
Ejercicio Intermitente y quema de grasas: es lo que parece?
Publicado 2 de agosto de 2013, 13:02

Continuando con nuestra temáticas acerca del ejercicio intermitente y su utilización en ambientes de fitness y salud, varias décadas posteriores a los trabajos de Essén que ya analizamos en una entrada de blog anterior, una serie de estudios de
Christmass (1999) tuvieron como objeto comparar el uso neto de sustrato a través de
un período sostenido de ejercicio intenso intermitente y ejercicio submáximo
continuo con similar VO2 y gasto energético globales. Curiosamente,
estos trabajos de Christmass no son citados (en cambio, sí citan los de Essén) por
los defensores del ejercicio intermitente como el método de excelencia para la
quema de grasas. ¿Por qué? Veremos…
Para uno de sus estudios, Christmass
utilizó a 7 sujetos activos (24 años de edad; 180 cm de altura; 73 kg de peso
corporal; 56 VO2máx, valores promedio) que realizaron 2 protocolos
de ejercicio en cinta ergométrica (ejercicio de carrera): uno de los protocolos
consistió en ejercicio intermitente intenso durante 90’ (intensidad: 120% VO2pico;
rango de velocidad: 19,5-20,9 km·h-1; 0% de inclinación). El formato
era de 12” (ejercicio intenso) seguido de 18” de pausa (descanso completamente
pasivo), es decir, una relación trabajo:pausa de 1:1,5. El protocolo de
ejercicio continuo submáximo en cinta ergométrica también, fue realizado al 60% del VO2pico
(rango de velocidad: 9.3-10.6 km·h-1) durante 90’. Ambos protocolos
fueron realizados con 7 días de diferencia. Algunos de los resultados de dicho
estudio se pueden observar en la Figura 1.
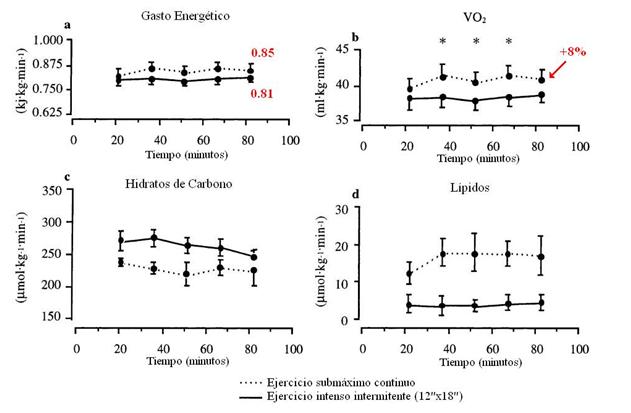
Figura 1. a, gasto energético; b, consumo de oxígeno (VO2); c, tasa de oxidación de CHO; d, tasa de oxidación de lípidos durante el ejercicio submáximo continuo (líneas punteadas) y durante el ejercicio intenso intermitente (líneas sólidas). * P < 0.05 diferencia significativa entre los protocolos de ejercicio.
Allí podemos observar que el gasto energético no fue diferente entre ambos protocolos y se mantuvo constante desde los 22,5 minutos hasta el final del experimento. El VO2 fue igualado en los primeros momentos y al final del ejercicio (7,5-37,5 minutos y 82,5 minutos), pero ligeramente más alto (+8%) fue durante el ejercicio continuo desde los 37,5 hasta los 67,5 minutos. La intensidad global del ejercicio, representada como una proporción del VO2pico fue de 68.1±2.5% VO2pico para el ejercicio intermitente (casi un +10% que el estudio de Essén), y 71.8±1.8% VO2pico para el ejercicio continuo. La respuesta de la FC fue de 89% de la FCmáx para el ejercicio intermitente y del 84% de la FCmáx para el ejercicio continuo. Aunque el gasto energético fue casi el mismo durante ambos protocolos, el R fue más alto durante el ejercicio intermitente (0.99) comparado al ejercicio continuo (0.93). Recordemos que este investigador utilizó una calorimetría indirecta para dicho análisis, un método que fue utilizado por Essén también y que tiene cierto cuestionamiento de su uso en esfuerzos de alta intensidad no estables. La tasa de oxidación de lípidos fue casi 3 veces menor durante el ejercicio intermitente (6.9±0.9 µmol·kg-1·min-1) comparado al ejercicio continuo (17.9±1.6 µmol·kg-1·min-1). El ritmo de oxidación de los hidratos de carbono fue más alta durante el ejercicio intermitente (264±5 µmol·kg-1·min-1) comparado al ejercicio continuo (229±6 µmol·kg-1·min-1). Ambas tasas de oxidación de lípidos y de CHO se mantuvo constante entre los 22,5 y los 90 minutos.
