
Efectos de 3 días de Reposo en Cama sobre las Respuestas Fisiológicas al Ejercicio Progresivo en Atletas y en Hombres Sedentarios
W. C Byrnes2, J. Smorawinski1, K. Nazar2, H. Kaciuba-Uscilko2, E. Kaminska1, A. Kodrzycka2, B. Bicz2 y J. E Greenleaf3
1Department of Sport Medicine, Academy of Physical Education, 61-871 Poznan.
2Department of Applied Physiology, Medical Research Centre, Polish Academy of Sciences, 02-106 Warsaw, Polonia.
3Laboratory for Human Environmental Physiology, National Aeronautics and Space Administration Ames Research Center, Moffett Field, California 94035-1000.
Artículo publicado en el journal PubliCE, Volumen 0 del año 2001.
Publicado 22 de diciembre de 2004
Resumen
Palabras clave: tolerancia al ejercicio, umbral de lactato en sangre, catecolaminas, hormonas, actividad de la renin
INTRODUCCION
El reposo prolongado en cama (BR) provoca una reducción en el rendimiento durante el ejercicio como resultado de una desmejora en el transporte de oxígeno (7, 16, 17) y en la termorregulación (15, 22), así como también por disturbios en el metabolismo intermediario (3, 30), y cambios adversos en la estructura y la función musculoesquelética (4). Tanto los ajustes cardiovasculares como los ajustes metabólicos al ejercicio están controlados por el sistema nervioso autónomo y por el sistema endocrino. Además, un aumento en la secreción de la hormona del crecimiento y de la testosterona luego del ejercicio, provocan un efecto prolongado por medio de la estimulación de los procesos anabólicos en los músculos esqueléticos y en otros tejidos. Por ello, para un mejor entendimiento de los mecanismos de desacondicionamiento producidos por el BR, es necesario aclarar el funcionamiento de los sistemas de regulación nerviosa y hormonal.
La importancia del sistema nervioso simpático (SNS) en la determinación de la tolerancia al trabajo y en la capacidad aeróbica durante el desacondicionamiento fue enfatizada por Sullivan y cols. (34), quienes mostraron que la administración de dobutamina (una droga sintética que imita los efectos de las hormonas adrenales) a sujetos que realizaron BR evitó la disminución del consumo de oxígeno pico (VO2 pico) y atenuó el incremento en la concentración de lactato ([LA]) durante el ejercicio. Sin embargo, hay pocos datos sobre la influencia del desacondicionamiento provocado por el BR sobre la respuesta del sistema nervioso simpático al ejercicio. Engelke y Convertino (14) demostraron un marcado incremento en la concentración plasmática de norepinefrina ([NE]) post-ejercicio máximo, luego de 16 días de BR sin que se observaran cambios en la concentración plasmática de epinefrina ([Epi]), mientras que Sullivan y cols. (34) hallaron, luego de 21 días de BR, una mayor [Epi] arterial durante el ejercicio submáximo sin cambios significativos en la [NE] tanto con el ejercicio submáximo como con el ejercicio máximo. Los datos microneurográficos durante la realización de ejercicios isométricos con el antebrazo indicaron que la actividad nerviosa simpática de los músculos del antebrazo (MSNA) luego de 14 días de BR, fue similar a la observada antes del BR, pero la respuesta de la MSNA a la isquemia muscular, inducida por la detención de la circulación luego del ejercicio, estuvo atenuada (24). Estos hallazgos indican que los reflejos metabólicos en los músculos esqueléticos que activan al SNS, pueden ser atenuados por el desacondicionamiento provocado por el BR.
Poco se sabe acerca de los efectos del BR sobre las respuestas endocrinas al ejercicio. McCall y cols. (27) utilizaron series de flexiones plantares isométricas luego del BR y hallaron que el incremento en la concentración de hormona de crecimiento bioanalizable estuvo inhibida, sugiriendo que la falta de actividad y/o la descarga de los músculos esqueléticos provoca una disrupción del eje aferente músculo–pituitario, lo que provoca una modulación de la liberación de la hormona de crecimiento. Estos autores no hallaron ninguna influencia del BR sobre los niveles, tanto de reposo como de ejercicio, de la hormona de crecimiento inmunoanalizable ([hGH]), testosterona, cortisol u hormonas tiroideas. Parece no haber datos acerca de los efectos del BR sobre la respuesta del sistema renina-angiostesina al ejercicio, aunque se ha reportado consistentemente (18) un incremento en la actividad de la renina plasmática (PRA) durante el BR.
Las respuestas fisiológicas al ejercicio dependen del nivel de aptitud física de los sujetos; sin embargo, excepto para el consumo máximo de oxígeno (VO2 máx.), hay pocos datos disponibles acerca del impacto del nivel de aptitud física (VO2 máx. inicial) sobre las respuestas neuroendócrinas al ejercicio luego del BR. Algunos datos indican que la magnitud de disminución en el VO2 máx. durante el BR es mayor en sujetos altamente entrenados que en aquellos con una baja capacidad de trabajo (9, 10, 31, 35). Sin embargo, hasta la fecha no se ha confirmado una correlación positiva entre el VO2 máx. inicial por kilogramo de masa corporal y el porcentaje de disminución en el VO2 máx. (21), al menos en sujetos que se ejercitaran en posición supina, lo cual minimiza la influencia de los factores ortostáticos sobre los ajustes cardiovasculares (10).
El presente estudio fue diseñado para evaluar las siguientes hipótesis: 1) el BR a corto plazo influencia las respuestas neurohormonales al ejercicio y 2) las respuestas metabólicas, cardiorrespiratorias y neurohormonales al ejercicio luego del BR dependen del nivel y del modo de actividad física habitual de los sujetos y de su capacidad de trabajo. Por lo tanto, se compararon las respuestas de los parámetros cardiorrespiratorios, la [LA] sanguíneo, las concentraciones plasmáticas de Epi, NE, hGH, testosterona, cortisol y PRA al ejercicio progresivo entre hombres sedentarios y atletas entrenados en fuerza y resistencia luego de 3 días de desacondicionamiento provocado por BR.
