
Influencia de la Duración de la Recuperación sobre el Rendimiento de Esprints Repetidos en Bicicleta Ergométrica
Gavin Moir1, Michael H Stone2, Mark Glaister1, Andrew M Stewart3 y Michael Hughes4
1Department of Physical Education, Sport, & Leisure Studies, University of Edinburgh, Edinburgh, Reino Unido.
2Sports Science, United States Olympic Committee, Colorado Springs, Colorado 80903, Estados Unidos.
3School of Health Science, Faculty of Health and Environmental Sciences, Ngaa, Kaawai Oranga, Unitec, Auckland, Nueva Zelanda.
4School of Sport, PE & Recreation, UWIC, Cardiff, Reino Unido.
Artículo publicado en el journal PubliCE, Volumen 0 del año 2005.
Publicado 4 de agosto de 2008
Resumen
Palabras clave: ejercicio intermitente, producción de potencia, esprints repetidos, recuperación
INTRODUCCION
Durante un solo esprint corto (6 segundos) la provisión de energía se mantiene principalmente a partir de fuentes anaeróbicas (degradación de la fosfocreatina [PCr] y glucólisis) (17, 26), con un aporte mínimo (<10%) del metabolismo aeróbico (5, 37). La capacidad de producir y mantener una alta producción de potencia durante esprints repetidos, tal como se requiere en muchos eventos deportivos (ej., badminton, básquetbol, hockey, fútbol, squash), está determinada por la naturaleza y duración de los períodos de recuperación intermedios.
Durante la recuperación, el VO2 permanece elevado para restablecer la homeostasis a través de mecanismos como la recuperación de las reservas de O2 en los tejidos, la resíntesis de PCr, el metabolismo del lactato, y la remoción del fosfato inorgánico intracelular (Pi). Si bien la recuperación completa está determinada por la duración de cada uno de estos mecanismos, la recuperación de la producción de potencia máxima está asociada principalmente con la resíntesis de PCr (6, 7, 13, 23, 25, 46).La influencia de la duración de la recuperación en el rendimiento de esprints repetidos ha sido investigada previamente utilizando una serie de metodologías diferentes que utilizan períodos de recuperación de 30 s o mayores (3, 24, 56). Sin embargo, en muchos deportes, los períodos de recuperación intermitentes pueden, en algunos casos, durar algo más que unos pocos segundos. De hecho, en los deportes como el badminton y el squash, es habitual que los períodos de recuperación promedio sean tan cortos como de 10 s (32, 36). Por lo tanto, el objetivo de este trabajo fue estudiar la influencia de la duración de la recuperación sobre diferentes mediciones de trabajo de esprints múltiples, utilizando los períodos de recuperación típicos, semejantes a los empleados frecuentemente en muchos deportes de campo y de cancha. Aunque las investigaciones de trabajo en esprints repetidos realizadas en el laboratorio no logran reproducir correctamente las demandas de rendimiento de los deportes competitivos, las mismas representan en la actualidad, los mejores medios para valorar en forma directa la respuesta fisiológica a este tipo de trabajo.
METODOS
Enfoque Experimental del Problema
Siguiendo las recomendaciones de Capriotti et al. (12) y Glaister et al. (18), luego de la realización de 2 pruebas de familiarización, los sujetos realizaron 2 protocolos de pruebas con esprints repetidos con diferentes períodos de recuperación, y un período de descanso de al menos 24 horas entre cada protocolo. El protocolo 1 consistió en veinte esprints de 5 segundos separados por períodos de recuperación de 10 segundos. El protocolo 2 consistió en veinte esprints de 5 segundos separados por períodos de recuperación de 30 segundos. La confiabilidad del diseño y test-retest de las mediciones de producción de potencia (intervalo de coeficiente de correlación intraclase [ICC]: 0,80 - 0,97) y fatiga (intervalo de ICC: 0,81 – 0,83) utilizados en estos protocolos ha sido informada previamente (18, 19). Se solicitó a los sujetos que mantuvieran su dieta normal a lo largo del período de evaluación, que evitaran comer y beber durante la hora previa a cada prueba, y que no realizaran ejercicio activo durante 24 horas antes de cada prueba.
