
1Health and Exercise Science Research Unit, School of Applied Sciences, University of Glamorgan, Trefforest, Pontypridd, CF37 1DL, Wales, UK.
2Department of Clinical Biochemistry, Royal Glamorgan Hospital, LLantrisant, Reino Unido, CF728XR.
Resumen
INTRODUCCION
El ejercicio en bicicleta ergométrica a alta intensidad ha sido muy utilizado para determinar índices de rendimiento muscular durante el ejercicio máximo. Tradicionalmente, la fuerza resistiva seleccionada para dicho test es determinada a partir de la masa corporal total del individuo para una bicicleta ergométrica Monark cargada por fricción (i.e., 75 g/kg) (1). Estudios más recientes (2-5) han demostrado que las fuerzas tradicionales pueden ser demasiado bajas para producir rendimientos máximos y que la optimización de los protocolos puede producir mayores PPO.
Conceptualmente, seleccionar la fuerza resistiva óptima de acuerdo a la masa corporal total puede no ser el mejor enfoque. La masa libre de grasa, masa magra o masa muscular activa pueden ser mejores alternativas (4). Van mil et al. (6) sugirieron que el rendimiento óptimo durante el ejercicio en bicicleta ergométrica de alta intensidad está altamente relacionado a la masa magra corporal del individuo, o a la masa de músculos esqueléticos implicados en el ejercicio. Claramente, los individuos de igual masa corporal pueden tener composiciones corporales muy diferentes (7). Debido a que la masa corporal y no la composición corporal es el índice más comúnmente usado para la fuerza resistiva, puede ocurrir una sobreestimación o subestimación de la carga «correcta». En apoyo a estos hallazgos recientes investigaciones conducidas en nuestro laboratorio han demostrado un claro incremento en la PPO cuando los sujetos fueron optimizados para la masa magra (8). Sin embargo, el incremento observado en la potencia puede comprometer la integridad del músculos esquelético. El daño muscular inducido por el ejercicio luego de diferentes tipos de ejercicio (9-11) ha sido ampliamente reportado en humanos. El incremento en los niveles de proteínas miocelulares en el suero que ocurre en forma simultánea o luego del ejercicio puede ser usado como indicador de daño muscular (10, 12). Este daño conduce a una pérdida temporal de la capacidad de ejercicio de los músculos para la producción de fuerza, lo cual tiene implicancias para el incremento del dolor muscular y puede estar relacionado a signos histológicos de daño muscular. El daño muscular puede ser detectado por un incremento transitorio en la concentración sérica de proteínas musculares tales como la S-CK y S-Mb.
El propósito de este estudio fue comparar los cambios en los niveles de S-CK, S-Mb y [La–]b luego de una serie de 30 s a alta intensidad en bicicleta ergométrica cuando las fuerzas resistivas fueron derivadas de la TBM y FFM.
METODOS
Sujetos y Diseño Experimental
Dieciocho estudiantes universitarios aparentemente sanos se ofrecieron voluntariamente como sujetos. Antes de las evaluaciones, todos los sujetos leyeron, completaron y firmaron un formulario de consentimiento informado. El estudio fue aprobado por el comité de ética local.
Todos los sujetos estaban completamente habituados a los procedimientos experimentales antes de las evaluaciones. Los períodos de familiarización fueron realizados en tres ocasiones, duplicando las condiciones de recolección de datos experimentales. El estudio fue diseñado usando un diseño aleatorio, doble ciego y con entrecruzamiento. Dos días de descanso (ninguna actividad física) precedieron a cada test y los sujetos asistían al laboratorio luego de una noche de ayuno en un intento de atenuar cualquier influencia de la dieta sobre el rendimiento. A través de todo el estudio y tres semanas antes de la recolección de los datos, los sujetos se abstuvieron de utilizar suplementación dietaria adicional y no fueron registradas desviaciones con respecto a sus hábitos alimenticios normales (Nutri – check, Reino Unido).