METODOS
Sujetos
Doce estudiantes varones desentrenados saludables, 10 atletas entrenados en resistencia (ciclistas) y 10 físicoculturistas fueron voluntarios para este estudio, luego de dar su consentimiento escrito (Tabla 1). El protocolo del estudio fue aprobado por el Comité de Ética de la Academia de Educación Física en Poznan, Polonia. Los atletas entrenados en resistencia eran ciclistas amateurs quienes habían estado entrenando regularmente durante 5.5±2.7 (DE) años, y su distancia promedio de entrenamiento era de 100km/semana. Los físicoculturistas tenían una experiencia en el entrenamiento de la fuerza de 3.6±1.8 (DE) años, y en el momento del estudio entrenaban al menos 3 hs/semana con un programa que incluía press de banca, prensa de piernas y sentadillas.
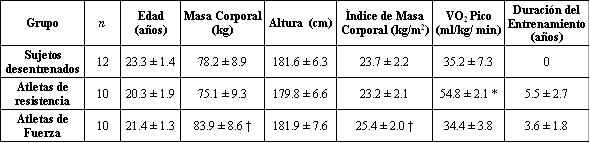
Tabla 1. Características de los sujetos. Los valores son presentados como
medias±DE; n, nro. de sujetos. VO2 pico, consumo de oxigeno pico. *
Diferencia significativa entre los atletas de resistencia y los desentrenados o
los atletas de fuerza, p<0.001. † Diferencia significativa entre los atletas de
fuerza y los atletas de resistencia, p<0.05.
Procedimientos
El BR fue realizado en el hospital para estudiantes en Poznan donde los sujetos se reportaron 2-3 días después de su última sesión de entrenamiento. Se eligió un período de reposo en cama de 3 días debido a que la exposición a unos pocos días de inactividad es suficiente para atenuar el rendimiento en el ejercicio (18). También, dicho período de reposo es el que se prescribe para el tratamiento de una lesión y de enfermedades ligeras. Durante el BR se les permitió a los sujetos deambular por no más de 20 minutos por día (para ducharse e ir al baño); el resto del día los sujetos leyeron libros, escucharon la radio y vieron televisión en posición supina. Un grupo de enfermeras les proporcionó a los sujetos los cuidados necesarios durante las 24hs. A los sujetos se les permitió beber agua ad libitum y su dieta consistió de tres comidas por días preparadas en la cocina del hospital con un total calórico de 12000kJ/día (50% carbohidratos, 35% grasas y 15% proteínas).
Antes y después del BR, entre las 8:30 y las 10:00 am y luego de 2 horas de un desayuno liviano, los sujetos realizaron un test progresivo en cicloergómetro en posición vertical (sentados). La primera carga fue de 50W y se incremento en 50W cada 3 minutos hasta el agotamiento, cada etapa estuvo separada por intervalos de reposo de 1 minuto. Durante el ejercicio se midieron continuamente la ventilación (VE BTPS) y el intercambio de gases respiratorios utilizando un Sistema Cardiopulmonar para el Ejercicio (MedGraphics, St. Paul, MN), a la vez se registró la frecuencia cardiaca (HR) mediante la utilización de un dispositivo Sport Tester (PE 3000, Polar Electro, Kempele, Finland). El volumen latido (SV) y el gasto cardiaco fueron medidos durante las cargas submáximas hasta los 100-150W por medio de un cardiógrafo de impedancia (26), utilizando un dispositivo de monitoreo diseñado en nuestro laboratorio (11). La validez de las mediciones del SV en reposo fue determinada por medio de la utilización de un ecocardiógrafo (r=0.72; n=21, p<0.001) (12), y durante el ejercicio por medio de la utilización del método de respiración de CO2 (r=0.72; n=10; p<0.01).
Inmediatamente después de cada carga, se extrajeron muestras de sangre de 3ml por medio de un catéter fijo insertado 30 minutos antes del comienzo del ejercicio en la vena antecubital, para la determinación de la concentración sanguínea de LA y concentraciones plasmáticas de Epi y NE. Antes y después del ejercicio se extrajeron 5ml adicionales de sangre para la determinación de PRA, [hGH] y de las concentraciones de testosterona y cortisol.
Métodos de Análisis
La [LA] sanguíneo se midió enzimáticamente utilizando instrumental comercial (Boehringer, Mannheim, Alemania), mientras que la [Epi] y la [NE] en plasma fueron medidas por medio del método radioenzimatico de DaPrada y Zurcher (13) utilizando el test de Catecolaminas producido por Immunotech (Praga, República Checa). La [hGH] plasmática se determinó por medio de radioinmunoensayo utilizando instrumental HGH-IRMA MI-131 (Polatom, Swierk, Polonia), las concentraciones plasmáticas de cortisol y testosterona se determinaron con el instrumental para radioinmunoensayo de Orion Diagnostica (Espoo, Finlandia), y la PRA se determinó por medio de radioinmunoensayo utilizando instrumental Immunotech Angiostesin I (Prague, República Checa). Los errores de análisis intra-ensayo (coeficientes de variación) para la Epi, NE, hGH, PRA, cortisol y testosterona fueron 10.8, 8.7, 4.2, 4.4, 7.2 y 3.5% respectivamente.
Cálculos
Las cargas de ejercicio asociadas con un rápido incremento en el lactato sanguíneo, y en las concentraciones plasmáticas de Epi y NE fueron definidas como los umbrales para estas variables y calculadas utilizando regresión lineal por segmentos (log carga de ejercicio vs. log [La], [Epi] o [NE]), de acuerdo con Beaver y cols. (2).
