
El Rol de la Masa Muscular Activa sobre la Translocación Cardiovascular Inducida por el Ejercicio
Stylianos N Kounalakis, George P Nassis, Maria D Koskolou y Nickos D Geladas
Department of Sport Medicine and Biology of Exercise, Faculty of Physical Education and Sport Science, University of Athens, Greece.
Artículo publicado en el journal PubliCE del año 2014.
Publicado 20 de mayo de 2014
Resumen
Palabras clave: ciclismo prolongado, regulación cardiovascular
INTRODUCCION
Durante el ejercicio prolongado en estado estable, se produce un progresivo incremento en la frecuencia cardíaca (HR) y una reducción en el volumen sanguíneo (SV) seguido de una caída en el gasto cardíaco (CO) y en la presión arterial media (MAP). Este fenómeno, conocido como translocación cardiovascular (CVdrift), puede afectar negativamente el rendimiento (Ganio et al., 2006). Se sabe que el CVdrift es afectado por al hipertermia (Gonzalez-Alonso et al., 1998), la deshidratación (Montain and Coyle, 1992) y el incremento en la frecuencia cardíaca (Fritzsche et al., 1999). Además, la acumulación de sangre en la periferia (Rowell et al., 1969) parece contribuir al CVdrift. Las interacciones entre estos factores pueden aumentar el CVdrift (Gonzalez-Alonso et al., 2000). A pesar de la contribución de los factores previamente mencionados en la aparición del CVdrift , su causa no se comprende totalmente (Coyle and Gonzalez-Alonso, 2001). El CVdrift es más pronunciado durante el ciclismo que durante la carrera a pesar de la mayor carga térmica, deshidratación e incremento de la frecuencia cardíaca observadas durante la carrera (Nassis and Geladas, 2002), lo que sugiere que la masa muscular activa desempeña algún tipo de rol. Se ha especulado que la magnitud de la masa muscular involucrada en el ejercicio puede ser un factor importante que contribuya al CVdrift y en definitiva a la desmejora del rendimiento durante un esfuerzo muscular prolongado. La mayor participación muscular resulta en una mayor activación simpática, la cual es estimulada por reflejos musculares y por comandos centrales, tal como lo indica la acción eléctrica muscular y el índice de esfuerzo percibido (Franke et al., 2000; Nobrega et al., 1994; Pawelczyk et al., 1997). Por un lado, el consumo de oxígeno (VO2) durante la realización de ejercicios con una pierna es más de la mitad del medido durante el ejercicio de ciclismo a dos piernas, lo cual presumiblemente puede inducir diferencias marcadas en el turnover de energía muscular y en las respuestas metabólicas (Lewis et al., 1985). Por otro lado, Klausen et al (1982) indicaron que durante la realización de 8-10 min de ciclismo con una y dos piernas a la misma intensidad relativa (70% del VO2pico, específico para cada una de las condiciones), las respuestas del CO y de la HR no fueron estadísticamente diferentes. Consecuentemente, la respuesta hemodinámica no se vería afectada por la masa muscular, siempre que la intensidad del ejercicio sea proporcional a la máxima capacidad por pierna. Para distinguir el efecto de la masa muscular del de la intensidad del ejercicio, se debería adoptar un paradigma de ejercicio con 1 y 2 piernas al mismo consumo de oxígeno absoluto. Si bien se han utilizado paradigmas en los cuales los sujetos se ejercitaron a la misma tasa de trabajo proporcional (el consumo de oxígeno y la intensidad relativa son diferentes, Jensen-Urstad et al., 1994), no existen datas respecto de las respuestas de la HR y del CO durante la realización de ejercicios prolongados llevados a cabo al mismo VO2 por pierna.
En el presente estudio se exploró el rol de la masa muscular activa sobre el CVdrift. Se hipotetizó que durante el ciclismo a una y dos piernas al mismo VO2 absoluto por pierna, el CVdrift sería mayor durante el ejercicio de ciclismo con dos piernas debido a un mayor comando central. Además se hipotetizó que el mayor CVdrift durante el ejercicio de ciclismo a dos piernas se debería a la mayor frecuencia cardíaca.
METODOS
Sujetos
Doce sujetos con una edad de 23.4 ± 3.8 años, una masa corporal de 77.8 ± 7.3 kg y un VO2pico de 3.52 ± 0.52 L·min-1 (medias ± DE), fueron voluntarios para participar en este estudio. Todos los sujetos dieron su consentimiento informado por escrito para participar en los experimentos luego de que se le informaran los detalles de los procedimientos y los posibles riesgos.
Sesiones Pre-Experimentales
Antes de las pruebas principales, cada sujeto visitó el laboratorio cuatro a cinco veces, para su familiarización con el instrumental, la metodología y la evaluación antropométrica. Además, cada sujeto realizó una prueba de ejercicio submáxima a dos piernas (4 etapas de 4 minutos de duración) o un test de ejercicio máximo en un cicloergómetro con cupla electromagnética (Lode RH, the Netherlands) para determinar la relación entre el costo de oxígeno, la carga de trabajo y el VO2pico. El VO2pico fue definido como el mayor valor de VO2, promediado cada 30s, siempre que se cumplieran con los siguientes criterios: (1) se alcance una meseta en el VO2 a pesar del incremento en la carga, (2) se alcance un índice de intercambio respiratorio mayor a 1.1 y (3) se alcance una concentración de lactato sanguíneo mayor de 9 mmol·L-1 (Accutrend, USA) 3 minutos después de la finalización del ejercicio.