Respecto a las
concentraciones de los metabolitos plasmáticos, el lactato fue similar en
reposo y se mantuvo constante desde los 15’ hasta el final del ejercicio en
ambos protocolos (Figura 2). La concentración de lactato fue más alta durante
el ejercicio intermitente. En ambos protocolos, el piruvato se incrementó desde
el reposo hasta los 15’. En el ejercicio intermitente, el piruvato fue
constante entre los 15’ y 75’, y luego al final se elevó. En cambio, en el
ejercicio continuo, se mantuvo constante desde los 15’ hasta el final. El
piruvato entonces fue más alto en el ejercicio intermitente comparado al
ejercicio continuo.
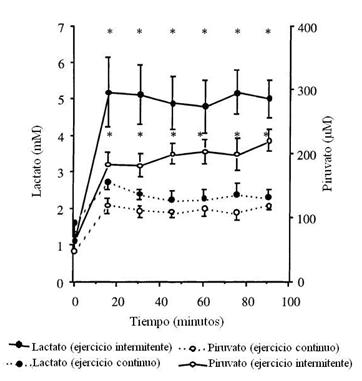
Figura 2. Concentración plasmática de lactato y piruvato durante el ejercicio submáximo continuo (líneas punteadas) y durante el ejercicio intenso intermitente (líneas sólidas). * P < 0.0001 diferencia significativa entre los protocolos de ejercicio.
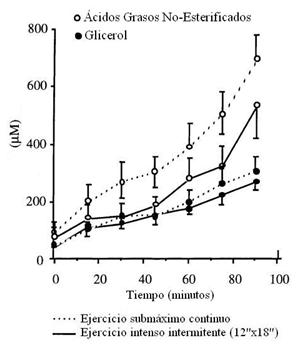
Figura 3. Concentración plasmática de
los ácidos grasos no esterificados y de glicerol durante el ejercicio submáximo continuo (líneas
punteadas) y durante el ejercicio intenso intermitente (líneas sólidas). * P < 0.05 diferencia significativa entre
los protocolos de ejercicio.
La concentración de glicerol aumentó desde el reposo y a lo largo del ejercicio, en ambos protocolos sin diferencias significativas (ver Figura 3). La concentración de ácidos grasos no estirificados (~AGL) aumentó en forma continua desde los 15’ y durante el ejercicio en ambos protocolos, a pesar de haber una menor oxidación de lípidos durante el ejercicio intermitente.
Otro estudio de Christmass (1999) en el que también analiza las mismas variantes que en el estudio anterior, pero donde en donde un grupo de sujetos sanos realizaron 2 protocolos de carrera intermitente en cinta ergométrica. Estos 2 protocolos tenían una similar velocidad de carrera (22 km·h-1 igual al 109±5% del VO2pico) y una similar duración (40 minutos) e igual relación trabajo y pausa 1:1,5 (IC, intermitente corto: 6”:9”; e IL, intermitente largo, 24”:36). Como podemos ver en la Figura 2, la oxidación de los lípidos fue menor durante el IL comparado al IC (11µmol·kg-1·min-1 vs 30µmol·kg-1·min-1, respectivamente), aunque no hubo cambios a lo largo del ejercicio en ambos protocolos. Pero, sí hubo una mayor oxidación de CHO en todo momento durante el protocolo de ejercicio IL comparado al IC. Y el R fue más alto durante el IL (0.95) comparado al IL (0.88). La concentración de glicerol en plasma aumentó desde el reposo hasta los 32' en ambos protocolos. Sin embargo, el glicerol fue menor (P < 0.05) en el ejercicio IL que en el IC (197±19 µM vs 246±19 µM, respectivamente).