Análisis Estadísticos
La evaluación estadística de las diferencias entre los datos pre y post BR fueron realizados utilizando el análisis de varianza de dos vías para mediciones repetidas. Los dos factores fueron la condición evaluada y las medidas repetidas de los parámetros cardiorrespiratorios, el lactato sanguíneo, y las concentraciones hormonales durante el ejercicio. Cuando se obtuvo un valor F significativo, se utilizo el t-test de Student para ubicar las diferencias entre las medias. El mismo test fue utilizado para la evaluación de las diferencias entre los valores pre y post BR para la carga máxima de ejercicio, VO2 pico, el umbral de lactato sanguíneo y los umbrales de catecolaminas plasmáticas. Se realizó una comparación entre los grupos utilizando el test no paramétrico de Whitney-Mann. Se utilizó el análisis de regresión lineal para evaluar la relación entre el VO2 pico inicial y su declinación durante el BR. Se aceptó un nivel de significancia de p<0.05. Todos los resultados son presentados como medias±DE, excepto en donde se menciona.
RESULTADOS
El BR redujo la carga máxima de ejercicio y el VO2 pico en todos los grupos (Tabla 2) sin provocar un cambio en la masa corporal de los sujetos (datos no presentados). La reducción en el VO2 pico fue más pronunciada en los sujetos entrenados en resistencia (17%, p<0.01) que en los sujetos entrenados en fuerza o en los sujetos sedentarios (10%; p<0.01). Sin embargo, el VO2 pico post BR en los atletas de resistencia siguió siendo más alto (p<0.001) que el valor inicial de los sujetos sedentarios. Los valores individuales de la reducción en el VO2 pico (l/min) inducida por el BR correlacionó positivamente con el VO2 pico inicial expresado en litros por minuto (Figura 1; r=0.73, p<0.001). También se hallaron correlaciones significativas cuando las reducciones en el VO2 pico fueron expresadas como porcentaje de los valores iniciales (r=0.64, p<0.001), cuando el VO2 pico y su reducción fue expresado por kilogramo de masa corporal (r=0.52, p<0.01), y cuando las reducciones del VO2 pico por kilogramo de masa corporal se expresaron como porcentajes de los valores iniciales (r=0.40, p<0.01).
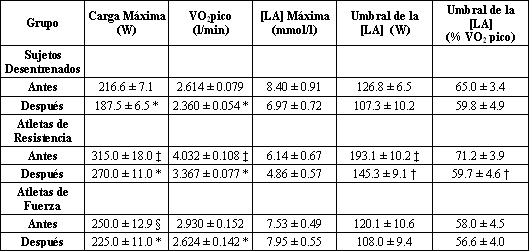
Tabla 2. Caga máxima de ejercicio, VO2 pico, y [LA] sanguíneo alcanzado
durante el test y umbral de la [LA] antes y después del reposo en cama. Los
valores son presentados como medias±EE. [LA], concentración de lactato. *
Significativamente diferente con respecto a los valores anteriores al reposo en
cama, p<0.01. † Significativamente diferente con respecto a los valores
anteriores al reposo en cama, p<0.001. ‡ Diferencia significativa entre los
atletas de resistencia y los sujetos desentrenados o los atletas de fuerza,
p<0.001. § Diferencia significativa entre los atletas de fuerza y los sujetos
desentrenados, p<0.05.
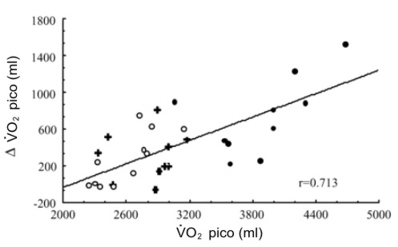
Figura 1. Relación entre el consumo de oxigeno pico inicial (VO2
pico) y su reducción (∆VO2 pico) luego del reposo en cama. ○, sujetos
sedentarios; ●, atletas entrenados en resistencia; +, atletas entrenados en
fuerza.
Los valores de la VE en reposo y durante el ejercicio submáximo fueron similares antes y después del BR en todos los grupos. La VE máxima disminuyó significativamente debido al BR (94.7±6.9 vs. 74.5±3.8l/min; p<0.05) en los atletas de resistencia, pero el cociente entre la VE y el consumo de oxígeno (VO2) se mantuvo sin cambios. Antes del BR el índice de intercambio respiratorio (RER) en reposo fue significativamente mayor (p<0.01) en los atletas entrenados en resistencia que en los sujetos sedentarios o en los atletas de fuerza. Luego del BR los valores del RER en reposo y durante las cargas submáximas fueron mayores que antes del BR en ambos grupos de atletas, pero no en los sujetos sedentarios (Figura 2).
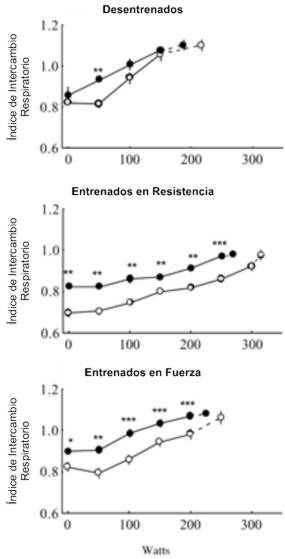
Figura 2. Índices de intercambio respiratorio durante el ejercicio
progresivo antes (○) y después (●) del reposo en cama en los tres grupos de
sujetos. Los valores son presentados como medias±EE. Los últimos puntos
(conectados con líneas punteadas) representan los valores medios con la máxima
carga de ejercicio. Diferencias significativas entre los valores pre y post
reposo en cama: * p<0.05, ** p<0.01, *** p<0.001.
Antes del BR la HR y el SV pre ejercicio no diferían significativamente entre los grupos. Luego del BR la HR de reposo se incrementó significativamente y el SV se redujo en los sujetos sedentarios y en los entrenados en resistencia. Durante el ejercicio submáximo la HR se incrementó y el SV se redujo como consecuencia del BR, solamente en los sujetos desentrenados (Figura 3). En ningún grupo el BR modificó el gasto cardiaco durante el ejercicio.