Procedimiento
Tres días antes de la prueba principal, se les pidió a los sujetos que registren sus dietas y el ejercicio realizado y que replicaran estos hábitos antes de cada condición. Además, se les pidió a los sujetos que se abstengan de realizar actividad física, consumir cafeína o alcohol en el día previo a los experimentos. Los experimentos se llevaron a cabo a la misma hora del día, al menos tres horas después de que los sujetos consumieran una comida liviana. Luego de que los sujetos vaciaran sus vejigas, se midió la masa corporal (Bilance Salus, Italy) y se fijaron los electrodos de electromiografía (EMG) y de electrocardiografía (ECG) a los extensores de la pierna (vasto lateral y vasto medial) y al pecho, respectivamente. Además, se colocó un termistor en el recto de los sujetos, a una profundidad de 13-15 cm del esfínter. Luego de esta preparación, los sujetos realizaron una entrada en calor de 8 min en el cicloergómetro a un 50% de su VO2pico. Luego de 5 min de estiramientos, se determinó la fuerza isométrica máxima de extensión de la rodilla (MCV) utilizando un dispositivo para el entrenamiento de la fuerza modificado conectado a una celda de carga calibrada. Cada sujeto realizó tres pruebas máximas de cinco segundos con un minuto de recuperación entre cada prueba, para obtener así la máxima señal de EMG. Si no se conseguía una diferencia mayor al 5% entre las pruebas, se llevaba a cabo una cuarta prueba y luego se promediaban los valores de las pruebas con mayores valores. Posteriormente, los sujetos se relajaron durante 20 min en el cicloergómetro mientras se colocaba el instrumental. Se recolectaron muestras de sangre capilar, por duplicado, de 10μL y de 50-60μL en la yema del dedo índice derecho para la determinación de la concentración de hemoglobina (Hb) y del hematocrito (Hct), respectivamente. En esta primera condición, los sujetos pedalearon a 80rpm durante 50-55 min con ambas piernas al 60% del VO2pico obtenido durante el test de ciclismo con las dos piernas. La segunda condición fue llevada a cabo tres días después y la carga externa fue ajustada para que se alcanzara la mitad del VO2 observado en la primera condición. Se utilizó el mismo consumo de oxígeno por pierna, en lugar de la mitad de la carga de trabajo (Watt), ya que esto último podría provocar una mayor respuesta metabólica y un mayor turnover energético en la condición de ciclismo con 1 pierna (Jensen-Urstad et al., 1994). La intensidad absoluta de ejercicio fue de 126 ± 1 Watts en el ejercicio de ciclismo con dos piernas, mientras que en la condición de ejercicio de ciclismo con una pierna, la intensidad absoluta de ejercicio fue ~9% menor de la mitad de la intensidad absoluta de ejercicio de ciclismo con dos piernas (carga de trabajo: 126/2-9% = 57 Watts), calculada de acuerdo a lo propuesto por Jensen-Urstad et al., 1994. Durante la condición de ciclismo a una pierna, la pierna que no realizaba ejercicio descansó entre los pedales, sobre el marco del cicloergómetro, y el retorno del pedal se produjo mediante un resorte eléctrico sujeto al manubrio. Los sujetos se familiarizaron con el montaje experimental para el ciclismo con una pierna un día antes del experimento y durante 20 minutos. Para asegurar el desarrollo del CVdrift los sujetos se ejercitaron sin ventilador y sin consumir fluidos. En todos los experimentos, la temperatura ambiente fue de 23.5 ± 0.4ºC y la humeada relativa ambiente fue de 56.2 ± 4.1%.
El CO se determinó entre los minutos 15-22, 30-37 y 47-54 utilizando el método de re-inhalación exponencial de CO2 (Defares, 1958) con mediciones triplicadas separadas por 2 min para asegurar el estado estable fisiológico. En el caso de que las mediciones difirieran más del 5%, se realizó una cuarta maniobra y se promediaron los resultados de todas las pruebas. El VO2 se midió respiración por respiración y los datos se promediaron cada 10 s entre el minuto 1-8 se ejercicio y a partir de allí cada 2-3 min antes de cada determinación del CO. La oxigenación muscular (saturación de oxígeno, StO2) y el volumen sanguíneo (hemoglobina total, HbT) se midieron mediante espectroscopia infrarroja. A los minutos 15, 25 y 40 de ejercicio se registraron los datos de la actividad electromiográfica. Se realizaron mediciones por duplicado del presión sanguínea en reposo y al minuto 25 de ejercicio (antes de la medición del CO) y antes del final del ejercicio utilizando un sistema automático (Criticon Dinamap, Finland) en posición estándar, con el brazo del sujeto sostenido por el investigador a la altura del corazón. En el caso de que la diferencia entre las mediciones fuera mayor a 5 mmHg, se realizó una tercera medición. La frecuencia cardíaca (Polar S- 810, Finland) y al temperatura corporal se registraron cada 5 minutos. Además, la HR fue registrada latido a latido en un período de 5 minutos en reposo y entre los minutos 8-13 y 37-42 de ejercicio. Estos datos de la HR fueron utilizados para evaluar la variabilidad de la HR (HRV). En los mismos períodos, se midió la frecuencia respiratoria (Rf) a 500 Hz, con una termocupla de respuesta rápida (TSD 202A, Biopac systems, USA) unida a la válvula de respiración. El flujo sanguíneo en el antebrazo (FskBF) fue registrado mediante flujometría Laser Doppler en reposo y durante un minuto a los minutos 15 y 44 de ejercicio. La sensación de fatiga local (piernas) y general de los sujetos se registró cada 5 minutos utilizando la escala de esfuerzo percibido de Borg (RPE).