Sujetos
En este estudio participaron voluntariamente veinticinco varones estudiantes de educación física y ciencias del deporte de la Universidad de Edimburgo. Los valores promedio±DS de edad, altura, masa corporal, y porcentaje de grasa corporal estimado (protocolo de pliegues cutáneos en 7 sitios) (28) fueron 20,6±1,5 años, 177,2±5,4 cm, 78,2±8,2 kg, y 11,9±4,7 %, respectivamente. La aprobación ética fue concedida por la Universidad de Edimburgo, y todos los sujetos dieron su consentimiento informado por escrito. Antes de comenzar, todos los sujetos completaron un cuestionario de historial de entrenamiento, el cual reveló que todos habían participado activamente en deportes durante un promedio de aproximadamente 13 años y que el 92 % participaban actualmente en algún tipo de deporte que implicaba esprints repetidos.
Equipamiento
Todos los protocolos de evaluación fueron realizados en una bicicleta ergométrica frenada por fricción (Modelo 814E, Monark, Varberg, Suecia) que fue equipada con punteras estándar y asegurada al piso del laboratorio. La llanta de la rueda de la bicicleta ergométrica fue modificada mediante la incorporación de 90 tiras negras y blancas que fueron colocadas en interfase con una computadora a través de un opto-sensor fotoreflectivo para posibilitar un registro de alta frecuencia de la velocidad angular de la rueda.
Los gases respiratorios fueron analizados utilizando un sistema automatizado de análisis de gases respiración por respiración en línea (Vista Mini CPX, Gold Edition, Vacu-Med, Ventura, CA). El analizador fue calibrado antes de cada prueba utilizando oxígeno y anhídrido carbónico gaseosos de concentraciones conocidas (BOC gaseas, Manchester, UK), y el flujómetro fue calibrado utilizando una jeringa de 3 L (Vacu-Med). Los valores de frecuencia cardíaca fueron monitoreados continuamente a lo largo de las pruebas por medio de radiotelemetría de corto alcance (Sport-tester, Polar Electro Oy, Kempele, Finlandia). El analizador de gases fue conectado en interfase con una computadora que proporcionó la información en línea (respiración a respiración) de VO2, VCO2, índice de intercambio respiratorio (RER), y frecuencia cardíaca.
Las concentraciones de lactato en sangre fueron establecidas mediante un dispositivo portátil Lactate Pro (Arkray, KDK, Japan). La confiabilidad (r=0,99) y la validez de este equipo han sido informadas previamente (40). Todas las muestras de sangre fueron tomadas del lóbulo de la oreja hiperemizado.
Procedimientos de Evaluación
A excepción de la diferencia en la duración de la recuperación entre cada esprint (10 s vs. 30 s), la metodología aplicada en los dos protocolos de prueba de esprints repetidos fue la misma. Antes del comienzo de cada prueba, los sujetos realizaron un calentamiento de 5 minutos a 60 rpm contra una resistencia en la rueda de 1,0 kg. Antes de la primera prueba se estableció la altura del asiento que cada sujeto utilizaría en el calentamiento y en las pruebas y luego esta altura se mantuvo constante. La resistencia de la rueda fue establecida para todas las pruebas en 0,075 kg por kilogramo de masa corporal, y las rotaciones de la misma fueron registradas a una frecuencia de 18,2 Hz.
Inmediatamente después del calentamiento, los sujetos realizaron dos esprints de 5 segundos de práctica contra una resistencia igual a la que sería utilizada durante la prueba. La prueba comenzó luego de un período de descanso estacionario adicional de 3 minutos. Antes de cada prueba, se solicitó a los sujetos que permanecieran sentados en el asiento durante la misma, que comenzaran cada esprint con los pedales siempre en la misma posición, que permanecieran inmóviles durante los períodos de recuperación, y que dieran siempre el máximo esfuerzo.
Se proporcionó a los sujetos una cuenta regresiva de 5 segundos antes de cada esprint; el comienzo y el final fueron indicados por una señal de audio producida por una computadora. Los sujetos fueron estimulados verbalmente para dar el máximo esfuerzo durante cada esprint. Después del último esprint, los sujetos pedalearon a 60 rpm durante 5 minutos contra una resistencia de 1,0 kg. Se utilizó un metrónomo (Seiko UK, Tokio, Japón) para indicar la cadencia en esta parte de la prueba.
