
¿Está la Recuperación Regulada por Factores Centrales o por Factores Periféricos? Rol del Cerebro en la Recuperación Luego de Ejercicios de Sprint Intermitente
Is Recovery Driven by Central or Peripheral Factors? A Role for the Brain In Recovery Following Intermittent-Sprint Exercise
Geoffrey M. Minett1,2 y Rob Duffield3
1Brisbane, QLD, Australia
2Institute of Health and Biomedical Innovation, Queensland University of Technology, Kelvin Grove, Brisbane, QLD, Australia
3Sport and Exercise Discipline Group, UTS: Health, University of Technology Sydney, Lindfield, Sydney, NSW, Australia
Artículo publicado en el journal Revista de Entrenamiento Deportivo, Volumen 30, Número 1 del año 2016.
Publicado 25 de abril de 2016
Resumen
Palabras clave: Deportes de equipo, futbol americano, fútbol, fatiga, sistema nervioso central, función contráctil, daño muscular
Abstract
Keywords: team-sports, football, soccer, fatigue, central nervous system, contractile function, muscle damage
INTRODUCCIÓN
Los atletas de deportes de equipo experimentan diferentes niveles de fatiga transitoria (aguda) y residual (continua) durante los períodos de entrenamiento intenso y de competición (Borresen y Lambert, 2009). Caracterizados por series repetidas de ejercicio de sprint intermitente, los deportes de equipo realizados en el campo plantean grandes demandas fisiológicas (Spencer et al, 2005a), neuromusculares (Rampinini et al, 2011; Duffield et al, 2012) y perceptuales (Impellizzeri et al, 2004). Estas tensiones relativas se agravan cuando las series sucesivas comprometen la tasa de recuperación innata, y como consecuencia pueden limitar el rendimiento resultante (Bishop et al, 2008). Es importante acelerar artificialmente la evolución natural de los procesos de regeneración por medio de estrategias de recuperación para la preparación de las series de entrenamiento o de competencia subsiguientes (Barnett, 2006). Sin embargo, la comprensión de los mecanismos que regulan la recuperación es fundamental para determinar los efectos probablemente beneficiosos de estas intervenciones; especialmente dado que el marco teórico de las investigaciones sobre la recuperación del rendimiento en deportes tiene un alcance limitado y está enfocado específicamente en la periferia (Nédélec et al, 2013).
Se han defendido varias modalidades de recuperación para tratar el daño muscular inducido por el ejercicio (EIMD) y el dolor muscular de aparición tardía asociado (DOMS), pero su eficacia ha sido el blanco de considerables críticas (Barnett, 2006,; Bishop et al, 2008; MacRae et al, 2011; Nédélec et al, 2013) y de revisión sistemática (Bleakley et al, 2012; Leeder et al, 2012; Bieuzen et al, 2013; Poppendieck et al, 2013). Si bien tales estrategias de recuperación podrían ser ampliamente diferenciadas como fisiológicas (ej., crioterapia, hidroterapia, masaje, compresión, sueño), farmacológicas (por ejemplo, antinflamatorios no esteroideos) o nutricionales (por ejemplo, suplementos dietéticos), todas apuntan a limitar las perturbaciones continuas post ejercicio y eventos inflamatorios s dentro de las células de los músculos que realizan los ejercicios. Este enfoque periférico enfatiza la importancia de un retorno acelerado a la integridad estructural y a la capacidad funcional desde abajo de la unión neuromuscular. Sin embargo, hay evidencia creciente que demuestra que existen tasas desasociadas de recuperación física y fisiológica luego de la realización de ejercicios de sprint intermitente, particularmente el retorno de fuerza neuromuscular a pesar del aumento continuo en los marcadores sanguíneos de EIMD (Pointon et al, 2012; Minett et al, 2013). En este sentido, es necesario tener una mayor consideración sobre el papel del sistema nervioso central (CNS) en la recuperación del rendimiento físico.
Teóricamente, la tasa de recuperación post ejercicio está relacionada con la magnitud de la carga impuesta sobre los sistemas fisiológico y neuromuscular por la serie de ejercicios (Nédélec et al, 2012). Por lo tanto, se asume que las bases fisiológicas de la recuperación dependen de la recuperación oreversión de estas tensiones en la periferia. Muy por el contrario, la regulación del rendimiento durante el ejercicio ha sido interpretada cada vez más como un fenómeno integrador, multifacético (Rnicker et al, 2011; Noakes, 2012) y en este proceso el cerebro tiene un rol central (Gandevia, 2001). Mientras que se plantean algunas discusiones acerca de si la regulación del rendimiento en el ejercicio es un evento consciente (Marcora, 2008) o un evento anticipador (Marino, 2004)-derivado de orígenes aferentes o eferentes (St Clair Gibson y Noakes, 2004); todo indica que los cambios en la regulación del CNS y el reclutamiento de unidades motoras están asociados con la fatiga y con la reducción del rendimiento de ejercicios de sprint intermitente (Billaut, 2011; Girard et al, 2011). Conceptualmente, si el cerebro tiene un lugar central en el proceso de la disminución del rendimiento (es decir, fatiga), lógicamente también desempeñaría algún papel en la recuperación post ejercicio (De Pauw et al, 2013). En tal sentido, esta revisión busca discutir la recuperación del CNS junto con los factores periféricos después de los ejercicios de sprint intermitente propios de deportes de equipo.
Fatiga y Necesidad de Recuperación después de Ejercicios de Sprints Intermitentes
Resaltando el punto anterior, es importante identificar primero la necesidad de recuperación antes de discutir los potenciales conductores desde el punto de vista central (nervios/cerebro) y periférico (muscular). Definida clásicamente como una reducción en la capacidad de generación de fuerza muscular inducida por el ejercicio, la fatiga puede ser atribuida a una falla contráctil periférica, a un output sub-óptimo del cortex motor (fatiga supraspinal) y/o a imputs aferentes alterados (fatiga espinal) que inervan la musculatura activa (Gandevia, 2001).
Independientemente, se han utilizado simulaciones de laboratorio de ejercicios de sprint intermitente para examinar los mecanismos de fatiga relacionados con los deportes de equipo (Pointon et al, 2012; Minett et al, 2013) y han informado una definición operacional de fatiga que estaría asociada con una disminución en la velocidad máxima de sprint o producción de potencia entre los esfuerzos múltiples (Bishop, 2012). Notablemente, sin embargo, las observaciones de fatiga, o por lo menos de reducciones en la intensidad del ejercicio que se realizan a campo son más abstractas. Los análisis de tiempo-movimiento resaltan la adopción de estrategias de ritmo en los deportes de equipo y sugieren tanto reducciones "transitorias" en la tasa de trabajo después esfuerzos intensos como disminuciones progresivas en las distancias/velocidades recorridas a medida que los partidos avanzan (Mohr et al, 2003; Duffield et al, 2009; Coutts et al, 2010; Akenhead et al, 2013).
Si bien las perturbaciones periféricas en el músculo esquelético, el funcionamiento cardiovascular y la tensión metabólica tradicionalmente están vinculados con la fatiga transitoria (Bangsbo et al, 2006), es cada vez mas frecuente la consideración de la participación del CNS en las estrategias de ritmo alteradas (Roelands et al, 2013). De hecho, los atletas de deportes de equipo pueden alterar brevemente sus patrones de movimiento en respuesta a contextos de competencias y a demandas fisiológicas acumuladas, de modo de preservar los esfuerzos de alta intensidad a medida que el juego progresa (Bradley y Noakes, 2013). Alternativamente, los conceptos de fatiga residual siguen estando predominantemente dentro del dominio de los mecanismos regulados periféricamente, como el flujo sanguíneo, la recuperación del glucógeno muscular y la eliminación de desechos metabólicos (Bangsbo et al, 2006). Como se discutió en una reciente revisión de recuperación en fútbol, Nédélec et al. (2012) resaltaron las disminuciones luego del partido que se observan en el tiempo de sprint, en la altura de salto y en la fuerza de contracción voluntaria máxima (MVC) que llegó a 36% a las 96 horas. La recuperación incompleta antes de las series de ejercicio siguientes probablemente sea perjudicial para el sprint intermitente- (Spencer et al, 2005b) y para el desempeño en las habilidad deporte específicas (Minett et al, 2012a), aunque la evolución en el tiempo de los cambios en la recuperación física y sus orígenes centrales y periféricos respectivos no están todavía claros (Bishop, 2008).