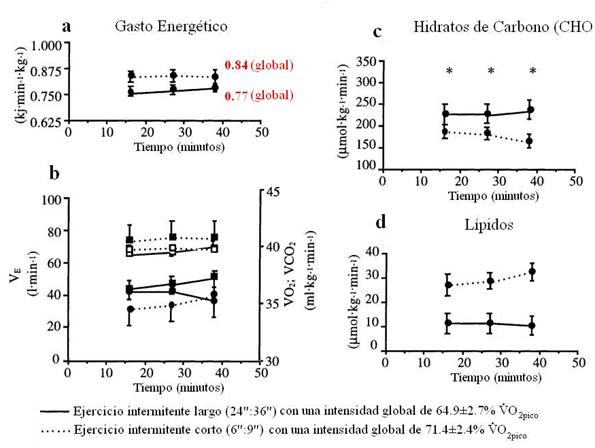
Figura 4. a,
Gasto energético; b, volumen expirado (VE)(□),
consumo de oxígeno (VO2) (●), y
producción de dióxido de carbono (VCO2)
(∎); c,
tasa de oxidación de CHO; d, tasa de oxidación de lípidos,
durante el ejercicio intermitente largo (24":36") y corto
(6":9").
* P
< 0.05 Diferencia significativa entre los protocolos de ejercicio.
La concentración de los
AGL fue más alta al final del ejercicio respecto al valor de los 10' iniciales,
pero a pesar de ese aumento y de una menor tasa de oxidación de lípidos durante
el ejercicio IL, no hubo diferencias entre ambos protocolos (537±74 µM y 883±148
µM, para el IL e IC, respectivamente). Con el lactato y el piruvato ocurrieron diferencias
significativas, siendo más alto tanto el lactato como el piruvato en plasma en
el ejercicio IL que en el IC (ver Figura 3).
Esto significa que la oxidación de los lípidos es aproximadamente 3 veces menor
y la oxidación de los CHO 1.3 veces mayor durante el ejercicio IL comparado al
ejercicio IC. Por lo tanto, estos aumentos en la disponibilidad del lactato y
el piruvato, como producto de una rápida glucólisis durante el ejercicio
intermitente de alta intensidad, puede resultar en una caída en la oxidación de
los lípidos, inhibiendo el transporte de ácidos grasos de cadena liviana hacia la
mitocondria. Christmass M.A. concluyó que factores
metabólicos dentro de las fibras musculares oxidativas eran los responsables de
la diferencia en la utilización de los sustratos entre el ejercicio IL y el
ejercicio IC; y que la duración del período de trabajo determina la respuesta
metabólica al ejercicio intermitente.
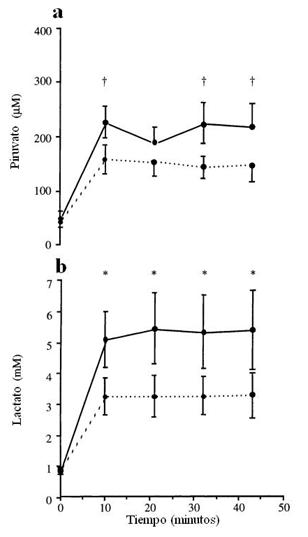
Figura 5.Concentraciones plasmáticas de Piruvato (a) y de lactato (b) durante el ejercicio intermitente largo (24":36") y corto (6":9"). † P < 0.01, * P < 0.001 Diferencias significativas entre los protocolos de ejercicio.
Una variante que Christmass hizo un par de años después (2001) sobre este patrón de protocolos corto (6”:9”) y largo (24”:36), fue introducir un turno de 10 minutos de ejercicio intenso intermitente (24”:36”) y luego descansar 45 minutos en forma pasiva, y luego realizó los dos protocolos anteriores. Encontró que esta variante (la del ejercicio previo, en forma intermitente e intensa), cambió sustancialmente el uso de sustrato durante el subsecuente ejercicio intermitente, ya que la oxidación de CHO fue menor (P ≤ 0.05) y la oxidación de los lípidos fue más alta (P ≤ 0.01) durante el protocolo con ejercicio previo comparado al protocolo sin ejercicio previo. En contraste, el patrón de oxidación de sustratos durante el ejercicio intermitente corto fue al parecer similar entre los 2 experimentos (ver Tabla 1).
