
El por qué de la glucólisis: (lenta y rápida)
Publicado 23 de abril de 2013, 13:08

Autor: Raul Herrero
La glucólisis
La mayoría de los profesionales que estamos en las ciencias del ejercicio conocemos la importancia de la glucólisis como sistema de producción de energía. Sin embargo pocos nos preguntamos para qué existe. Si la célula puede usar todos sus combustibles oxidativamente, ¿por qué existe la glucólisis anaeróbica?
Comúnmente diríamos: porque la presión parcial (PP) de O2 en la mitocondria no es suficiente o por qué no puede entregar energía a una alta velocidad. Pero es interesante saber cuál debería ser la presión de O2 en la mitocondria como para que la glucosa se degrade oxidativamente? o ¿cuál es la PP que hay cuando se degrada en forma parcial anaeróbicamente?
Lo más probable es que la PP de O2 no sea la única variable de la problemática.
Pues bien, la PP de O2 en la mitocondria es de aproximadamente 1mmHg y en el citoplasma de unos 5mmHg. Pero para que esta presión sea efectiva durante el ejercicio primero deben hacerse los ajustes circulatorios y respiratorios que conllevan a la vasodilatación arteriolar que permite la adecuada llegada (disponibilidad) del oxígeno al tejido. Por ello tendríamos un retraso en los ajustes de los sistemas transportadores de O2 lo que aparentemente justificaría la glucólisis anaeróbica en los primeros segundos de un ejercicio sobre todo intenso. Pero en otro sentido, ¿cómo justificaríamos con este argumento el hecho de estar en ritmo de trabajo aeróbico hacer un cambio de ritmo que nos vuelve a poner anaeróbicos? El argumento previamente explicado no sería del todo válido.
También tratándose de los comienzos de un ejercicio sería válido decir que hay una inercia metabólica (lo cual es cierto) ya que para poner en marcha la glucólisis solo requiere activar unas 10 enzimas mientras que poner en marcha la vía oxidativa implica más del doble de reacciones, pero este argumento solo sería válido al comienzo del ejercicio ya que una vez en ritmo aeróbico la inercia metabólica no podría explicar un cambio de ritmo con acumulación de lactato generado por la glucólisis.
Ensayemos entonces una opción válida que justifique la producción láctica cuando cambiamos el ritmo a partir de una fase aeróbica estable: la potencia de un sistema energético se mide en función de la capacidad de generar determinada cantidad de moles de ATP por segundo y claramente el sistema anaeróbico es mucho más potente que el aeróbico. Esta velocidad de producción de energía depende de las necesidades momentáneas de ATP y a mayor necesidad de ATP por segundo se necesita mayor potencia metabólica. De aquí está la cuestión por la cual el sujeto comienza a producir / acumular lactato cuandoacelera por encima de la fase estable de un ejercicio aeróbico.
El sistema energético aeróbico no puede proveer moles de ATP a la velocidad que sus músculos lo requieren por lo que usa la artimaña anaeróbica (glucólisis).
Esto se ve justificado más aún con la idea de que en el paso de la glucólisis que cataliza la reacción de gliceraldehido 3 fosfato (G3P) a 1,3 difosfoglicerato (1-3DPG), una molécula llamada NAD+ toma los H+ que libera dicha reacción y se reduce a NADH+H y para que esta reacción pueda seguir llevándose a cabo con el solo propósito de que la glucólisis pueda avanzar sin interrupciones, es necesario la presencia de NAD+ en el citoplasma. Sin embargo esta molécula ahora se encuentra como NADH+H por lo que no puede continuar tomando H+ hasta que se desprenda de ellos.
En situación de reposo o ejercicio liviano Brooks propuso que se genera una glucólisis lenta y que estos hidrógenos se envían a la mitocondria para ser procesados. En cambio durante contracciones musculares potentes este se produce a una velocidad tal que no le permite a los sistemas de transporte enviar este NADH a la mitocondria para sacarle estos H+ (reoxidarlo) y retransformarlo en NAD+ así que la célula debe recurrir a algún mecanismo lo suficientemente rápido que le permita oxidar el NADH+H+a NAD+ y así reusarlo.