Las determinaciones de lactato en sangre se realizaron inmediatamente antes del primer esprint, después del esprint número 10, luego del esprint número 20, y 5 minutos después de las pruebas. Los índices individuales de esfuerzo percibido (RPE) fueron supervisados durante cada prueba utilizando una escala de 15 puntos (8). Las lecturas del RPE fueron registradas luego de los esprints 5, 10, 15, y 20.
Los valores de producción de potencia durante cada esprint fueron corregidos según la inercia de la rueda siguiendo los procedimientos establecidos por Lakomy (29) y promediados en intervalos de 1 segundo. A partir de estos datos, se obtuvieron para cada esprint los valores de producción de potencia máxima (PP) y media (MP). Los valores de producción de potencia a lo largo de cada protocolo de prueba de esprints repetidos fueron obtenidas como mediciones de PP máxima (PPmax), MP máxima (MPmax), PP promedio (PPmedia), y MP promedio (MPmedia). Siguiendo las recomendaciones realizadas por Glaister et al. (19), en cada prueba se calculó la fatiga a partir de la MP, utilizando el registro de disminución de rendimiento diseñado por Fitzsimons et al. (16): fatiga = 100 – [(producción de potencia total/producción de potencia ideal) × 100)], donde, producción de potencia total = suma de los valores de MP de todos los esprints y producción de potencia ideal = número de esprints × MPmax.
Análisis Estadísticos
Todos los análisis estadísticos fueron realizados utilizando el Paquete Estadístico para las Ciencias Sociales (SPSS for Windows, SPSS, Inc., Chicago, IL). Las medidas de tendencia central y dispersión se presentan como Media±DS. Los efectos de la duración de la recuperación en los valores de producción de potencia, RPE, y de lactato en sangre fueron evaluados a través de análisis de varianza de de 2 vías con mediciones repetidas. En caso de encontrar interacción, se realizaron los análisis a posteriori de Bonferonni, para determinar cuales eran los valores que provocaban las diferencias significativas. Las diferencias entre protocolos de las mediciones de fatiga, VO2, RER, frecuencia cardíaca, y el resumen de los datos de producción de potencia, fueron evaluados utilizando test-t de muestras apareadas. Los análisis mencionados proporcionaron límites de confianza de 95% para todo los efectos.
RESULTADOS
Producción de Potencia
En la Figura 1 se presentan los datos de MP de los protocolos de prueba de esprints repetidos, y en la Tabla 1 se presenta un resumen de todos los datos de producción de potencia. No se encontraron diferencias significativas (p<0,05) entre los 2 protocolos de evaluación en los valores de producción de potencia registrados durante los esprints 1-3. Sin embargo, mientras que en el protocolo 1 los valores de PPmax y MPmax correspondieron al 92% de los valores alcanzados durante el esprint 1, el valor correspondiente en el protocolo 2 fue sólo de 24%.
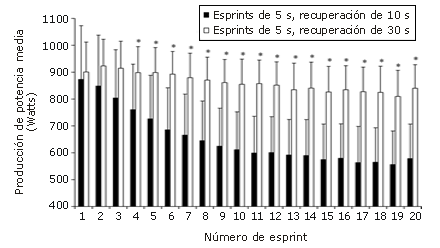
Figura 1. Datos correspondientes a la producción de potencia media obtenidos
a partir de los protocolos de ciclismo con 20 esprints de 5 segundos utilizando
diferentes períodos de recuperación. Los valores se expresan como
medias. Las barras corresponden a la desviación estándar (DS).
* Significativamente diferente (p<0,05) respecto del protocolo con período
de recuperación de 10 s.
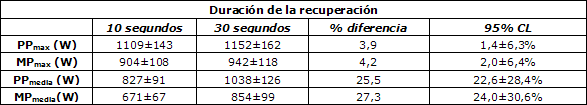
Tabla 1. Resumen de los datos de producción de potencia
provenientes de los protocolos de ciclismo con 20 esprints de 5 segundos con
períodos de recuperación diferentes (10 s o 30 s). * Nota: Los
datos de producción de potencia máxima (PP) y producción
de potencia media (MP) corresponden a mediciones obtenidas en cada esprint.
PPmax (máxima producción de potencia máxima)
MPmax (máxima producción de potencia media), PPmedia
(producción de potencia máxima promedio), y MPmedia
(producción de potencia media promedio) representan datos provenientes
de los tests de esprint múltiples; CL= Límites de confianza.