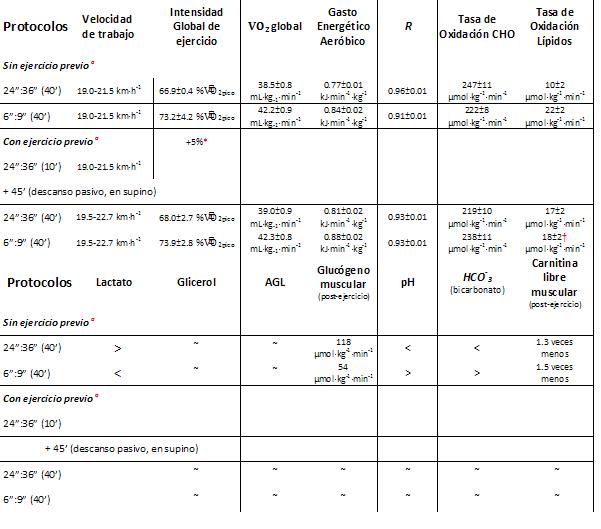
Tabla 1. Descripción de los protocolos y sus resultados. aLa intensidad global durante el período de ejercicio en el protocolo sin ejercicio previo fue de 114% VO2pico y durante el protocolo con ejercicio previo fue de 111% VO2pico. *Respecto al protocolo 1 (sin ejercicio previo). † Más alto respecto al ejercicio sin ejercicio previo. ≈ sin cambios significativos.
Una de las principales
conclusiones que se pueden hacer respecto a estos resultados de los estudios de
Christmass es que contradicen los hallazgos de Essen (que analizamos la semana
pasada) en lo referente a la oxidación de lípidos, ya que en el último caso (el
estudio de Christmass) fue 3 veces menor la tasa de oxidación durante el
ejercicio intermitente respecto al ejercicio continuo. Como una suerte de
objeción, se puede decir que los ritmos de oxidación de los CHO y los lípidos
alcanzados durante el ejercicio intenso intermitente y ejercicio submáximo
continuo no son equivalentes en todos los niveles del ejercicio, a pesar de un
gasto energético global similar. Una explicación podría ser que al haber una
depleción del glucógeno (cuando cae 20-30 µmol·kg-1
de peso corporal) el suministro de energía proveniente del glucógeno se ve
reducido, con lo cual actúa como un factor limitante en el ejercicio prolongado
(ejemplo, deportes como el fútbol). También, un dato llamativo del estudio de
Christmass es que el R y el lactato
fueron más altos durante el ejercicio intermitente comparado al ejercicio
continuo con igual potencia global, lo cual coincide con la tasa más alta de
oxidación de CHO y más baja la oxidación de lípidos, en contraste con los
resultados de Essen. De todas formas, la
diferencia entre estos estudios, está en la intensidad global de ejercicio, lo
que influencia esencialmente la diferencia de utilización de sustrato entre los
protocolos de ejercicio. Mientras que en el estudio de Essen era del ≈55% VO2máx,
en el estudio de Christmass es del ≈70% VO2pico, es decir, bastante
mayor, como resultado de una intensidad del período de trabajo más alto (120% VO2pico,
en el estudio de Christmass, y 100% % VO2máx, en el estudio de
Essen) durante el ejercicio intermitente, además que uno se realizó en
cicloergómetro (Essen) y otro en cinta ergométrica (Christmass). Por lo tanto,
una mayor utilización de CHO (y una menor utilización de lípidos) con el
aumento de la intensidad está bien establecido históricamente, y una fuerte
relación entre la mayor intensidad de ejercicio y el valor R está demostrado también que ocurra durante el ejercicio
intermitente. La tasa más rápida de oxidación de CHO alcanzada en el ejercicio
intermitente fue acompañada por casi el 100% de aumento del lactato y el
piruvato. Y lógicamente, un mayor ritmo de glucogenólisis durante el ejercicio
intermitente está asociado a una mayor intensidad de ejercicio en el período de
trabajo. A esto se le debe agregar, la baja disponibilidad de O2 en
el músculo, lo cual potencia a la glucólisis y a la glucogenólisis, además de
aumentos citosólicos de ADP, AMP y Pi. El otro factor clave fue que
el suministro de O2 requerido durante el ejercicio intermitente no
pudo igualar a la demanda de O2, como se ve reflejado en la caída
más baja de la HbO2 (ver Figura 6) durante el período de trabajo
intermitente, y su correlación con la velocidad de carrera.
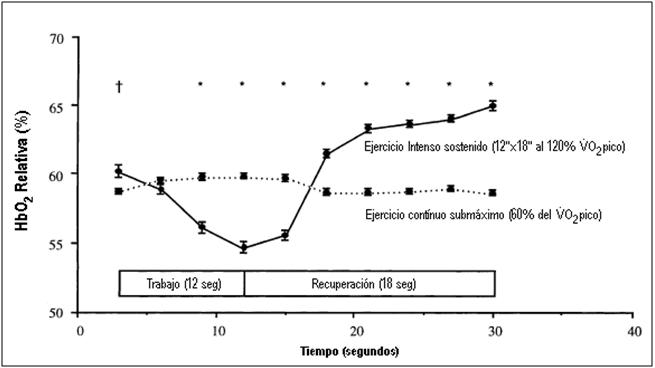
Figura 6. Oxigenación relativa del músculo cuádriceps durante el período de
análisis (un ciclo de trabajo y pausa) de datos en el ejercicio intermitente
sostenido y ejercicio continuo submáximo. † P
< 0.05, * P < 0.001, diferencias significativa entre los protocolos
de ejercicio (Christmass, 1999).
