
Efecto de la Adaptación a las Grasas a Corto Plazo sobre el Entrenamiento de Alta Intensidad
Louise M Burke, John A Hawley, Nigel K Stepto, Andrew L Carey, Heidi M Staudacher y Nicola K Cummings
0Exercise Metabolism Group, School of Medical Sciences, Faculty of Health Sciences, RMIT University, Bundoora, 3083, Australia.
0Sports Science and Sports Medicine, Australian Institute of Sport, Belconnen 2616, Australia.
Artículo publicado en el journal PubliCE, Volumen 0 del año 2013.
Publicado 4 de febrero de 2013
Resumen
Palabras clave: Ciclismo, entrenamiento intervalado, dieta con alto contenido de grasas
INTRODUCCION
El efecto de la adaptación a las grasas sobre la capacidad de realizar ejercicio aeróbico de alta intensidad no está claro (5,13,14,16,19). Sin embargo, esta estrategia nutricional tiene el potencial de aumentar la resistencia sustancialmente a través de los cambios inducidos por la dieta en los patrones de utilización de combustibles, que promueve la oxidación de grasas y la reposición del glucógeno muscular (5). De hecho, nosotros hemos informado previamente que 5 días de una elevada ingesta de grasas junto con entrenamiento de resistencia prolongado duplican la tasa de oxidación de grasas durante el ciclismo de moderada intensidad (5).
Inmediatamente antes de los eventos de resistencia importantes, los atletas de competición frecuentemente realizan entrenamiento (24) y prácticas nutricionales (10) especializados. Durante este período, se aumenta la intensidad de entrenamiento, con una reducción concomitante en el volumen (24). Los atletas de resistencia también pueden escoger realizar una dieta corta (3-5 días) con elevado contenido de grasa, seguida por un régimen de consumo de carbohidratos tradicional en los 2 días previos al evento (9). Sin embargo, en los individuos desentrenados, la ingesta de una dieta con alto contenido de grasas durante 3-5 días afecta la capacidad de realizar ejercicios, incluso cuando la intensidad del entrenamiento es tan baja como 70% del consumo de oxígeno máximo (VO2máx) (6,7). Nosotros también hemos observado previamente que, en comparación con una dieta rica en CHO, los atletas altamente entrenados con una dieta con elevado contenido de grasas, experimentan síntomas de letargo y mayor fatiga que son particularmente evidentes durante las sesiones de entrenamiento de alta intensidad (5).
Lamentablemente, en nuestra investigación previa, no cuantificamos las demandas metabólicas del entrenamiento de alta intensidad ni las respuestas subjetivas de los atletas a los diferentes regímenes dietéticos (5). Otros investigadores que también han estudiado el efecto de adaptación a las grasas en el metabolismo y rendimiento tampoco supervisaron las respuestas de entrenamiento de sus sujetos moderadamente entrenados (16) o bien entrenados (19). Si los atletas se benefician de regímenes nutricionales/entrenamiento específicos, es necesario que la intensidad del entrenamiento pueda mantenerse mientras consumen una dieta rica en grasas (12). Por consiguiente, el objetivo de ésta investigación fue determinar si los atletas de resistencia competitivos pueden realizar sesiones de entrenamiento de alta intensidad, típicas de los deportistas que se incorporan a una puesta a punto, mientras consumen una dieta con elevado contenido de grasas.
MÉTODOS
Sujetos
Siete ciclistas competitivos de sexo masculino altamente entrenados o triatletas (edad 24 ± 6 años, peso 75,3 ± 5,8 kg, Consumo de oxígeno máximo (VO2max) 5,0 ± 0,5 Lmin-1, producción de potencia aeróbica máxima (PPO) 404 ± 40 W, Media±SD) con antecedentes (>4 años) de entrenamiento de resistencia regular (20 ± 6 h sem-1) fueron reclutados para participar en este estudio. Los sujetos fueron informados de los procedimientos experimentales y posibles riesgos antes de proporcionar su consentimiento por escrito siguiendo las recomendaciones del Comité de Ética de Investigaciones Humanas de la Universidad de RMIT. Los participantes eran atletas de nivel nacional o internacional (tres de ellos habían competido en el Campeonato Mundial Triathlon Ironman de 2000 en Hawai a pocas semanas de realizar este estudio).
Determinaciones Preliminares
Cada sujeto realizó un test de ciclismo máximo incremental hasta el agotamiento en una bicicleta ergométrica con freno magnético (Lode Excalibur, Groningen, Países Bajos). El protocolo del test (11) y los procedimientos de recolección de gases (4) han sido descriptos previamente en detalle. Los resultados del test de máxima intensidad se utilizaron para determinar las producciones de potencia que correspondían a 65% y 85% de VO2max, que fueron las intensidades seleccionadas para las sesiones de entrenamiento en laboratorio (descriptas a continuación).