Los cambios físicos y bioquímicos observados durante el ejercicio de sprint intermitente han sido interpretados tradicionalmente en términos de capacidad metabólica (Glaister, 2005). De hecho, las menores concentraciones de fosfocreatina (Dawson et al, 1997), la menor regeneración de ATP por la vía glucolítica (Gaitanos et al, 1993) y la acumulación creciente de H+ (Bishop et al, 2003) se han asociado con un menor rendimiento en sprint intermitente. Si bien, también se han observado disminuciones en la excitabilidad muscular luego de ejercicios de sprint intermitente (Obispo, 2012), las perturbaciones metabólicas se recuperan rápidamente en minutos (Glaister, 2005). En concordancia con esto, cuando se experimentan reducciones sostenidas en el rendimiento (>2h) durante entrenamiento o sesiones de competencias consecutivas, se ha sugerido que el factor limitante de la producción de potencia pico en ejercicios de sprint intermitente es el daño muscular inducido por el ejercicio (EIMD) (Twist y Eston, 2005). Esta hipótesis es lógica, aunque limitada por una aparente desconexión entre la expresión de marcadores de daño muscular y la recuperación de la función neuromuscular luego de ejercicios de sprint intermitentes (Pointon et al, 2012; Minett et al, 2013). Además, dado el efecto del dolor muscular de aparición tardía (DOMS) subsiguiente en la alteración del reclutamiento neuromuscular o en los patrones de activación (Cheung et al, 2003), parecería que el foco de la recuperación no debería ser solamente el periférico (De Pauw et al, 2013).
El mejor indicador de la recuperación post ejercicio es la habilidad del músculo de producir fuerza, i.e., los resultados de rendimiento. La secuenciación y la intensidad óptimas de activación del músculo y los patrones de reclutamiento son importantes para el desarrollo de producción de potencia máxima (Ross et al, 2001), y por lo tanto se consideran importantes factores de regulación que contribuyen con el rendimiento sostenido en sprint intermitente (Girard et al, 2011). Las reducciones en las señales EMG luego de un ejercicio simulado de fútbol parecen coincidir con una menor contracción voluntaria máxima (MVC) y sugieren una inhibición del impulso nervioso que sigue siendo evidente 24 hs post ejercicio (Rampinini et al, 2011). Si bien recomendamos tener cierta cautela cuando se interpreta directamente la EMG superficial durante la actividad dinámica (Farina, 2006), se han observado disminuciones en la activación voluntaria (VA) determinadas mediante la técnica de contracción interpolada (interpolated twitch) después de ejercicios de sprint repetidos (Racinais et al, 2007; Perrey et al, 2010). Notablemente, los resultados obtenidos a campo son mixtos, y en algunos casos de deportes basados en sprints intermitentes la activación voluntaria (VA) se ve afectada luego de los partidos (Girard et al, 2008, 2010), pero no en todos (Duffield et al, 2012). Estas diferencias probablemente se explican tanto por el tipo de deporte es decir, la carga específica de la tarea realizada, así como por las diferencias metodológicas. Específicamente, los cambios en la función neuromuscular que se observan luego del ejercicio son afectados por la duración de la medición (Girard et al, 2013) y la proximidad a la finalización de la serie de ejercicios (Froyd et al, 2013). En concordancia con esto, es concebible que la fatiga neuromuscular pueda ser subestimada en las mediciones realizadas en el campo (Froyd et al, 2013), particularmente en aquellos casos en que es difícil el acceso inmediato a los participantes luego del juego.
En síntesis, las disminuciones relacionadas al rendimiento en sprint intermitente parecen ser un fenómeno multifacético proveniente de vías neuromusculares, contráctiles y metabólicas (Girard et al, 2011; Bishop, 2012). Si bien la inclusión de mediciones neurológicas avanzadas ha aportado una valiosa visión sobre las contribuciónes de la fatiga central y periférica producidas inmediatamente después del ejercicio de sprint intermitente (Girard et al, 2013), es necesario comprender completamente la cinética de la recuperación de estos mecanismos. Además, considerando la especificidad de las respuestas fisiológicas frente una cierta tarea de ejercicio, es necesario obtener una mejor comprensión de la cinética de la recuperación luego de los ejercicios de sprint intermitente, particularmente en el campo. El papel del cerebro, y por lo tanto del SNC, ha sido reconocido ampliamente como integral en la regulación de los ejercicios realizados al propio ritmo (Tucker, 2009; Roelands et al, 2013), aunque no se ha considerado adecuadamente cual es el rol que el cerebro desempeña en la recuperación
.
Recuperación Periférica después de Ejercicios de Sprints Intermitentes
Los cambios en el rendimiento físico luego de ejercicios de sprint intermitente prolongados son representados de la mejor manera por las reducciones en la velocidad máxima, la distancia total recorrida y los esfuerzos de carrera de alta intensidad (Mohr et al, 2003; Coutts et al, 2010). Estos cambios en los marcadores de rendimiento a campo coinciden temporalmente con una disminución en la función del musculo esquelético, deducida de las observaciones de menor rendimiento en MVC y CMJ (Cormack et al, 2008; McLellan et al, 2011; Duffield et al, 2012). Frecuentemente se propone que las disminuciones en la función del músculo esquelético después de los ejercicios de sprint intermitente son provocadas por un rango de factores inducidos periféricamente, entre los que se incluyen: el agotamiento del glucógeno intramuscular; aumento en las concentraciones de metabolitos en el musculo y en la sangre; alteraciones en las funciones de las bombas de Ca++ o Na+-K+; aumento en el daño del músculo esquelético; aumentos excesivos en las temperaturas endógenas del músculo y del core; y reducción en la función circulatoria por un menor volumen de sangre e hipo hidratación (Duffield y Coutts, 2011; Bishop, 2012; Nédélec et al, 2012). Como señalan Coutts et al. (En prensa), ya sea en conjunto o de manera aislada, muchas de estas alteraciones inducidas por el ejercicio en el estado fisiológico reducen la producción de fuerza post ejercicio y se postula que afectan la tasa de recuperación del rendimiento subsiguiente. Se supone que aliviar estas perturbaciones fisiológicas facilitaría el retorno de la fuerza contráctil neuromuscular, y también el rendimiento en sprint intermitente.
Es marcado el nivel de estrés glucolítico y oxidativo que se produce durante el ejercicio de sprint intermitente, con disminuciones considerables en la disponibilidad de sustratos de energía, lo que se sabe que produce una disminución en la fuerza del músculo esquelético (Girard et al, 2011). En concordancia con esto, también se supone que las limitaciones en el suministro de energía, ya sea de fosfato inorgánico o de concentraciones de glucógeno muscular almacenadas, impiden la recuperación después de la actividad en deportes de equipo (Nédélec et al, 2012). Aunque la concentración de fosfocreatina (disminuida entre 35-55% del valor de reposo luego de un esfuerzo de sprint de 6 s) está asociada con la producción de potencia (Bishop, 2012), los períodos de recuperación breves y variables experimentados en los deportes de equipo no siempre permiten que se realice la resíntesis. Independientemente, dada la evolución de la regeneración de fosfocreatina, no se considera un factor en la recuperación de competencias o series de entrenamiento que están separadas por horas o días. De manera contraria, Rrustrup et al. (2006) informaron disminuciones en el glucógeno intramuscular de 42±6% en jugadores de fútbol, con agotamiento o casi agotamiento de las reservas de glucógeno en ~55% de las fibras de tipo I y ~25-45% de las fibras tipo II, lo que permite explicar las disminuciones agudas en la velocidad de sprint luego de un partido. Notablemente la resíntesis de glucógeno muscular, después de una actividad de deporte de equipo es lenta y puede permanecer atenuada durante 2-3 días (Nédélec et al, 2012). Tales resultados resaltan la importancia de la nutrición en la recuperación post ejercicio (Burke et al, 2006); por lo tanto es importante destacar que las reservas de glucógeno muscular permanecen afectadad 24 horas después de un partido del fútbol, independientemente de la ingesta de carbohidratos y deben ser reconocidas como un factor en la supresión de fuerza sostenida luego de un partido (Bangsbo et al, 2006; Krustrup et al, 2011).