Mediciones Antropométricas
Fueron determinadas la talla, masa corporal y composición corporal usando balanzas calibradas (Seca, Reino Unido), estadíometro (Seca, Reino Unido) y procedimientos de pesaje hidrostático, respectivamente. La masa corporal fue medida con una apreciación de 0,1 kg y la talla con una apreciación de 0,1 cm. La densidad corporal fue determinada usando técnicas de pesaje hidrostático previamente descritas por Behnke et al. (13). La grasa corporal relativa fue estimada a partir de la densidad corporal utilizando las ecuaciones de Siri (14). El volumen pulmonar residual fue medido usando el método modificado de respiración de oxígeno (15). La FFM fue determinada sustrayendo la masa grasa de la TBM.
Test de Fuerza Velocidad
Fue realizado un test de fuerza velocidad una semana antes del test de 30 s en bicicleta ergométrica para determinar las fuerzas resistivas óptimas para la TBM y FFM, usando procedimientos descritos por Jaskolska et al. (5). Brevemente, el test consistió de 6 esprints máximos de corta duración (6-8 s) contra fuerzas resistivas asignadas aleatoriamente (70, 75, 80, 90 y 95 g/kg). Las series de ejercicio sucesivas estuvieron separadas por un período de descanso de 5 min. La carga que produjo el mayor valor de PPO para TBM y FFM fue considerada óptima y fue usada en el test de 30 s.
Test de 30 s en Bicicleta Ergométrica
Una bicicleta ergométrica (Monark 864) fue calibrada antes de la recolección de los datos, de acuerdo a lineamientos descritos por Coleman (16). Para las pruebas de fuerza velocidad y 30 s fue usado el mismo procedimiento de calibración. Cada sujeto regresó a realizar el test restante con una semana entre las condiciones experimentales. Este período de tiempo fue propuesto para facilitar una recuperación completa. Todos los sujetos eran físicamente muy activos, utilizaron contracciones concéntricas durante las evaluaciones y estaban completamente familiarizados con las condiciones experimentales, de este modo cualquier efecto de serie repetida observable sería mínimo.
La altura del asiento fue ajustada para que la flexión parcial de la rodilla estuviera entre 170 a 175° (los 180° denotan una posición extendida de la rodilla) durante la pedaleada hacia abajo. A través de la duración del test los pies fueron sujetados en los pedales a través de punteras. Todos los sujetos permanecieron sentados durante el test y fueron verbalmente alentados para realizar un esfuerzo máximo. Todos los sujetos realizaron una entrada en calor estandarizada de 5 min de acuerdo a procedimientos previamente descritos (5).
Los índices de rendimiento fueron determinados a partir de las revoluciones de la rueda usando un programa computacional corregido para la inercia (16). La transferencia de datos fue realizada usando un sensor convenientemente montado y un registrador de potencia adherido a la horquilla del ergómetro. La frecuencia de muestreo del sensor fue de 1,8 Hz. La validez y confiabilidad de la bicicleta ergométrica como un test de potencia muscular ha sido reportada con un valor de r=0,93 (17). Los registros de frecuencia cardíaca para cada sujeto fueron medidos constantemente usando un sistema de telemetría de corto alcance (Sport Tester 3000, Polar Electro, Finlandia).
Muestras Sanguíneas
Fueron recolectadas muestras sanguíneas en duplicado en la misma hora del día y por el mismo investigador en un intento de controlar las variaciones biológicas y entre sujetos (18). En un intento de controlar los cambios en el volumen plasmático, todas las muestras en reposo fueron tomadas luego de 30 min del reposo en posición supina. Las muestras recolectadas inmediatamente post-ejercicio fueron tomadas con los sujetos en posición supina sobre una camilla clínica para minimizar el riesgo de mareos. Este procedimiento fue seguido de ambos protocolos. Además, tanto las muestras capilares como venosas fueron corregidas para los cambios en el volumen plasmático usando las ecuaciones de Dill y Costill (19).