Fatiga
Los valores promedio de fatiga registrados durante los protocolos 1 y 2 fueron 25,3±6,0% y 9,2±4,5%, respectivamente. El intervalo de confianza de 95% para el verdadero valor de la diferencia en la fatiga entre los 2 protocolos fue 14,1-18,2%.
Consumo de Oxígeno
En la Figura 2 se observan los datos del VO2 de uno de los sujetos durante los 2 protocolos de evaluación. En la Tabla 2 se presenta un resumen de los datos por grupo. Si bien no se encontraron diferencias significativas entre los protocolos en el VO2 promedio durante los esprints, se observó una diferencia de 15,9% entre los protocolos en el VO2 promedio obtenido durante la recuperación (intervalo de confianza de 95%: 12,3-19,4%).
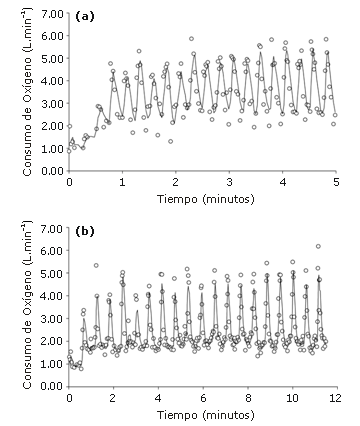
Figura 2. Datos de consumo de oxígeno pertenecientes
a un sujeto, obtenidos a partir de a) 20 esprints de 5 segundos con 10 s de
recuperación y b) 20 esprints de 5 s con 30 s de recuperación.
Nota: Los círculos vacíos representan los valores de VO2
obtenidos respiración a respiración y las líneas representan
el promedio de VO2 obtenido a partir de tres movimientos respiratorios.
Índice de Intercambio Respiratorio
En la Tabla 2 se sintetiza la respuesta del RER a los 2 protocolos. De modo general, en el protocolo 2 el RER fue 9,3 % más bajo durante los esprints (intervalo de confianza de 95%: 7,5-11,2%) y 6,6% más bajo en la recuperación (intervalo de confianza de 95%: 4,7-7,5%). En la Figura 3 se presentan los datos pertenecientes a uno de los sujetos obtenidos mediante los 2 protocolos.
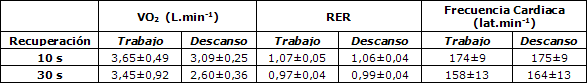
Tabla 2. Valores medios de VO2, RER y frecuencia
cardíaca obtenidos con los protocolos de ciclismo de 20 esprints de 5
s y períodos de recuperación de 10 o 30 s.* RER=índice
de intercambio respiratorio.
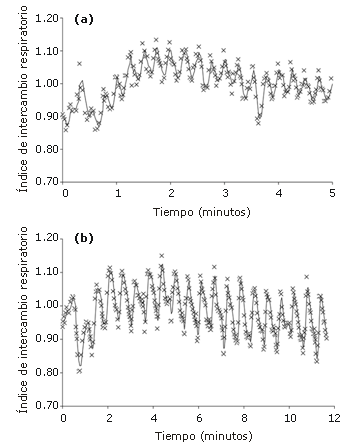
Figura 3. Datos del índice de intercambio respiratorio pertenecientes
a un sujeto, obtenidos a partir de a) 20 esprints de 5 segundos con 10 segundos
de recuperación y b) 20 esprints de 5 segundos con 30 segundos de recuperación.
Nota: Las cruces representan valores de RER obtenidos respiración por
respiración, y las líneas representan el RER promedio de 3 movimientos
respiratorios.
Frecuencia Cardiaca
Los efectos de la duración del período de recuperación en la frecuencia cardíaca se muestran en la Figura 4, y en la Tabla 2 se presenta un resumen de los datos grupales. De manera general, las frecuencias cardíacas medias durante los esprints fueron 10,1% más altas en el protocolo 1 que en el protocolo 2 (intervalo de confianza de 95%: 7,6-12,7%), durante la recuperación fueron observadas diferencias similares (6,7%) (intervalo de confianza de 95%: 4,9-9,1%).