Métodos Analíticos y Equipamiento
Para la medición del VO2 se utilizó un sistema de medición del metabolismo (MedGraphics, CPX-D, USA), el cual fue calibrado antes del ejercicio con dos mezclas diferentes de gases. El CO se determinó mediante el método de re-inhalación de CO2 (Defares, 1958) modificado por DaSilva et al (1985) y Nassis y Geladas (2002). Los sujetos respiraron de una bolsa que contenía 1.5-2 veces el volumen corriente a una frecuencia de 50 respiraciones por minuto, establecida con el uso de un metrónomo. De acuerdo con el método de re-inhalación exponencial de CO2, el CO se calcula utilizando la ecuación indirecta de Fick. Las tres mediciones del CO en los momentos establecidos fueron promediadas. El coeficiente de variación con diferentes intensidades de ejercicio varió de 2.4 a 3.6%, lo cual aseguró que las mediciones del CO fueron confiables, dados que los valores obtenidos con el método directo se encuentran entre el 5-10% (Warburton et al., 1999).
Los cambios en la StO2 y en la HbT fueron registrados utilizando espectroscopia infrarrojo cercano (NIRS) (In Spectra325, Hutchinson Technology Inc, USA). Los principios operativos del dispositivo de NIRS pueden encontrarse en diversos artículos (Kounalakis et al. 2008). La sonda de 25mm fue ajustada y asegurada mediante un parche adhesivo a la piel afeitada y limpia en el aspecto medial del músculo recto femoral. Antes de cada medición, el dispositivo de NIRS fue calibrado utilizando un estándar de luz dispersa validado con referencias estándar equivalentes al 38% y 90% de saturación de hemoglobina.
La actividad del EMG, que indició el nivel relativo de actividad muscular, y la MVC de los extensores de la rodilla fueron medidas con la rodilla en un ángulo de 60º (0º = extensión completa). Se utilizaron correas de velcro para estabilizar al sujeto y la respuesta mecánica fue registrada mediante el uso de una celda de tensión (LC-500F, Kyowa, Japan) y mediante un dinamómetro (SS- 25, Biopac, USA). El EMG fue registrado utilizando electrodos circulares (Kendall-Arbo, Germany) (ancho de banda de 20-500 Hz, sensibilidad de 0.08 mV) ubicados en el vientre de los músculos vasto lateral (VL) y vasto medial (VM) con una distancia inter-electrodo de 20 mm. Antes de la colocación de los electrodos, la piel fue cuidadosamente rasurada y limpiada con alcohol. Se aplicó un filtro de 50 Hz a los datos del EMG para evitar la interferencia de las fuentes eléctricas. Las señales del EMG y de la tensión muscular fueron digitalizadas on-line (TEL 100, BIOPAC System, USA) con una frecuencia de muestreo de 1 kHz, y fueron guardados para los análisis. Las señales del EMG fueron filtradas con un filtro de paso alto a una frecuencia de corte de 0.2 Hz y luego fueron suavizadas con un filtro de paso bajo con una frecuencia de corte de 400 Hz (Acknowledge 7.3.3 Software, Biopac USA). Se realizaron los mismos análisis del EMG para 15 ciclos consecutivos durante el ejercicio de ciclismo. Todos los datos del EMG fueron normalizados dividiendo cada valor obtenido durante el ejercicio de ciclismo por el valor promedio del EMG obtenido durante la MVC y expresados como porcentajes.
Para los análisis de la HRV se utilizó la transformación de Fourier en un período de 5 min, lo cual se considera como una valoración de la influencia autonómica del corazón (Carter et al., 2003). Una unidad ECG100 (Biopac, USA) amplificó la señal proveniente de los electrodos, las cuales fueron digitalizadas on-line a una frecuencia de 1 kHz. Las frecuencias de 0.04-0.15 Hz (LF-ms2) y de 0.15-0.40 Hz (HF-ms2) fueron normalizadas respecto del espectro total de potencia y de muy bajas frecuencias (<0.04 Hz, VLF-ms2): LFn=LF/(potencia total - VLF) ×100 y HFn= HF/(potencia total-VLF) ×100. El índice LF/HF fue utilizado como un índice del grado de activación simpática (Carter et al., 2003). El análisis espectral de la señal respiratoria fue llevado a cabo mediante el mismo procedimiento. Estos espectros fueron utilizados para valorar la frecuencia respiratoria y para ubicar el componente respiratorio en el análisis de la potencia espectral de la variabilidad de los intervalos de la frecuencia respiratoria.