Sangre Venosa
Fueron recolectados 10 ml de sangre venosa de la vena antecubital del antebrazo usando el sistema Vacutainer (Becton Dickinson, Rutherford, NJ, Estados Unidos). Luego de la recolección las muestras fueron inmediatamente colocadas en hielo, se permitió que se congelaran y fueron centrifugadas a 3500 rpm durante 10 min. Luego fue extraído el suero y fue colocado en tubos eppendorf (contenedores plásticos) y fueron almacenados a -80° antes de los análisis bioquímicos.
Sangre Capilar
Fueron recolectadas muestras sanguíneas en duplicado a partir del lóbulo de la oreja derecha usando un tubo capilar. Las muestras fueron inmediatamente analizadas para determinas las concentraciones de [La–]b utilizando un analizador electroquímico calibrado automáticamente (Analox PGM7 Champion, Londres, Reino Unido).
Creatinquinasa (S-CK)
Las concentraciones de S-CK fueron medidas utilizando el dispositivo Kodac Ektachem Clinical Slide (CK). Las densidades de reflexión fueron monitoreadas durante la incubación. La velocidad de cambio en la densidad de reflexión fue luego convertida a mediciones de actividad enzimática. El coeficiente de variación durante la medición fue de 4,4%.
Determinación de la Mioglobina (S-Mb)
La concentración de S-Mb fue analizada usando el Chiron Diagnostics ACS: 180® Automated Chemiluminescence System. La concentración de mioglobina fue determinada por medio de la relación directa de la cantidad presente en la muestra del paciente y la cantidad relativa de unidades de luz (RLUs) detectada por el sistema. El coeficiente de variación establecido para la medición fue de 3,5%.
Análisis Estadísticos
Las diferencias en los componentes de la potencia entre TBM y FFM fueron analizadas usando test t de Student. Fue usado un test ANOVA de dos factores para mediciones repetidas con correcciones de Bonferroni para investigar diferencias en las concentraciones de S-CK, S-Mb, S-cTnI, dentro y entre los protocolos a través de las tres etapas de muestreo sanguíneo. La significancia fue establecida a un nivel p<0,05. Todos los cálculos estadísticos fueron realizados usando el programa SPSS para Windows (SPSS, Surrey, Reino Unido) y todos los datos fueron reportados como valores medios±DS.
RESULTADOS
Las características fisiológicas y de edad de los sujetos son presentadas en la Tabla 1. Los valores de producción de potencia generados durante el estudio para los protocolos TBM y FFM son presentados en la Tabla 2. Fueron encontradas diferencias significativas (p<0,05) en PPO entre los protocolos TBM y FFM (953±114 vs. 1020±134 W, respectivamente). También fueron encontradas diferencias significativas (p<0,01) entre las cadencias de pedaleo y las fuerzas resistivas (134±8 vs. 141±7 rev./min; 6±1 vs. 5±1 kg, respectivamente). No fueron encontradas diferencias (p>0,05) entre los protocolos para la producción de potencia media (MPO), índice de fatiga (FI%), trabajo realizado (WD) o frecuencia cardíaca. Las concentraciones de S-CK, S-Mb, S-cTnI y [La–]b registradas durante el estudio son presentadas en la Tabla 3. Fueron encontradas diferencias significativas para S-CK desde la condición de reposo hasta la condición inmediatamente post-ejercicio durante los protocolos TBM y FFM (p<0,01; p<0,05, respectivamente). Los valores de [La–]b fueron también significativamente diferentes desde la condición de reposo hasta las condición inmediatamente post-ejercicio y 24 h post-ejercicio, tanto para los protocolos TBM como FFM. No fueron registradas diferencias significativas durante ninguno de los muestreos sanguíneos para S-Mb o S-cTnI dentro de los grupos (p>0,05). Sin embargo, fueron observadas diferencias significativas inmediatamente post-ejercicio entre los protocolos TBM y FFM para las concentraciones de S-Mb (p<0,05), el protocolo TBM registró los valores más altos.
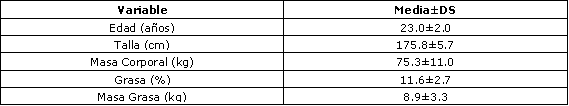
Tabla 1. Edad y características antropométricas de los sujetos (n=18).
