
Efectos de la Dieta sobre los Triacilglicéridos Musculares y el Rendimiento en Resistencia
David L Costill1, Raymond D Starling1, Todd A Trappe1, Allen C Parcell1, Chad G Kerr1 y William J Fink1
Human Perfomance Laboratory, Ball State University, Muncie, Indiana, U.S.A.
Artículo publicado en el journal PubliCE, Volumen 0 del año 1997.
Publicado 14 de julio de 2003
Resumen
Palabras clave: glucógeno muscular, rendimiento en ciclismo a intensidad autorregulada, glucosa sérica
INTRODUCCIÓN
Las investigaciones previas han demostrado que hay una relación directa entre la concentración de glucógeno muscular y la capacidad de resistencia (3). Así, es importante para un atleta de resistencia ingerir cantidades adecuadas de carbohidratos antes y después del entrenamiento y/o la competición (7). De manera contraria, menos es conocido acerca de la contribución de los triacilglicéridos musculares al gasto energético durante el ejercicio y acerca de las necesidades alimentarias de grasa de los atletas de resistencia.
Las investigaciones recientes, usando tecnología de isótopos estables, han demostrado que tanto como el 20-25 % del gasto energético durante el ejercicio puede ser derivado de los triacilglicéridos musculares durante el ejercicio prolongado a intensidades moderadas (29). Además, los estudios con biopsia en humanos, han reportado reducciones de 20-50 % en la concentración de triacilglicéridos intramusculares durante el ejercicio prolongado a intensidades entre ≈ 25 y 75 % del máximo consumo de oxígeno (VO2 máx.) (4, 5, 8, 9, 14, 16). De manera contraria, algunos investigadores han demostrado disminuciones menores (18, 20, 32) o no han reportado cambios (32) en la concentración de triacilglicéridos musculares durante el ejercicio prolongado. No obstante, podría ser importante para un atleta de resistencia repletar las reservas de triacilglicéridos musculares después del ejercicio. Para nuestro conocimiento, ninguna investigación ha examinado el efecto de diferentes composiciones dietarias sobre la concentración de triacilglicéridos musculares durante un período agudo después de un ejercicio prolongado.
De este modo, el propósito de esta investigación fue examinar los efectos de una dieta alta en carbohidratos y una dieta alta en grasas sobre el almacenamiento de triacilglicéridos musculares 24 h después de 120 min de ciclismo al 65 % del VO2 máx. Un propósito secundario fue examinar los efectos de estas ingestas diferentes de carbohidratos y grasas sobre la resíntesis de glucógeno muscular y el rendimiento en ciclismo a intensidad autorregulada.
MATERIALES Y MÉTODOS
Sujetos
Siete ciclistas entrenados del sexo masculino participaron en la investigación. Los promedios de edad, talla, peso, y porcentaje de grasa corporal de los sujetos fueron 25 ±3 (DS) años, 181 ±7 cm, 76 ±9 kg, y 10 ±4 %, respectivamente. Su VO2 máx. fue 4.50 ±0.35 L/min. Cada sujeto fue informado de los riesgos, el esfuerzo y los beneficios del estudio antes de firmar un formulario de consentimiento por escrito que había sido aprobado por la Junta de Revisión Institucional.
Evaluaciones Experimentales
Durante esta investigación, cada sujeto completo dos pruebas experimentales que estuvieron separadas por 10-12 días. El protocolo de evaluación experimental fue completado en un período de 2 días. Por los dos días antes de la prueba 1, los sujetos registraron su rutina de dieta y ejercicio de modo que la misma pudiera ser repetida durante la prueba 2 para reducir las diferencias en las concentraciones pre-evaluación de sustratos intramusculares entre las pruebas experimentales. En el día 1 de la evaluación experimental, cada sujeto completo una serie de 120 min de ciclismo al 65 % del VO2 máx. en la mañana (0600-0800) después de una noche de ayuno. Antes e inmediatamente después de la serie de ciclismo de 120 min, fue obtenida una biopsia del músculo vasto lateral, usando una aguja de biopsia percutanea (1) con succión modificada (12).