Es decir, existe una pequeña hipoxia en los tejidos durante el período de trabajo, lo cual provoca la utilización temprana de CHO y, por lo tanto, incide en la utilización de los lípidos. También, la duración del período de trabajo durante el ejercicio intermitente de intensidad constante influencia el patrón del uso de sustrato. Cuánto más largo es el período de trabajo, menos disponibilidad de O2 existe, lo cual implicará una tasa alta de metabolismo de los CHO y una disminución de la oxidación de los lípidos ocurrirá. Este panorama cambia cuando se realiza un ejercicio previo y se descansa 45’ en forma pasiva como ya vimos, donde aumenta la oxidación de los lípidos y disminuye la depleción de los CHO. Así, esta ‘preservación’ de los depósitos de CHO en el ejercicio prolongado con ejercicio previo (tan sólo 10’ de ejercicio intenso intermitente) podría servir como estrategia de una entrada en calor con dicho objetivo de ahorro de CHO.
Un mecanismo para explicar esto podría ser el siguiente: las catecolaminas podrían contribuir al aumento en el metabolismo energético observado durante la pausa del ejercicio (exceso de consumo de oxígeno post-esfuerzo) a través de un aumento tanto de triglicéridos/AGL circulantes y oxidación de lípidos. Una mayor oxidación de AGL y menor actividad del ciclo del ácido cítrico durante el descanso pasivo después de un breve ejercicio intenso intermitente, podría haber impedido el subsiguiente flujo glucolítico durante los ejercicios intermitentes corto y largo por medio de aumentos en el acetil CoA y el citrato.
Este aumento de la acetil CoA durante la pausa provoca el aumento de acetilcarnitina y a su vez menor carnitina libre pre-ejercicio. Así, niveles altos de carnitina libre pre-ejercicio después de un breve ejercicio intenso seguido de un descanso pasivo observado en el estudio de Christmass indican una formación mínima de acetilcarnitina. Pero no hay una explicación definitiva del por qué del enlentecimiento del metabolismo de los CHO cuando el ejercicio intermitente largo (24”:36”) es precedido por un breve ejercicio intenso y posterior recuperación pasiva. Una prueba de ello, es que con similar intensidad en el período de trabajo (114% VO2pico, en el protocolo sin ejercicio previo y subsiguiente descanso pasivo, y 111% VO2pico con ejercicio previo y descanso pasivo, e idéntica duración total de trabajo (16’), la diferencia en la oxidación global de las grasas corporales durante el ejercicio intermitente largo realizado sin ejercicio breve intenso, comparado con ejercicio previo intenso y recuperación pasiva, podría ser una causa de un/os factor/es dentro del mismo músculo (ver Figura 7).
Nota: La enzima carnitina palmitiltransferasa I (CPTI) de la membrana mitocondrial externa elimina la coenzima A de la molécula de acil-CoA y, a la vez, la une a la carnitina situada en el espacio intermembrana, originando acetilcarnitina, la cual penetra a la membrana interna mitocondrial y gana acceso a una β-oxidación. El CoA queda libre en el citosol para poder activar otro ácido graso.
Una situación parecida al ejercicio intermitente, es el ejercicio de alta intensidad, donde la disponibilidad de AGL se encuentra reducida. Pero también, se ha demostrado que esta menor disponibilidad de AGL puede solamente explicar en parte la reducida oxidación de lípidos en esas condiciones. Como se ha visto en estudios previos, la concentración plasmática de AGL y el ritmo de aparición de AGL (RaAGL) fueron muy bajos a esta intensidad (85% V2máx, carrera continua, por ejemplo). Y aún infundiendo intralípidos y heparina, la oxidación de lípidos solamente aumentaba muy poco, incluso menos que a intensidades moderadas; lo cual, esto indica que la oxidación de AGL es al menos en parte regulada a nivel muscular.