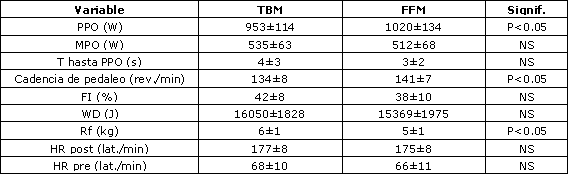
Tabla 2. Perfiles de producción de potencia en bicicleta ergométrica
tanto para los protocolos TBM como FFM, tiempo para alcanzar el PPO, cadencia de
pedaleo, índice de fatiga, trabajo realizado, fuerza resistiva y frecuencias
cardíacas máximas.
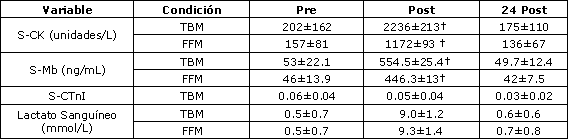
Tabla 3. Concentraciones de creatinquinasa (S-CK), mioglobina (s-Mb),
troponina cardíaca (S-CTnI) y lactato sanguíneo ([La–]b) medidas en reposo,
inmediatamente post-ejercicio y 24 h post-ejercicio para los protocolos FFM y
TBM. † Diferencias significativas a un nivel p<0.05.
DISCUSION
La troponina cardíaca no fue detectada en ningún participante a través de ninguna de las tres condiciones experimentales, tanto para el protocolo FFM como TBM, lo cual proporciona evidencia para sostener que la pérdida de S-CK ocurre predominantemente desde el músculo esquelético y que no ocurrió daño cardíaco. Los incrementos en las concentraciones séricas de proteínas intracelulares son generalmente aceptados como buenos indicadores de ruptura de las fibras musculares, y daño o permeabilidad de las membranas de las células musculares (20-22). Los mecanismos responsables del escape de proteínas intramusculares a la sangre son pobremente entendidos en la actualidad. Sin embargo, Pyne (22) ha propuesto dos mecanismos, que incluyen daño mecánico al músculo esquelético y a la membrana celular. Otros investigadores han descrito otros mecanismos (12, 23-25). Estos mecanismos incluyen daño metabólico, tasa insuficiente de producción de ATP, isquemia o hipoxia muscular, alteraciones en la concentración de iones y producción de radicales libres que resulta en peroxidación de las membranas celulares.
Las mayores concentraciones de S-CK y S-Mb observadas para el protocolo TBM, en comparación con el FFM pueden estar relacionadas a las mayores fuerzas resistivas que fueron utilizadas en el primer protocolo. Estas observaciones fueron consistentes cuando el protocolo TBM fue asignado aleatoriamente como la primera o segunda condición experimental. Los resultados de este estudio indican que las fuerzas resistivas significativamente menores que son usadas en el protocolo FFM y las mayores cadencias de pedaleo obtenidas pueden estar reflejando efectividad y eficiencia en las relaciones fuerza velocidad para el protocolo FFM en comparación con el TBM. Estas observaciones coinciden con sugerencias de Wilkie (26), quien estableció que la fuerza debería equiparar la capacidad del músculo con el objetivo de explotar completamente la relación fuerza velocidad. Los hallazgos también indican que el protocolo FFM puede estar causando menos daño mecánico a las fibras musculares y membranas celulares a pesar de que el valor de PPO registrado es mayor cuando se lo compara con el del protocolo TBM.
Kyrolainen et al. (11) midieron la concentración de anhidrasa carbónica III (S-CAIII) y encontraron que la respuesta de la proteína al rendimiento de salto fue curvilínea. Estos autores sugirieron que era requerida una determinada intensidad o umbral de ejercicio antes de que ocurriera la salida de proteínas. En este estudio no fueron encontradas relaciones entre las producciones de potencia registradas para los protocolos TBM y FFM y las concentraciones séricas de S-CK o S-Mb. Esto puede ser explicado por el hecho de que las potencias medidas y las concentraciones de proteínas observadas por Kyrolainen et al. (11) fueron obtenidas usando atletas de potencia. Los sujetos en el presente estudio fueron estudiantes universitarios y en comparación eran probablemente menos potentes. Además la duración de las evaluaciones fue menor y S-CAIII, una enzima citosólica, puede estar más concentrada en las fibras tipo I (27). S-Mb ha sido observada en altas concentraciones en las fibras tipo I y II (28). Debido a que los tipos de fibras no fueron establecidos en este estudio, es difícil realizar una comparación entre las pruebas.