Después de la biopsia post-ejercicio, en un diseño transversal y al azar los sujetos empezaron un período de 12 h de una dieta alta en carbohidratos (Altos-CHO; 14.97 ± 0.43 MJ, 83% de la energía (E%) de los carbohidratos, 5 E% de las grasas, 12 E% de las proteínas) o de una dieta alta en grasas (Altas-Grasas; 15.02 ± 0.43 MJ, 16 E% de los carbohidratos, 68 E% de las grasas, 16 E% de las proteínas. Toda la comida ingerida fue proporcionada a los sujetos durante el estudio. La energía ingerida por cada sujeto fue la suma del gasto energético calculado durante la serie de 120 min de ciclismo y una estimación de su gasto energético diario en reposo (15), excluyendo el gasto para cualquier otra actividad diaria. Las calorías ingeridas estuvieron equitativamente separadas a través de todo el período de alimentación de 12 horas (i.e., desayuno, almuerzo, cena). Los sujetos ingirieron el desayuno en la hora siguiente después de terminar la serie de ciclismo de 120 min.
Después de un ayuno de 12 h después de una noche, cada sujeto completo luego una prueba de tiempo de intensidad autorregulada sobre un cicloergómetro isoquinético (Cybex MET-100, Ronkonkoma, NY). Este ergómetro está diseñado para fijar la cadencia de pedaleo, pero permite una resistencia variable como respuesta al esfuerzo del sujeto. Inmediatamente antes de la prueba de tiempo, fue obtenida otra biopsia del músculo vasto lateral. La prueba de tiempo empezó después de 5 min de entrada en calor a 150 W. Todos los sujetos completaron 1600 kJ de trabajo durante la prueba de tiempo, la cual simuló ≈ 2 h de ciclismo. Los sujetos fueron instruidos para dar un esfuerzo máximo que fue similar al esfuerzo por tiempo durante las competiciones de ciclismo de ruta que realizaban los sujetos. Además, cada sujeto completo una prueba de tiempo de familiarización 7-10 días antes de la primera prueba experimental, para minimizar cualquier efecto de aprendizaje. El ergómetro tenía una interfase a una computadora que les permitió a los sujetos ver el trabajo total completado, sin ninguna retroalimentación con respecto al tiempo total transcurrido, también fue registrado el tiempo total requerido para completar la prueba de ciclismo de intensidad autorregulada. Además, los gases respiratorios y la frecuencia cardiaca, por medio de radiotelemetría (Polar Vantage XL, Polar Electro, Port Washington, NY) fueron obtenidos cada 200 kJ. Una muestra sanguínea fue obtenida de una vena del antebrazo antes de la prueba y cada 400 kJ a través de toda la duración de la misma.
Técnicas Analíticas
Las muestras obtenidas por medio de la biopsia fueron rápidamente diseccionadas para estar libres de grasa y tejido conectivo visible y luego fueron congeladas y almacenadas en nitrógeno líquido (-190 °C) hasta los análisis posteriores para medir concentraciones de glucógeno y triacilglicéridos. Antes de estos análisis, las muestras musculares fueron deshidratadas por congelación por 20 h y luego fueron pesadas en una balanza electrónica de una sensibilidad de 0.001 mg. El glucógeno muscular total fue determinado en triplicado después de la hidrólisis del músculo en ácido hidroclórico (26). Los residuos de glucosa resultantes de la hidrólisis fueron medidos fluorometricamente con el uso de la reacción de la hexoquinasa-glucosa-6-fosfatasa (27). Los triacilglicéridos musculares totales fueron determinados en cuadriplicado con un tamaño de muestra en peso seco (media ±DS) de 1.3 ±0.5 mg. Fue usada una modificación del método del cloroformo-metanol de Frayn y Maycock (13) para extraer los triacilglicéridos de las muestras musculares. Las extracciones fueron secadas, hidrolizadas en KOH alcohólico, neutralizadas con HCl, y la concentración de triacilglicéridos fue luego determinada por medio de la medición del glicerol liberado espectrofotometricamente usando un método enzimático comercialmente disponible (kit no. 337, Sigma Chemical, St. Louis, MO).