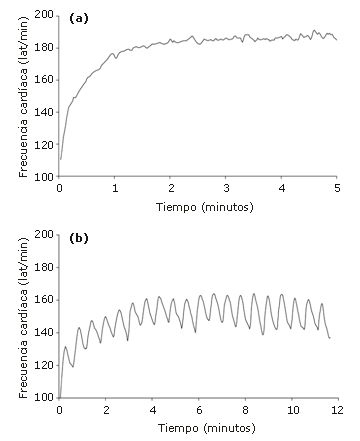
Figura 4. Valores de frecuencia cardíaca correspondientes a un sujeto
obtenidos a partir de a) 20 esprints de 5 segundos con 10 segundos de recuperación
y b) 20 esprints de 5 segundos con 30 s de recuperación. Nota: Las líneas
representan la frecuencia cardíaca promedio de 3 mediciones.
Lactato Sanguíneo
La respuesta del lactato sanguíneo a los protocolos de evaluación se ilustra en la Figura 5. No se observaron diferencias significativas en las concentraciones medias de lactato en sangre entre los 2 protocolos al comienzo de cada prueba. Sin embargo, se observaron diferencias de 24,2% (intervalo de confianza de 95%: 2,4-46,0%), 29,0% (intervalo de confianza de 95%: 12,7-45,3%), y 40,1% (intervalo de confianza de 95%: 22,7 – 57,5%) después de los esprints 10 y 20, y 5 minutos después de cada prueba, respectivamente.
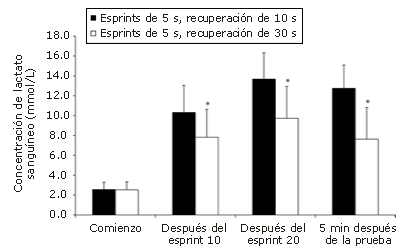
Figura 5. Valores correspondientes a la concentración de lactato sanguíneo
determinados en los protocolos de ciclismo con 20 esprints de 5 segundos utilizando
diferentes períodos de recuperación. Los datos expresados corresponden
a los valores medios mientras que las barras corresponden a la desviación
estándar (SD). * Presenta diferencias significativas (p<0.05) respecto
del protocolo con 10 s de recuperación.
Índices de Esfuerzo Percibido
El RPE aumentó progresivamente a lo largo de ambos protocolos (ver Figura 6). Entre los 2 protocolos se observaron diferencias en el RPE promedio de 11,1% (intervalo de confianza de 95%: -1,1 a 23,3%), 12,3% (intervalo de confianza de 95%: 2,3-22,3%), 10,4% (intervalo de confianza de 95%: 1,3-19,5%), y 8,0% (intervalo de confianza de 95%:: -0,6 a 16,6%) después de los esprints 5, 10, 15, y 20, respectivamente.
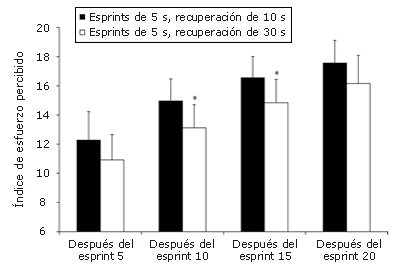
Figura 6. Valores de los índices de esfuerzo percibido observados durante
los protocolos de ciclismo con 20 esprints de 5 segundos utilizando diferentes
períodos de recuperación. Los datos se informan como valores medios;
las barras corresponden a la desviación estándar (SD). *Presenta
diferencias significativas (p<0,05) con la condición con 10 s de recuperación.
DISCUSION
El objetivo de este trabajo fue estudiar la influencia de la duración de la recuperación (10 y 30 segundos) sobre diferentes mediciones del rendimiento de esprints repetidos. La duración de la recuperación ejerció un efecto significativo sobre los valores de producción de potencia máximos (PPmax y MPmax) y medios (PPmedia y MPmedia), donde las diferencias en los valores de producción de potencia máximos podrían ser el resultado del efecto de potenciación que se produjo durante los primeros esprints en el protocolo 2. Las potenciaciones de producción de potencia a corto plazo se han observado en varias investigaciones referentes a este tipo de ejercicio (10, 21, 24, 42, 50), pero no se ha establecido aún cual es la causa de las mismas (1, 20, 48). La falta de alguna potenciación en la producción de potencia observada en el protocolo 1 sugiere que los períodos de recuperación de 10 segundos no proporcionaron la suficiente recuperación de la homeostasis para permitir que el efecto se produjera.