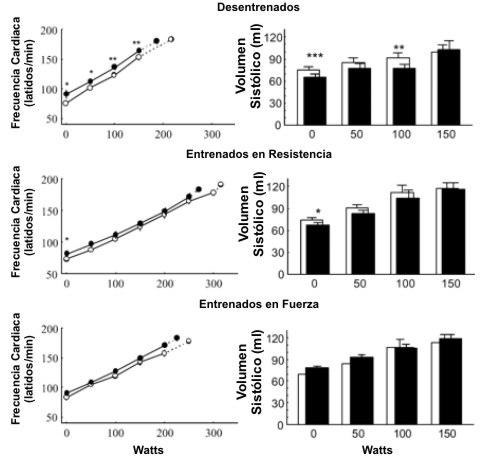
Figura 3. Frecuencia cardiaca y volumen sistólico durante el ejercicio
progresivo antes (○ y barras blancas) y después (● y barras sólidas) del reposo
en cama en los tres grupos de sujetos. Los valores son presentados como medias±
EE. Los últimos puntos (conectados con líneas de puntos) representan los valores
medios obtenidos con la carga máxima de ejercicio. Diferencias significativas
entre los valores pre y post reposo en cama: * p<0.05, ** p<0.01; *** p<0.001.
El BR no cambió la [LA] máxima en ninguno de los grupos (Tabla 2; Figura 4); sin embargo, los valores submáximos para la [LA] fueron mayores después del BR y el umbral de la [LA] en sangre cambió a cargas más bajas en los atletas de resistencia (Tabla 2).
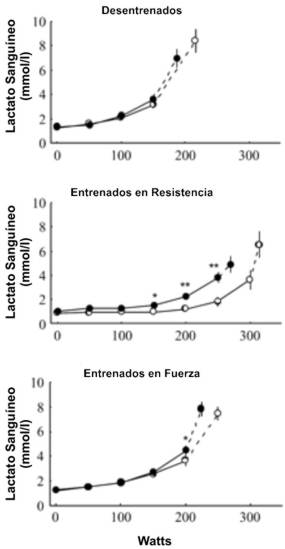
Figura 4. Concentraciones de lactato sanguíneo durante el ejercicio
progresivo antes (○) y después (●) del reposo en cama en los tres grupos de
sujetos. Los valores son presentados como medias±EE. Los últimos puntos
(conectados con líneas de puntos) representan los valores medios obtenidos con
la carga máxima de ejercicio. Diferencias significativas entre los valores pre y
post reposo en cama: * p<0.05, ** p<0.01; *** p<0.001.
De manera similar a la [LA] sanguíneo, la [Epi] y de [NE] en plasma se incrementaron exponencialmente durante el ejercicio (Figuras 5 y 6). Antes del BR el umbral calculado para la NE fue mayor (p<0.01) en ambos grupos de atletas en comparación con los sujetos sedentarios, sin diferencias significativas entre los atletas de resistencia y los atletas de fuerza. Los umbrales para la Epi plasmática fueron similares en todos los grupos. Ni las [Epi] y [NE] pre ejercicio, ni las del ejercicio fueron afectadas significativamente por el BR, pero el umbral de la NE plasmática fue reducido por el BR (p<0.05) en los atletas de resistencia. El umbral de NE post BR en los atletas siguió siendo mayor (p<0.01) que en los sujetos sedentarios. Hubo una correlación significativa (r=0.48, p<0.01) entre los cambios inducidos por el BR en el umbral de la [LA] y el umbral de la [NE] para todos los sujetos. El coeficiente de correlación en los atletas de resistencia fue de 0.78 (p<0.01). El umbral de la Epi plasmática no fue afectado por el BR.
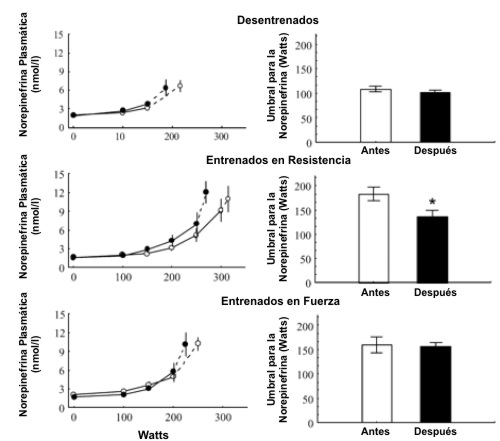
Figura 5. Concentraciones plasmáticas de norepinefrina durante el ejercicio
progresivo y umbral de la norepinefrina antes (○ y barras blancas) y después (●
y barras sólidas) del reposo en cama en los tres grupos de sujetos. Los valores
son presentados como medias±EE. Los últimos puntos (conectados con líneas de
puntos) representan los valores medios obtenidos con la carga máxima de
ejercicio. Diferencias significativas entre los valores pre y post reposo en
cama, * p<0.05.
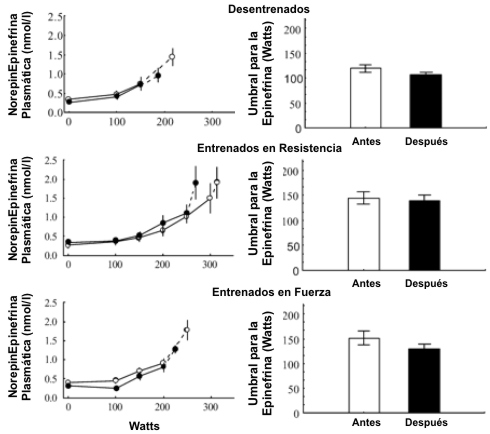
Figura 6. Concentraciones plasmáticas de epinefrina durante el ejercicio
progresivo y umbral de la epinefrina antes (○ y barras blancas) y después (● y
barras sólidas) del reposo en cama en los tres grupos de sujetos. Los valores
son presentados como medias±EE. Los últimos puntos (conectados con líneas de
puntos) representan los valores medios obtenidos con la carga máxima de
ejercicio.