La temperatura rectal (Tre) fue medida utilizando un termistor (Yellow Springs, USA) conectado a un termómetro telemétrico (Yellow Springs, Model 46, USA). La FskBF fue estimada a partir de la velocidad microvascular de las células sanguíneas medida mediante una sonda Laser Doppler (LDF 100A, BIOPAC System, USA) ubicada en el lado dorsal del brazo derecho y asegurada para que mantuviera la misma posición y el contacto con la piel durante la prueba experimental. El Laser Doppler fue calibrado apropiadamente antes de su utilización.
La sonda laser Doppler fue conectada a un modulo de flujo LDF100A (BIOPAC, USA). Todas las señales del sensor fueron primero elaboradas por un dispositivo UIM100A de interface A/D (BIOPAC) que fue conectado a la unidad de adquisición de datos MP100A (BIOPAC) para su almacenamiento y posterior análisis. La concentración de Hb fue determinada mediante el método de cianmetahemoglobina (Dr. Lange Mini-Cuvette, LKM 143, Germany). Los valores del Hct fueron obtenidos a través de un lector (Hawksley, UK) luego de 4 min de centrifugado a 11500 rpm. Los coeficientes de variación en análisis repetidos fueron del 1.8% y del 1.7% para la Hb y el Hct respectivamente.
Cálculos
El SV (mL·latido-1) se calculó como el cociente entre el CO y la HR. La MAP se calculó a partir de la presión sanguínea sistólica (SP) y diastólica (DP): MAP=DP+1/3·(SP-DP). La resistencia periférica (TRP) se calculó como el cociente MAP·CO-1, y la diferencia arteriovenosa de oxígeno (a-vO2Diff) se calculó como el cociente VO2·CO-1. Los cambios en el volumen sanguíneo y en el volumen plasmático fueron calculados utilizando los datos de la Hb y del Hct (Dill and Costill, 1974).
Análisis Estadísticos
Se utilizó el análisis de varianza de dos vías para medidas repetidas en ambos factores (Statistica 5.0, USA). También se utilizó el análisis de covarianza para medidas repetidas para el ΔSV con el consumo de oxígeno como covarianza, para la HR con el Tre como covarianza y para la HRV con la Rf como covarianza. Se utilizó el test de Tukey para asignar las diferencias específicas en el análisis de varianza. Los datos se presentan como medias ± DE. Se utilizó el análisis de regresión múltiple por pasos utilizando como variable dependiente los cambios en la HR y la Tre, la actividad EMG, el cociente LF/HF y los cambios en el BV y en el PV como variables independientes. El mismo procedimiento se llevó a cabo con los datos del SV como variable dependiente y ∆HR, ∆Tre, ∆BV, ∆SBF, LF/HF y ΔHbT como variables independientes. El nivel de significancia fue establecido a 0.05.
RESULTADOS
La media de VO2 fue de 1.04 ± 0.09 y 2.1 ±0.12 L·min-1 para las condiciones de ejercicio con una y dos piernas, respectivamente; sin cambios en el tiempo. La intensidad para el ciclismo con dos piernas fue del 58% del VO2pico y la intensidad respectiva para la condición de ciclismo a una pierna fue calculada de acuerdo con lo propuesto por Kausen et al (1982) y fue de aproximadamente el 42% del VO2pico obtenido con el ejercicio a una pierna.
El CO fue mayor en la condición de ciclismo con dos piernas (p<0.01) con una tendencia a declinar (caída de 0.9L) hacia el final del ejercicio (p = 0.07, Figura 1a). En ambas condiciones el SV se redujo luego del minuto 35 de ejercicio, en la condición de ciclismo a dos piernas, y luego del minuto 50 en la condición de ciclismo a una pierna (Figura 1b). Luego del minuto 50 de ejercicio el ΔSV fue -20.8 ±0.8 ml·latido-1 en la condición de ejercicio a dos piernas y de -13.3 ± 1.3 mL·latido-1 en la condición de ciclismo a una pierna. La respuesta de la HR tuvo un curso inverso al de la SV, observándose diferencias significativas entre las condiciones (p<0.05) (Figura 1c). La translocación de la HR continuó siendo mayor en la condición de ciclismo a dos piernas cuando el porcentaje del VO2 fue utilizado como covarianza (p = 0.04).
La MAP fue de 101.0 ± 1.3 y de 100.4 ± 1.0 mmHg en las condiciones de ciclismo a una y dos piernas, respectivamente, sin diferencias significativas en el tiempo. La TPR fue mayor (p<0.05) al minuto 20 de la condición de ejercicio a una pierna (8.42 ± 0. 38 vs. 5.92 ± 0.27 mmHg·L·min-1) así como también al minuto 50 de ejercicio (9.2 ± 0.41 vs. 6.26 ± 0.31 mmHg·L·min-1). En contraste, la a-vO2diff fue mayor en la condición de ciclismo a dos piernas (8.7, 8.6 y 9.2 y 11.2, 11.8 y 12.4 mL de O2 por 100 mL de sangre para las condiciones de ciclismo a una y dos piernas a los minutos 20, 35 y 50, respectivamente, p<0.01).
Las diferencias entre las condiciones en la StO2 fueron del 7-10% (p<0.05) a partir del comienzo del ejercicio, mientras que las respectivas diferencias en la HbT no alcanzaron diferencias significativas. Ambas variables se incrementaron constantemente a partir del minuto 25 de ejercicio (Figura 2).