Además del compromiso en la disponibilidad/provisión de sustratos de energía experimentado luego de una actividad de sprint intermitente, las contracciones excéntricas repetidas de alta potencia producidas durante el ejercicio de deportes de equipo perturban la función del músculo esquelético y producen marcadas respuestas de daño muscular (McLellan et al, 2011). Las rupturas mecánicas de las fibras musculares dependen de las tareas, aunque probablemente están relacionadas con la cantidad de aceleración, desaceleración, cambio direccional y contacto entre jugadores que se producen (es decir, derribos o colisiones) (McLellan et al, 2011; Duffield et al, 2012). Notablemente, el daño muscular inducido por el ejercicio (EIMD) se manifiesta en una menor producción de fuerza voluntaria que se asocia con una expresión elevada de proteínas intracelulares (por ejemplo, creatina quinasa y proteína C reactiva), inflamación, rango restringido de movimiento y dolor muscular (Cheung et al, 2003). Aunque generalmente se acepta que bajar los perfiles sanguíneos de daño muscular puede acelerar la recuperación deportiva, los mecanismos que explican el retorno de la función del músculo esquelético son algo ambiguos (Howatson y Van Someren, 2008). Tal como se observa en la Figura 1, la evolución en el tiempo de los marcadores asociados con la recuperación de la fuerza voluntaria luego de ejercicios de sprint intermitente, no refleja las mediciones obtenidas en sangre. Es interesante destacar que los marcadores de EIMD no están estrechamente asociados con el dolor muscular (Nosaka et al, 2002; Prasartwuth et al, 2005), aunque se ha informado que la recuperación perceptual está relacionada con la recuperación de la velocidad máxima de sprint (Cook y Beaven, 2013). Si bien esto plantea interrogantes en términos de fundamentos fisiológicos sobre el dolor muscular, las relaciones débiles entre EIMD y el rendimiento neuromuscular pueden sugerir la existencia potencial de otros reguladores de la recuperación diferentes a los factores periféricos (daño muscular o metabólico) exclusivamente.
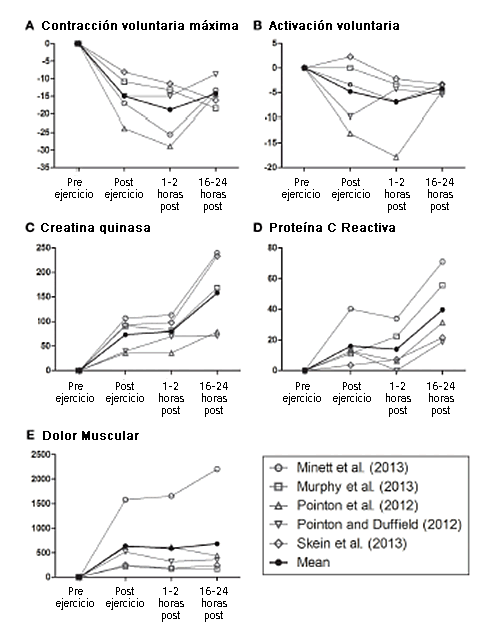
Figura1. Cambio en la evolución en el tiempo de (A) contracción voluntaria máxima de extensores de piernas, (B) activación voluntaria de extensores de piernas, (C) creatina quinasa, (D) proteína C reactiva, y (E) dolor muscular percibido luego de ejercicios de sprint intermitente prolongados. Estos datos fueron obtenidos de los estudios de Pointon y Duffield (2012), Pointon et al. (2012); Skein et al. (2013), Minett et al. (2013), y Murphy et al. (2013). Además se presenta un valor medio recopilado de cada uno de los estudios mencionados.
Finalmente, si bien la relación entre el estado de hidratación y el rendimiento en sprint intermitente sigue siendo conflictiva (Edwards y Noakes, 2009), los déficits de fluidos de 2-4% son comunes luego de la práctica de deportes de equipo (Duffield y Coutts, 2011). Hay informes que demuestran que la hipo hidratación moderada tiene efectos limitados en la potencia anaeróbica y el rendimiento en salto vertical (Hoffman et al, 1995; Cheuvront et al, 2006); sin embargo, es necesario tener cierta cautela al interpretar estos datos porque estos protocolos de evaluación reflejan sólo componentes seleccionados del rendimiento de deportes de equipo. En particular, los efectos perjudiciales de un elevado estrés cardiovascular (i.e, reducción en el volumen plasmático relacionado a la hidratación) sobre la reducción del rendimiento de ejercicios autorregulados (Périard et al, 2011) pueden ser bien aplicables a los eventos prolongados de deportes de equipo. Teniendo en cuenta el énfasis que se pone en la recuperación de fluidos luego de un partido (Reilly y Ekblom, 2005), el efecto del estado de hidratación en la recuperación debe ser despreciable cuando existe un período de tiempo extenso entre las sesiones consecutivas (Nédélec et al, 2012). No obstante, el papel de la hidratación en la recuperación no debe ser pasado por alto, porque se ha sugerido que los cambios en la osmolaridad extracelular influyen en la cinética de la glucosa y de la leucina (Keller et al, 2003). Además, las asociaciones psicológicas negativas (conscientes o de cualquier otro tipo) derivadas de un esfuerzo perceptual mayor provocado en un estado de hipo hidratación pueden afectar la fatiga mental (Devlin et al, 2001; Mohr et al, 2010). Ahora bien, pocos estudios han informado el retorno a la euhidratación luego de un partido (o de un estado de hipo hidratación) junto a alguna medida de rendimiento relacionada a sprint intermitente o a deportes de equipo.
En síntesis, las perturbaciones fisiológicas producidas dentro del músculo esquelético periférico durante ejercicios prolongados de sprint intermitente son marcadas y variadas (Bangsbo et al, 2006; Duffield y Coutts, 2011; Nédélec et al, 2012). Mientras muchos de los factores periféricos vinculados clásicamente con la fatiga durante el rendimiento agudo parecen recuperar la homeostasis rápidamente, las interrelaciones entre el agotamiento de glucógeno muscular, EIMD y la hidratación pueden ser prolongadas y preocupan a los atletas que emprenden sesiones o competencias espalda contra espalda. Sin embargo, no se ha demostrado que las tasas de cambio en los marcadores de recuperación (ej., MVC, CK, CRP, y MS) coincidan, mientras que los marcadores de hidratación y de recuperación del rendimiento luego de un partido están notablemente ausentes en la literatura. En consecuencia, parecería que a pesar del enfoque común de las intervenciones de recuperación sobre estos marcadores, ningún factor por si solo puede representar un mecanismo causal directo que explique la tasa de retorno del rendimiento físico. Más bien, puede ser relevante la regulación integradora de las perturbaciones de todo el cuerpo basadas en estos factores periféricos junto a la regulación central.
Recuperación de la función del CNC luego de ejercicios de sprint intermitente.