Las concentraciones plasmáticas de las hormonas, tanto en reposo como en ejercicio, antes y después del BR se presentan en la Tabla 3. Antes del BR no hubo diferencias entre los grupos en las concentraciones de reposo o post ejercicio para el cortisol y testosterona, así como también para la PRA, mientras que los niveles plasmáticos post ejercicio de la hGH fueron significativamente mayores (p<0.05) en los atletas que en los sujetos sedentarios. En todos los grupos el ejercicio causo un incremento significativo en la [hGH] plasmática. El BR no afecto la [hGH] en reposo, mientras que los valores post ejercicio fueron significativamente menores en ambos grupos de atletas. Asimismo, los incrementos en la [hGH] inducidos por el ejercicio fueron disminuidos por el BR en los atletas de resistencia (p<0.01), mientras que hubo una tendencia (p<0.07) hacia un menor incremento en los físicoculturistas. De esta manera las diferencias entre los grupos en los valores de la [hGH] post ejercicio no fueron significativos después del BR.
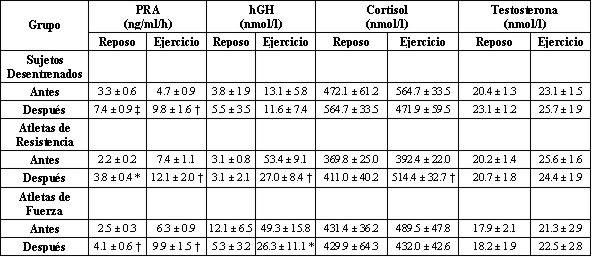
Tabla 3. Actividad de la renina en plasma y concentraciones de la hormona
del crecimiento, cortisol y testosterona en reposo y después del ejercicio,
antes y después de 3 días de reposo en cama. Los valores son presentados como
medias±EE. PRA, actividad de la renina en plasma; hGH, hormona del crecimiento
humana. * Significativamente diferente con respecto a los valores previos al
reposo en cama, p<0.05. † Significativamente diferente con respecto a los
valores previos al reposo en cama, p<0.01. ‡ Significativamente diferente con
respecto a los valores previos al reposo en cama, p<0.001.
Antes del BR el ejercicio no causó un incremento significativo en el cortisol plasmático en ningún grupo, mientras que después del BR hubo un incremento significativo (p<0.05) inducido por el ejercicio en la concentración de cortisol en los atletas de resistencia.
El ejercicio, tanto antes como después del BR, causó una elevación significativa (p<0.01) en la concentración plasmática de testosterona en todos los grupos, pero el BR no causo cambio alguno en los niveles de testosterona, tanto en reposo como post ejercicio. En todos los grupos el BR elevó significativamente (p<0.01) la PRA de reposo; los mayores valores se observaron en el grupo de sujetos sedentarios y difirieron significativamente (p<0.01) de los valores observados en ambos grupos de atletas. En los atletas entrenados en resistencia y en fuerza, los incrementos en la PRA inducidos por el ejercicio fueron mayores después que antes de BR (p<0.01 y p<0.05 respectivamente).
DISCUSION
Los presentes datos muestran claramente que solo 3 días de reposo en cama pueden causar una reducción marcada en la tolerancia al ejercicio manifestada por medio de la reducción en la carga máxima de ejercicio y el correspondiente VO2 pico, que ocurrió durante el ejercicio progresivo. El efecto más pronunciado del desacondicionamiento provocado por el BR fue hallado en los atletas entrenados en resistencia, los cuales tenían la capacidad aeróbica más alta. Los atletas entrenados en fuerza, quienes no diferían significativamente de los sujetos sedentarios en su VO2 pico, tuvieron también una reducción similar en la tolerancia al ejercicio luego del BR. El porcentaje de declinación en el VO2 pico en los últimos dos grupos (~10%) es comparable al reportado luego del desacondicionamiento por BR de corta duración en sujetos con una gran aptitud física, pero que no entrenaban regularmente, mientras que la reducción en el VO2 pico de casi el 17% en los atletas entrenados en resistencia en el presente estudio coincide con niveles obtenidos luego de 2-3 semanas de reposo en cama (7). Los coeficientes de correlación significativos confirmaron la relación entre la capacidad de ejercicio ambulatorio y la reducción en el VO2 pico post BR en un grupo relativamente grande de 32 sujetos.
El desacondicionamiento provocado por el BR involucra efectos de cambios en la posición corporal y comúnmente una reducción en la actividad física. La contribución de los cambios posturales a los efectos del BR probablemente fue similar en los tres grupos de sujetos. Sin embargo, el grado de reducción en la actividad física fue ciertamente mayor en los atletas de resistencia que en los sujetos sedentarios. En los físicoculturistas el entrenamiento está dirigido a mejorar principalmente su masa muscular y su fuerza, pero no su capacidad aeróbica, por lo tanto su actividad física habitual se vio también más reducida por el BR que en los sujetos sedentarios, pero la evaluación realizada en este estudio no fue específica para su régimen de entrenamiento. De esta manera se esperaba una reducción más pronunciada del rendimiento durante el ejercicio aeróbico en los atletas de resistencia que en los sujetos entrenados en fuerza o los sujetos sedentarios, así como también la falta de una gran diferencia entre los dos últimos grupos.
Se ha reportado que las alteraciones en las funciones cardiacas y vasculares inducidas por el desacondicionamiento provocado por el BR prolongado son los factores responsables de la disminución del rendimiento durante el ejercicio (7, 16, 17), sin embargo no son claros los mecanismos que producen la reducción del rendimiento durante el ejercicio luego de 3 días de reposo en cama. En el presente estudio, el gasto cardiaco máximo no fue medido; sin embargo, en los sujetos sedentarios, luego del BR, los valores del SV submáximo se redujeron y la HR se incrementó. Si se asume que el SV durante el ejercicio progresivo alcanza el nivel máximo a cargas submáximas, se podría esperar que el gasto cardiaco máximo se redujera en este grupo. Por otro lado, en ninguno de los grupos de atletas el BR alteró la HR o el SV durante el ejercicio hasta 150W. Esto sugiere que la reducción en el gasto cardiaco máximo no es el factor principal responsable por la declinación en el VO2 pico. Sin embargo, el SV en atletas puede incrementarse progresivamente durante ejercicios progresivos sin llegar a una meseta (20). Por ello, en base a los valores submáximos del SV, no se puede estimar el gasto cardiaco máximo y de esta manera la contribución de la reducción del gasto cardiaco máximo a la limitación del rendimiento luego del BR no puede ser excluida.
