto femoral (RF), vasto medial (VM), vasto lateral (VL) e gastrocnémios medial e lateral (GM/GL), de acordo com as contribuições indicadas pela anatomia funcional para a flexão da anca, extensão do joelho e flexão plantar. Cada sujeito realizou um total de três ensaios para cada um dos tipos de saltos MIV considerados, nomeadamente sem CM (SJ - Squat Jump), com CM longo (CMJ - Counter Movement Jump) e com CM curto (DJ - Drop Jump).
Procedimentos operacionais
Durante os ensaios foram registadas as atividades EMG de superfície dos grupos musculares selecionados, com recurso a sistema eletromiográfico sem fios Biotell 99 da Glonner Electronic GmbH à frequência de 2000 Hz e as forças de reação do solo com recurso a plataforma de força AMTI BP2416-4000CE à frequência de 1000 Hz. Foram detetadas as fases de movimento a partir dos perfis de força registados e selecionados o melhor salto de cada tipo de MIV de acordo com o tempo máximo de voo por ausência de contacto com a plataforma de força. Os sinais brutos EMG foram sujeitos a retificação de onda completa e filtrados com filtro passa-baixo Butterworth de 2ª ordem e frequência de corte 2,3 Hz para obtenção da envolvente linear, aplicado no sentido direto e inverso para evitar deslocamento de fase Winter (2009). Os sinais obtidos foram correlacionados (CR) para deteção de atividade sinergística, agonista e antagonista, análise de componentes principais (ACP) para deteção de componentes não correlacionados explicativos da maior variabilidade e correlação cruzada normalizada (CCRN) para deteção de correlações máximas e atraso eletromecânico. Assim, a coordenação expressa pelo desfasamento determinada pela atividade muscular mantém-se uma questão em aberto na explicação do desempenho obtido.
Correlação EMGs
De modo a analisar a relação existente entre a atividade registada dos músculos selecionados, nomeadamente sinergista, antagonista e desfasamento temporal foram correlacionados por pares os sinais EMGij com i, j=1, 2, 3, 5, 6 e i≠j os canais utilizados para monitorização dos músculos RF, VM, VL, GM e GL, sendo os canais 4, 7 e 8 utilizados para deteção e eliminação de artefactos nos restantes canais. Foram também calculados os coeficientes de correlação de Pearson correspondentes às retas ajustadas de acordo com o método dos mínimos quadrados às atividades EMGij nos diagramas de fase, Vaughan (1999),
Correlação cruzada EMGs
Atendendo ao desfasamento temporal de máxima correlação entre as atividades EMGij procedeu-se também à correlação cruzada por pares dos sinais registados para avaliação da correlação máxima e do desfasamento temporal de máxima correlação, de acordo com Winter (2009).
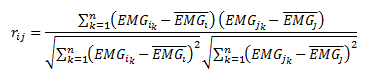
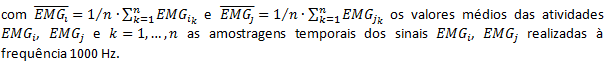
Correlação cruzada EMGs
Atendendo ao desfasamento temporal de máxima correlação entre as atividades EMGij procedeu-se também à correlação cruzada por pares dos sinais registados para avaliação da correlação máxima e do desfasamento temporal de máxima correlação, de acordo com Winter (2009).
Enquanto a correlação de Pearson fornece um único coeficiente, a correlação cruzada permite obter uma série de coeficientes de correlação para cada instante de desfasamento temporal entre os sinais x(t) e y(t) correlacionados de acordo com a expressão,
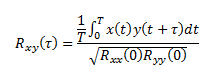
Na versão digital implementada em Matlab R2010b (The MathWorks Inc., Natick, MA, USA) a expressão utilizada para correlação dos sinais foi,
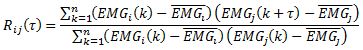
com τ=m∙∆t, m=1,⋯,2n-1 e ∆t=0,001s.
Análise de componentes principais EMGs
Embora as correlações de Pearson e as correlações cruzadas permitam aferir o grau de coatividade e, no caso das correlações cruzadas, o desfasamento temporal de maior coatividade, o número de comparações emparelhadas para músculos é que é da ordem , crescendo por isso de forma acentuada com . Tomando como referência o trabalho de Winter (1987) para 16 músculos, este corresponderia a 120 correlações, Vaughan (1999).