La diferencia significativa de MP entre los 2 protocolos reflejó la influencia considerable que tiene la duración de la recuperación sobre la fatiga. Más aún, en el protocolo 2, los sujetos pudieron mantener una producción de potencia alta, a pesar del alto nivel de acidosis metabólica. Se ha informado que concentraciones del ion hidrógeno elevadas interfieren con la actividad de diferentes enzimas involucradas en los procesos de generación de adenosín trifosfato (ATP). Sin embargo, la recuperación de la producción de potencia máxima está asociada principalmente con la resíntesis de PCr (6, 7, 13, 23, 25, 46), cuya fase inicial no sería afectada por el ambiente metabólico (43, 44, 52). Aunque la disponibilidad de PCr puede ser un factor limitante en el rendimiento, incluso antes de que las reservas de la misma sean consumidas totalmente (47), es probable que los períodos de recuperación de 30 segundos del protocolo 2 permitan que la PCr haga una contribución considerable a la resíntesis de ATP a lo largo de cada período de trabajo. Previamente se ha informado que la PCr aportaría el 80% de la provisión total de ATP en condiciones anaeróbicas durante la última serie de 10 esprints máximos de 6 segundos (con períodos de recuperación de 30 segundos) (17). En contraste, dado que se ha informado que el tiempo medio de resíntesis de PCr es de 22 segundos o más (7, 22, 30, 34), es poco probable que los períodos de recuperación de 10 segundos del protocolo 1 hayan sido suficientes para permitirle a la PCr mantener una contribución importante para la producción de potencia más allá de algunos de los primeros esprints.
La idea que sostiene que la producción de potencia durante los protocolos con esprints repetidos estaba regulada predominantemente por la disponibilidad de PCr, aporta la explicación más probable para las diferencias significativas encontradas entre los protocolos en las concentraciones de lactato en sangre. En efecto, la menor disponibilidad de PCr asociada con el protocolo 1 plantearía mayores demandas sobre la glucólisis anaeróbica para mantener la tasa de provisión de ATP requerida. Aunque la duración de la recuperación también influye en la eliminación del lactato, el tiempo medio para este proceso es de aproximadamente 9 minutos (35, 45) y por lo tanto tendría un efecto pequeño durante los cortos períodos de recuperación del presente estudio. Investigaciones previas sobre la respuesta fisiológica que se produce frente al trabajo de esprints repetidos han informado una aparente inhibición de la glucólisis durante los esprints repetidos (4, 17, 33, 37, 39, 49). Aunque el lactato en sanguíneo es solo un reflejo del equilibrio entre la producción y la degradación del mismo, el incremento progresivo en el lactato sanguíneo a lo largo de ambos protocolos plantea la necesidad de realizar investigaciones adicionales exhaustivas que estudien la inhibición de la glucólisis durante este tipo de ejercicio.
Si bien la disponibilidad de PCr aporta una explicación probable para las diferencias en la producción de potencia y fatiga encontradas entre los protocolos, los resultados también podrían ser explicados a través de diferencias entre los protocolos en la acumulación intracelular de Pi. El Pi ha sido implicado recientemente como la mayor causa de fatiga durante el ejercicio de alta intensidad como resultado de su efecto inhibitorio sobre la liberación de calcio en el retículo sarcoplásmico (2, 15, 53). Durante la recuperación, la remoción del Pi intracelular acumulado a través de la fosforilación del adenosín difosfato y de la creatina es un proceso dependiente de oxígeno (27). Así, los períodos de recuperación más largos del protocolo 2 probablemente habrían aumentado la remoción del Pi, promoviendo así un mayor acoplamiento excitación-contracción durante los esprints.
A pesar de que los períodos de recuperación más extensos del protocolo 2 permitieron que se alcanzara un mayor consumo de oxígeno, se observaron diferencias entre los protocolos en el RER durante los esprints y durante los períodos de recuperación. Debido a que el VO2 durante los esprints no fue afectado por la duración de la recuperación, probablemente las diferencias en el RER fueron el resultado de diferencias en el VCO2 como consecuencia del tamponamiento del ion hidrógeno.