La reducción en el rendimiento no puede atribuirse a una reducción en el intercambio de gases pulmonares, ya que la VE durante el ejercicio máximo no fue diferente de los valores pre BR en los sujetos sedentarios y en los atletas de fuerza. En los sujetos entrenados en resistencia, la VE máxima se redujo proporcionalmente a la reducción en el VO2, ya que el cociente VE/VO2 no fue afectado. La falta de influencia del desacondicionamiento provocado por el BR sobre la VE máxima fue reportada previamente (8, 9, 31).
Parece probable que un ajuste inadecuado en la circulación periférica durante el ejercicio o una desmejora en la capacidad aeróbica de los músculos contribuya a la limitación de la capacidad de trabajo luego del BR a corto plazo. Esta hipótesis esta respaldada por el aumento en la [LA] sanguíneo durante el ejercicio submáximo y el más bajo umbral de LA luego del BR en los atletas de resistencia. Esto esta coincide con datos previos (9, 25) que muestran un mayor incremento en la [LA] sanguíneo con cargas submáximas luego de 5-10 días de reposo en cama. Convertino y cols. (9) también reportaron que el umbral anaeróbico, detectado en base a la ventilación pulmonar durante el ejercicio progresivo, apareció con una intensidad más baja de ejercicio luego de 10 días de reposo en cama. Sullivan y cols. (34) también hallaron una reducción en el umbral anaeróbico luego de 3 semanas de BR, menos en los sujetos que fueron tratados con dobutamina.
La mayor producción de LA y el cambio del umbral pueden también ser el resultado de una contribución incrementada de los carbohidratos al proceso de producción de energía, como ocurre luego de una dieta alta en carbohidratos (36). Luego del BR, tanto en reposo como durante el ejercicio submáximo, el RER se elevó en ambos grupos de atletas. Sin embargo, a pesar de una elevación similar en el RER luego del BR en los sujetos entrenados en resistencia y en fuerza, el umbral de [LA] sanguíneo solamente se redujo marcadamente en los primeros. Se ha reportado previamente (3, 8, 30) un incremento del RER luego de reposo en cama prolongado, pero el mecanismo de este efecto no es claro. La dieta durante el BR en el presente estudio, no contuvo excesivas cantidades de carbohidratos en comparación con la dieta habitual de los sujetos, pero la no realización de ejercicio los 2 días previos al estudio y durante los 3 días de BR pudo resultar en una acumulación de glucógeno y/o en una reducción de la actividad de las enzimas musculares involucradas en la oxidación de ácidos grasos.
El BR no afecto la [Epi] y [NE] pre ejercicio en ninguno de los grupos, indicando que la inhibición de la actividad simpática basal previamente reportada después del desacondicionamiento (32, 33) está atenuada en sujetos sentados en un cicloergómetro antes del ejercicio. Las concentraciones de catecolaminas plasmáticas luego del ejercicio máximo fueron similares antes y después de los 3 días de BR, lo que contrasta con los resultados de Engelke y Convertino (14) quienes hallaron que la concentración plasmática de NE luego del agotamiento fue un 64% mayor luego del BR, sin embargo la duración del BR fue mayor (16 días) que en el presente estudio. Coincidiendo con reportes previos (6, 29), nuestros datos demostraron un patrón exponencial en los cambios de las concentraciones de catecolaminas plasmáticas durante el ejercicio progresivo con umbrales similares a los de la [LA]. En los sujetos sedentarios y en los entrenados en fuerza, las concentraciones de catecolaminas con las cargas submáximas y los umbrales de las catecolaminas fueron similares antes y después del BR; sin embargo, en los atletas de resistencia la concentración plasmática de NE con las cargas submáximas tendió a ser mayor luego del BR y el umbral para la NE fue significativamente menor. De esta manera el efecto de desacondicionamiento provocado por 3 días de BR sobre la respuesta del SNS al ejercicio fue evidente solamente en los sujetos entrenados en resistencia.
Parece probable que el cambio en el umbral de la [NE] hacia menores cargas de ejercicio, refleja una activación más temprana del SNS, contribuyendo a un incremento en la producción de lactato, ya que los cambios inducidos por el BR en los umbrales de NE y LA estuvieron significativamente correlacionados. Hay una aparente discrepancia entre esta observación y los hallazgos de Sullivan y cols. (34), quienes reportaron que la administración de dobutamina durante el BR evitó la caída del rendimiento en el ejercicio y la disminución del umbral anaeróbico. Sin embargo, la aplicación de infusiones diarias de dobutamina a lo largo de las 3 semanas de BR probablemente simuló el efecto del entrenamiento aeróbico sobre el sistema cardiovascular y sobre las enzimas oxidativas en el músculo esquelético.