Las muestras sanguíneas fueron centrifugadas, y el suero fue luego almacenado a -20 °C hasta los análisis posteriores. Las concentraciones de glucosa sérica, ácidos grasos libres (kit no. 990-75401, Wako Pure Chemical Industries, Osaka, Japan), y de glicerol fueron medidas en todas las muestras séricas por medio del uso de métodos enzimáticos comercialmente disponibles.
Los gases respiratorios fueron analizados para identificar las fracciones de oxígeno (model S-3A, Applied Electrochemistry, Sunnyvale, CA) y dióxido de carbono (model LB-2, SensorMedics, Anaheim, CA) usando analizadores electrónicos. El volumen de gas fue determinado por medio de un contador de gas seco (Parkinson Cowan). A partir de estos datos, fue calculado en consumo de oxígeno (VO2) y el índice de intercambio respiratorio (RER).
Análisis Estadísticos
Para examinar posibles diferencias entre las pruebas experimentales para las concentraciones musculares de glucógeno y triacilglicéridos y para todas las variables dependientes obtenidas a través de toda la prueba de ciclismo por tiempo fue usado un análisis de varianza a dos vías para mediciones repetidas (tratamiento x tiempo). Cuando eran obtenidos valores F significativos, era administrado un análisis post hoc Tukey para localizar las diferencias entre las medias. Además, las posibles diferencias entre las pruebas experimentales para el tiempo total de rendimiento fueron examinadas usando un test t apareado. La significancia fue aceptada al nivel P <0.05.
RESULTADOS
Glucógeno y Triacilglicéridos Musculares
La concentración total de triacilglicéridos musculares antes de la serie de ciclismo de 120 min. no fue significativamente diferente entre los grupos de dietas Altos-CHO y Altas-Grasas (Tabla 1). Además, las concentraciones después de la prueba de 120 min no fueron significativamente diferentes en comparación con los valores correspondientes a los grupos Altos-CHO y Altas-Grasas. La concentración de triacilglicéridos musculares 24 h después de la prueba fue significativamente más alta para el grupo Altas-Grasas en comparación con el grupo Altos-CHO (Tabla 1). La concentración de glucógeno muscular total antes de la serie de ciclismo de 120 min. no fue significativamente diferente entre los grupos Altos-CHO y Altas-Grasas (Tabla 1). La concentración de glucógeno muscular después de la prueba de 120 min. fue significativamente más baja en comparación con los valores pre-ejercicio para ambos grupos Altos-CHO y Altas-Grasas. Además, la concentración de glucógeno muscular fue significativamente más alta para el grupo Altos-CHO en comparación con el grupo Altas-Grasas 24 h después de la prueba de 120 min. (Tabla 1).
Prueba de Ciclismo de Intensidad Autorregulada
El tiempo total para completar la prueba de tiempo de 1600 kJ fue significativamente mayor durante la dieta Altas-Grasas (139.3 ±7.1 min) en comparación con la dieta Altos-CHO (117.1 ±3.2 min). La Figura 1 exhibe el tiempo transcurrido cada 200 kJ a través de toda la prueba de tiempo para las dietas Altas-Grasas y Altos-CHO. Además, el VO2 expresado como un porcentaje del VO2 máx. no fue significativamente diferente entre las dietas Altas-Grasas y Altos-CHO durante los primeros 1000 kJ de la prueba de tiempo (Figura 2). De manera contraria, el porcentaje del VO2 máx. mantenido fue significativamente más bajo durante la dieta Altas-Grasas en comparación con la dieta Altos-CHO en 1200 (59 ±3 vs. 75 ±3%), 1400 (59 ±6 vs. 76 ±4%), y 1600 (53 ±8 vs. 84 ±5%) kJ, respectivamente. Como en los datos del VO2, los valores de frecuencia cardiaca durante los primeros 1000 kJ de la prueba de tiempo no fueron significativamente diferentes entre las dietas Altas-Grasas y Altos-CHO (Figura 2). La frecuencia cardiaca fue significativamente más baja durante la dieta Altas-Grasas en comparación con la dieta Altos-CHO a 1200 (143 ±3 vs. 156 ±5 latidos/min), 1400 (139 ±6 vs. 162 ±4 latidos/min), y 1600 (133 ±9 vs. 167 ±4 latidos/min) kJ, respectivamente. El RER fue significativamente más bajo en todos los puntos de análisis durante la prueba de tiempo para las dietas Altas-Grasas vs. Altos-CHO (Figura 2). El RER promedio durante la prueba de tiempo para las dietas Altos-CHO y Altas-Grasas fue de 0.89 ± 0.01 y 0.82 ± 0.01, respectivamente.