Protocolo experimental
Después de un día (d-0) de dieta estandarizada (58% CHO: 9,7 g de CHO kg-1 d-1, 27% grasa, 15% de proteínas; energía total 0,25 MJ kg-1 d-1) y de entrenamiento suave (2-4 h de ciclismo de ruta de intensidad submáxima), los participantes comenzaron dos tratamientos dietarios de 3 días siguiendo un diseño cruzado aleatorizado, con un período de limpieza de 18 días entre cada dieta. Cada sujeto o fue asignado a una dieta alta en grasas (>65% de energía; HIFAT), baja en CHO (<20% de energía) o a una dieta isoenergetica (0,25 MJkg-1 BM) alta en CHO (70-75% de energía; HICHO), baja en grasas (<15% de energía). La ingesta de energía en cada dieta fue estimada, en base a los informes proporcionados por los mismos participantes de ingesta habitual y a cálculos factoriales de gasto de energía anticipado, para mantener el equilibrio energético de los sujetos. Se estimaron las dietas habituales de estos sujetos para que fueran similares en la composición de macronutrientes a la dieta normal (es decir, aprox. 10 g kg-1d-1 de CHO, proporcionando 60% de energía). Las dietas fueron diseñadas para aumentar al máximo, o por lo menos igualar, la energía absorbible; la ingesta de fibra se mantuvo en una ingesta diaria media de 50g y se mantuvo en 5-10 g en los días entre los tratamientos. Los alimentos que contenían carbohidratos con un índice glucémico muy bajo o con un elevado contenido de almidón resistente fueron evitados. Se proporcionaron todas las comidas y bocadillos a los sujetos, con dietas individualizadas según las preferencias de alimentos. Por lo menos una comida diaria fue consumida bajo supervisión en el laboratorio y los alimentos restantes para cada período de 24-h se proporcionaron en paquetes previamente preparados. Los sujetos realizaron un registro de alimentos, en el cual debían informar todos los alimentos y bebidas ingeridos diariamente para aumentar al máximo el cumplimiento de las dietas prescritas. Los alimentos no consumidos fueron devueltos y se descontaron de la ingesta total de ese día. Además, diariamente se llevaron archivos de entrenamiento en un diario proporcionado a cada sujeto para asegurar que el entrenamiento fuese similar durante ambos períodos de intervención dietética. Aunque los sujetos eran conscientes del tratamiento que estaban recibiendo (porque no era completamente posible enmascarar las dietas), el investigador responsable de la recolección de los datos desconocía el orden de tratamientos.
Para determinar el efecto de los tratamientos dietéticos en el metabolismo de los sustratos durante el ejercicio, se establecieron dos sesiones de entrenamiento intervalado de alta intensidad en cada programa de entrenamiento de los atletas. Estas sesiones estandarizadas se realizaron bajo vigilancia en el laboratorio en una bicicleta ergométrica marca Lode. La primera sesión de entrenamiento intervalado se realizó después de un ayuno de toda la noche y antes del comienzo de cada intervención dietética (d-1). El objetivo de esta sesión fue causar un marcado descenso en la concentración del glucógeno muscular (25) y comenzar una diferenciación rápida entre tratamientos dietéticos basados en la capacidad de los sujetos de reponer el glucógeno consumido. Una segunda sesión de laboratorio se realizó en el último día (d-4) de cada tratamiento (es decir, después de 3 d de intervención dietética), también después de un ayuno de toda la noche.
El día de la sesión de entrenamiento intervalado, los sujetos arribaron al laboratorio entre las 07:00 y 08:00 h después de un ayuno de toda la noche de 10-a 12-h. Fueron pesados y se les insertó una cánula de Teflón (20 G, OptivaTM, Johnson & Johnson Intl., Bruselas, Bélgica) en la vena antecubital del antebrazo para la obtención de muestras de sangre continua rápida a través de una llave de cierre estéril (válvula Safsite, B. Braun Médical Inc., Lakeside, PA). En este momento, se obtuvo una muestra de 10-mL de sangre. Después de cada extracción, la cánula fue llenada con 1-2 mL de solución salina estéril 0,9% para mantener la vena despejada. Después de descansar calladamente sentados durante 10 min, los sujetos se subieron a la bicicleta ergométrica y comenzaron a pedalear para entrar en calor durante 20-min con una carga de trabajo constante que produjo aproximadamente 65% de VO2max. Se tomó una muestra de sangre adicional durante los últimos 60 seg de este ejercicio de pedaleo. Después de un descanso de 10-min durante el cual los sujetos permanecieron sentados en la bicicleta ergométrica, los sujetos comenzaron la sesión de entrenamiento intervalado que consistió en series de trabajo (8 x 5 min) a 86 ± 2% VO2max (323 ± 32W) con un período de recuperación activa de 60-s (100 W) entre las series (relación trabajo: descanso de 5:1). Una sesión de entrenamiento de este tipo es típica de un entrenamiento realizado por atletas de ultra resistencia durante un entrenamiento de puesta a punto (Hawley JA, observaciones inéditas). Las condiciones del laboratorio fueron mantenidas a 20°C y 46% humedad relativa. Los sujetos fueron refrescados con un ventilador (velocidad del viento de aprox.7 ms-1) y se les proporcionó agua sin límites a lo largo de la sesión de entrenamiento. Durante los últimos 10 min del pedaleo de 20-min y a lo largo de las series de trabajo 1, 4, y 8 de la sesión de entrenamiento intervalado, se evaluaron los gases pulmonares. Al finalizar la 1ra, 4ta y 8va repeticiones se obtuvieron muestras de sangre (~10 mL) y se monitoreó la frecuencia cardíaca (HR) continuamente por medio de telemetría (Accurex Plus; Polar Electro OY, Kempele, Finlandia). Al finalizar cada serie de trabajo se registraron los índices de esfuerzo percibido subjetivos (RPE) para las piernas en una escala de 6-20 (2).
En los días 2 y 3 de cada tratamiento dietético, los sujetos mantuvieron su entrenamiento normal el cual fue registrado en un libro de registros. Inmediatamente luego de finalizar una sesión de entrenamiento, los sujetos clasificaron el entrenamiento según una escala de 7 puntos dónde 1 indicaba que el sujeto “se había sentido bien durante todas las partes del entrenamiento" y 7 indicaba que se "había sentido terrible a lo largo del entrenamiento." Durante la intervención dietética cruzada, los sujetos reprodujeron su programa de entrenamiento externo al laboratorio. Al final del d-1 y d-4 de cada tratamiento, los sujetos completaron el cuestionario de Perfil de Estado de Animo (POMS) (17) para establecer cualquier cambio psicológico asociado con la intervención dietética.
Análisis
Tasas de oxidación de sustratos.