La disminución de la respuesta de la hGH al ejercicio luego del BR es consistente con los datos reportados por McCall y cols. (27); estos investigadores utilizaron ejercicios isométricos realizados con grupos musculares pequeños, lo cual, antes del BR, indujo un incremento en la hGH bioanalizable pero no en la hGH inmunoanalizable. En nuestro estudio los incrementos en la hGH inducidos por el ejercicio fueron mucho mayores en ambos grupos de atletas que en los sujetos sedentarios. Sin embargo, en el grupo de atletas entrenados en fuerza hubo tres sujetos con niveles de hGH muy altos, tanto antes como después del BR. Si estos sujetos hubiesen sido excluidos en los cálculos, el valor medio de incremento inducido por el ejercicio en este grupo podría haber sido menor que en los atletas de resistencia, pero aun sería mayor que en los sujetos sedentarios. La reducción en las respuestas hormonales al ejercicio luego del BR ocurrió en ambos grupos de atletas, pero fue más pronunciada en los sujetos entrenados en resistencia. Estos datos sugieren que la magnitud de la respuesta de la hGH al ejercicio máximo en cicloergómetro depende de la intensidad absoluta de ejercicio, y que la reducción de esta respuesta inducida por el BR puede ser resultado de un agotamiento más temprano. Sin embargo, no puede excluirse la posibilidad de la interrupción del mecanismo reflejo de activación de la liberación de hGH, como lo sugirieran McCall y cols. (27).
Aunque la activación del sistema renina-angiostesina luego del BR está bien documentado (19), no hay datos acerca del efecto del desacondicionamiento por BR sobre la respuesta de la PRA al ejercicio. Los presentes resultados indican que, aparte de la elevación en los valores de PRA de reposo y post ejercicio en todos los sujetos, el BR causó un aumento en la respuesta de la PRA al ejercicio en los atletas, a pesar de una menor carga máxima de ejercicio. La estimulación β-adrenergica del aparato juxtaglomerular del riñón es probablemente el mecanismo principal responsable del incremento en la liberación de renina durante el ejercicio (23). Por ello parece que el incremento en la respuesta de la PRA luego del BR puede ser atribuido a la sensibilización de los receptores β-adrenérgicos, como se sugiriera previamente en experimentos con infusión de agonista β-adrenérgico en reposo (1, 28). La activación temprana del SNS, como lo indicó la disminución del umbral de la [NE] luego del BR en los atletas de resistencia, pudo también haber facilitado la mayor respuesta de la PRA al ejercicio en estos sujetos.
En ninguno de los sujetos el BR modificó la concentración plasmática de testosterona, tanto de reposo como post ejercicio, lo que coincide con los resultados de McCall y cols. (27). Sin embargo, en contraste con los resultados de McCall, en nuestro estudio los niveles de cortisol post ejercicio se elevaron significativamente en los atletas de resistencia a pesar de las menores cargas de trabajo luego del BR.
En resumen, estos datos muestran que la tolerancia al trabajo, la capacidad aeróbica y el umbral anaeróbico estuvieron disminuidos en los sujetos entrenados en resistencia luego de solamente 3 días de BR, mientras que en sujetos sedentarios y entrenados en fuerza, los cambios fueron menos pronunciados. Los tres días de BR incrementaron la PRA de reposo en los tres grupos de sujetos. En los atletas de resistencia el BR modificó las respuestas neuroendócrinas al ejercicio provocando un incremento en la [NE], una atenuación del incremento en la [hGH] y una elevación acentuada de la PRA y el cortisol. El efecto de 3 días de BR sobre las respuestas hormonales al ejercicio no fue significativo en los atletas entrenados en fuerza y en los sujetos sedentarios. Aparentemente la capacidad aeróbica inicial determina la magnitud de estos efectos. Se necesitan más estudios para determinar si estos efectos dependen del nivel de actividad específica del entrenamiento de resistencia que precede al BR o de factores genéticos asociados con la capacidad aeróbica.
Agradecimientos
Este estudio estuvo parcialmente respaldado por la Beca 4 PO5D040 12 del Comité del Estado de Polonia para la Investigación Científica.
Dirección para el pedido de reimpresiones y otra correspondencia
H. Kaciuba-Uscilko, Dept. of Applied Physiology, Medical Research Centre, Pol. Acad. Sci., 5 Pawinskiego str., 02-106 Warsaw, Polonia; (correo electrónico: kaciuba@cmdik.pan.pl).
Referencias
1. Barbe, P, Galitzky J, Thalamas C, Langin D, Lafontan M, Senard JM, and Berlan M (1999). Increase in epinephrine-induced responsiveness during microgravity simulated by head-down bed rest in humans. J Appl Physiol 87: 1614-1620
2. Beaver, WL, Wasserman K, and Whipp BJ (1985). Improved detection of lactate threshold during exercise using a log-log transformation. J Appl Physiol 59: 1936-1940
3. Blanc, S, Normand S, Pahiaudi C, Fortrat JO, Laville M, and Gharib C (2000). Fuel homeostasis during physical inactivity induced by bed rest. J Clin Endocrinol Metab 85: 2223-2233
4. Bloomfield, SA (1997). Changes in musculoskeletal structure and function with prolonged bed rest. Med Sci Sports Exerc 29: 197-206
5. Chase, GA, Grave C, and Rowell LB (1966). Independence of changes in functional and performance capacities attending prolonged bed rest. Aerospace Med 37: 1232-1238
6. Chwalbinska-Moneta, J, Krysztofiak H, Ziemba A, Nazar K, and Kaciuba-Uscilko H (1996). Threshold increase in plasma growth hormone in relation to plasma catecholamines and blood lactate concentrations during progressive exercise in endurance trained athletes. Eur J Appl Physiol 73: 117-120