Atendendo ao objetivo de procura do menor número de componentes capazes de explicar a máxima variação dos fatores explicados, suportado pelo princípio de igual simplicidade apresentado por Bernstein (1967) que postula o controlo pelo sistema nervoso central em algum nível de sinergia em virtude da incrível complexidade de controlo individualizado de cada músculo, foi aplicada a análise de componentes principais (ACP) às envolventes dos sinais EMGs retificados e filtrados dos músculos selecionados.
A capacidade da análise de componentes principais (ACP) para identificar relações entre variáveis e a obtenção de combinações lineares entre estas pode contribuir de forma substancial para a identificação de padrões entre os dados, para a redução dimensional e a deteção de variáveis responsáveis pela máxima variabilidade dos sinais, melhorando assim a sua interpretação, Joliffe (2002).
Para obter as componentes principais que explicam a máxima variabilidade dos sinais, desloca-se o sistema de coordenadas para a média x_c=x-x ̅, estabelecem-se as novas direções como combinações lineares das anteriores z_i=u_i'(x-x ̅ ) e selecionam-se os coeficientes u_i que maximizam a variância. Sendo S a estimativa da matriz de covariância Σ, I a matriz unitária d×d, λ_i um escalar e u_i um vetor coluna d×1, as soluções do sistema de equações homogéneas (S-λ_i I) u_i=0 implicam (S-λ I)=0. Esta equação possui d soluções escalares λ_i designados como valores próprios de S que representam as variâncias das novas variáveis z_i. Uma vez resolvido o sistema de equações homogéneas para os diferentes valores próprios λ_i, obtêm-se para cada λ_i uma família de vetores próprios ortogonais u_i u_j=0 de variáveis não correlacionadas, selecionando-se normalmente os vetores próprios com comprimento unitário u_i u_j=1 para constituição de um sistema ortonormado, de acordo com Sá (2003). A redução dimensional dos componentes principais pode ser realizada, entre outros, por seleção dos componentes responsáveis pela explicação de um determinado nível de variância, por exemplo 95%, com base no critério de Gutman-Kaiser, eliminando os valores próprios estandardizados inferiores à unidade, ou com base no teste do scree plot, por representação gráfica dos valores próprios, eliminando os valores próprios em que o declive do gráfico se torna aproximadamente horizontal.
RESULTADOS
Correlação EMGs
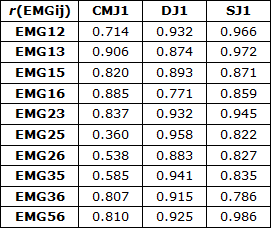
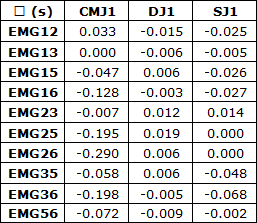
As correlações simples (r) das envolventes EMG, Figura 1 e Tabela 1, permitiram detetar coatividades superiores em DJ relativamente a SJ e estas coatividades superiores a CMJ com maior sinergia em DJ relativamente a SJ e CMJ que apresentam vários ciclos correspondentes a desfasamento temporal de atividade. As correlações simples em DJ foram superiores às registadas em SJ e CMJ para EMG25, EMG26, EMG35 e EMG36 correspondentes às atividades musculares EMG2 do reto femoral, EMG3 do vasto lateral, EMG5 do gastrocnémio lateral e EMG6 do gastrocnémio medial. Já em SJ as correlações simples foram superiores a CMJ e DJ em EMG12, EMG13, EMG23 e EMG56, sendo EMG1 correspondente à atividade muscular do vasto medial. As correlações simples máximas foram obtidas em CMJ para EMG13, em DJ para EMG25 e em SJ para EMG56.
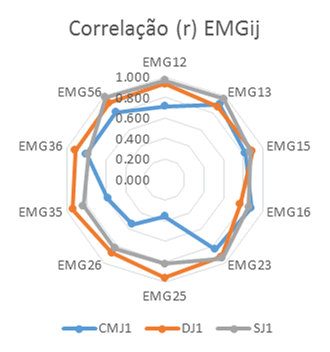
Figura 1. Distribuição radial das correlações simples (r) das atividades EMGij.