Las tasas de oxidación de CHO y de grasas (g-min-1) del cuerpo entero se calcularon a partir de los valores de VCO2 y VO2 utilizando los valores de RER no proteicos (18). En este sentido, asumimos que la cantidad de proteína oxidada es pequeña y que otros procesos metabólicos que involucran la producción y/o utilización de O2 y CO2 (por ejemplo, gluconeogenesis a partir de proteínas, formación de cuerpos cetónicos y lipogénesis) son despreciables en comparación con la oxidación de glucosa y de ácidos grasos (18). Tales suposiciones parecen razonables: incluso cuando consumían la dieta de rica en grasas, los sujetos estaban ingiriendo ~200 g CHO d-1. Estas ecuaciones se basan en la premisa que VO2 y VCO2 reflejan con precisión el consumo de O2 y producción de CO2 de los tejidos. En los sujetos altamente entrenados similares a los que participaron en la presente investigación, se ha demostrado previamente que la calorimetría indirecta es un método válido para cuantificar las tasas de oxidación de los sustratos durante el ejercicio activo a ~85% de VO2max (21). Las tasas de oxidación de grasas (µmol kg-1 min-1) se determinaron convirtiendo la tasa de oxidación del triacilglicéridos en g-min-1 a su equivalente molar, asumiendo que el peso molecular promedio de un triacilglicérido humano es 855,3 gmol-1 y multiplicando la tasa de oxidación molar del triglicérido por 3, porque cada molécula contiene 3 moles de ácido graso. Las tasas de oxidación de CHO (µmol kg-1 min-1) se determinaron convirtiendo la tasa de oxidación de CHO en g min-1 a su equivalente molar.
El gasto de energía total durante el las sesiones de entrenamiento en el laboratorio fue estimado a partir de los valores de VO2 y VCO2 en estado estable asumiendo un valor de 37,5 kJ g-1 y 16,9 kJ g-1 para las grasas y CHO, respectivamente.
Análisis de los metabolitos en sangre
Las muestras de sangre (~3 mL) fueron analizadas inmediatamente para determinar las concentraciones de glucosa y lactato utilizando un analizador automatizado Stat Plus 2300 (Yellow Springs Instruments, Yellow Springs, OH). Una alícuota de 3 mL de sangre fue colocada en un tubo que contenía un conservante que consistía en etilen glicol-bis (ß-aminoetil éter)-N, N, N, N'- ácido tetraacético y glutatión reducido en solución salina normal, fue suavemente mezclada y centrifugada en una centrífuga (J6-MC Beckman Instruments Inc., Yellow Spring, CA), a 4500 revoluciones durante 8 min. Luego se analizó el plasma para determinar la concentración de ácidos grasos libres (FFA) por medio de un método enzimático colorimétrico (Wako, NEFAC code 279-75409, Tokio, Japón). La sangre restante fue colocada en un tubo con heparina de litio y fue centrifugada a 4500 revoluciones durante 15 min; luego 500 µL de plasma fueron transferidos a un tubo con 500 µL de ácido perclórico 3M enfriado con hielo, el mismo fue agitado vigorosamente en un agitador magnético y fue centrifugado; 800 µL de sobrenadante fueron transferidos a un tubo con 200 µL de hidróxido de potasio 6 M, fueron mezclados y centrifugados a 10000 revoluciones durante 5 min. El sobrenadante resultante fue analizado para evaluar la concentración plasmática de glicerol por medio de un análisis enzimático fluorométrico (20).
Análisis estadístico
Los datos de las sesiones de entrenamiento en el laboratorio fueron analizados usando un ANOVA de mediciones repetidas (dieta x día x tiempo). El volumen de entrenamiento y RPE durante las sesiones de entrenamiento en las dos dietas diferentes, fueron analizados con el Test-t de muestras apareadas. Todos los análisis fueron realizados usando el software Statistica para Windows versión 5,1 (StatSoft Inc., 1997, Tulsa, OK). Todos los datos se expresan en forma de Media±SD y la significancia se fijó en P <0,05.
RESULTADOS
Cumplimiento de la dieta y del entrenamiento
Los sujetos cumplieron de manera excelente con los dos tratamientos dietéticos: las ingestas medias de CHO informadas del d-1 al d-4 fueron 11,00 ± 0,03 y 2,60 ± 0,07 g kg-1 d-1 para HICHO y HIFAT, respectivamente y tal como se pretendía, fueron significativamente diferentes (P <0,001). Las ingestas de grasas informadas para el mismo período fueron 1,01 ± 0,03 g kg-1 d-1 para HICHO y 4,62 ± 0,23 g kg-1 d-1para HIFAT (P <0,001). El tiempo total de entrenamiento fuera del laboratorio cumplido durante los períodos de l intervención fue 820 ±201 y 788 ± 196 min para HICHO y HIFAT, respectivamente (NS). De estos, 608 ± 198 y 578 ± 172 min fueron destinados a ciclismo (NS) y 210 ± 176 y 210 ± 172 min fueron destinados a "otros entrenamientos" (natación, carrera o sobrecarga) en HICHO y HIFAT, respectivamente (NS).
Frecuencia cardíaca, índice de esfuerzo percibido durante el entrenamiento y perfil de estado de ánimo (POMS)
Los índices de esfuerzo percibido durante el entrenamiento fuera del laboratorio fueron mayores tanto para el ciclismo (P <0,05) como para todos los "otros entrenamientos" (P <0,01) durante la dieta HIFAT en comparación con la dieta HICHO. No se observaron diferencias en las puntuaciones globales de POMS en día d-1 en los dos tratamientos dietéticos (143 ± 71 y 134 ± 120). Sin embargo, los valores de POMS fueron mayores en el d-4 de la dieta HIFAT (232 ± 89) en comparación con el d-4 de la dieta HICHO (94 ± 90, P <0,01). En el d-1 de ambos tratamientos dietéticos, las puntuaciones individuales de POMS para la fatiga fueron similares (42 ± 21 contra 35 ± 20 para HICHO y HIFAT, respectivamente) pero fueron mayores en el d-4 de HIFAT (66 ± 18, P <0,01).