Aunque se supone que la disminución en el impulso del CNC luego de los ejercicios de sprint intermitente es uno de los mayores inhibidores del rendimiento de sprint luego de un partido y del desempeño de las contracciones musculares voluntarias (MVC) (Rampinini et al, 2011), la relativa invasividad y los impedimentos de tiempo que tienen las mediciones neurofisiológicas [por ejemplo, tomografía computada (CT), imaginología por resonancia magnética (MRI), estimulación magnética transcraneal (TMS); electroencefalografía (EEG) y espectroscopia cercana al infrarrojo (NIRS)] restringen el uso aplicado. En consecuencia, las pseudo mediciones de la función del CNC, tales como EMG y estimaciones de VA vía estimulación periférica o cortical, son metodologías cada vez más utilizadas, particularmente dentro de los protocolos de simulación de deportes de equipo que se utilizan en instalaciones de laboratorio (Pointon et al, 2012; Girard et al, 2013; Minett et al, 2013). Estas mediciones ofrecen estimaciones inferidas de la función del CNC, principalmente reclutamiento muscular y señalización de las neuronas motoras para que inerven de manera óptima las fibras musculares (Gandevia, 2001). Pese a que el propósito de esta discusión no es revisar estas mediciones per se, cualquier esfuerzo por entender el papel del CNC en la recuperación debe basarse en las inferencias realizadas con tales datos. Pocos estudios han evaluado directamente la función cerebral luego de ejercicios de sprint intermitente prolongado y esto representa un área para las investigaciones futuras (Minett et al, 2013).
La EMG superficial es una metodología común que se utiliza para examinar los patrones de reclutamiento neuromuscular durante el ejercicio y puede ser interpretada como una medida sustituta de comando motor descendente (Billaut, 2011). Notablemente, existe una correlación directa entre las disminuciones en las amplitudes de EMG y la producción de potencia durante ciclos de sprint repetidos de 5 s con recuperación mínima (<30s) (Billaut y Smith, 2010). De manera similar como paralelismo, las alteraciones en las estrategias de ritmo durante el ejercicio de resistencia prolongado aparentemente coinciden con cambios en el reclutamiento de unidades motoras inferido por medio de la señalización de EMG (St Clair Gibson et al, 2001; Billaut et al, 2013). Considerado las demandas neuromusculares de deportes de equipo (Cormack et al, 2008; McLellan et al, 2011), y evidencias de ritmo para conservar los esfuerzos de alta intensidad (Coutts et al, 2010; Skein et al, 2011), podría presumirse que la recuperación del rendimiento físico luego de sprint intermitente es afectada por los patrones de reclutamiento muscular subsiguientes. El análisis de actividad EMG [raíz cuadrada del valor medio de los cuadrados (RMS)] durante una contracción muscular voluntaria (MVC) del extensor de la rodilla luego de realizar protocolos de simulación de sprint intermitente de deportes de equipo, aunque sin demandas específicas de los deportes de equipo, permitió observar disminuciones post ejercicio sostenidas durante un lapso de 1 a 24 horas en la actividad de las unidades motoras (Pointon y Duffield, 2012; Pointon et al, 2012; Minett et al, 2013). Estos resultados parecen corroborar los datos de Rampinini et al. (2011), lo que sugiere que existiría una relación entre la MVC reducida y el rendimiento en sprint con la actividad EMG del día siguiente (RMS y relación RMS/PPA) después de un partido de fútbol de 90 minutos. Notablemente, la ausencia de cambio observada en las características de la onda M después de ejercicios de sprint intermitente autorregulados (Rampinini et al, 2011; Minett et al, 2013) es indicativa de excitación sostenida en el sarcolema (Allen et al, 2008) e implicaría adicionalmente que los mecanismos centrales participarían en la regulación de la recuperación post ejercicio.
Aunque hay considerables factores periféricos de confusión que pueden afectar la transmisión y propagación de la señal EMG (Farina, 2006), la estimulación nerviosa periférica se ha utilizado para evaluar inferencias de contribuciones centrales para la recuperación de la fatiga muscular después de sprints intermitentes autorregulados (Skein et al, 2011; Pointon y Duffield, 2012; Pointon et al, 2012; Minett et al, 2013) y ejercicios de deportes de equipo (Rampinini et al., 2011; Duffield et al, 2012; Murphy et al, 2013; Skein et al, 2013). Usando estametodología, nosotros y otros autores obtuvimos resultados mixtos, con alteraciones en VA durante esfuerzos repetidos isométricos (es decir, 8-15) de MVC, lo que demuestra duraciones limitadas (Duffield et al, 2012,; Skein et al, 2012b) y variables del déficit de activación agudo (1-2 h) (Pointon et al, 2012; Minett et al, 2013) y prolongado (24 h) (Rampinini et al, 2011; Pointon y Duffield, 2012). Estas alteraciones en VA serían tarea-específicas de la carga del ejercicio y del ambiente encontrado; no obstante, en condiciones de fatiga inducida por el ejercicio, una incapacidad sostenida del CNC para regular adecuadamente la musculatura esquelética durante los esfuerzos voluntarios subsiguientes, podría tener un efecto perjudicial en la recuperación del rendimiento de sprint intermitente (Rampinini et al, 2011; Bishop, 2012).
Como evidencia observacional de esto, la Figura 1 destaca la evolución en el tiempo de la recuperación post ejercicio de MVC, VA, y de marcadores relacionados al daño muscular en las 24 h posteriores al ejercicio de sprint intermitente prolongado (Pointon y Duffield, 2012; Pointon et al, 2012; Minett et al, 2013; Murphy et al, 2013; Skein et al, 2013). Como podemos observar, la evolución en el tiempo, en forma de U, de la medición de MVC post ejercicio representa la supresión y regeneración de la fuerza contráctil muscular. Notablemente, esto coincide temporalmente con la supresión y recuperación de VA. En cambio, observaciones similares no se observan en el daño muscular o en los marcadores de percepción de dolor. Dicho esto, es necesario reconocer la ausencia de otros numerosos marcadores potenciales de recuperación fisiológica en esta figura. De todos modos, el papel del cerebro en la recuperación sigue siendo incierto, ya que la activación voluntaria (VA) estimada a través de la estimulación de nervios periférico reflejaría output espinal pero no necesariamente output supraespinal (i.e, corteza motora) (Taylor, 2009). Aunque no específica de deportes, la literatura relevante, destaca que el flujo corticomotor submáximo determinado mediante TMS contribuye con la reducción de MVC a las 4h posteriores de una maratón simulada. Estas disminuciones en la regulación motoneuronal descendente se propusieron como explicación para la reducción observada en el torque de contracción central (Ross et al, 2009). Sin embargo, es necesario realizar investigaciones futuras que incluyan el uso conjunto de TMS, EEG y NIRS para confirmar los cambios en la actividad de las neuronas motoras como factor en la recuperación neuromuscular luego de ejercicios de sprint intermitentes autorregulados (De Pauw et al, 2013; Minett et al, 2013).
A pesar de la sugerencia mencionada previamente de que una mayor regulación central sea responsable de conducir la recuperación de la función contráctil esquelética (es decir, después del ejercicio), hay escasez de evidencia directa que pueda demostrar estas demandas. En particular, el uso de medidas EEG para destacar un funcionamiento cerebral alterado como evidencia de cambios en la evolución de la recuperación post ejercicio ha recibido poca atención (De Pauw et al. 2013). Ciertamente, los aumentos temporales en la actividad EEG alfa observada durante y después del ejercicio han sido vinculados con una menor activación cortical que quizás sea indicativa de fatiga central (Kubitz y Mott, 1996; Crabbe y Dishman, 2004). Notablemente, De Pauw et al. (2013) informaron que la inmersión en agua fría (CWI) luego de ejercicios de ciclismo prolongado en condiciones de calor puede acelerar la recuperación de la menor actividad beta en las áreas del cerebro asociadas con las actividades somato sensoriales. Si bien un mayor procesamiento cerebral regional en la corteza insular (BA 13) y en el giro supramarginal (BA 40) reflejaría una mayor excitación, la CWI no produjo beneficios en el rendimiento en la prueba contrarreloj subsiguiente 1 hora post ejercicio (De Pauw et al, 2013). Por consiguiente, es difícil diferenciar la respuesta emocional o somática a la sensación del agua fría (Schneider et al, 2010) durante las intervenciones post ejercicio de cualquier función cognoscitiva relacionada a los mecanismos de recuperación. No obstante, dado el enfoque de De Pauw et al. (2013) el objetivo del estudio fue examinar el funcionamiento cerebral y la recuperación en ciclistas, y no los marcadores de rendimiento. El uso de numerosas determinaciones neurofisiológicas ej., EEG, EMG, NIRS y TMS) durante intervalos de recuperación más largos es un área para las investigaciones futuras.