Tabla 1. Sustratos intramusculares antes, inmediatamente después, y 24 h después
de una serie de ciclismo de 120 min.
Los valores están expresados como medias ±DS en mmol/kg de peso seco. Altos-CHO,
altos carbohidratos; Altas-Grasas, altas grasas. * Significativamente diferente
con respecto al valor pre-prueba, P <0.05. † Significativamente diferente con
respecto al valor post-prueba, P <0.05. ‡ Significativamente diferente con
respecto al valor 24 h después del grupo Altos-CHO, P <0.05.
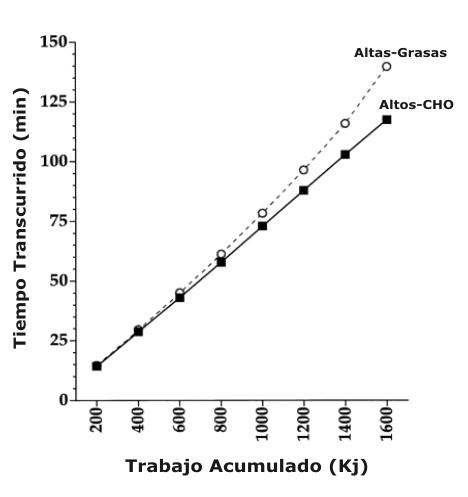
Figura 1. Tiempo de rendimiento transcurrido a través de toda la prueba de
ciclismo por tiempo de 1600 kJ para las dietas altas en carbohidratos (Altos-CHO)
y altas en grasas (Altas-Grasas).
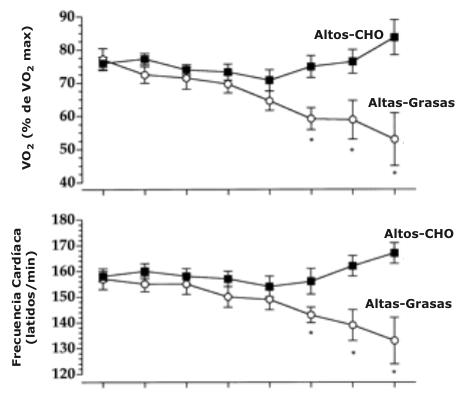

Figura 2. Datos de los porcentajes de la captación máxima de oxígeno (VO2
máx.; arriba), frecuencia cardiaca (mitad), e índice de intercambio respiratorio
(RER; abajo) durante la prueba de ciclismo por tiempo de 1600 kJ para las dietas
Altos-CHO y Altas-Grasas. VO2 máx., captación de oxígeno. *Significativamente
diferente con respecto al valor Altos-CHO, P < 0.05. † Efecto de tratamiento
significativo, P < 0.05.