La HR durante el ejercicio de ciclismo de 20-min antes de cada sesión de entrenamiento intervalado en el laboratorio presentó un valor medio de 130 lat min-1 y no fue diferente entre los tratamientos dietéticos. Durante las sesiones de entrenamiento intervalado, la HR aumentó típicamente de 155 lat min-1 al final de la primera serie de trabajo, a ~165 lat min-1 al final de la última repetición: este aumento fue similar en las condiciones de tratamiento. El RPE (piernas) fue similar en el d-1 para HICHO y HIFAT (14,1 ± 1,4 y 14,8 ± 1,5) y en el d-4 de HICHO (13,8 ± 1,8). Sin embargo, el RPE fue significativamente mayor en el d-4 de HIFAT (16,0 ± 1,3) en comparación con todas las otras condiciones (P <0,05). Un participante no pudo realizar la sesión de entrenamiento en el laboratorio en el d-4 de la dieta de HIFAT. En este caso, él realizó las primeras cuatro series de trabajo a la intensidad prescrita (85% de VO2max) pero completó las últimas cuatro repeticiones a una tasa de trabajo reducida (~70% de VO2max).
Medición de los gases pulmonares y oxidación de combustibles.
La Tabla 1 resume las mediciones de gases pulmonares tomadas durante el ejercicio de ciclismo de 20-min a 65% de VO2max y a lo largo de las series de trabajo 1, 4, y 8 de las sesiones de entrenamiento intervalado realizadas a 86 ± 2% de VO2max. El VO2 fue similar en todos los ejercicios de ciclismo de 20 min (~3,2 L min-1), aumentando a ~4,4 ± 0,3 L min-1durante las series de trabajo de alta intensidad (P <0,001). Los valores de RER que habían sido similares durante los ejercicios de ciclismo de 20-min en ambos días en el tratamiento HICHO y en el d-1 de la dieta HIFAT, fueron menores en el d-4 de la dieta rica en grasas (0,79 ± 0,03 contra 0,85 ± 0,03, P <0,01). Durante las series de trabajo de alta intensidad, RER disminuyó de la serie 1 a la serie 8 tanto en HICHO y como en el d-1 de HIFAT (P <0,01). Sin embargo, en el d-4 de HIFAT, RER fue menor en todas las series de trabajo en comparación con los puntos de tiempo similares para todas las otras condiciones (P <0,01) y no varió significativamente de la primera a la última serie de trabajo.
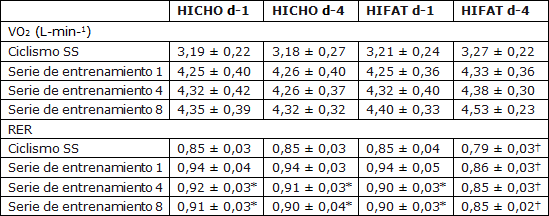
Tabla 1. Mediciones de intercambio de gases pulmonares durante las sesiones de ejercicio realizadas en el laboratorio. Todos los valores se presentan en forma de Media ±SD. N=7. VO2=Consumo de oxígeno; HICHO= Dieta con elevado contenido de carbohidratos; HIFAT=Dieta con elevado contenido en grasas; Ciclismo SS= Ejercicio de ciclismo de 20 min a una carga de trabajo constante correspondiente a 65% del VO2max. *Presenta diferencias significativas con la serie de trabajo 1 (P <0,01). † Presenta diferencias significativas con HICHO d-1, HICHO d-4, y HIFAT d-1 (P <0,01).
La Figura 1 muestra las tasas de oxidación de CHO (Panel A) y la oxidación de grasas (Panel B) determinadas durante los ejercicios de ciclismo de 20 min y las series de trabajo 1, 4, y 8 de la sesión de entrenamiento intervalado. Las tasas de oxidación de CHO fueron menores (P <0,01) durante el ejercicio de ciclismo de 20 min y en cada serie de trabajo de la sesión de entrenamiento en el d-4 de HIFAT en comparación con todas las otras condiciones. Por consiguiente, las tasas de oxidación de las grasas (Panel B) fueron significativamente mayores (P <0,05) en el d-4 de HIFAT para el ejercicio de ciclismo de 20-min y a lo largo de las series de trabajo 1, 4, y 8 en comparación con todas las otras medidas. La Figura 2 muestra la contribución de la oxidación de las grasas y de CHO con el gasto total de energía durante el ciclismo de 20-min y el valor promedio a los largo de las series de trabajo 1, 4, y 8 de la sesión de entrenamiento intervalado de alta intensidad. El gasto de energía total no se vio afectado por el tratamiento dietético ni durante el ejercicio de ciclismo de 20 min a 65% de VO2max (~880 J kg min-1) ni durante las sesiones de entrenamiento de alta intensidad (~1225 J kg min-1; Figura. 2). Sin embargo, la contribución de la oxidación de las grasas con el gasto de energía total aumentó de 45,8 ± 11,1% a 66,4 ± 11,9% (P <0,05) durante el ciclismo de 20 min y de 22,3 ± 9,8 a 43,1 ± 8,5% durante el ejercicio a 85% de VO2max (P <0,05; Figura 2) en el d-4 de la dieta con elevado contenido de grasas.
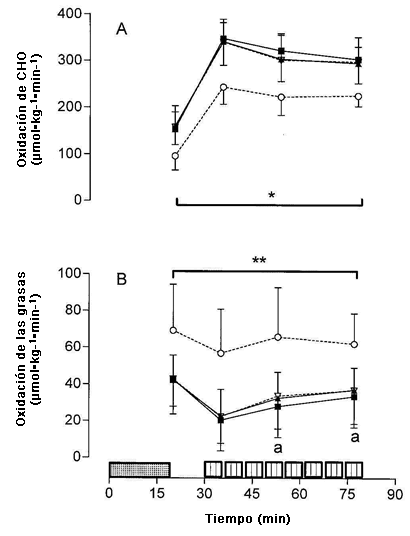
Figura 1. Tasa de oxidación de carbohidratos (Panel A) y de grasas (Panel B) determinadas durante el ejercicio de ciclismo de 20 min a 65% del consumo de oxígeno máximo y luego de las series de trabajo 1, 4 y 8 del entrenamiento intervalado de alta intensidad, para el d-1 de la dieta rica en carbohidratos (▲), d-4 de la dieta rica en carbohidratos (■), d-1 de la dieta rica en grasas (◊) y d-4 de la dieta rica en grasas (○); N=7. *Se observó un efecto significativo principal de la dieta P <0,05; ** Se observó un efecto significativo principal de la dieta, P <0,01;a Significativamente mayor que la serie de trabajo 1, P <0,05.