Aunque el rol del cerebro como un factor de contribución para la recuperación neuromuscular (tal como se requiere luego de las competencias y del entrenamiento) sigue siendo una pregunta sin respuesta, esto nos permite razonar que cualquier disminución en la capacidad física durante series consecutivas de actividades de deportes de equipo todavía puede ser explicada por modelos periféricos de rendimiento físico. Después de todo, las perturbaciones en la fisiología pre-ejercicio (por ejemplo, temperatura del core y reservas de glucógeno) se reflejan en patrones de reclutamiento neuromuscular alterados y disminuciones en los resultados de rendimiento (Bangsbo et al, 2006; Skein et al, 2012b). Sin embargo, aquí tenemos un problema, dado que las perturbaciones centrales y periféricas luego del ejercicio (entre las que se incluyen respuestas circulatorias, metabólicas y termodinámicas) descriptas como perjudiciales para el rendimiento agudo retornan rápidamente a valores dentro de los rangos homeostáticos normales, y por lo tanto no siempre coinciden con la lenta recuperación de la capacidad física. Tales eventos pueden considerar EIMD como la manera de regular la recuperación del rendimiento (Howatson y Van Someren, 2008; Twist y Eston, 2009), aunque es igualmente factible que los aumentos simultáneos en la liberación de citoquinas sensitivas del CNS (IL-1, IL-6, y TNFa) puedan aumentar las sensaciones de fatiga (Ament y Verkerke, 2009). Quedan pendientes de discusión, aunque son muy factibles las alteraciones potenciales en la recuperación percibida, que a su vez pueden afectar la fisiología neural lo que indicaría que la recuperación post ejercicio recae en una emoción proveniente del cerebro (Noakes, 2012). Sin embargo, hasta la fecha ningún estudio de investigación ha logrado aislar con éxito las respuestas perceptivas a la recuperación para mitigar el rendimiento afectado; aunque los efectos de placebos informados con varias ayudas ergogénicas propuestas pueden dar peso al rol de la recuperación percibida en el rendimiento deportivo. Notablemente, como se observa en la Figura 1 la supresión y recuperación de MVC no coincide ni está aparentemente relacionada con el aumento en el dolor muscular percibido. Evidentemente el problema sobre el dolor percibido y la recuperación percibida no es similar y las futuras investigaciones deben incluir escalas explícitas relacionadas a la recuperación i.e, REST-Q (Kellmann y Klaus-Dietrich, 2000).
Aunque la evidencia presentada destaca la desconexión entre la función inferida del CNS (i.e, VA) y varios marcadores fisiológicos periféricos (i.e, marcadores de EIMD) durante la recuperación post ejercicio, no contamos con evidencia explícita. Sin embargo, dentro de este marco teórico investigaciones adicionales realizadas en ambientes extremos podrían ayudar a aclarar este argumento, particularmente donde los factores de estrés externos exacerben demandas inducidas por el ejercicio sobre el SNC. Sabemos que el ejercicio en ambientes extremos (es decir, calor o hipoxia) reduce el rendimiento en ejercicios de sprint intermitente (Drust et al, 2005; Billaut et al, 2013). Se afirma que tales ambientes no sólo agravan las respuestas fisiológicas frente al ejercicio, si no que también invocar una precoz y mayor retroalimentación negativa del reclutamiento del músculo esquelético (Tucker et al, 2006; Noakes y Marino, 2007). Por ejemplo, ejercicios de sprint intermitente con grandes cargas de calor exógeno y endógeno provocan reducciones en la distancia recorrida, junto con mayores reducciones en MVC y VA post ejercicio que los estados fisiológicos con menor temperatura (Minett et al, 2011, 2012b; Skein et al, 2012a). Notablemente, el alivio de esa excesiva carga de calor endógena tiene como resultado el mantenimiento de MVC y VA con un mejor rendimiento físico (Minett et al, 2011, 2012b; Skein et al, 2012a). De manera similar, también se ha observado que la eliminación de esa carga de calor endógena luego del ejercicio acelera la recuperación de VA y el reclutamiento del músculo esquelético luego de ejercicios de sprint intermitente en condiciones de calor (Pointon et al, 2012; Minett et al, 2013).
Dos cosas son evidentes de la literatura anterior; en primer lugar, que en los ambientes extremos existe un rol mayor de la regulación central en la manifestación de fatiga post ejercicio; y en segundo lugar que la eliminación de los efectos asociados al ambiente extremo por intervenciones de recuperación externas acelera la recuperación de la función de neuromuscular a través de un mejor funcionamiento mediado centralmente (Pointon et al, 2012; Minett et al, 2013). En concordancia, los ambientes extremos (por ejemplo, calor) destacan un contexto en el cual la recuperación podría estar regulada tanto por los cambios dentro del CNS como por la restauración de estado fisiológico periférico (De Pauw et al, 2013). Las intervenciones de recuperación que tienen como objetivo mejorar la función del SNC también pueden ser más eficaces en estas condiciones y pueden tener un impacto mayor en la preparación del atleta para las series de ejercicios subsiguientes. No se sabe con claridad si esta mejora está regulada por la reversión del estado fisiológico para promover un mayor reclutamiento del SNC (es decir, sistema de retroalimentación), o si es la mejora en el estado fisiológico del cerebro la que promueve un mayor reclutamiento del SNC de la periferia. Las intervenciones como la inmersión en agua fría después de realizar ejercicios en condiciones de calor pueden afectar el funcionamiento del SNC a través de la remoción de la carga térmica junto con la preservación del estado fisiológico periférico. Por el contrario, las intervenciones que apuntan a la provisión de fluidos y ayudas nutricionales apropiadas i.e aminoácidos de cadena ramificada pensadas para mejorar la función del cerebro (Blomstrand, 2006) pueden de hecho actuar directamente en el cerebro para mejorar el reclutamiento subsiguiente del musculo esquelético y beneficiar la recuperación. Tales interrogantes deben ser respondidos, de modo que sea posible dar mejores recomendaciones sobre las prácticas para la recuperación en el campo.
CONCLUSIÓN
El ejercicio de sprint intermitente prolongado plantea grandes demandas fisiológicas, metabólicas, neuromusculares y de percepción. Se ha realizado mucho esfuerzo para comprender la evolución en el tiempo de la recuperación y las intervenciones propuestas para acelerar la velocidad de recuperación del rendimiento. Mientras que las perturbaciones periféricas entre las que se incluyen EIMD, el agotamiento del glucógeno y estados de hidratación son perjudiciales para la fuerza contráctil, las diferentes tasas de recuperación de rendimiento y de los marcadores fisiológicos sugieren que la recuperación no puede ser explicada exclusivamente por factores periféricos. El conocimiento actual sobre los efectos del ejercicio de sprint intermitente y la recuperación en el cerebro es escaso; aunque dada la atenuación observada en VA y las alteraciones en los patrones de reclutamiento neuromuscular, la contribución del SNC debe ser considerada importante para el proceso de recuperación. En el mismo sentido, aunque especulativo, esta interacción de perturbaciones bioquímicas, neurológicas y centrales luego de ejercicios de sprint intermitente respaldaría el conocimiento anecdótico acerca de la recuperación innata como específica de la intensidad y duración. No obstante, se necesitan datos adicionales para delinear los manejos centrales y periféricos de recuperación luego de ejercicio de sprint intermitente y poder planear y estructurar las intervenciones de recuperación adecuadas y basadas en evidencia.