Tabela 1. Correlações simples (r) das atividades EMGij em CMJ1, DJ1 e SJ1.
Correlação cruzada EMGs
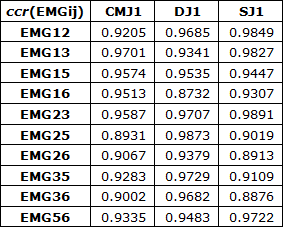
As correlações cruzadas (ccr) das envolventes EMG, Figura 2 e Tabela 2, permitiram também detetar coatividades superiores em DJ relativamente a SJ e ambos coatividades superiores a CMJ. As correlações cruzadas (ccr) em DJ foram superiores às registadas em CMJ e SJ para EMG25, EMG26, EMG35 e EMG36. Já em EMG12, EMG13, EMG23 e EMG56 as correlações registadas em SJ foram superiores às registadas em DJ e CMJ. As correlações cruzadas (ccr) máximas foram obtidas em CMJ para EMG13 com desfasamento nulo, em DJ para EMG25 com desfasamento 19 ms e em SJ para EMG23 com desfasamento 14 ms, Tabela 3 e Figura 4.
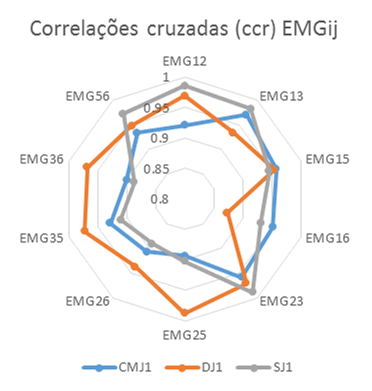
Figura 2. Distribuição radial das correlações cruzadas (ccr) das atividades EMGij.
Tabela 2. Correlações cruzadas (ccr) das atividades EMGij em CMJ1, DJ1 e SJ1.
Correlação simples e cruzada EMGs
As correlações cruzadas (ccr) apresentam valores superiores às correlações simples (r), Figura 3 e Tabelas 1 e 2, indicando que a correlação máxima entre os diferentes sinais EMG ocorre com desfasamento temporal superior nos casos em que r e ccr apresentam maiores diferenças.
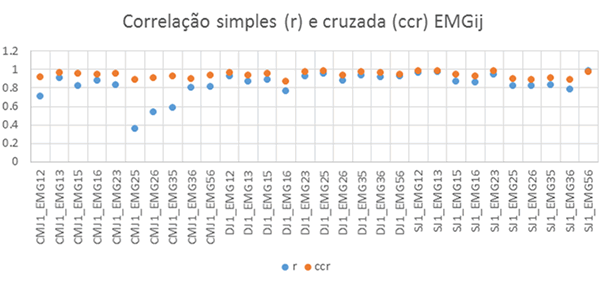
Figura 3. Correlações simples (r) e cruzadas (ccr) das atividades EMGij em CMJ, DJ e SJ.
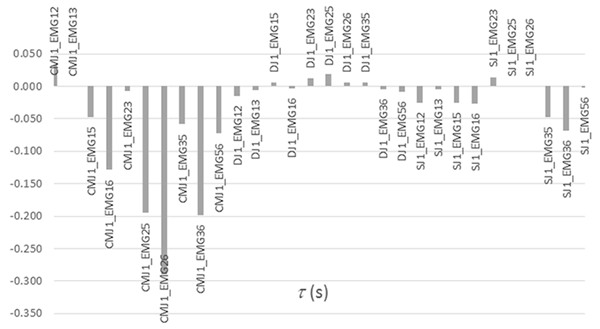
Figura 4. Desfasamento temporal (s) de ccr das atividades EMGij em CMJ1, DJ1 e SJ1.
Tabela 3. Desfasamento temporal (s) de ccr das atividades EMGij em CMJ1, DJ1 e SJ1.