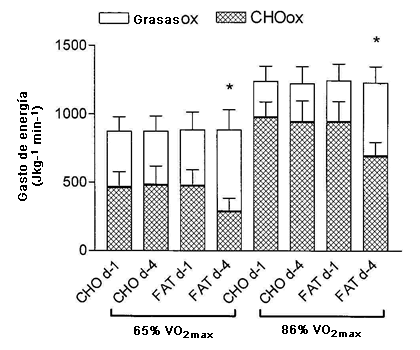
Figura 2. Gasto de energía determinado durante la entrada de calor de 20 min a 65% del consumo de oxígeno máximo y durante el entrenamiento intervalado de alta intensidad realizado a 86% del consumo de oxígeno máximo para el d 1 y d 4 de las intervenciones dietarias. * Presenta diferencias significativas con el d1 y d4 de la dieta alta en carbohidratos y el d1 de la dieta alta en grasas, P<0,05.
Parámetros de la sangre y del plasma.
La Figura 3 muestra la concentración de glucosa (Panel A) y la concentración del lactato en sangre (Panel B) en reposo, al final del ejercicio de ciclismo de 20 min, e inmediatamente al finalizar las series de trabajo 1, 4, y 8 de las sesiones de entrenamiento intervalado. La concentración de glucosa sanguínea permaneció en todo momento entre 3,8 y 4,5 mmol l-1 y no fue diferente entre los tratamientos dietéticos o con el tiempo (Figura 3A). Se observó una interacción dieta x día x tiempo (P <0,05) tal que la concentración de lactato sanguíneo durante la sesión de entrenamiento intervalado fue menor en el d-4 de HIFAT en comparación con todos los otros puntos de tiempo (Figura 3B).
Las concentraciones plasmáticas de FFA (Panel A) y glicerol plasmático (Panel B) en reposo, después del ejercicio de ciclismo de 20 min e inmediatamente después de las series de trabajo 1, 4 y 8 de la sesión de entrenamiento intervalado se presentan en la Figura 4. En comparación con el d-1, las concentraciones plasmáticas de FFA fueron mayores en el d-4 para las intervenciones dietéticas HICHO y HIFAT (P <0,01, Figura 4A). Se observó una interacción significativa dieta x día x tiempo (P <0,05) para el glicerol plasmático. La concentración de glicerol plasmático (Panel B) se incrementó con el tiempo de modo que la concentración fue más alta en el d-4 de HIFAT después del ejercicio de ciclismo de 20 min y después de las series de trabajo 4 y 8 de la sesión de entrenamiento de alta intensidad (P <0,01). La concentración plasmática de glicerol también fue más alta después de las series de trabajo 4 y 8 en comparación con la primera repetición en el d-4 de HIFAT (P <0,01; Figura 4B).
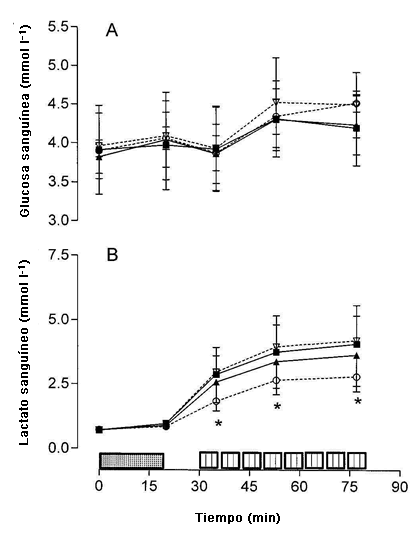
Figura 3. Concentraciones de glucosa (A) y lactato (B) sanguíneos determinados antes y después del ejercicio de ciclismo de 20 min a 65% del consumo de oxígeno máximo y luego de las series de trabajo 1,4 y 8 de la sesión de entrenamiento intervalado de alta intensidad para el d-1 de la dieta rica en carbohidratos (▲), d-4 de la dieta rica en carbohidratos (■), d-1 de la dieta rica en grasas (◊) y d-4 de la dieta rica en grasas (○). * Valores significativamente menores que los obtenidos en los mismos puntos de tiempo del d 1 y d 4 de la dieta alta en carbohidratos y el d 1 de la dieta alta en grasas, P < 0,05.
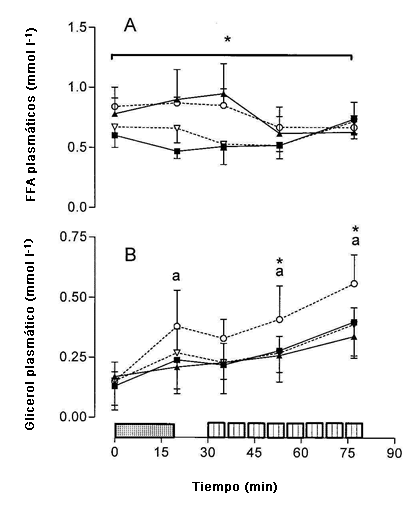
Figura 4. Concentraciones plasmáticas de FFA (A) y glicerol (B) determinados antes y después del ejercicio de ciclismo de 20 min a 65% del consumo de oxígeno máximo y luego de las series de trabajo 1,4 y 8 de la sesión de entrenamiento intervalado de alta intensidad para el d-1 de la dieta rica en carbohidratos (▲), d-4 de la dieta rica en carbohidratos (■), d-1 de la dieta rica en grasas (◊) y d-4 de la dieta rica en grasas (○). * Valores significativamente diferentes de los obtenidos en la serie 1, P < 0,01. a Significativamente mayor que los valores de d 1 y d 4 de la dieta alta en carbohidratos y el d 1 de la dieta alta en grasas, P < 0,01.