Un gran problema que existe cuando se intenta determinar la producción de potencia máxima de corta duración es la dependencia de la producción de potencia sobre la velocidad de contracción (29-31). Debido a que la relación entre fuerza y velocidad tiene una forma curvilínea inversa, la potencia, el producto de la fuerza y la velocidad, tiene una relación parabólica, alcanzando un valor máximo a una determinada velocidad óptima intermedia. De este modo, la contracción a velocidades más lentas o más rápidas puede reducir la producción de potencia. Durante el presente estudio las fuerzas resistivas fueron constantes, y la contracción muscular de naturaleza concéntrica, y cualquier incremento en la cadencia de pedaleo para una dada resistencia a través de un tiempo específico resultaría en un incremento de la potencia. Sin embargo, hay diferentes factores que necesitan consideración respecto al flujo de proteínas. Estos incluyen al transporte de proteínas desde el intersticio del músculo hasta el espacio intravascular a través de los vasos linfáticos, y su velocidad de clearence. Ha sido demostrado que el flujo linfático y sanguíneo desde los músculos está muy afectado por la actividad muscular (32, 33). Las mayores concentraciones de S-CK observadas luego del protocolo TBM fueron probablemente el resultado de una mayor alteración de la membrana de las células musculares y posiblemente aceleró el transporte al suero cuando fueron comparados los protocolos TBM y FFM.
Mair et al. (34) sugirió que si el estrés mecánico es mayor que la capacidad del músculo para resistir, va a ocurrir una ruptura del aparato contráctil, miofibrillas y daño mecánico a la membrana plasmática y al retículo sarcoplasmático. Esto puede conducir luego a una alteración de la homeostasis intracelular del calcio y de la función contráctil. Estas observaciones parecen ser consistentes con las menores concentraciones halladas de S-CK y S-Mb luego del protocolo FFM. El flujo temprano de proteínas musculares puede también ser atribuido al incremento temprano en la permeabilidad de las membranas plasmáticas de las fibras musculares. Las fuerzas resistivas usadas durante el protocolo TBM pueden causar más trauma mecánico en los primeros estadios del test, acelerando así la permeabilidad de la membrana.
Esta sugerencia puede ser apoyada por las altas cadencias de pedaleo registradas para el protocolo FFM, indicando facilidad de movimiento y de este modo menos trauma mecánico en el inicio del ejercicio. Van der Meulen et al. (35) demostraron que la cantidad de daño muscular estructural inducido por el ejercicio fue significativamente menor que la cantidad de daño estimado en base a la liberación enzimática intracelular a la sangre. Estos investigadores también concluyeron que el incremento en la actividad plasmática de las enzimas musculares puede reflejar cambios en la permeabilidad de la membrana. McNeil et al. (36) demostraron que las alteraciones de la membrana causadas por la imposición de fuerza mecánica en estas estructuras frágiles son comunes en el músculo en ejercicio.