Análise de componentes principais
A análise de componentes principais permitiu detetar um componente principal (PC) explicativo de maior variabilidade de EMG, 92,2% em DJ relativamente a SJ com 90,6% e ambos variabilidade superior a CMJ com 78,7%, sendo o segundo PC responsável pela explicação de maior variabilidade em CMJ com 15,3% relativamente a SJ com 6,7% e ambos variabilidade superior a DJ com 4,9%, Tabela 4. Em termos acumulados são necessários 3 PC para explicação de 97,72% da variabilidade em CMJ e apenas 2 PC para explicação de 97,09% em DJ e 97,28% da variabilidade em SJ. DJ e SJ apresentam também valores próprios i superiores de PC1 relativamente a CMJ, apresentando CMJ valor próprio de PC2 superior a DJ e SJ, Figura 5. No que respeita à distribuição pelos componentes PC1, PC2 e PC3 ilustrados na Figura 6, DJ e SJ apresentam também menor dispersão relativamente a CMJ.
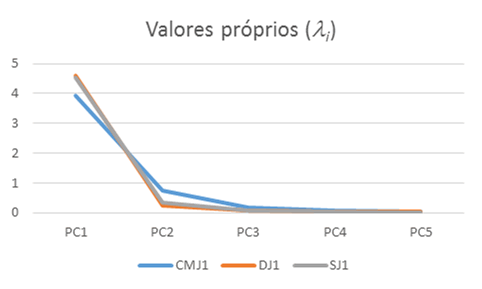
Figura 5. Valores próprios dos componentes principais EMGij em CMJ1, DJ1 e SJ1.
Tabela 4. Variância explicada pelos componentes principais EMGij em CMJ1, DJ1 e SJ1.
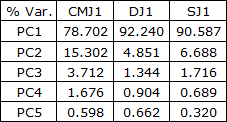
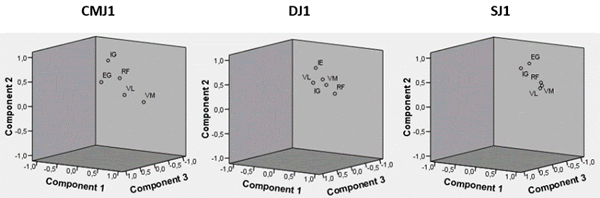
Figura 6. Valores próprios dos componentes principais EMGij em CMJ1, DJ1 e SJ1.
CONCLUSÕES E TRABALHOS FUTUROS
Neste estudo apresentam-se vários trabalhos que apontam para as limitações da anatomia funcional (AF) na sua capacidade de explicar de forma rigorosa a atuação muscular (AM) e para a observação de AM complexas e inesperadas em estudos invasivos (EI). De acordo com a importância da compreensão das AM, procurando colmatar as lacunas da AF e o caráter invasivo dos métodos utilizados principalmente em animais, apresentam-se as vantagens da utilização de EMG de superfície (EMGs), bem como as suas limitações, em virtude da variabilidade inerente aos sujeitos analisados, ao método e à atividade em análise. Atendendo à predominância nas ações naturais do CAE muscular e ao registo em estudos invasivos de comportamentos inesperados nos CAE, a atenção do estudo focou-se neste fenómeno, tendo-se realizado a sua avaliação com recurso a saltos de máxima impulsão vertical (MIV) em virtude da maior expressão dos CAE nos saltos MIV e do seu caráter padrão, reprodutibilidade e simplicidade. Em virtude da variabilidade registada em estudos baseados na análise dos sinais EMGs e do caráter subjetivo das comparações qualitativas, procedeu-se à aplicação de três métodos distintos correspondentes à correlação de Pearson (CR), correlação cruzada normalizada (CCRN) e análise de componentes principais (ACP) dos sinais EMGs, para avaliação da consistência dos métodos aplicados e dos resultados obtidos. Os três métodos revelaram-se consistentes, apresentando correlações superiores em DJ e SJ relativamente a CMJ, tanto pelo método CR como pela CCRN, com maiores diferenças nos casos em que a CCRN máxima ocorre com maior desfasamento temporal, tendo-se encontrado também por aplicação da ACP, 1 PC explicativo da maior variabilidade em DJ e SJ relativamente a CMJ. Assim a CR pode ser aplicada nos casos em que as correlações ocorrem com menor desfasamento temporal, recomendando-se a CCRN nos casos em que se pretende analisar as correlações com maior desfasamento temporal. Já no caso em que o número de músculos a analisar seja superior, recomenda-se a aplicação da ACP em virtude da capacidade de detetar um menor número de fatores não correlacionados explicativos da maior variabilidade dos sinais e eliminando a redundância nos sinais originais fortemente correlacionados. Atendendo ao caráter linear dos métodos utilizados, e apesar das relações encontradas, cuja inexistência impediria a sua deteção, a utilização de métodos não lineares em complemento dos métodos lineares para classificação dos sinais EMGs dos vários músculos selecionados e a deteção de um menor número de fatores relativamente ao número de músculos analisados pode significar que o sistema neuromuscular poderá não utilizar um programa para cada sinal EMG, tal como defendem Vaughan (1999) relativamente ao caminhar e Rodrigues et al. (2012) para os saltos de máxima impulsão vertical.