DISCUSIÓN
Las mejoras en la capacidad de resistencia durante el entrenamiento a largo plazo (7 semanas) son notablemente perjudicadas cuando los sujetos desentrenados ingieren una dieta rica en grasas en comparación con una dieta con elevado contenido de carbohidratos (13). A corto plazo (<6 d) la adaptación a una dieta con elevado contenido de grasas y bajo contenido de carbohidratos también es perjudicial para el rendimiento de resistencia submáxima en individuos desentrenados (9,12,13). Phinney et al. (19) también propusieron que habría una limitación en la intensidad del ejercicio que podría ser manejada por "ciclistas de élite" después de una dieta con elevado contenido de grasas. Sin embargo, como se ha señalado recientemente (12), es fundamental para los atletas de competición que la capacidad de entrenamiento no se vea afectada en las fases de pre-competencia o de puesta a punto cuando se siguen prácticas nutricionales (10) y/o de entrenamiento (24) especializadas. Así, el primer hallazgo del presente estudio fue que los atletas de resistencia de competición altamente entrenados pudieron realizar sesiones de entrenamiento intervalado aeróbico de alta intensidad, mientras consumían una dieta con elevado contenido de grasas, y bajo contenido en carbohidratos. Según nuestros conocimientos, las tasas de trabajo absolutas (~325 W) y relativas (86% de VO2max) mantenidas por nuestros sujetos durante las sesiones de entrenamiento en el laboratorio están entre las más altas informadas en la literatura para personas que consumían una dieta con elevado contenido de grasas.
Las elevadas tasas de oxidación de grasas fueron proporcionales a las impresionantes tasas de trabajo absolutas sostenidas por nuestros sujetos durante el entrenamiento intervaldo de alta intensidad después de 4 d de adaptación a las grasas. Previamente, nosotros informamos una tasa de oxidación de grasas de todo el cuerpo de 57 µmol kg-1 min-1 para los atletas de resistencia de competición (VO2 pico 65 mL kg-1.min-1) que realizaron ejercicios de ciclismo a 70% de VO2max (234 W) después de 5 días de una dieta rica en grasas (5). En concordancia con estos valores, las tasas de oxidación de grasas para los sujetos en la investigación presente fueron 69 ± 25 µmol kg-1 min-1 durante el ejercicio de ciclismo a 65% de VO2max (232 ± 23 W). Sin embargo, a pesar de un aumento en la intensidad del ejercicio de 65% de VO2max a 85% de VO2max (323 ± 32 W), las tasas de oxidación de grasas no disminuyeron: en las producciones de potencia más altas, las tasas de oxidación de grasas aun presentaban un valor medio de 61 µmol kg-1 min-1. Esto indica que después de la dieta rica en grasas, los sujetos pudieron oxidar mejor los lípidos durante el ejercicio de alta intensidad para compensar (probablemente) sus bajas reservas de glucógeno muscular. Apoyando ésta discusión, la concentración de glicerol en sangre fue mayor durante los ejercicios de ciclismo a 65% y 85% de VO2max después de 4 d de la dieta rica en grasas en comparación con la dieta rica en carbohidratos. Lamentablemente, sin biopsias musculares y/o técnicas de seguimiento/marcaje, es imposible de determinar la(s) fuente(s) de la grasa adicional utilizada durante el ejercicio de alta intensidad después de la adaptación a las grasas.
La comparación de datos de los sujetos en la investigación actual con el modelo integrado presentado por Brooks y Mercier (3) revela una contribución sustancialmente mayor de las grasas con el gasto de energía. Hay varias posibles razones para esta discrepancia. Primero, los sujetos en el estudio presente eran todos atletas altamente entrenados en ultra-resistencia que realizaban entrenamiento prolongado activo en el momento de la investigación (algunos habían competido recientemente en el Campeonato Mundial Ironman en Hawai). Aparte de las investigaciones de Romijn et al. (21,22), los trabajos previos que habían estudiado el metabolismo de combustibles, en su mayoría habían empleado a individuos moderadamente entrenados. Aunque los valores de VO2max de los sujetos en el estudio de Burke et al. (5) eran similares a los del presente experimento, los valores de VO2max exclusivamente no proporcionan información válida sobre el estado de entrenamiento de un individuo. Segundo, nosotros hemos informado previamente (5) que la adaptación a las grasas junto con entrenamiento de resistencia activo disminuye el contenido de glucógeno muscular en reposo a ~250 mmol kg de peso seco-1. En ese estudio, las sesiones de entrenamiento de laboratorio no fueron supervisadas y los sujetos tuvieron dificultad para completar las sesiones establecidas (5). De manera contraria, en la presente investigación los sujetos completaron las sesiones de entrenamiento establecidas, con (probablemente) niveles de glucógeno muscular pre ejercicio muy bajos: las tasas de oxidación de sustratos en el modelo presentado por Brooks y Mercier (3) corresponden a individuos evaluados con sus reservas de glucógeno completas. Tomados en conjunto, nosotros estamos seguros que nuestros datos representan los límites superiores para la oxidación de grasas durante el ejercicio activo.