La concentración de glucosa antes de la prueba de ciclismo por tiempo no fue significativamente diferente entre las dietas de Altos-CHO (5.0 ± 0.05 mmol/l) y Altas-Grasas (5.0 ± 0.08 mmol/l) (Figura 3). De manera contraria, la concentración de glucosa fue significativamente más baja en el grupo Altas-Grasas en comparación el grupo Altos-CHO a 400 (4.4 ± 0.2 vs. 4.8 ± 0.2 mmol/l), 800 (4.0 ± 0.2 vs. 4.5 ± 0.2 mmol/l), 1200 (3.6 ± 0.2 vs. 4.2 ± 0.2 mmol/l), y 1600 (3.1 ± 0.3 vs. 4.1 ± 0.2 mmol/l) kJ, respectivamente. La concentración de ácidos grasos libres antes la prueba de ciclismo por tiempo no fue significativamente diferente entre las dietas Altos-CHO (0.38 ±0.04 mmol/l) y Altas-Grasas (0.54 ±0.06 mmol/l) (Figura 3). Sin embargo, la concentración de ácidos grasos libres fue significativamente más alta en el grupo Altas-Grasas en comparación con el grupo Altos-CHO a 1200 (1.37 ± 0.14 vs. 1.03 ± 0.12 mmol/l) 1600 (1.76 ± 0.24 vs. 1.35 ± 0.21 mmol/l) kJ, respectivamente. La concentración de glicerol antes de la prueba de ciclismo por tiempo no fue significativamente diferente entre las dietas Altos-CHO (0.053 ± 0.015 mmol/l) y Altas-Grasas (0.062 ± 0.012 mmol/l) (Figura 3). De manera contraria, la concentración de glicerol fue significativamente más alta para el grupo Altas-Grasas en comparación con el grupo Altos-CHO a 400 (0.31 ± 0.03 vs. 0.14 ± 0.02 mmol/l), 800 (0.45 ± 0.04 vs. 0.22 ±0.04 mmol/l), 1200 (0.57 ± 0.07 vs. 0.33 ± 0.04 mmol/l), y 1600 (0.60 ± 0.06 vs. 0.48 ± 0.05 mmol/l) kJ, respectivamente.
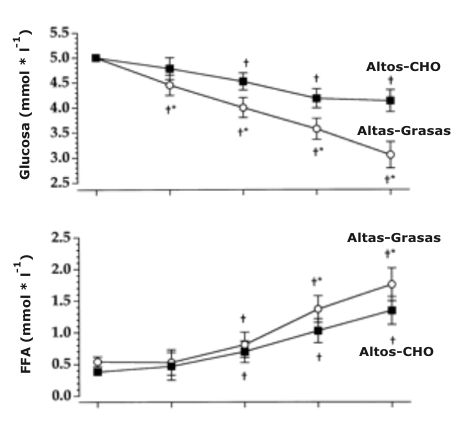
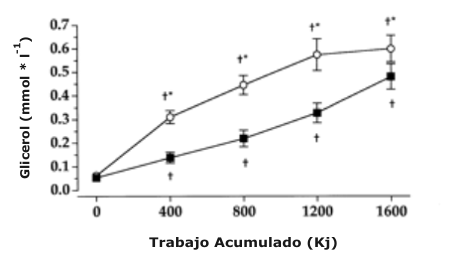
Figura 3. Datos de la glucosa sérica (arriba), ácidos grasos libres (FFA;
mitad) y glicerol (abajo) durante la prueba de ciclismo por tiempo de 1600 kJ
para las dieta Altos-CHO y Altas-Grasas.* Significativamente diferente con
respecto al valor Altos-CHO, P < 0.05. † Significativamente diferente con
respecto al valor pre-prueba, P < 0.05.
DISCUSIÓN
Los resultados de la presente investigación demuestran que no hubo una diferencia significativa en la concentración de triacilglicéridos musculares antes y después de una serie de ciclismo prolongada a intensidad submáxima. No obstante, la ingestión de una dieta alta en grasas durante el período de alimentación de 24 h después de la serie de ciclismo, incremento la concentración de triacilglicéridos musculares en un 36%. Además, la ingestión de 9.8 vs. 1.9 g de carbohidratos/kg de peso corporal durante el período de alimentación de 24 h después de la serie de ciclismo resultó en un mayor almacenamiento de glucógeno y una subsiguiente mejoría en el rendimiento en una prueba de ciclismo a intensidad autorregulada.
Durante la serie de ciclismo de 120 min al 65% del VO2 máx. hubo una reducción (P > 0.05) en la concentración de los triacilglicéridos musculares de un 6 a un 11% en las dietas Altos-CHO y Altas-Grasas, respectivamente (ver Tabla 1). Otros investigadores han demostrado porcentajes de cambio similares en la concentración de triacilglicéridos musculares durante el ejercicio prolongado (18, 20, 32). De manera contraria al presente estudio y a los estudios antes mencionados, otros investigadores han demostrado reducciones de un 20-50% en la concentración de triacilglicéridos musculares durante ejercicio prolongado a intensidades de entre un ≈55 y 75% del VO2 máx. (4, 5, 8, 9, 14, 16). Las diferencias entre estos estudios en el modo de ejercicio, intensidad y duración podrían explicar las discrepancias en los hallazgos. Además, las concentraciones intramusculares pre-ejercicio (11), el almacenamiento heterogéneo de grasa (10), y las diferencias en la utilización entre los grupos musculares (28) podría influenciar los cambios en los triacilglicéridos musculares durante el ejercicio prolongado.