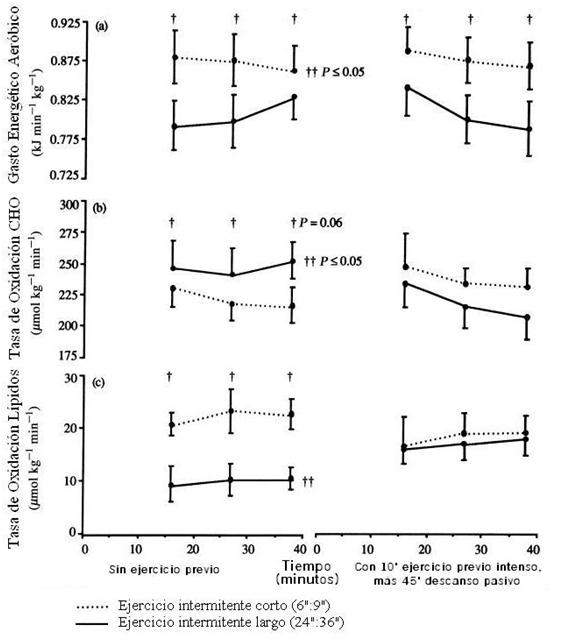
Figura 7. Gasto energético, tasa de oxidación de CHO y tasa de oxidación de lípidos durante el ejercicio intermitente con ciclo de trabajo:pausa de 24”:36” (líneas sólidas) o 6”:9” (líneas punteadas). Un grupo completó el ejercicio intermitente sin ejercicio previo, y otro grupo con ejercicio previo intenso (10’) más 45’ de descanso pasivo antes de realizar el ejercicio intermitente (40’). Los valores son promedios ± (DS). † Diferencia significativa entre los protocolos (ejercicio intermitente corto vs largo). †† Diferencia significativa entre los experimentos (con o sin ejercicio previo).
Un aumento de la intensidad del ejercicio podría reducir la oxidación de lípidos en parte por inhibición del transporte mitocondrial de ácidos grasos de larga cadena (AGLC). Así un mayor flujo glucolítico podría reducir ese transporte de AGLC por inhibición de la carnitina palmotil-transferasa I (CTP I). Dicha enzima es esencial para transferir los AGLC a través de la membrana mitocondrial y es inhibida por la malonil CoA. Pero hay hallazgos que indican de una menor transferencia de AGLC hacia las mitocondrias en presencia de un ‘acelerado’ flujo glucolítico (como ocurre en el ejercicio intermitente) en humanos es poco probable que implique la inhibición de la CPT I por la malonil CoA. Entonces una rápida formación de acetilcarnitina con el mayor flujo glucolítico, podría depletar los niveles de carnitina libre, provocando así un mecanismo para la inhibición de la oxidación de lípidos cuando el metabolismo de los CHO está ‘acelerado’. Así, una limitación en la transferecnia de AGLC podría ser responsable de la declinación en la oxidación de lípidos en paralelo con la caída en la disponibilidad de carnitina libre durante el ejercicio intenso.
Nota: Los AGLC son ácidos grasos de 14 a 24 carbonos, derivados de la lipólisis del tejido adiposo (o por hidrólisis intracelular de triglicéridos) es el mayor combustible oxidado por los músculos activos, especialmente cuando el ejercicio es prolongado y la intensidad de baja a moderada. La regulación del metabolismo de los AGLC en el músculo ocurre en varios pasos celulares, incluyendo transporte de AGLC a través de la membrana plasmática, transporte de AGLC dentro del citosol, y metabolismo intracelular de los AGLC. La entrada a la mitocondria es un sitio de regulación. Los AGLC requieren de un transportador para entrar a la mitocondria (a través de la carnitina), lo cual puede limitar la oxidación de los AGLC durante el ejercicio de alta intensidad. Una vez que está en la mitocondria, los AGLC se someten a la β-oxidación e ingresan al ciclo de Krebs y producen energía para formar ATP.
Nota: carnitina palmotil-transferasa I (CTP I): Enzima de la membrana interna mitocondrial. Transporta AGLCs uniéndolos a moléculas de la carnitina.
Y según la Figura 8, podemos observar que el contenido de carnitina libre, en el estudio de Christsmass, aumenta 1.3 veces más post-ejercicio en el ejercicio intermitente largo (con ejercicio previo+descanso) comparado al ejercicio intermitente (sin ejercicio previo). Esto ocurrió en presencia de un menor metabolismo de CHO y una mayor oxidación de lípidos durante el ejercicio intermitente largo con ejercicio previo comparado al ejercicio intermitente largo sin ejercicio previo. En contraste, un aumento de 1.7 veces más alto post-ejercicio del nivel de carnitina durante el ejercicio intermitente corto (con ejercicio previo+descanso) respecto al ejercicio intermitente corto (sin ejercicio previo) fue observado a pesar de similares tasas de oxidación de lípidos (ver Figuras 6 y 8).