Dadas las tasas elevadas de oxidación de grasas después de una dieta con elevado contenido de grasas como las observadas en el estudio presente, es interesante especular sobre la intensidad máxima de ejercicio que es posible sostener cuando las grasas son el combustible principal. Aunque se ha sugerido que la oxidación exclusiva de grasas no puede sostener un ejercicio con una intensidad muy superior a 50% de VO2max (8), esta hipótesis sigue sin demostración. Según nuestros conocimientos, el único estudio que investigó sistemáticamente el efecto de la intensidad del ejercicio en el metabolismo de los sustratos en sujetos varones altamente entrenados fue el estudio de Romijn et al. (22). Usando técnicas de isótopos estables junto con calorimetría indirecta, los autores observaron que pedalear a 65% de VO2max producía las mayores tasas de oxidación de grasas en el cuerpo entero (43 µmol kg-1 min-1). Notablemente, en ese estudio (22), el ejercicio realizado a 25% y 85% de VO2max produjo tasas de oxidación de grasas totales similares (27-30 µmol kg-1 min-1).
En la presente investigación, un sujeto no finalizó la sesión de entrenamiento intervalado en el laboratorio, en el d 4 de la dieta rica en grasas. El valor del RER para éste individuo al final de la cuarta repetición a 86% de VO2max fue 0,79 (~30% de energía de los carbohidratos y 70% de las grasas). Para que este sujeto finalizara las cuatro repeticiones finales de 5 min, la producción de potencia fue disminuida a una intensidad equivalente a ~70% de VO2max. Astrand et al. (1) informaron previamente que cuando los sujetos desentrenados comienzan el ejercicio con bajas reservas de glucógeno muscular iniciales, solo pueden manejar una carga de trabajo que alcance el ~85% del VO2max logrado en la condición control. No está completamente claro por qué la capacidad de realizar ejercicios intensos disminuye cuando la grasa es la fuente de combustible predominante. La disponibilidad de carbohidratos como el glucógeno en el músculo activo puede ser importante (3), al igual que el hecho que los carbohidratos son necesarios para mantener los intermediarios del ciclo de los ácidos tricarboxílicos en un nivel necesario para sostener la capacidad oxidativa del músculo (23). Sin embargo, es probable que la razón principal por la cual los carbohidratos son el combustible preferido durante el ejercicio de alta intensidad es que la energía obtenida por unidad de oxígeno consumido es mayor que la de las grasas (8).
Aunque los sujetos en el estudio presente pudieron completar el entrenamiento establecido mientras consumían una dieta con elevado contenido de grasas, esto se asoció con un aumento en el esfuerzo percibido. Tanto el ciclismo de ruta como todas las otras sesiones de entrenamiento que no se realizaron en el laboratorio se caracterizaron por índices significativamente más altos de percepción de esfuerzo de entrenamiento, cuando los sujetos consumieron una dieta con elevado contenido de grasas en comparación a cuando consumieron una dieta con elevado contenido de carbohidratos. Los índices de percepción de esfuerzo de las piernas fueron significativamente mayores durante el entrenamiento intervalado luego de 4 días de la dieta de rica en grasas en comparación con todos los otros días.
En la presente investigación, para evaluar el impacto de los cambios dietéticos en el entrenamiento y en la vida diaria, se realizó un cuestionario de POMS a los sujetos al final de cada día. El puntaje global de POMS fue significativamente más alto en el d-4 de la dieta rica en grasas en comparación con todos los otros días para ambas intervenciones dietéticas. Solamente otro estudio realizó un cuestionario POMS a "atletas" mientras consumían una dieta rica en grasas e intentaban mantener su plan de entrenamiento normal. En esa investigación, Keith et al. (15) observaron una elevación en los puntajes del estado de ánimo total pero no observaron diferencias en el componente de fatiga del cuestionario cuando sus participantes de sexo femenino consumían durante 7 días una dieta con bajo contenido de carbohidratos en comparación a cuando consumían una dieta rica en carbohidratos. Parte de la causa de la diferencia en los resultados entre el estudio actual y el de Keith et al. (15) podría ser que los sujetos en nuestro estudio eran todos atletas altamente entrenados que estaban trabajando con objetivos de competición y mantenían su intensidad de entrenamiento normal. Por otro lado, los "atletas" (VO2max 55 mL kg-1 min-1) en el estudio de Keith et al. (15) podrían haber reducido la intensidad de su bajo volumen de entrenamiento (~110 km sem-1) y haber realizado ejercicios con la misma percepción de esfuerzo tanto en la dieta rica en carbohidratos como en la dieta con bajo contenido de los mismos.
Una conclusión práctica de los resultados del estudio presente es que los atletas de resistencia de competición altamente motivados y entrenados, pueden realizar tanto sesiones de entrenamiento intervalado de alta intensidad como sesiones de resistencia prolongadas de intensidad submáxima consumiendo una dieta con elevado contenido de grasas y bajo contenido de carbohidratos durante 4 días. Sin embargo, en comparación con una dieta rica en carbohidratos isoenergetica, las sesiones de entrenamiento generales y los entrenamientos de laboratorio de alta intensidad se asociaron con mayores índices de esfuerzo percibido. Por consiguiente, cuando se realiza un entrenamiento y prácticas nutricionales especializadas (es decir, una puesta a punto), nosotros recomendamos que los atletas trabajen con medidas objetivas de intensidad de entrenamiento (es decir, producción de potencia) en lugar de con factores subjetivos (es decir, índices de esfuerzo percibido). Por otra parte, los atletas deben ser conscientes que si comienzan un corto período de periodización nutricional, ellos experimentarán un aumento general en la fatiga y un sentimiento reducido de bienestar. Quizás la conclusión más interesante del estudio presente es que después de la adaptación a corto plazo a una dieta rica en grasa y con bajo contenido de carbohidratos, los atletas de competición altamente entrenados pudieron completar el ejercicio aeróbico de alta intensidad. Pedalear a intensidades altas absolutas (~325 W) y relativas (es decir, 86% de VO2max) después de la adaptación a las grasas produjo tasas de oxidación de grasas que se encuentran entre las más altas que hayan sido informadas en la literatura (es decir, >60 µmol kg-1 min-1) para esa producción de potencia. Sin embargo, la oxidación de grasas sola no puede sostener el ejercicio a producciones de potencia que requieren >60-65% de VO2max, incluso en atletas altamente entrenados adaptados a una dieta con elevado contenido de grasas.