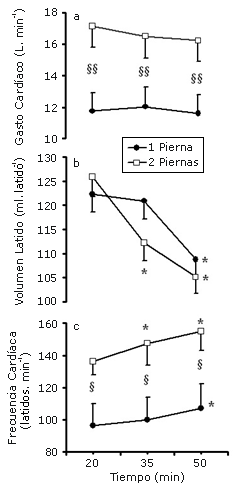
Figura 1. Gasto cardíaco, volumen latido y frecuencia cardíaca durante 55 min de ciclismo con dos y una pierna a un consumo de oxigeno similar por pierna (n = 12). § y §§ denotan diferencias significativas entre las condiciones; p<0.05 y p<0.01, respectivamente. *Denota diferencias a partir del minuto 20 de ejercicio, p<0.05.
La actividad del EMG integrado fue mayor tanto para el VM como para el VL en la condición de ciclismo a dos piernas (Figura 3) debido a la mayor carga externa aplicada. El cociente LF/HF se incrementó desde el reposo en ambas condiciones observándose mayores valores en la condición de ciclismo a dos piernas (2.98 ± 0.45, 5.23 ± 0.54 y 5.84 ± 0.58 para la condición de ciclismo a una pierna vs 3.40 ± 0. 39, 8.04 ± 0.49 y 8.22 ± 0.86 para la condición de ciclismo a dos piernas en reposo y entre los minutos 9-14 y 35-40 de ejercicio, respectivamente).
El porcentaje de reducción en el volumen plasmático y en el volumen sanguíneo a partir de los valores pre ejercicio fue significativo en la condición de ciclismo a dos piernas, mientras que las diferencias entre las condiciones se observaron únicamente en la primera variable (Tabla 1).
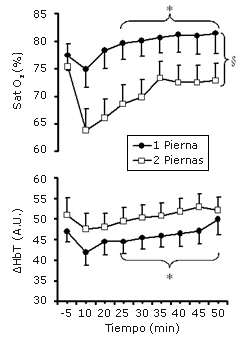
Figura 2. Saturación muscular de oxígeno (StO2) y cambios en el volumen sanguíneo (ΔHbT), durante 55 min de ciclismo con dos y una pierna a un consumo de oxígeno similar por pierna (n = 12). § denota diferencias significativas entre las condiciones; p<0.05. *Denota diferencias a partir del minuto 10 de ejercicio, p<0.05.
En ambas condiciones, la temperatura rectal medida al final del ejercicio fue mayor que la temperatura rectal medida en reposo (p<0.05). Al minuto 35 de ejercicio y de allí en adelante, la Tre fue mayor en la condición de ciclismo a dos piernas (Tabla 1). El FskBF se incrementó a lo largo del ejercicio en ambas condiciones, pero no se hallaron diferencias entre las condiciones. La RPE general y local fue diferente entre las condiciones luego del minuto 30 de ejercicio. Los valores medios de la RPE local y general para las condiciones de ciclismo a una y dos piernas fueron de 12 ± 0.5 vs 13.7 ± 0.3 y 10.5 ± 0.4 vs 11.8 ± 0.4, respectivamente.
En la condición de ciclismo a dos piernas, el análisis de regresión múltiple reveló que el ΔSV se vio influenciado (r2 = 0.99, p < 0.001) por el ΔHR (p<0.01), el ΔBV (p = 0.04) y el ΔHbT (p = 0.02), mientras que el ΔHR (r2 = 0.88, p < 0.01) fue influenciado por el ∆Tre (p < 0.01) y por el ∆ de actividad EMG (p < 0.01). En la condición de ciclismo a una pierna, el ∆SV fue explicado (r2 = 0.99, p < 0.001) por el ∆HR (p < 0. 01), mientras que ninguno de los factores examinados explicó el ∆HR.
DISCUSION
El principal hallazgo de este estudio fue que el ciclismo con una mayor masa muscular incrementa el CVdrift, lo cual fue indicado por un mayor incremento en la HR a través del protocolo, y la mayor caída del SV al final del ejercicio con dos piernas y en comparación con el ejercicio a una pierna. El mayor incremento en la HR estuvo acompañado por la mayor respuesta simpática y se relacionó principalmente con la actividad del EMG y el RPE. Estos factores indican una mayor activación central en el ejercicio con dos piernas, bajo las presentes condiciones experimentales.
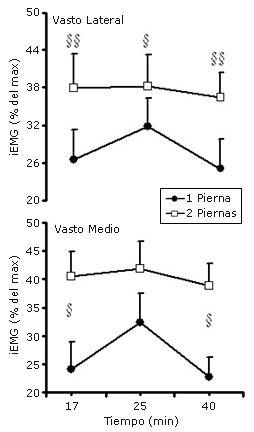
Figura 3. EMG integrado (iEMG) en el vasto lateral y en el vasto medial a los 17, 25 y 40 minutos de ejercicio con dos y una pierna a un consumo de oxígeno similar por pierna (n = 12). § y §§ denotan diferencias significativas entre las condiciones; p<0.05 y p<0.01, respectivamente. *Denota diferencias a partir del minuto 17 de ejercicio, p<0.05.