El mecanismo que encontró la célula es reducir el piruvato, que acepta los H+ del NADH+H y transformarlo en lactato de esta forma volvemos a tener NAD+ rápidamente disponible para que la glucólisis no se interrumpa. Ver reacción general figura 1.
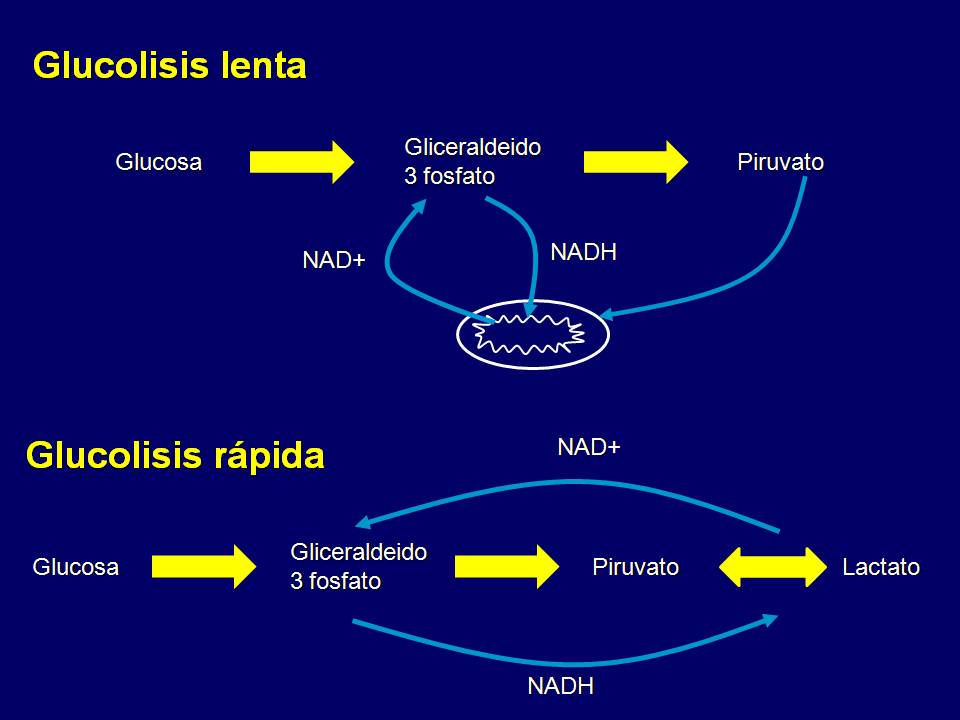
En conclusión como se ve en los comienzos de un ejercicio aún no siendo lo suficientemente intenso, produciremos lactato como consecuencia de la falta de los ajustes que aumenten la PP de O2en la mitocondria más el ajuste de la inercia metabólica. Mientras que cuando ya estamos en una fase estable aeróbica y aumentamos el ritmo lo suficiente la producción de lactato aumentará como consecuencia de que las necesidades momentáneas de ATP superan la capacidad de la célula muscular para sintetizar el mismo por la vía aeróbica, sumado a la incapacidad de la mitocondria de transportar el NADH+H y usar los H+ para la síntesis aeróbica de ATP. Podríamos concluir diciendo que la glucólisis es una oxido reducción interna.
Bibliografía:
Styer,L (1988) Biochemistry (2nd ed).W.H.Freeman,SanFransisco,CA.
Astrand,P. O. and Rodahl (1986) Textbook of work physiology (3er ed) McGraw-Hill.
Lehninger. Principios de bioquímica.David L. Nelson and Michael M. Cox.Edi. Omega.
Role of mitochondrial lactate dehydrogenase and lactate oxidation in the intracellular lactate shuttle. GEORGE A. BROOKS*, HERVE´ DUBOUCHAUD, MARCIA BROWN, JAMES P. SICURELLO, AND C. ERIC BUTZProc. Natl. Acad. Sci. Vol. 96, pp. 1129–1134, February 1999