Este estudio fue subsidiado mediante un subsidio otorgado por Nestle Australia al Departamento de Nutrición de Deportiva del Instituto Australiano de Deporte. Agradecemos al Dr. Iñigo Mujika, Fabien Gargano, Robyn Hunter y Bronwyn Jones por el excelente soporte técnico.
Dirección de Contacto
John A. Hawley, Ph.D., Department of Human Biology and Movement Science, RMIT University, P.O. Box 71, Bundoora, 3083, Victoria, Australia; E-mail: john.hawley@rmit.edu.au.
Referencias
1. ASTRAND, P. O., I. HALLBACK, R. HEDMAN, and B. SALTIN (1963). Blood lactates after prolonged, severe exercise. J. Appl. Physiol. 18:619-622
2. BORG, G (1975). Simple rating method for estimation of perceived exertion. In: Physical Work and Effort, G. Borg (Ed.). New York: Pergamon, pp. 39-46
3. BROOKS, G. A., and J. MERCIER (1994). Balance of carbohydrate and lipid utilization during exercise: the “crossover” concept. J. Appl. Physiol. 76:2253-2261
4. BRUCE, C. R., M. E. ANDERSON, S. F. FRASER, et al (2000). Enhancement of 2000-m rowing performance after caffeine ingestion. Med. Sci. Sports. Exerc. 32:1958-1963
5. BURKE, L. M., D. J. ANGUS, G. R. COX, et al (2000). Effect of fat adaptation and carbohydrate restoration on metabolism and performance during prolonged cycling. J. Appl. Physiol. 89:2413-2421
6. CHRISTENSEN, E. H., and O. HANSEN (1939). Arbeitsfahigkeit und Ernarung (Physical performance and nutrition). Scand. Arch. Physiol. 81: 160-171
7. GALBO, H., J. J. HOLST, and N. J. CHRISTENSEN (1979). The effect of different diets and of insulin on the hormonal response to prolonged exercise. Acta. Physiol. Scand. 107:19-32
8. GOLLNICK, P. D (1985). Metabolism of substrates: energy substrates during exercise as modified by training. Fed. Proc. 44:353-357
9. HAWLEY, J. A., F. BROUNS, and A. E. JEUKENDRUP (1998). Strategies to enhance fat utilization during exercise. Sports Med. 25:241-257
10. HAWLEY, J. A., and W. G. HOPKINS (1995). Aerobic glycolytic and aerobic lipolytic power systems. A new paradigm with implications for endurance and ultra endurance events. Sports Med. 19:240-250
11. HAWLEY, J. A., and T. D. NOAKES (1992). Peak sustained power output predicts VO2max and performance time in trained cyclists. Eur. J. Appl. Physiol. 65:79-83
12. HELGE, J. W (2000). Adaptations to a fat-rich diet: effects on endurance performance in humans. Sports Med. 30:347-357
13. HELGE, J. W., E. A. RICHTER, and B. KIENS (1996). Interaction of training and diet on metabolism and endurance during exercise in man. J. Physiol. 492:293-306
14. HELGE, J. W., B. WULFF, and B. KIENS (1998). Impact of a fat-rich diet on endurance in man: role of dietary period. Med. Sci. Sports Exerc. 30:456-461
15. KEITH, R. E., K. A. O’KEEFFE, D. L. BLESSING, and G. D. WILSON (1991). Alterations in dietary carbohydrate, protein, and fat intake and mood state in trained female cyclists. Med. Sci. Sports Exerc. 23:212-216
16. LAMBERT, E. V., D. P. SPEECHLY, S. C. DENNIS, and T. D. NOAKES (1994). Enhanced endurance in trained cyclists during moderate intensity exercise following 2 weeks adaptation to a high fat diet. Eur. J. Appl. Physiol. 69:287-293
17. MCNAIR, D. N., and M. LORR (1971). Droppleman: Profile of Moods States Manual. San Diego, CA: Educational and Industrial Testing Service
18. PERONNET, F., and D. MASSICOTTE (1991). Table of nonprotein respiratory quotient: an update. Can. J. Sport Sci. 16:23-29
19. PHINNEY, S. D., B. R. BISTRIAN, W. J. EVANS, E. GERVINO, and G. L. BLACKBURN (1983). The human metabolic response to chronic ketosis without caloric restrictions: preservation of submaximal exercise capacity with reduced carbohydrate oxidation. Metabolism 32: 769-776
20. PINTER, J. K, J. A. HAYASHI, and J. A. WATSON (1967). Enzymatic assay of glycerol, dihydroxyacetone and glyceraldehyde. Arch. Bio-chem. Biophys. 121:404
21. ROMIJN, J. A., E. F. COYLE, J. HIBBERT, and R. R. WOLFE (1992). Comparison of indirect calorimetry and a new breath 13C/12C ratio, method during strenuous exercise. Am. J. Physiol. Endocrinol. Metab. 263:E64-E71
22. ROMIJN, J. A., E. F. COYLE, S. SIDOSSIS, et al (1993). Regulation of endogenous fat and carbohydrate metabolism in relation to exercise intensity and duration. Am. J. Physiol. Endocrinol. Metab. 265:E380-E391
23. SAHLIN, K, A. KATZ, and S. BROBERG (1990). Tricarboxylic acid cycle intermediates in human muscle during prolonged exercise. Am. J. Physiol. Cell Physiol. 259:C834-C841
24. SHEPLEY, B., J. D. MACDOUGALL, N. CIPRIANO, J. R. SUTTON, M. A. TARNAPOLSKY, and G. COATES (1992). Physiological effects of tapering in highly trained athletes. J. Appl. Physiol. 72:706-711
25. STEPTO, N. K, D. T. MARTIN, K. E. FALLON, and J. A. HAWLEY (2001). High metabolic demands of intense aerobic interval training in competitive cyclists. Med. Sci. Sports Exerc. 33:303-310