Referencias
1. Akenhead, R., Hayes, P. R., Thompson, K. G., and French, D. (2013). Diminutions of acceleration and deceleration output during professional football match play. /. Sci. Med. Sport 16, 556-561. doi: 10.1016/j.jsams.2012.12.005
2. Allen, D. G., Lamb, G. D., and Westerblad, H. (2008). Skeletal muscle fatigue: cellular mechanisms. Physio!. Rev. 88, 287-332. doi: 10.1152/physrev.00015.2007
3. Ament, W., and Verkerke, G. J. (2009). Exercise and fatigue. Sports Med. 39, 389-422. doi: 10.2165/00007256-200939050-00005
4. Bangsbo, J., Mohr, M., and Krustrup, P. (2006). Physical and metabolic demands of training and match-play in the elite football player. /. Sports Sci. 24, 665-674. doi: 10.1080/02640410500482529
5. Barnett, A. (2006). Using recovery modalities between training sessions in elite athletes: does it help? Sports Med. 36, 781-796. doi: 10.2165/00007256-200636090-00005
6. Bieuzen, E, Bleakley, C. M., and Costello, J. T. (2013). Contrast water therapy and exercise induced muscle damage: a systematic review and meta-analysis. PLoS ONE 8:e62356. doi: 10.1371/journal.pone.0062356
7. Billaut, E (2011). "Electromyography assessment of muscle recruitment strategies during high-intensity exercise," in Advances in Applied Electromyography Rijeka, ed J. Mizrahi
8. Billaut, E, Kerris, J. P., Rodriguez, R. E, Martin, D. T., Gore, C. J., and Bishop, D. J. (2013). Interaction of central and peripheral factors during repeated sprints at different levels of arterial 02 saturation. PLoS ONE 8:e77297. doi: 10.1371/journal.pone.0077297
9. Billaut, E, and Smith, K. (2010). Prolonged repeated-sprint ability is related to arterial Oj desaturation in men. Int. J. Sports Physiol. Perform. 5, 197-209.
10. Bishop, D. J. (2012). Fatigue during intermittent—sprint exercise. Clin. Exp. Pharmacol. Physiol. 39, 836-841. doi: 10.1111/j.l440-1681.2012.05735.x
11. Bishop, D., Lawrence, S., and Spencer, M. (2003). Predictors of repeated-sprint ability in elite female hockey players. /. Sci. Med. Sport. 6, 199-209. doi: 10.1016/S1440-2440
12. Bishop, P. A., Jones, E., and Woods, A. K. (2008). Recovery from training: a brief review. /. Strength. Cond. Res. 22, 1015-1024. doi: 10.1519/JSC.0b013e31816eb518
13. Bleakley, C, McDonough, S., Gardner, E., Baxter, G. D., Hopkins, J. T., and Davison, G. W. (2012). Cold-water immersion.
14. Blomstrand, E. (2006). A role for branched-chain amino acids in reducing central fatigue. /. Nutr. 136, 544S-547S.
15. Borresen, J., and Lambert, M. I. (2009). The quantification of training load, the training response and the effect on performance. Sports Med. 39, 779-795. doi: 10.2165/11317780-000000000-00000
16. Bradley, P. S., and Noakes, T. D. (2013). Match running performance fluctuations in elite soccer: indicative of fatigue, pacing or situational influences? /. Sports Sci. 31, 1627-1638. doi: 10.1080/02640414.2013.796062
17. Burke, L. M., Loucks, A. B., and Broad, N. (2006). Energy and carbohydrate for training and recovery. / Sports Sci. 24, 675-685. doi: 10.1080/02640410500482602
18. Cheung, K., Hume, P. A., and Maxwell, L. (2003). Delayed onset muscle soreness. SportsMed. 33, 145-164. doi: 10.2165/00007256-200333020-00005
19. Cheuvront, S. N., Carter, R. 3rd., Haymes, E. M., and Sawka, M. N. (2006). No effect of moderate hypohydration or hyperthermia on anaerobic exercise performance. Mea Sci. Sports Exerc. 38, 1093-1097. doi: 10.1249/01.mss.0000222838. 74015.15
20. Cook, C. J., and Beaven, C. M. (2013). Individual perception of recovery is related to subsequent sprint performance. Br. J. Sports Med. 47, 705-709. doi: 10.1136/bjsports-2012-091647
21. Cormack, S. J., Newton, R. U., and McGuigan, M. R. (2008). Neuromuscular and endocrine responses of elite players to an Australian rules football match. Int. J. Sports Physiol. Perform. 3, 359-374.
22. Coutts, A. J., Mendham, A. E., and Duffield, R. - in press. (In Press). Training and nutritional needs of the older team sport athlete, in Nutrition and Performance in Masters Athletes, ed P. Reaburn. Boca Raton: CRC Press.
23. Coutts, A. J., Quinn, J., Hocking, J., Castagna, C, and Rampinini, E. (2010). Match running performance in elite Australian Rules Football. / Sci. Med. Sport. 13, 543-548. doi: 10.1016/j.jsams.2009.09.004
24. Crabbe, J. B., and Dishman, R. K. (2004). Brain electrocortical activity during and after exercise: a quantitative synthesis. Psychophysiology 41, 563-574. doi: 10.HH/j.1469-8986.2004.00176.x
25. Dawson, B., Goodman, C, Lawrence, S., Preen, D., Polglaze, T., Fitzsimons, M., et al. (1997). Muscle phosphocreatine repletion following single and repeated short sprint efforts. Scand. J. Med. Sci. Sports 7, 206-213. doi: 10.111 l/j.1600-0838.1997.tb00141.x
26. De Pauw, K., Roelands, B., Marusic, U., Tellez, H. E, Knaepen, K., and Meeusen, R. (2013). Brain mapping after prolonged cycling and during recovery in the heat. / Appl. Physiol. 115, 1324-1331. doi: 10.1152/japplphysiol. 00633.2013
27. Devlin, L. H., Fraser, S. E, Barras, N. S., and Hawley, J. A. (2001). Moderate levels of hypohydration impairs bowling accuracy but not bowling velocity in skilled cricket players. / Sci. Med. Sport. 4, 179-187. doi: 10.1016/S1440-2440
28. Drust, B., Rasmussen, P., Mohr, M., Nielsen, B., and Nybo, L. (2005). Elevations in core and muscle temperature impairs repeated sprint performance. Acta Physiol Scand, 183, 181-190. doi: 10.1111/j.l365-201X.2004.01390.x
29. Duffield, R., and Coutts, A. J. (2011). "Fatigue and the regulation of exercise intensity during team sport matches," in Regulation of Fatigue in Exercise, ed E Marino.
30. Duffield, R., Coutts, A. J., and Quinn, J. (2009). Core temperature responses and match running performance during intermittent-sprint exercise competition in warm conditions. / Strength Cond. Res. 23, 1238-1244. doi: 10.1519/JSC.0b013e318194e0bl
31. Duffield, R., Murphy, A., Snape, A., Minett, G. M., and Skein, M. (2012). Post-match changes in neuromuscular function and the relationship to match demands in amateur rugby league matches. / Sci. Med. Sport. 15, 238-243. doi: 10.1016/j.jsams.2011.10.003
32. Edwards, A. M., and Noakes, T. D. (2009). Dehydration: cause of fatigue or sign of pacing in elite soccer? SportsMed. 39, 1-13. doi: 10.2165/00007256-200939010-00001
33. Farina, D. (2006). Interpretation of the surface electromyogram in dynamic contractions. Exerc. SportSci. Rev. 34, 121-127. doi: 10.1249/00003677-200607000-00006
34. Froyd, C, Millet, G. Y., and Noakes, T. D. (2013). The development of peripheral fatigue and short-term recovery during self-paced high-intensity exercise. /. Physiol. 591, 1339-1346. doi: 10.1113/jphysiol.2012. 245316
35. Gaitanos, G. C, Williams, C, Boobis, L. H., and Brooks, S. (1993). Human muscle metabolism during intermittent maximal exercise. / Appl. Physiol. 75, 712-719.