Aunque no hubo una diferencia significativa en los triacilglicéridos musculares antes y después de la serie de ciclismo de 120 min, la ingestión de una dieta alta en grasas durante el subsiguiente período de alimentación de 24 h incrementó la concentración de triacilglicéridos musculares en un 36% (ver Tabla 1). De manera contraria, una dieta alta en carbohidratos no cambio significativamente la concentración de triacilglicéridos musculares. Para nuestro conocimiento esta es la primera investigación que examino el efecto de la dieta sobre la concentración de triacilglicéridos musculares 24 h después de un ejercicio prolongado. Otros investigadores han examinado el efecto de la composición de la dieta sobre la concentración de triacilglicéridos musculares en humanos durante períodos de 5 días a 4 semanas de duración (18, 21).
Con el uso de una dieta similar a la usada en la presente investigación, Jansson y Kaijser (18) examinaron el efecto de 5 días de una dieta alta en carbohidratos (75% de la energía) o alta en grasas (69% de la energía) sobre la concentración de triacilglicéridos musculares. Aunque no fueron obtenidas biopsias pre-dieta para examinar el porcentaje de cambios, la concentración de triacilglicéridos musculares del músculo vasto lateral en reposo, después del período de alimentación de 5 días fue 80% más alta con la dieta alta en grasas (90.7 ±20.1 mmol/kg de peso seco) en comparación con la dieta alta en carbohidratos (50.4 ±7.4 mmol/kg de peso seco). Esta diferencia promedio no fue estadísticamente significativa. Después de 4 semanas de una dieta alta en grasas (54% de la energía), se ha demostrado que la concentración de triacilglicéridos del músculo vasto lateral se incrementó un 56% (de 30 ±4 a 47 ±8 mmol/kg de peso seco) en un grupo de 10 hombres sanos (21). Durante un período subsiguiente de 4 semanas de una dieta baja en grasas (29% de la energía), Kiens et al. (21) reportaron que la concentración de triacilglicéridos musculares disminuyó un 13%, a 41 ±7 mmol/kg de peso seco.
La habilidad de incrementar la concentración de triacilglicéridos podría estar vinculada con la actividad de la lipoproteína lipasa. Esta enzima cataliza la hidrólisis de triacilglicéridos en el lecho capilar del tejido adiposo y el músculo esquelético (25). Varios investigadores han demostrado un incremento de la actividad de la lipoproteína lipasa del músculo esquelético después de varios días (17) o semanas (21, 31) de una dieta alta en grasas. La actividad de la lipoproteína lipasa del músculo esquelético podría estar vinculada con los niveles de insulina circulante. Ha sido demostrado que la actividad de la lipoproteína lipasa del músculo esquelético disminuye cuando las concentraciones de insulina se elevan (22). Así, podría esperarse que una dieta que no aumente los niveles de insulina, incremente la actividad de la lipoproteína lipasa del músculo esquelético, la cual facilitaría el almacenamiento de triacilglicéridos intramusculares.
Cuando se compara el presente estudio con otros estudios que utilizaron períodos de dieta más largos, es interesante señalar que solo 1 día de dieta alta en grasas durante la presente investigación resultó en un incremento muy grande en la concentración de triacilglicéridos musculares (18, 21). Además de la dieta, la actividad de la lipoproteína lipasa del músculo esquelético podría haberse incrementado después del ejercicio prolongado (25, 32). Ha sido demostrado que la actividad de la lipoproteína lipasa del músculo esquelético se incrementa de dos a tres veces inmediatamente después de una serie prolongada de ejercicio vigoroso (23, 24, 30). Además, ha sido demostrado que la actividad de la lipoproteína lipasa del músculo esquelético se eleva significativamente 4 h después de 60 min de ejercicio con una pierna (22). Así, durante la presente investigación la combinación de una dieta alta en grasas y la serie previa de ciclismo de 120 min podría haber tenido un efecto sinérgico con respecto al incremento de la concentración de triacilglicéridos musculares .