La activación central pudo haber afectado la respuesta simpática, derivando en un incremento de la frecuencia cardíaca y de la vasoconstricción en los lechos vasculares musculares (Pawelczyk et al., 1997; Schibye et al., 1981). El mayor RPE observado en la condición de ciclismo a dos piernas, luego del minuto 30 de ejercicio respalda la hipótesis de la mayor activación central (Norton et al., 1999). Además, la actividad del EMG fue mayor en la condición de ciclismo a dos piernas (Figura 2), lo cual significa un mayor reclutamiento de unidades motoras motivado por una mayor activación central (Franke et al., 2000; Nobrega et al., 1994; Schibye et al., 1981). La asociación de la activación muscular, registrada como actividad EMG, y la HR fue confirmada por el análisis de regresión múltiple por pasos (n = 48), utilizando los cambios en la HR como variable dependiente, y los cambios en la Tre, el iEMG, la BV y en el RPE como variables independientes. Los análisis de regresión revelaron que, además de la Tre, la actividad del EMG también explicó casi el 50% de la variabilidad en la HR durante el ciclismo a dos piernas (ponderación beta = 0.49 para el EMG, R2 = 0.88, p < 0.01).
La mayor activación simpática también fue indicada por el mayor cociente LF/HF en el ejercicio a dos piernas. Este incremento en el tono simpático observada durante la condición de ejercicio a dos piernas fue probablemente necesario para mantener la perfusión y la presión y para evitar la hipotensión (Calbet et al., 2004). En efecto, al igual que en el presente estudio, en un estudio previo no se observaron diferencias en la MAP entre las condiciones de ciclismo a una y dos piernas (Savard et al., 1989) a pesar de la menor TPR y la mayor concentración de catecolaminas plasmáticas durante el ejercicio con mayor masa muscular activa (Jensen-Urstad et al., 1994). Por ejemplo, Savard et al (1989) observaron que durante la realización de ejercicios dinámicos, el excedente noradrenalina (una medida indirecta del tono simpático) es proporcional a la fracción de masa muscular total reclutada sin cambios en la MAP y no está asociado con los cambios en el flujo sanguíneo hacia las extremidades. El mayor VO2 durante el ejercicio a dos piernas, en comparación con el ciclismo a una pierna, está asociado con la mayor reducción en los niveles de StO2 en los músculos activos (Figura 2a). El incremento en la a-vO2Diff observado en la condición de ciclismo a dos piernas implica un adecuado suministro de oxígeno hacia los músculos, ya que no se observaron diferencias significativas en el flujo sanguíneo muscular entre las condiciones, lo cual fue indicado por valores similares en la HbT (Figura 2b). Estas observaciones concuerdan con los estudios que han reportado una mayor desaturación (Jensen-Urstad et al., 1994), una mayor vO2Diff (Jensen-Urstad et al., 1994; Lewis et al., 1983) y un similar flujo sanguíneo hacia las piernas (Jensen-Urstad et al., 1994) con una mayor masa muscular involucrada en el ejercicio. Norton et al (1999) reportaron que durante el ciclismo prolongado, el incremento en el input aferente desde los músculos hacia el área cardiovascular es también es importante para el desarrollo del CVdrift, y en este sentido (Kjaer et al., 1999; Romer et al., 2007), los mayores niveles de desaturación observados en el presente estudio en la condición de ciclismo a dos piernas, podrían exagerar el incremento en la frecuencia cardíaca y, por lo tanto, en la reducción del SV. La relación significativa hallada entre el incremento de la frecuencia cardíaca y el ∆StO2 (r = 0.51, p<0. 05) respalda esta hipótesis.
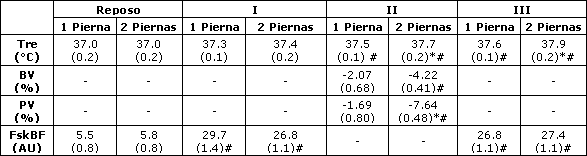
Tabla 1. Cambios en la temperatura rectal (Tre), volumen sanguíneo (BV) y volumen plasmático (PV) a partir del reposo y flujo sanguíneo en el antebrazo (FskBF) a los 20 (I), 35 (II) y 50 (III) minutos de ejercicio para la Tre, a los 35 min (II) para el BV y el PV, y a los 15 (I) y 45 (III) de ejercicio para el FskBF. Los valores son medias (± EE). *Denota diferencias entre las condiciones, p<0.05. # Diferente del reposo, p<0.05- AU: unidades arbitrarias.
La hipertermia es otro posible factor que puede afectar el CVdrift. Más específicamente, la mayor Tre observada durante el ciclismo a dos piernas (0.3º) podría haber acelerado el ritmo cardíaco (Rubin, 1987) posiblemente induciendo una mayor respuesta simpática (Gonzalez-Alonso et al., 1999), tal como lo indicó el cociente LF/HF en la condición de ejercicio a dos piernas. Sin embargo, la Tre no puede explicar completamente el diferente curso de la HR entre las condiciones debido a: (a) la HR fue mayor en el ejercicio a dos piernas a la misma Tre y (b) las diferencias estadísticas en la HR entre las dos condiciones se mantuvo cuando la Tre fue utilizada como covarianza en el análisis de varianza de la HR.