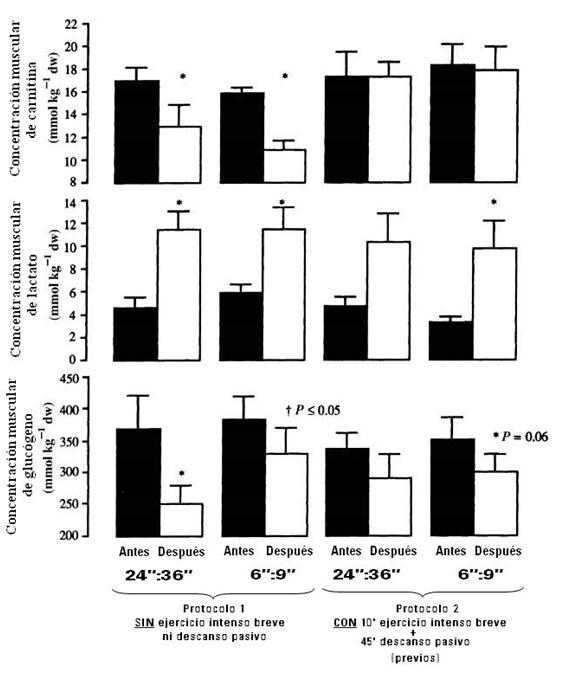
Figura 8. Concentración del músculo esquelético de carnitina, lactato y glucógeno antes (∎) y después (ÿ) de ejercicio intermitente con un ciclo de trabajo:pausa de 24”:36” (largo) o de 6”:9” (corto). Un grupo lo realizó sin ejercicio previo (40’ de ejercicio intermitente, Protocolo 1) y otro grupo lo realizó con un ejercicio breve intenso intermitente (24”:36”) de 10’ seguido de un descanso pasivo de 45’, antes de realizar 40’ de ejercicio intermitente (Protocolo 2). Los valores son promedios ±SD. † Diferencia significativa entre los protocolos. * Diferencia significativa antes, comparado a después, del ejercicio intermitente.
Es decir, la caída en el contenido de carnitina libre muscular fue casi idéntica en ambos protocolos de ejercicio (Figura 8) a pesar de más del doble más bajo del ritmo de oxidación de lípidos durante el ejercicio intermitente largo (24”:36”). Por lo tanto, estos resultados no apoyan la propuesta de que el flujo rápido (acelerado) glucolítico reduce la disponibilidad de carnitina libre y así inhibir la oxidación de lípidos bajo condiciones de ejercicio intermitente (como sí lo refería Essén en sus estudios). En ese estudio de Christmass no hubo diferencias en los valores pre- y post-ejercicio en el contenido de carnitina libre muscular entre el ejercicio intermitente largo y corto durante el protocolo con ejercicio previo seguido de descanso, en una respuesta similar a la del protocolo sin ejercicio previo. Recordemos que la concentración de carnitina en sus 2 formas (acetilcarnitina y carnitina libre) dentro del músculo se mantiene constante aún durante el ejercicio, donde aumenta en su forma de acetilcarnitina. Y la caída de la carnitina libre en el protocolo 1 (Ver figura 8), significa que el ritmo de formación de acetil-CoA fue mayor que la tasa de oxidación dentro del ciclo de Krebs. En cambio, con el ejercicio previo (protocolo 2) la formación y oxidación de acetil-CoA se balanceó y, por lo tanto, la carnitina no decayó (Figura 8). Por lo tanto, los datos del estudio de Christmass (2001) indican que la disponibilidad de carnitina libre es poco probable que haya influenciado las tasas de oxidación de lípidos durante el ejercicio intermitente corto o largo, aunque se desconocen las causas de la caída de la oxidación de lípidos durante el ejercicio intermitente largo. Un estudio reciente de Benjamin T. Wall (2011) donde se ingirió en forma oral L-carnitina, se demostró que se podía incrementar el contenido muscular de carnitina (a través de la dieta, +21% de aumento, durante 24 semanas) y que resultaba en un ahorro de CHO durante el ejercicio de baja intensidad (50% del VO2máx en forma continua, y por tanto, se utilizan más lípidos como sustrato energético) y hay una mejor igualación del flujo glucolítico, mitocondrial y del complejo piruvato deshidrogenasa, en los ejercicios de alta intensidad (80% del VO2máx), con la consecuente reducción de generación de energía anaeróbica de ATP. Estos cambios, a su vez, fueron acompañados de un mejor rendimiento (+35% de aumento).