36. Gandevia, S. C. (2001). Spinal and supraspinal factors in human muscle fatigue. Physiol. Rev. 81,1725-1789.
37. Girard, O., Bishop, D. J., and Racinais, S. (2013). Neuromuscular adjustments of the quadriceps muscle after repeated cycling sprints. PLoS ONE 8:e61793. doi: 10.1371/journal.pone.0061793
38. Girard, O., Lattier, G., Maffiuletti, N. A., Micallef, J. P., and Millet, G. P. (2008). Neuromuscular fatigue during a prolonged intermittent exercise: application to tennis. /. Electromyogr. Kinesiol. 18, 1038-1046. doi: 10.1016/j.jelekin.2007.05.005
39. Girard, O., Mendez-Villanueva, A., and Bishop, D. (2011). Repeated-sprint ability -Parti. SportsMed. 41, 673-694. doi: 10.2165/11590550-000000000-00000
40. Girard, O., Micallef, J. P., Noual, J., and Millet, G. P. (2010). Alteration of neuromuscular function in squash. /. Sci. Med. Sport. 13, 172-177. doi: 10.1016/j.jsams.2008.11.002
41. Glaister, M. (2005). Multiple sprint work. Sports Med. 35, 757-777. doi: 10.2165/00007256-200535090-00003
42. Hoffman, J. R., Stavsky, H., and Folk, B. (1995). The effect of water restriction on anaerobic power and vertical jumping height in basketball players. Int. J. Sports Med. 16,214-218. doi: 10.1055/s-2007-972994
43. Howatson, G., and Van Someren, K. A. (2008). The prevention and treatment of exercise-induced muscle damage. Sports Med. 38, 483-503. doi: 10.2165/00007256-200838060-00004
44. Impellizzeri, F. M., Rampinini, E., Coutts, A. J., Sassi, A., and Marcora, S. M. (2004). Use of RPE-based training load in soccer. Med. Sci. Sports Exerc. 36, 1042-1047. doi:10.1249/01.MSS.0000128199.23901.2F
45. Keller, U., Szinnai, G., Bilz, S., and Berneis, K. (2003). Effects of changes in hydra-tion on protein, glucose and lipid metabolism in man: impact on health. Eur. J. Clin. Nutr. 57, S69-S74. doi: 10.1038/sj.ejcn.l601904
46. Kellmann, M., and Klaus-Dietrich, G. (2000). Changes in stress and recovery in elite rowers during preparation for the Olympic Games. Med. Sci. Sports Exerc. 32, 676-683. doi: 10.1097/00005768-200003000-00019
47. Knicker, A. J., Renshaw, I., Oldham, A. R., and Cairns, S. P. (2011). Interactive processes link the multiple symptoms of fatigue in sport competition. Sports Med. 41, 307-328. doi: 10.2165/11586070-000000000-00000
48. Krustrup, P., Mohr, M., Steensberg, A., Bencke, J., Kjoer, M., and Bangsbo, J. (2006). Muscle and blood metabolites during a soccer game: implications for sprint performance. Med. Sci. Sports Exerc. 38, 1165-1174. doi: 10.1249/01.mss.0000222845.89262.cd
49. Krustrup, P., 0rtenblad, N., Nielsen, J., Nybo, L., Gunnarsson, T. P., Iaia, F. M., et al. (2011). Maximal voluntary contraction force, SR function and glycogen resynthesis during the first 72 h after a high-level competitive soccer game. Eur. f. Appl. Physiol 111, 2987-2995. doi: 10.1007/s00421-011-1919-y
50. Kubitz, K. A., and Mott, A. A. (1996). EEG power spectral densities during and after cycle ergometer exercise. Res. Q. Exerc. Sport. 67, 91-96. doi: 10.1080/02701367.1996.10607929
51. Leeder, J., Gissane, C, van Someren, K., Gregson, W., and Howatson, G. (2012). Cold water immersion and recovery from strenuous exercise: a meta-analysis. Br. f. SportsMed. 46, 233-240. doi: 10.1136/bjsports-2011-090061
52. MacRae, M. B. A., Cotter, J. D., andLaing, R. M. (2011). Compression garments and exercise. SportsMed. 41, 815-843. doi: 10.2165/11591420-000000000-00000
53. Marcora, S. M. (2008). Do we really need a central governor to explain brain regulation of exercise performance? Eur. J. Appl Physiol 104, 929-931. doi: 10.1007/s00421-008-0818-3
54. Marino, F. E. (2004). Anticipatory regulation and avoidance of catastrophe during exercise-induced hyperthermia. Comp. Biochem. Physiol B Biochem. Mol Biol 139, 561-569. doi: 10.1016/j.cbpc.2004.09.010
55. McLellan, C. P., Lovell, D. I., and Gass, G. C. (2011). Markers of postmatch fatigue in professional rugby league players. /. Strength. Cond. Res. 25, 1030-1039. doi: 10.1519/JSC.0b013e3181cc22cc
56. Minett, G. M., Duffield, R., Billaut, E, Cannon, J., Portus, M. R., and Marino, F. E. (2013). Cold-water immersion decreases cerebral oxygenation but improves recovery after intermittent-sprint exercise in the heat. Scand. J. Med. Sci. Sports. doi: 10.1111/sms.l2060. [Epub ahead of print].