Referencias
1. Aragón-Vargas, L. F., Gross, M. M. (1997). Kinesiological factors in vertical jump performance: differences among individuals. Journal of Applied Biomechanics, 13, 24-44.
2. Asmussen, E., Bonde-Petersen, F. (1974). Storage of elastic energy in skeletal muscles in man. Acta Physiol. Scand., 91, 385-392.
3. Basmajian, J. V., & De Luca, C. J. (1985). Muscles alive: Their functions revealed by electromyography (5th. Ed.). Baltimore: Williams & Wilkins.
4. Bartlett, R. (1999). Sports biomechanics: Reducing injury and improving performance. London: E&FN Spon.
5. Bernstein, N. A. (1967). The co-ordination and regulation of movements. Oxford: Pergamon Press.
6. Blazevich, A. (2011). The Stretch-Shortening Cycle (SSC). In M. Cardinale, R. Newton, R. & K. Nosaka (Eds.). Strength and Conditioning: Biological Principles and Practical Applications (Section II, Chap. 2.8, pp. 209-221). West Sussex, UK: Wiley-Blackwell.
7. Bobbert, M. F., Mackay, M., Schinkelshoek, P. A., Huijing, & van Ingen Schenau, G.J. (1986). Biomechanical analysis of drop and countermovement jumps. Eur J Appl Physiol, 54, 566-573.
8. Bobbert, M. F., van Ingen Schenau, G.J. (1988). Coordination in vertical jumping. Journal of Biomechanics, 21, 249-262.
9. Bobbert, M. F., Gerritsen, K. G. M., Litjens, M. C. A., & van Soest, A. J. (1996). Why is countermovement jump height greater than squat jump height? Medicine and Science in Sports and Exercise, 28, 1402-1412.
10. Bobbert, M. F., Casius L. J. R. (2011). Spring-like leg behaviour, musculoskeletal mechanics and control in maximum and submaximum height human hopping. Phil. Trans. R. Soc. B., 366, 1516-1529.
11. Cavagna, G. A., Saibene, P. F., & Margaria, R. (1965). Effect of negative work on the amount of positive work performed by an isolated muscle. Journal of Applied Physiology, 20, 157-158.
12. Cavagna, G. A., Komarek, L., Citterio, G., & Margaria, R. (1971). Power output of the previously stretched muscle. In Medicine and Sport, 6. Biomechanics II, 159-167.
13. Gregor, R. J. (Ed.) (1997). Mechanics and energetics of the stretch-shortening cycle [special issue]. Journal of Applied Biomechanics, 13(4).
14. Gowitzke, B.A. (1984). Muscles alive in sport. In M. Adrian & H. Deutsch (Eds.), Biomechanics. Eugene, Ore.: Microform.
15. Hamill, J., Knutzen, K.M. (2009). Biomechanical basis of human movement (3rd Ed. ). Philadelphia, PA: Lippincott Williams & Wilkins
16. Herzog, W., & Leonard, T. R. (2000). The history dependence of force production in mammalian skeletal muscle following stretch-shortening and shortening-stretch cycles. Journal of Biomechanics, 33, 531-542.
17. Jolliffe, I. T. (2002). Principal Component Analysis (2. ed). New York, NY: Springer-Verlag.
18. Knudson, D. (2007). Fundamentals of Biomechanics (2. ed). New York, NY: Springer.
19. Komi, P. V., & Bosco, C. (1978). Utilization of stored elastic energy in leg extensor muscles by men and women. Medicine and Science in Sports, 10(4), 261-265.