Uno de los contrastes más llamativos entre los 2 protocolos de esprints repetidos se observó en la respuesta de la frecuencia cardíaca. En el protocolo 2, el patrón de respuesta de la frecuencia cardíaca fue similar al previamente informado para 10 esprints máximos de 6 segundos realizados en cinta caminadora intercalados con períodos de recuperación de 30 segundos (24). En contraste, la respuesta de la frecuencia cardíaca observada en el protocolo 1 fue mucho más parecida a la que se observa en un protocolo de ejercicio continuo. Si bien los períodos de recuperación de 10 segundos no aportaron el tiempo suficiente para que se diera alguna recuperación apreciable de la frecuencia cardiaca entre los esprints, el mismo efecto de no-recuperación estuvo ausente en las respuestas del VO2 y del RER al protocolo. Estos resultados tienen implicancias para aquellos estudios que han utilizado las relaciones frecuencia cardíaca submáxima –VO2 para estimar el costo de oxígeno en diferentes deportes que implican la realización de esprints repetidos (9, 14, 31).
Finalmente, las diferencias observadas entre los protocolo en los RPE y en la concentración de lactato sanguíneo adicionan sustento a la idea de que existe una unión entre estas variables (41, 51). Sin embargo, mientras que las mediciones de los RPE y del lactato aumentaron progresivamente a lo largo de ambos protocolos, en el protocolo 2, las mediciones de producción de potencia indicaron una cantidad relativamente pequeña de fatiga. Esta disparidad entre el lactato sanguíneo y fatiga aporta un apoyo adicional a la evidencia reciente de indica que habría un efecto mínimo de la acidosis sobre el acoplamiento excitación-contracción a temperaturas fisiológicas (11, 38, 54, 55).
Aplicaciones Prácticas
El término deportes con esprints repetidos ha sido acuñado para describir los patrones de actividad intermitentes de muchos deportes de campo y de pista. Aunque las investigaciones sobre las respuestas fisiológicas a este tipo de actividad todavía se encuentran en etapas tempranas, el trabajo que se produce en los esprints repetidos tiene un rol potencial como herramienta de entrenamiento y como un medio para evaluar las capacidades de rendimiento de atletas involucrados en deportes con esprints repetidos. De esta manera, los entrenadores y atletas necesitan ser conscientes de los efectos que pueden provocar cambios relativamente pequeños en la duración de la recuperación sobre las respuestas fisiológicas y de rendimiento frente a este tipo de ejercicio.
Agradecimientos
Los autores agradecen al Consorcio para el Desarrollo de la Universidad de Edimburgo por su aporte de subsidios para este proyecto. Los autores también agradecen al Dr. Simon Coleman por su ayuda invalorable en el desarrollo del software para las pruebas de esprints repetidos.
Referencias
1. Abbate, F., A.J. Sargeant, P.W. Verdijk, and A. De Haan (2000). Effects of high-frequency initial pulses and posttetanic potentiation on power output of skeletal muscle. J. Appl. Physiol. 88:3540
2. Allen, D.G., A.A. Kabbara, and H. Westerblad (2002). Muscle fatigue: The role of intracellular calcium stores. Can. J. Appl. Physiol. 27:8396
3. Balsom, P.D., J.Y. Seger, B. Sjodin, and B. Ekblom (1992). Maximal-intensity intermittent exercise: Effect of recovery duration. Int. J. Sports Med. 13:528533
4. Bangsbo, J (1996). Regulation of muscle glycogenolysis and glycolysis during intense exercise: In vivo studies using repeated intense exercise. In: Biochemistry of Exercise IX. R.J. Maughan and S.M. Shirreffs, eds. Champaign, IL: Human Kinetics. pp. 261275
5. Bogdanis, G.C., M.E. Nevill, L.H. Boobis, H.K. Lakomy, and A.M. Nevill (1995). Recovery of power output and muscle metabolites following 30 s of maximal sprint cycling in man. J. Physiol. 482:467480
6. Borg, G (1970). Perceived exertion as an indicator of somatic stress. Scand. J. Rehab. Med. 2:9298