McNeil et al. (36) comentaron que en algunos casos ocurre la recuperación de estas rupturas de la membrana localizadas. En el presente estudio fueron registrados incrementos significativos en S-CK tanto para los protocolos TBM como FFM, los valores más altos registrados para el protocolo TBM pueden indicar un proceso de recuperación más lento reflejado por un mayor daño muscular y/o de la membrana. El mayor trabajo y fuerza resistiva, usados para el protocolo TBM puede incrementar el agotamiento de energía tal como es medido por el ATP. Cerny (39) sugirieron que la energía demandada por los músculos que trabajan redujo la cantidad de ATP disponible para la integridad de la membrana, lo cual condujo a una pérdida de enzimas celulares y a daño celular. Esto está en contra del protocolo FFM. Las mayores cadencias de pedaleo y menores fuerzas resistivas pueden haber resultado en un uso más económico de la energía derivada de los sistemas fosfágenos. El incremento en la PPO observado para el protocolo FFM puede también indicar un mecanismo más eficiente de resíntesis de ATP, lo cual puede contribuir a que ocurra un menor daño muscular a partir de los mecanismos descritos por Cerny et al. (39). Esta sugerencia puede explicar en parte la falta de diferencias significativas en la producción de [La–]b entre los protocolos TBM y FFM. Los valores de [La–]b indican que la glucólisis anaeróbica estuvo activa durante ambos protocolos y sugiere que la magnitud de activación fue similar para ambos grupos a pesar de los incrementos en la PPO registrados para el protocolo FFM.
Conclusiones
En conclusión, el protocolo FFM produjo una PPO significativamente mayor en comparación con el protocolo TBM. Las diferencias en la PPO estuvieron reflejadas por incrementos significativos en S-CK tanto en los protocolos TBM como FFM desde el reposo hasta la condición inmediatamente post-ejercicio, registrándose las mayores concentraciones con el protocolo TBM. No fueron registradas diferencias en las concentraciones de S-Mb entre los protocolos para ninguna de las condiciones experimentales. Sin embargo, la concentración S-Mb durante el protocolo TBM fue significativamente mayor, inmediatamente post-ejercicio, cuando fueron comparados los protocolos TBM y FFM. El incremento observado en la PPO durante el protocolo FFM, y la falta de diferencias significativas entre las concentraciones de [La–]b, sugiere que este método de selección de fuerzas resistivas representa un índice más válido de la actividad ATP-PC y/o eficiencia muscular. Las concentraciones de S-CK y S-Mb registradas indican un daño muscular significativamente menor cuando el protocolo FFM fue comparado con protocolos que incluyen al componente graso de la masa corporal.
Dirección para el Envío de Correspondencia
Baker, JS (PhD), Health and Exercise Science Research Laboratory, School of Applied Science, University of Glamorgan, Pontypridd, Wales, UK, CF37 1DL. Tel.: 01443-482972; Fax: 01443-48225; correo electrónico: [email protected].
REFERENCIAS
1. Aylon, A., Inbar, O. Bar-Or, O (1974). Relationships among measurements of explosive strength and anaerobic power. In R. C. Nelson and C. A. Morehouse (Eds) International series on sports sciences, Vol 1 Biomechanics IV, 572-577
2. Dotan, R., and Bar- Or, O (1983). Load optimization for the Wingate anaerobic test. Eur J Appl Physiol 51:409-17
3. Winter, E. M., Brooks F. B. C. and Hamley, E. J (1989). Optimized loads for external power output during brief, maximal cycling. J Sport Sci 7:69-70
4. Inbar, O. Bar-Or, O. and Skinner, S (1996). The Wingate Anaerobic Test. Human Kinetics. Leeds