20. Komi, P. V., Belli, A., Huttunen, V., Bonnefoy, R., Geyssant, A., & Lacour, J. R. (1996). In vivo dynamics of human medial gastrocnemius muscle-tendon complex during stretch-shortening cycle exercise. Acta Physiological Scandinavica, 170, 127-135.
21. Komi, P. V., & Golhofer, A. (1997). Stretch reflexes can have an important role in force enhancement during SSC exercise. Journal of Applied Biomechanics, 13, 451-460.
22. Komi, P.V., Ishikawa, M., & Linnamo, V. (2011). Identification of Stretch-Shortening Cycles in different sports. In Vilas-Boas, Machado, Kim, Veloso (eds.). Portuguese Journal of Sports Sciences, 11(2), 31-33: Biomechanics in Sports.
23. Kubo, K., Kanehisa, H., Takeshita, D., Kawakami, Y., Fukashiro, S., & Fukunaga, T. (2000). Optic fibre as a transducer of tendomuscular forces. Eur. J. of App. Physiol., 72, 278-280.
24. Kurokawa, S., Fukunaga, T., Nagano, & Fukashiro, S. (2003). Interaction between fascicles and tendinous structures during counter movement jump investigated in vivo. Journal of Applied Physiology, 95, 2306-2314.
25. Marey, M., & Demeny, G. (1885). Locomotion humaine, mécanisme du saut. Comptes endus des séances de l’Académie des Sciences, 101, 485-494.
26. Margaria, R., Aghemo, P. & Rovelli, E. (1966). Measurement of muscular power (anaerobic) in man. Journal of Applied Physiology, 21, 1662-1664.
27. Perry, J., Burnfield, J. M. (2010). Gait Analysis: Normal and Pathological Function. Thorofare, NJ: SLACK Incorporated
28. Richmond, F. J. R. (1998). Elements of style in neuromuscular architecture. American Zoologist, 38, 729-742.
29. Rodrigues, C. M. B., Correia, M. V., Abrantes, J. M. C. S., & Nadal, J. (2012). Componentes principais da actividade EMG nos músculos dos membros locomotores humanos. Proceedings of XXIII Congresso Brasileiro de Engenharia Biomédica, 317-321.
30. Roberts, T. J., Marsh, R. L., Weyand, P. G., & Taylor, D. R. (1997). Muscular force in running turkeys: The economy of minimizing work. Science, 275, 1113-1115.
31. Sá, J. P. M. (2003). Applied Statistics using SPSS, STATISTICA and MATLAB. Berlin Heidelberg: Springer Verlag.
32. Sargent, D. A. (1921). The physical test of a man. Americ. Physical Ed. Rev., 26, 188-194.
33. Van Ingen Schenau, G. J., van Soest, A. J., Gabreels, F. J. M., & Horstink, M. (1995). The control of multi-joint movements relies on detailed internal representation. Human Movement Science, 14, 511-538.
34. Van Soest, A.J., Schwaab, A.L., Bobbert, M.T., e van Ingen Schenau, G.J. (1993). The influence of the biarticularity of the gastrocnemius muscle on vertical jumping achievement. Journal of Biomechanics, 26, 1-8
35. Vaughan,C. L., Davis, B. L. and O’Connor, J. C. (1999). Dynamic of Human Gait. Cape Town, South Africa: Kiboho Publishers
36. Weineck, J. (2008). Sportanatomie. Balingen, Germany: Spitta Verlag GmbH & Co. KG
37. Winter, D. A. & Yack, H. J. (1987). EMG profiles during normal human walking: stride-to-stride and inter-subject variability. Electroencephalography and clinical Neurophysiology,67, 402-411.
38. Winter, D.A. (2009). Biomechanics and Motor Control of Human Movement. (4a ed.). New Jersey: John Willey & Sons.
39. Zajac, F. E. (1991). Muscle coordination of movement: A perspective. Journal of Biomechanics, 26(S1), 109-124.
40. Zatsiorsky,V.M. & Kraemer, W.J., (2006). Science and Practice of Strength Training. Champaign, IL: Human Kinetics.