Con lo cual, los datos de los estudios de Christmass, no apoyan el concepto de que diferencias en la disponibilidad de oxígeno en el músculo, como resultado de diferencias en la duración del período de trabajo, sean la principal influencia sobre el metabolismo de los CHO durante el ejercicio intermitente intenso con constante período de trabajo. Es decir, los estudios de Christmass contradicen los estudios de Essén respecto al metabolismo de los lípidos en esfuerzos intermitentes, a pesar de pequeñas diferencias en los protocolos utilizados, pero también, los de Christmass se acercan mucho más a la 'realidad práctica' del ejercicio intermitente actual.
Referencias
- Christmass M.A., Dawson B., Passeretto P., Arthur P.G. A comparison of skeletal muscle oxygenation and fuel use in sustained continuous and intermittent exercise. Eur J Appl Physiol Occup Physiol, Oct 1999; 80(5): 423-35.
- Christmass M.A., Dawson B., Arthur P.G. Effect of work and recovery duration on skeletal muscle oxygenation and fuel use during sustained intermittent exercise.Eur J Appl Physiol Occup Physiol, Oct 1999; 80(5): 436-47.
- Clifford, P.S.; Tschakovsky, M.E. Rapid Vascular Responses to Muscle Contraction; Exerc Sport Sci Rev. 2008 Jan;36(1):25-9.
- Wray D.W., Donato A.J., Abhimanyu Uberoi J.P. Merlone, Richardson R.S. Onset exercise hyperaemia in humans: partitioning the contributors. J Physiol 565: 1053–1060. (2005).
- Carzola G., Petibois S.C., Leger L. Lattato ed esercizio: miti e realtà, Rivista SDS N° 53 ottobre-dicembre 2001, Romana Editrice.
- Wittenberg B.A., Wittenberg, J.B. Transport of oxygen in muscle.Ann. Rev. Physiol.51,857 -878 (1989).
- Christmass M.A., Dawson B., Arthur P.G. Goodman C. Brief intense exercise followed by passive recovery modifies the pattern of fuel use in humans during subsequent sustained intermittent exercise. Acta Physiol Scand 2001;172: 39-52.
- Essen, B. Studies on the regulation of metabolism in human skeletal muscle using intermittent exercise as an experimental model. Acta Physiol Scand Suppl. 454: 1-32. 1978.
- Maughan R., Gleeson M. Biochemical Basis of Sports Performance, The 1st Edition. Oxford University Press, 2004.
- Vuorimaa T., Vasankari T., Mattila K., Heinonen O., Häkkinen K., Rusko H.Serum hormone and myocellular protein recovery after intermittent runs at the velocity associated with VO2max. Eur J Appl Physiol Occup Physiol, (1999) 80: 575-581.
- Jeukendrup, A.E. (2002). Regulation of Fat Metabolism in Skeletal Muscle. Ann N Y Acad Sci 967, 217-235.
- Wolinsky I., Driskell J.A. Sports Nutrition; Energy Metabolism and Exercise. CRC Press, Taylor & Francis Group (2008).
- Hargreaves M., Spriet L.L. Exercise Metabolism. Human Kinetics. 2nd Edition (2006).
- Driskell J.A. Sports Nutrition; Fats and Proteins. CRC Press; Taylor & Francis Group (2007).
- Sidossis L.S., Gastaldelli A., Klein S., Wolfe R.R. Regulation of plasma fatty acid oxidation during low- and high-intensity exercise. Am J Physiol 1997; 272: E1065-E1070.
- Plowman, S.A.; Smith, D.L Exercise physiology for health, fitness, and performance; 2nd ed.; San Francisco: Benjamin Cummings, Pearson Education, Inc . 2003.
- Wall BT, Stephens FB, Constantin-Teodosiu D, Marimuthu K, Macdonald IA, Greenhaff PL. Chronic oral ingestion of L-carnitine and carbohydrate increases muscle carnitine content and alters muscle fuel metabolism during exercise in humans. J Physiol. 2011 Feb 15;589(Pt 4):963-73.