5. Jaskolska, A., Goossens, P. Veenstra, B. Jakolski, A. and Skinner, J. S (1999). Comparison of treadmill and cycle ergometer measurements of force – velocity relationships and power outputs. Int J Sports Med 20:92-197
6. Van mil, E., Schoeber, N. Calvert, R. and Bar-Or, O (1996). Optimization of force in the Wingate Test for children with a neuromuscular disease. Med Sci Sport Exerc 28:1087-1092
7. McInnis, K. J., and Balady, G. J (1999). Effects of body composition on oxygen uptake during treadmill exercise: Body builders verses weight matched men. Res Quart Exerc Sport 70(2):150-156
8. Baker, J. S., Bailey, D. M. Williams, S. and Davies, B (1999). Validation of a modified aerobic shuttle run as a measure of anaerobic performance. Med Sci Sport Exerc 31(5):251
9. Balnave, C. D., and Thompson, M. W (1993). Effect of training on eccentric exercise induced muscle damage. J of Appl Physiol 75:1545-1551
10. Clarkson, M. M., Nosaka, K. and Braun, B (1993). Muscle function after exercise enduced muscle damage and rapid adaption. Med Sci Sport and Exerc 24:512-520
11. Kyrolainen, H., Timo, T. and Komi, P. V (1998). Muscle damage enduced by stretch shortening cycle. Med Sci Sport Exerc 30(3):415-420
12. Armstrong, R. B., Warren, G. L. and Warren, J. A (1991). Mechanisms of exercise induced muscle fibre injury. Sports Med 12(3):184-207
13. Behnke, A. R., and Wilmore, J. H (1974). Evaluation of body build and composition Englewood Cliffs. NJ: Prentice Hall inc. 1974:20-24
14. Siri, W. E (1956). Gross composition of the body In: Lawrence. J. H. and Tobias, C. A. Advances in Biological and Medical physics. IV. New York Academic Press
15. Wilmore, J. H., Vodak, P. A. Parr, R. B. Girandola, R. N. and Billing, J. E (1980). Further simplification of a method for determination of residual lung volume. Med Sci Sport and Exerc 12:216-218
16. Coleman, S (1996). Corrected Wingate Anaerobic Test. Cranlea users handbook. Cranlea and Co. England
17. Patton, J., Murphy, M. and Frederick, F (1985). Maximal power outputs during the Wingate anaerobic test. Inter J Sports Med 6:82-5.
18. Reilly, T., and Brooks, G. A (1982). Investigation of circadian rhythms in metabolic responses to exercise. Ergonomics 1093-1197
19. Dill, D. B., and Costill, D. L (1974). Calculation of the percentage changes in volumes of blood, plasma and red cells in dehydration. J of Appl Physiol 37:247-248
20. Ebbeling, C. B., and Clarkson, P. M (1989). Exercise induced muscle damage and adaptation. Sports Med 7:207-234
21. Janssen, G. M. E., Kuipers, H. Willems, G. M. Does, R. J. M. Janssen, M. P. E. and Geurten, P (1989). Plasma activity of muscle enzymes: quantification of skeletal muscle damage and relationship with metabolic variables. Inter J Sports Med 10:160-168
22. Pyne, D. B (1994). Exercise induced muscle damage and inflammation. Aus J Sci Med Sport 26: 49-58
23. Hortobagyi, T., and Denahan, T (1989). Variability in creatine kinase: methodological exercise, and clinically related factors. Inter J Sports Med 10:69-80
24. Roxin, L. E., Hedin, G. and Venge, P (1986). Muscle cell leakage of myoglobin after long term exercise and relation to individual performance. Inter J Sports Med 7:259-263
25. Sjodin, B., Westing, Y. H. and Apple, F.S (1990). Biochemical mechanisms for oxygen free radical formation during exercise. Sports Med 10(4):236-254
26. Wilkie, D. R (1960). Man as a source of mechanical power. Ergonomics 3:1-8
27. Vaananen, H. K., Leppilampi, M. Vuori, J and Takala, T. E. S (1986). Liberation of muscle carbonic anhydrase into serum during extensive exercise. J Appl Physiol 61:561-564