Los resultados de la presente investigación demostraron que una cantidad adecuada de carbohidratos debe ser ingerida después de un ejercicio exhaustivo para mantener la concentración de glucógeno muscular y prevenir una reducción en el rendimiento de resistencia subsiguiente. La ingestión de 9.8 g de carbohidratos/kg de peso corporal durante la dieta Altos-CHO resultó en la resíntesis del 93% del glucógeno utilizado a través de toda la serie de ciclismo de 120 min (ver Tabla 1). De manera contraria, solo el 13% del glucógeno utilizado a través de toda la serie de ciclismo de 120 min fue resintetizado cuando fueron ingeridos 1.9 g de carbohidratos/kg de peso corporal durante la dieta Altas-Grasas. Estos resultados son similares a los de otros estudios que han demostrado una resíntesis completa de glucógeno muscular durante un período de 24 h después de un ejercicio agotador cuando fueron ingeridos entre 8 y 10 g de carbohidratos/kg de peso corporal (2, 6). De manera similar a la presente investigación, Costill et al. (6) reportó que no hubo resíntesis de glucógeno alguna después de una carrera exhaustiva cuando fueron ingeridos 2.4 g de CHO x kg de peso corporal -1 x 24 h -1.
Con la resíntesis incompleta de glucógeno muscular durante la dieta Altas-Grasas, el rendimiento en ciclismo a intensidad autorregulada fue un 19% más bajo en comparación con el grupo Altos-CHO. Para nuestro conocimiento, nadie ha examinado el efecto de diferentes ingestas de carbohidratos durante un período de 24 h después de un ejercicio prolongado sobre el rendimiento en una prueba de ciclismo a intensidad autorregulada. Estos resultados son interesantes porque investigaciones recientes han demostrado que las pruebas por tiempo a intensidad autorregulada son una medición más confiable del rendimiento de lo que lo son las evaluaciones tradicionales de tiempo-fatiga (19). El almacenamiento inadecuado de glucógeno muscular resulto más probablemente en una reducción del rendimiento durante la dieta Altas-Grasas en comparación con la dieta Altos-CHO. Aunque nosotros no obtuvimos biopsias musculares post-tiempo, el RER y la concentración de glucosa sérica, más bajos y los niveles más altos de ácidos grasos libres y glicerol durante la dieta Altas-Grasas vs. la dieta Altos-CHO proporciono evidencia indirecta de una reducción en el almacenamiento de glucógeno y así un incremento en la contribución de las grasas al gasto energético del ejercicio. En conjunto, estos resultados demuestran que la ingesta de ≈ 10 g de carbohidratos/kg de peso corporal durante un período de 24 h después de un ejercicio agotador van a resintetizar las reservas de glucógeno muscular y a mejorar el subsiguiente rendimiento en ciclismo a intensidad autorregulada.
En conclusión, la concentración de triacilglicéridos musculares no fue significativamente diferente antes y después de 120 min de una serie de ciclismo submáximo. No obstante, la ingestión de una dieta alta en grasas incremento la concentración de triacilglicéridos musculares en un 36%, 24 h después de la serie de ciclismo. Además, una dieta alta en carbohidratos no incremento la concentración de triacilglicéridos musculares, pero incremento las reservas de glucógeno muscular e incrementó el rendimiento de una serie de ciclismo a intensidad autorregulada en comparación con una dieta alta en grasas.
Agradecimientos: Agradecemos a Doug E. Bolster y a Gary Lee por la asistencia técnica.
Nota: Esta investigación fue subvencionada en parte por una beca del Programa Interno de Becas de la Universidad de Ball State.
Dirección para Correspondencia: D. L. Costill, Human Performance Laboratory, Ball State Univ., Muncie, IN 47306.