7. Convertino, VA (1997). Cardiovascular consequences of bed rest: effect on maximal oxygen uptake. Med Sci Sports Exerc 29: 191-196
8. Convertino, VA, Hung J, Goldwater DJ, and DeBusk RF (1982). Cardiorespiratory responses to exercise in middle-aged men following 10 days of bed rest. Circulation 65: 134-140
9. Convertino, VA, Karst GM, Kirby CR, and Goldwater DJ (1986). Effect of simulated weightlessness on exercise-induced anaerobic threshold. Aviat Space Environ Med 57: 325-331
10. Convertino, VA, Stremel RW, Bernauer EM, and Greenleaf JE (1977). Cardiorespiratory responses to exercise after bed-rest in men and women. Acta Astronaut 4: 895-905
11. Cybulski, G, Ksiazkiewicz A, Lukasik W, Niewiadomski W, and Palko T (1996). Central hemodynamics and ECG ambulatory monitoring device with signals recording on PCMCIA memory cards. Med Biol Eng 1: 22-34
12. Cybulski, G, Miskiewicz Z, Szulc J, Torbicki A, and Pasierski T (1993). A comparison between the automatized impedance cardiography and pulsed-wave Doppler echocardiography methods for measurements of stroke volume (SV) and systolic time intervals (STI). J Physiol Pharmacol 44: 251-258
13. DaPrada, M, and Zurcher G (1979). Radioenzymatic assay of plasma and urinary catecholamines in man and various animal species. Physiological and pharmacological adaptations. In: Radioimmunoassay of Drugs and Hormones in Cardiovascular Medicine. edited by Albertini A, DaPrada M, and Pescar A.. Amsterdam
14. Engelke, KA, and Convertino VA (1996). Catecholamine response to maximal exercise following 16 days of simulated microgravity. Aviat Space Environ Med 67: 243-247
15. Ertl, AC, Dearborn AS, Weidhofer AR, Bernauer EM, and Greenleaf JE (2000). Exercise thermoregulation in men after 1 and 24-hours of 6 degrees head-down tilt. Aviat Space Environ Med 71: 150-155
16. Ferretti, G, Antonutto G, Denis C, Hoppeler H, Minetti AE, Narici MV, and Deplanches D (1997). The interplay of central and peripheral factors in limiting maximal O<sub>2</sub> consumption in man after prolonged bed rest. J Physiol (Lond) 501: 677-686
17. Ferretti, G, Giradis M, Moia C, and Antonutto G (1998). Effects of prolonged bed rest on cardiovascular oxygen transport during submaximal exercise in humans. Eur J Appl Physiol 78: 398-402
18. Fortney, SM, Schneider VS, and Greenleaf JE (1996). The physiology of bed rest. In: Handbook of Physiology. Environmental Physiology. Bethesda, MD: Am. Physiol. Soc. sect. 4, vol. II, chapt. 39, p. 889-936
19. Gledhill, N, Cox D, and Jamnik R (1994). Endurance athletes stroke volume does not plateau: major advantage is diastolic function. Med Sci Sports Exerc 26: 1116-1121
20. Greenleaf, JE, and Kozlowski S (1982). Reduction in peak oxygen uptake after prolonged bed rest. Med Sci Sports Exerc 14: 477-480
21. Greenleaf, JE, and Reese RD (1980). Exercise thermoregulation after 14 days of bed rest. J Appl Physiol 48: 72-78
22. Hespel, P, Lijnen P, Vanhees L, Fagard R, and Amery A (1996). Adrenoreceptors and the regulation of blood pressure and plasma renin during exercise. J Appl Physiol 60: 108-113
23. Kamiya, A, Iwase S, Michnikama D, Fua Q, and Mano T (2000). Muscle sympathetic nerve activity during handgrip and post-handgrip muscle ischemia after exposure to simulated microgravity in humans. Neurosci Lett 280: 49-52
24. Katkov, VE, Chestukhin VV, Shepter I, Troshin AZ, Zakharova NS, Sokolov YAA, and Petrov AA (1979). Short term immobilization of healthy men: right ventricular function and metabolism during graded exercise. Cor Vasa 21: 61-70
25. Kubicek, WG, Karnegis JM, Patterson RP, Witsoe DA, and Mattson RH (1966). Development and evaluation of an impedance cardiac output system. Aviat Space Environ Med 37: 1208-1212
26. McCall, GE, Goulet C, Grindeland RE, Hodgson JA, Bigbee AJ, and Edgerton VR (1997). Bed rest suppresses bioassayable growth hormone release in response to muscle activity. J Appl Physiol 83: 2086-2090
27. Melada, GA, Goldman RH, Luetscher JA, and Zager PG (1975). Hemodynamics, renal function, plasma renin and aldosterone in man after 5 to 14 days of bedrest. Aviat Space Environ Med 46: 1049-1055
28. Podolin, DA, Munger PA, and Mazzeo RS (1991). Plasma catecholamine and lactate response during graded exercise with varied glycogen conditions. J Appl Physiol 71: 1427-1433
29. Ritz, P, Acheson KJ, Gachon P, Vico L, Bernard JJ, Alexandre C, and Beaufrere B (1998). Energy and substrate metabolism during a 42-day bed-rest in a head-down tilt position in humans. Eur J Appl Physiol 78: 308-314
30. Saltin, B, Blomqvist G, Mitchell JH, Johnson RL, Wildenthal K, and Chapman CB (1968). Responses to exercise after bed rest and after training. Circulation 38, Suppl7: VIII-1-VII-78
31. Sigaudo, D, Fortrat JO, Allevard AM, Maillet A, Cottet-Emard JM, Vouillarmet A, Hughson RL, Gauquelin-Koch G, and Gharib C (1998). Changes in the sympathetic nervous system induced by 42 days of head-down bed rest. Am J Physiol Heart Circ Physiol 274: H1875-H1884
32. Smorawinski, J, Kaciuba-Uscilko H, Nazar K, Kubala P, Kaminska E, Ziemba AW, Adrian J, and Greenleaf JE (2000). Effects of three-day bed rest on metabolic, hormonal, and circulatory responses to an oral glucose load in endurance or strength trained athletes and untrained subjects. J Physiol Pharmacol 51: 279-289
33. Sullivan, MJ, Binkley PF, Unverferth DV, Ren JH, Boudoulas H, Bashore TM, Merola AJ, and Leier CV (1985). Preventing bed rest-induced physical deconditioning by daily dobutamine infusions. J Clin Invest 76: 1632-1642
34. Taylor, HL, Henschel A, Brozek J, and Keys A (1949). Effects of bed rest on cardiovascular function and work performance. Appl Physiol 2: 223-239
35. Weltman, A (1995). The Blood Lactate Response to Exercise. Champaign, IL: Human Kinetics