28. Zheng, A., Rahkila, J. Vuori, S. Takala, T. and Vaananen, H. K (1992). Quantification of carbonic anhydrase III and myoglobin in different fiber types of human psoas muscle. Histochemistry 97:77-81
29. Fenn., W. O. and Marsh, B. S (1935). Muscular force at different speeds of shortening. J Physiol ;85:277-297
30. Hill, A. V (1992). The maximal work and mechanical efficiency of human muscles and their most economical speed. J Physiol 56:19-41
31. Hill, A. V (1938). The heat of shortening and the dynamic constant of muscle. Proc Royal Soc B 126:136-195
32. Lindena, J., Kupper, W. and Trautschold, I (1984). Enzyme activities in thoracic duct lymph and plasma of anaesthetized, conscious, resting and exercised dogs. Eur J Appl Physiol 52:188-195
33. Lindena, J., Diedrichs, F. Wittenberg, H. and Trautschold, I (1986). Kinetic adjustment of enzyme catylitic concentrations in the extracellular space of the man, the dog and the rat. J Clin Chem Clin Biochem. 24:61-71
34. Mair, J,. Mayr, M. Muller, E. Haid, C. Artner-Dworzak, E. Laure, C. and Puschendorf, B (1995). Rapid adaptation to eccentric enduced muscle damage. Inter J Sports Med 16(6):352-356
35. Van der Meulen, J. H., Kuipers, H. and Drukker, J (1991). Relationship between exercise induced muscle damage and enzyme release in rats. J Appl Physiol 71(3):99-104
36. McNeil, P. L., Khakee, R (1992). Disruptions of muscle fiber plasma membranes: role in exercise enduced muscle damage. Amer J Path 140:1097-1109
37. Cheetham, M. E., Boobis, L. H. Brook's, S. and Williams, C (1986). Human muscle metabolism during sprint running. J of Appl Physiol 61:54-60
38. Hiroven, J., Rehunen, S. Rusko, H. and Harkonen, M (1987). Breakdown of high energy phosphate compounds and lactate accumulation during short term submaximal exercise. Eur J Appl Physiol 56:53-259
39. Cerny, F. J., and Haralambie, G (1983). Exercise induced loss of muscle enzymes. In Biochemistry of Exercise. Eds H.G. Knuttgen, J.A. Vogel, and J. Poortmans. Human Kinetics Inc. Champaign Illinois. 441-446
Beneficios cardiovasculares de un programa de ejercicio en casa en pacientes con enfermedad de células falciformes
Beneficios cardiovasculares de un programa de ejercicio en casa para pacientes con enfermedad de...
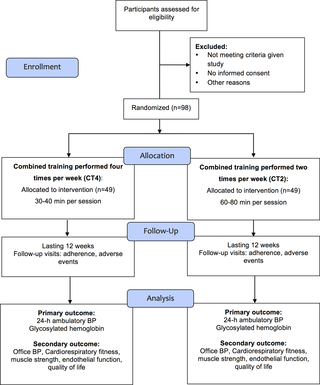
Efectos del entrenamiento combinado realizado dos o cuatro veces por semana sobre la presión arterial de 24 horas, hemoglobina glucosilada y otros resultados relacionados con la salud en personas mayores con hipertensión: Justificación y protocolo del estudio de un ensayo clínico aleatorio
Efectos del entrenamiento combinado en hipertensión y salud en adultos mayores El entrenamiento...
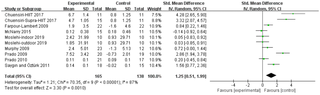
Efectos del ejercicio aeróbico y del ejercicio de resistencia sobre los índices físicos y los factores de riesgo cardiovascular en niños en edad escolar con sobrepeso y obesidad: una revisión sistemática y un metaanálisis
Ejercicio Aeróbico y de Resistencia: Soluciones para la Obesidad Infantil La obesidad infantil es...
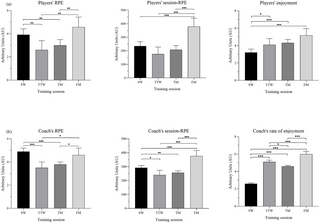
Problemas de bienestar: Su influencia en la RPE y el disfrute en el entrenamiento de waterpolo máster y la brecha jugador–entrenador
El estudio reciente sobre el bienestar y sus efectos en la carga de entrenamiento y disfrute en el...
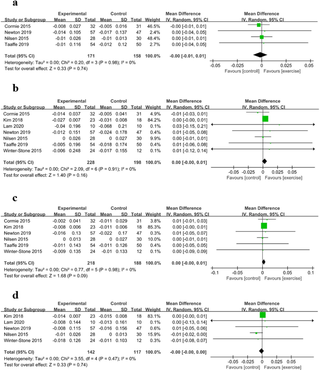
Efectos del ejercicio en la composición corporal de pacientes con cáncer de próstata que reciben terapia de privación androgénica: una revisión sistemática actualizada y un metaanálisis
Efectos del ejercicio en la composición corporal de pacientes con cáncer de próstata bajo terapia...