Similarmente, las diferencias observadas entre las condiciones experimentales respecto de la HR no son atribuibles a la mayor intensidad de ejercicio (~15%) en el ejercicio a dos piernas, debido a que las diferencias entre las condiciones se mantuvieron intactas cuando se llevó a cabo el análisis ANCOVA. Estos hechos sugieren que existen otros factores, además de la Tre y de la intensidad del ejercicio, que pueden desempeñar un rol en las diferencias observadas en incremento en la frecuencia cardíaca entre las dos pruebas.
En ambas condiciones experimentales se observó el CVdrift, pero fue más pronunciado durante el ciclismo a dos piernas (mayor incremento en ΔHR, 13 latidos·min-1). El incremento resultante en la frecuencia cardíaca podría reducir el tiempo de llenado cardíaco y explicar la caída del SV a lo largo del tiempo (mayor caída en el ΔSV de 7.5 ml·latido-1) (Franke et al., 2000). La alta correlación (n = 48; 12 sujetos x 4 puntos de medición) hallada entre los cambios en la HR y en el SV en ambas condiciones (r = -0.80 y -0.76 para el ejercicio a 2 y 1 pierna, p<0.01) concuerda con aquellos estudios que han utilizado un protocolo de ejercicio prolongado de ciclismo y carrera (Franke et al., 2000; Nassis and Geladas, 2002) y respalda el rol el incremento en la frecuencia cardíaca en el desarrollo de una translocación en el SV bajo circunstancias similares a las del presente estudio.
La mayor reducción del BV durante el ciclismo a dos piernas también pudo inducir un incremento en la frecuencia cardíaca a través de la activación de los barorreceptores (Norton et al., 1999) y así explicar la declinación en el SV (). Como respaldo al rol de la deshidratación sobre la reducción del SV, el análisis de regresión múltiple por pasos (n = 48, R2 = 0.97, p<0.01) mostró que, en la condición de ejercicio a dos piernas, además del ∆HR (ponderación beta 0.40, p = 0.03), y la HbT (ponderación beta 0.37, p = 0.02), la variabilidad en el ∆SV fue explicado solo por el ∆BV (ponderación beta de 0.2, p = 0.04), sugiriendo un rol de en la translocación de la HR.
Las posibles limitaciones del presente estudio incluyen las mediciones de la actividad del EMG y del RPE, los cuales no son índices directos de la activación central. Sin embargo, el comando central ha sido evaluado con índices similares a los utilizados en el presente estudio (Franke et al., 2000; Norton et al., 1999; Schibye et al., 1981). Además, los cambios en el volumen sanguíneo muscular fueron estimados con mediante NIRS a través de la HbT, lo cual sin embargo, es considerado como un índice del flujo sanguíneo muscular (Meyers et al., 2005).
CONCLUSIONES
En conclusión, durante los 55 min de ciclismo, el CVdrift fue mayor en la condición de ejercicio con dos piernas que en la condición de ejercicio a una pierna al mismo consumo de oxígeno por pierna. Esto implica que conjuntamente con el estatus térmico y la reducción en el volumen sanguíneo, el comando central desempeña un rol en la regulación cardiovascular durante la realización de ejercicios en estado estable con una gran masa muscular involucrada.
Puntos Clave
La magnitud de la masa muscular activa desempeña un rol crítico en el desarrollo de la translocación cardiovascular, cuando el consumo de oxígeno por pierna es el mismo.
Además del estatus térmico y de la reducción en el volumen sanguíneo, el comando central desempeña un rol en la regulación cardiovascular durante la realización de ejercicios en estado estable que involucren una gran masa muscular.
Agradecimientos
El proyecto fue co-financiado por una subvención de Educación Operacional de la European Social Fund y National Resources. Declaramos que los experimentos cumplen con las leyes griegas y todos los procedimientos tuvieron la aprobación del Comité de Ética local.
Referencias
1. Calbet, J., Jensen-Urstad, M., Van Hall, G., Holmberg, H., Rosdahl, H. and Saltin, B (2004). muscular vascular conductances during whole body upright exercise in humans. Journal of Physiology 5558, 319-331
2. Carter, J., Banister, E. and Blaber, A (2003). Effect of endurance exercise on autonomic control of heart rate. Sports Medicine 33, 33-46
3. Coyle, E. and Gonzalez-Alonso J (2001). Cardiovascular drift during prolonged exercise: new perspectives. Exercise and Sports Science Reviews 29, 88-92
4. DaSilva, G., Manshawi, A., Heigenhouser, G. and Jones, N (1985). Measurement of mixed venous carbon dioxide pressure by rebreathing during exercise. Respiratory Physiology 59, 379-392
5. Defares, J (1958). Determination of PvCO2 from the exponential CO2 rise during rebreathing. Journal of Applied Physiology 13, 159-64
6. Dill, D. and Costill, D (1974). Calculation of percentage changes in volumes of blood, plasma and red cells in dehydration. Journal of Applied Physiology 37, 247-248
7. Franke, W., Boettger, C., and McLean, S (2000). Effects of varying central command and muscle mass on the cardiovascular responses to isometric exercise. Clinical Physiology 20, 385-387