Referencias
1. Costill, D. L., P. D. Gollnick, E. D. Jansson, B. Saltin, and E. M. Stein (1973). Glycogen depletion pattern in human muscle fibres during distance running. Acta Physiol. Scand. 89: 374-383
2. Costill, D. L., W. M. Sherman, W. J. Fink, C. Maresh, M. Witten, and J. M. Miller (1981). The role of dietary carbohydrates in muscle glycogen resynthesis after strenuous running. Am. J. Clin. Nutr. 34: 1831-1836
3. Coyle, E. F (1991). Timing and method of increased carbohydrate intake to cope with heavy training, competition and recovery. J. Sports Sci. 9: 29-52
4. Evans, W. J., S. K. Phinney, and V. R. Young (1982). Suction applied to a biopsy maximizes sample size. Med. Sci. Sports Exercise 14: 101-102
5. Frayn, K. N., and P. F. Maycock (1980). Skeletal muscle triacylglycerol in the rat: methods for sampling and measurement, and studies of biological variability. J. Lipid Res. 21: 139-144
6. Harris, J. A., and F. G. Benedict (1919). A Biometric Study of Basal Metabolism in Man. Philadelphia, PA: Washington Square
7. Hurley, B. F., P. M. Nemeth, W. H. Martin III, J. M. Hagberg, G. P. Dalsky, and J. O. Holloszy (1986). Muscle triglyceride utilization during exercise: effect of training. J. Appl. Physiol. 60: 562-567
8. Jacobs, I., H. Lithell, and J. Karlsson (1982). Dietary effect of glycogen and lipoprotein lipase activity in skeletal muscle in man. Acta Physiol. Scand. 115: 85-90
9. Jansson, E., and L. Kaijser (1982). Effect of diet on the utilization of blood-borne and intramuscular substrates during exercise in man. Acta Physiol. Scand. 115: 19-30
10. Jeukendrup, A., W. H. M. Saris, F. Brouns, and A. D. M. Kester (1996). A new validated endurance performance test. Med. Sci. Sports Exercise 28: 266-270
11. Kiens, B., H. Lithell, K. J. Mikines, and E. A. Richter (1989). Effects of insulin and exercise on muscle lipoprotein lipase activity in man and its relation to insulin action. J. Clin. Invest. 84: 1124-1129
12. Ladu, M. J., H. Kapsas, and W. K. Palmer (1991). Regulation of lipoprotein lipase in muscle and adipose tissue during exercise. J. Appl. Physiol. 71: 404-409
13. Oscai, L. B., D. A. Essig, and W. K. Palmer (1990). Lipase regulation of muscle triglyceride hydrolysis. J. Appl. Physiol. 69: 1571-1577
14. Passonneau, J. V., and V. R. Lauderdale (1974). A comparison of three methods of glycogen measurement in tissues. Anal. Biochem. 60: 405-412
15. Passonneau, J. V., and O. H. Lowry (1993). A collection of metabolite assays. In: Enzymatic Analysis. A Practical Guide. Totowa, NJ: Humana, p. 111-128
16. Reitman, J., K. M. Baldwin, and J. O. Holloszy (1973). Intramuscular triglyceride utilization by red, white, intermediate skeletal muscle and heart during exhausting exercise. Proc. Soc. Exp. Biol. Med. 142: 628-631
17. Romijn, J. A., E. F. Coyle, L. S. Sidossis, A. Gastaldelli, J. F. Horowitz, E. Endert, and R. R. Wolfe (1993). Regulation of endogenous fat and carbohydrate metabolism in relation to exercise intensity and duration. Am. J. Physiol. 265 (Endocrinol. Metab. 28): E380-E391
18. Thompson, P. D., E. M. Cullinane, R. Eshleman, M. A. Kantor, and P. N. Herbert (1984). The effects of high-carbohydrate and high-fat diets on the serum lipid and lipoprotein concentrations of endurance athletes. Metabolism 33: 1003-1010
19. Turcotte, L. P., E. A. Richter, and B. Kiens (1995). Lipid metabolism during exercise. In: Exercise Metabolism, edited by M. Hargreaves. Champaign, IL: Human Kinetics, p. 99-130