57. Minett, G. M., Duffield, R., Kellett, A., and Portus, M. (2012). Effects of mixed-method cooling on recovery of medium-fast bowling performance in hot conditions on consecutive days. /. Sports Sci. 30, 1387-1396. doi: 10.1080/02640414.2012.709267
58. Minett, G. M., Duffield, R., Marino, F. E., and Portus, M. (2012). Duration-dependant response of mixed-method pre-cooling for intermittent-sprint exercise in the heat. Eur. f. Appl Physiol 111, 3655-3666. doi: 10.1007/s00421-012-2348-2
59. Minett, G. M., Duffield, R., Marino, F. E., and Portus, M. (2011). Volume-dependent response of precooling for intermittent-sprint exercise in the heat. Med. Sci. Sports Exerc. 43, 1760-1769. doi: 10.1249/MSS.0b013e318211be3e
60. Mohr, M., Krustrup, P., and Bangsbo, J. (2003). Match performance of high-standard soccer players with special reference to development of fatigue. /. Sports Sci. 21, 519-528. doi: 10.1080/0264041031000071182
61. Mohr, M., Mujika, I., Santisteban, J., Randers, M. B., Bischoff, R., Solano, R., et al. (2010). Examination of fatigue development in elite soccer in a hot environment: a multi-experimental approach. Scand. J. Med. Sci. Sports 20, 125-132. doi: 10.1111/J.1600-0838.2010.01217.X
62. Murphy, A. P., Snape, A. E., Minett, G. M., Skein, M., and Duffield, R. (2013). The effect of post-match alcohol ingestión on recovery from competitive rugby league matches. /. Strength. Cond. Res. 27, 1304-1312. doi: 10.1519/JSC.0b013e318267a5e9
63. Nédélec, M., McCall, A., Carling, C, Legall, E, Berthoin, S., and Dupont, G. (2012). Recovery in soccer: Part I: post-match fatigue and time course of recovery. Sports Med. 42, 997-1015. doi: 10.2165/11635270-000000000-00000
64. Nédélec, M., McCall, A., Carling, C, Legall, E, Berthoin, S., and Dupont, G. (2013). Recovery in soccer: part II-recovery strategies. Sports Med. 43, 9-22. doi: 10.1007/s40279-012-0002-0
65. Noakes, T. D. (2012). Fatigue is a brain-derived emotion that regulates the exercise behavior to ensure the protection of whole body homeostasis. Front. Physiol. 3:82. doi: 10.3389/fphys.2012.00082
66. Noakes, T. D., and Marino, E E. (2007). Arterial oxygenation, central motor output and exercise performance in humans. /. Physiol 585, 919-921. doi: 10.1113/jphysiol.2007.145110
67. Nosaka, K., Newton, M., and Sacco, P. (2002). Delayed-onset muscle soreness does not reflect the magnitude of eccentric exercise-induced muscle damage. Scand. f. Med. Sci. Sports 12, 337-346. doi: 10.1034/j.1600-0838.2002.10178.x
68. Périard, J. D., Cramer, M. N., Chapman, P. G., Caillaud, C, and Thompson, M. W. (2011). Cardiovascular strain impairs prolonged self-paced exercise in the heat. Exp. Physiol 96, 134-144. doi: 10.1113/expphysiol.2010.054213
69. Perrey, S., Racinais, S., Saimouaa, K., and Girard, O. (2010). Neural and muscular adjustments following repeated running sprints. Eur. J. Appl Physiol 109, 1027-1036. doi: 10.1007/s00421-010-1445-3
70. Pointon, M., and Duffield, R. (2012). Cold water immersion recovery after simulated collision sport exercise. Med. Sci. Sports Exerc. 44, 206-216. doi: 10.1249/MSS.0b013e31822b0977
71. Pointon, M., Duffield, R., Cannon, J., and Marino, E E. (2012). Cold water immersion recovery following intermittent-sprint exercise in the heat. Eur. J. Appl Physiol 112,2483-2494. doi: 10.1007/s00421-011-2218-3
72. Poppendieck, W., Faude, O., Wegmann, M., and Meyer, T. (2013). Cooling and performance recovery of trained athletes- a meta-analytical review. Int. J. Sports Physiol Perform. 8, 227-242.
73. Prasartwuth, O., Taylor, J. L., and Gandevia, S. C. (2005). Maximal force, voluntary activation and muscle soreness after eccentric damage to human elbow flexor muscles. /. Physiol 567, 337-348. doi: 10.1113/jphysiol.2005.087767
74. Racinais, S., Bishop, D., Denis, R., Lattier, G., Mendez-Villaneuva, A., and Perrey, S. (2007). Muscle deoxygenation and neural drive to the muscle during repeated sprint cycling. Med. Sci. Sports Exerc. 39, 268-274. doi: 10.1249/01. mss.0000251775.46460.cb
75. Rampinini, E., Bosio, A., Ferraresi, I., Petruolo, A.,Morelli, A., and Sassi, A. (2011). Match-related fatigue in soccer players. Med. Sci. Sports Exerc. 43, 2161-2170. doi: 10.1249/MSS.0b013e31821e9c5c
76. Reilly, T., and Ekblom, B. (2005). The use of recovery methods post-exercise. / Sports Sci. 23, 619-627. doi: 10.1080/02640410400021302
77. Roelands, B., de Koning, J., Foster, C, Hettinga, E, and Meeusen, R. (2013). Neurophysiological determinants of theoretical concepts and mechanisms involved in pacing. SportsMed. 8, 556-564. doi: 10.1007/s40279-013-0030-4
78. Ross, A., Leveritt, M., and Riek, S. (2001). Neural influences on sprint running: training adaptations and acute responses. Sports Med. 31, 409-425. doi: 10.2165/00007256-200131060-00002
79. Ross, E. Z., Middleton, N., Shave, R., George, K., and Nowicky, A. (2009). Corticomotor excitability contributes to neuromuscular fatigue following marathon running in man. Exp. Physiol 92, 417-426. doi: 10.1113/expphys-iol.2006.035972
80. Schneider, S., Askew, C. D., Abel, T., Mierau, A., and Strüder, H. K. (2010). Brain and exercise: a first approach using electrotomography. Med. Sci. Sports Exerc. 42, 600-607. doi: 10.1249/MSS.0b013e3181b76ac8
81. Skein, M., Duffield, R., Cannon, J., and Marino, E E. (2012). Self-paced intermittent-sprint performance and pacing strategies following respective precooling and heating. Eur. / Appl Physiol 112, 253-266. doi: 10.1007/s00421-011-1972-6
82. Skein, M., Duffield, R., Kelly, B. T., and Marino, E E. (2012). The effects of carbohydrate intake and muscle glycogen content on self-paced intermittent-sprint exercise despite no knowledge of carbohydrate manipulation. Eur. J. Appl Physiol 112,2859-2870. doi: 10.1007/s00421-011-2253-0
83. Skein, M., Duffield, R., Edge, J., Short, M. J., and Mundel, T. (2011). Intermittent-sprint performance and muscle glycogen after 30 h of sleep deprivation. Med. Sci. SportsExerc. 43, 1301-1311. doi: 10.1249/MSS.0b013e31820abc5a
84. Skein, M., Duffield, R., Minett, G. M., Snape, A., and Murphy, A. (2013). The effect of overnight sleep deprivation following competitive rugby league matches on post-match physiological and perceptual recovery. Int. J. Sports Physiol Perform. 8, 556-564.
85. Spencer, M., Bishop, D., Dawson, B., and Goodman, C. (2005). Physiological and metabolic responses of repeated-sprint activities: specific to field-based team sports. SportsMed. 35, 1025-1044. doi: 10.2165/00007256-200535120-00003
86. Spencer, M., Rechichi, C, Lawrence, S., Dawson, B., Bishop, D., and Goodman, C. (2005). Time-motion analysis of elite field hockey during several games Post-exercise recovery and the brain in succession: a tournament scenario. /. Sci. Med. Sport. 8, 382-391. doi: 10.1016/S1440-2440
87. St Clair Gibson, A., and Noakes, T. D. (2004). Evidence for complex system integration and dynamic neural regulation of skeletal muscle recruitment during exercise in humans. Br. }. Sports Med. 38, 797-806. doi: 10.1136/bjsm.2003. 009852
88. St Clair Gibson, A., Schabort, E. J., and Noakes, T. D. (2001). Reduced neuromus-cular activity and force generation during prolonged cycling. Am. J. Physiol. Regulatory Interactive Comp. Physiol. 281, R187-R196.
89. Taylor, J. L. (2009). Point: counterpoint: the interpolated twitch does/does not provide a valid measure of the voluntary activation of muscle. /. Appl. Physiol. 107, 354-355. doi: 10.1152/japplphysiol.91220.2008
90. Tucker, R. (2009). The anticipatory regulation of performance: the physiological basis for pacing strategies and the development of a perception-based model for exercise performance. Br. J. Sports Med. 43, 392-400. doi: 10.1136/bjsm.2008.050799
91. Tucker, R., Marie, T., Lambert, E. V., and Noakes, T. D. (2006). The rate of heat storage mediates an anticipatory reduction in exercise intensity during cycling at a fixed rating of perceived exertion. /. Physiol. 574, 905-915. doi: 10.1113/ jphysiol.2005.101733
92. Twist, C., and Eston, R. (2005). The effects of exercise-induced muscle damage on maximal intensity intermittent exercise performance. Eur. J. Appl. Physiol. 94, 652-658. doi: 10.1007/s00421-005-1357-9
93. Twist, C., and Eston, R. G. (2009). The effect of exercise-induced muscle damage on perceived exertion and cycling endurance performance. Eur. J. Appl. Physiol. 105, 559-567. doi: 10.1007/s00421-008-0935-z