8. Fritzsche, R., Switzer, T., Hodgkinson, B. and Coyle, E (1999). Stroke volume decline during prolonged exercise is influenced by the increase in heart rate. Journal of Applied Physiology 86, 799-805
9. Ganio, M., Wingo, J., Carroll, C., Thomas, M. and Kureton, K (2006). Fluid ingestion attenuates the decline in VO2peak associated with cardiovascular drift. Medicine and Science in Sports and Exercise 38, 901-909
10. Gonzalez-Alonso, J., Mora-Rodriguez, R. and Coyle, E (2000). Stroke volume during exercise: interaction of environment and hydration. American Journal of Physiology 2278, 321-330
11. Gonzalez-Alonso, J., Calbet, J.A. and Nielsen, B (1998). Muscle blood flow is reduced with dehydration during prolonged exercise in humans. Journal of Physiology 513, 895-905
12. Gonzalez-Alonso, J., Teller, C., Andersen, S., Jensen, F., Hyldig, T. and Nielsen, B (1999). Influence of body temperature on the development of fatigue during prolonged exercise in the heat. Journal of Applied Physiology 86, 1032-1039
13. Jensen-Urstad, M., Svedenhag, J. and Sahlin, K (1994). Effect of muscle mass on lactate formation during exercise in humans. European Journal of Applied Physiology 669, 189-195
14. Kjaer, M., Hanel, B., Worm, L., Perko, G., Lewis, S.F., Sahlin, K., Galbo, H. and Secher, N.H (1999). Cardiovascular and neuroendocrine responses to exercise in hypoxia during impaired neural feedback from muscle. American Journal of Physiology 2277, 76-85
15. Klausen, K., Secher, N., Clausen, J., Hartling, O. and Trap-Jensen, J (1982). Central and regional circulatory adaptations to one-leg training. Journal of Applied Physiology 552, 976-983
16. Kounalakis S, Keramidas M, Nassis G, Geladas N (2008). The role of muscle pump in the development of cardiovascular drift. European Journal of Applied Physiology 103, 99-107
17. Lewis, S., Snell, P., Taylor, W., Hamra, M., Graham, R., Pettinger, W. and Blomqvist, G (1985). Role of muscle mass and mode of contraction in circulatory responses to exercise. Journal of Applied Physiology 558, 146-151
18. Lewis, S., Taylor, W., Graham, R., Pettinger, W., Schutte, E. and Blomqvist, G (1983). Cardiovascular responses to exercise as functions of absolute and relative work load. Journal of Applied Physiology 554, 1314-1323
19. Meyers, D., Anderson, L., Seifert, R., Cooper, C., Beilman, G. and Mowlem, J (2005). Noninvasive method for measuring local hemoglobin oxygen saturation in tissue using wide gap second derivative near-infrared spectroscopy. Journal of Biomedical Optics 10, 3401-3407
20. Montain, J. and Coyle, F (1992). Influence of graded dehydration on hyperthermia and cardiovascular drift during exercise. Journal of Applied Physiology 73, 1340-1350
21. Nassis, G. and Geladas, N (2002). Cardiac output decline in prolonged dynamic exercise is affected by the exercise mode. Pflügers Archiv European Journal of Physiology 4445, 398-404
22. Nobrega, A., Williamson, W., Friedman, D., Araujo, C. and Mitchell, J (1994). Cardiovascular responses to active and passive cycling movements. Medicine and Science in Sports and Exercise 226, 709-714
23. Norton, K., Gallagher, K., Smith, S., Querry, R., Welch-O'Connor, R. and Raven, P (1999). Carotid baroreflex function during prolonged exercise. Journal of Applied Physiology 887, 339-347
24. Pawelczyk, J., Pawelczyk, R., Warberg, J., Mitchell, J. and Secher, N (1997). Cardiovascular and catecholamine responses to static exercise in partially curarized humans. Acta Physiologica Scandinavica 1160, 23-28
25. Romer, L.M., Haverkamp, H.C., Amann, M., Lovering, A.T., Pegelow, D.F. and Dempsey, J.A (2007). Effect of acute severe hypoxia on peripheral fatigue and endurance capacity in healthy humans. American Journal of Physiology 2292, 598-606
26. Rowell, L., Murray, J., Brengelmann, G. and Kraning, K (1969). Human cardiovascular adjustments to rapid changes in skin temperature during exercise. Circulation Research 224, 711-724
27. Rubin, S (1987). Core temperature regulation of heart rate during exercise in humans. Journal of Applied Physiology 662, 1997-2002
28. Savard, G., Richter, E., Strange, S., Kiens, B., Christensen, N., Saltin, B., et al (1989). Norepinephrine spillover from skeletal muscle during exercise in humans: the role of muscle mass. American Journal of Physiology 2257, 1812-1818
29. Schibye, B., Mitchell, J., Payne, F. and Saltin, B (1981). Blood pressure and heart rate response to static exercise in relation to electromyographic activity and force development. Acta Physiologica Scandinavica 1113, 61-66
30. Warburton, D., Haykowsky, M., Quinney, H., Humen, D. and Teo, K (1999). Reliability and validity of measures of cardiac output during incremental to maximal aerobic exercise. Part I: Conventional techniques. Sports Medicine 27, 23-41
31. Watanabe, S., Ishii, C., Takeyasu, N., et al (2005). Assessing muscle vasodilation using near infrared spectroscopy in cardiac patients. Circulation Journal 69, 802-814
32. Williamson, J., Nobrega, A., McColl, R., Mathews, D., Winchester, P., Friberg, L. and Mitchell, J (1997). Activation of the insular cortex during dynamic exercise in humans. Journal of Physiology 5503, 277-283