7. Boyle, P.M., C.A. Mahoney, and W.F.M. Wallace (1994). The competitive demands of elite male field hockey. J. Sports Med. Phys. Fitness. 34:235241
8. Brooks, S., M.E. Nevill, L. Meleagros, H.K. Lakomy, G.M. Hall, S.R. Bloom, and C. Williams (1990). The hormonal responses to repetitive brief maximal exercise in humans. Eur. J. Appl. Physiol. 60:144148
9. Bruton, J.D., J. Lannergren, and H. Westerblad (1998). Effects of CO2-induced acidification on the fatigue resistance of single mouse muscle fibers at 28 degrees C. J. Appl. Physiol. 85:478483
10. Capriotti, P.V., W.M. Sherman, and D.R. Lamb (1999). Reliability of power output during intermittent high-intensity cycling. Med. Sci. Sports Exerc. 31:913915
11. Cherry, P.W., H.K. Lakomy, L.H. Boobis, and M.E. Nevill (1998). Rapid recovery of power output in females. Acta Physiol. Scand. 164:7987
12. Christmass, M.A., S.E. Richmond, N.T. Cable, P.G. Arthur, and P.E. Hartmann (1998). Exercise intensity and metabolic response in singles tennis. J. Sports Sci. 16:739747
13. Duke, A.M., and D.S. Steele (2001). Mechanisms of reduced SR Ca2+ release induced by inorganic phosphate in rat skeletal muscle fibres. Am. J. Physiol. 281:C418C429
14. Fitzsimons, M., B. Dawson, D. Ware, and A. Wilkinson (1993). Cycling and running tests of repeated sprint ability. Aust. J. Sci. Med. Sport. 25:8287
15. Gaitanos, G.C., C. Williams, L.H. Boobis, and S. Brooks (1993). Human muscle metabolism during intermittent maximal exercise. J. Appl. Physiol. 75:712719
16. Glaister, M., M.H. Stone, A.M. Stewart, M. Hughes, and G.L. Moir (2003). Reliability of power output during short-duration maximal-intensity intermittent cycling. J. Strength Cond. Res. 17:781784
17. Glaister, M., M.H. Stone, A.M. Stewart, M. Hughes, and G.L. Moir (2004). The reliability and validity of fatigue measures during short-duration maximal-intensity intermittent cycling. J. Strength Cond. Res. 18:459462
18. Hamilton, A.L., M.E. Nevill, S. Brooks, and C. Williams (1991). Physiological responses to maximal intermittent exercise: Differences between endurance-trained runners and games players. J. Sports Sci. 9:371382
19. Harris, R.C., R.H. Edwards, E. Hultman, L.O. Nordesjo, B. Nylind, and K. Sahlin (1976). The time course of phosphorylcreatine resynthesis during recovery of the quadriceps muscle in man. Pflüg. Arch. 367:137142
20. Hitchcock, H.C (1989). Recovery of short-term power after dynamic exercise. J. Appl. Physiol. 67:677681
21. Holmyard, D.J., M.R. Cheetham, H.K.A. Lakomy, and C. Williams (1988). Effect of recovery duration on performance during multiple treadmill sprints. In: Science and Football. T. Reilly, A. Lees, K. Davids, and W.J. Murphy, eds. London: E & FN Spon. pp. 134142
22. Holmyard, D.J., M.E. Nevill, H.K.A. Lakomy, and J. Santana Pereira (1994). Recovery of power output after maximal treadmill sprinting. J. Sports Sci. 12:140
23. Jackson, A.S., and M.L. Pollock (1978). Generalised equations for predicting body density of men. Br. J. Nutr. 40:497504
24. Lakomy, H.K (1986). Measurement of work and power output using friction-loaded cycle ergometers. Ergonomics. 29:509517
25. Laurent, D., H. Reutenauer, J.F. Payen, A. Favre-Juvin, J. Eterradossi, J.F. Lebas, and A. Rossi (1992). Muscle bioenergetics in skiers: Studies using NMR spectroscopy. Int. J. Sports Med. 13:S150S152
26. Lothian, F., and M.R. Farrally (1995). A comparison of methods for estimating oxygen uptake during intermittent exercise. J. Sports Sci. 13:491497
27. Majumdar, P., G.L. Khanna, V. Malik, S. Sachdeva, M.D. Arif, and M. Mandal (1997). Physiological analysis to quantify training load in badminton. Br. J. Sports Med. 31:342345
28. McCartney, N., L.L. Spriet, G.J.F. Heigenhauser, J.M. Kowalchuk, J.R. Sutton, and N.L. Jones (1986). Muscle power and metabolism in maximal intermittent exercise. J. Appl. Physiol. 60:11641169
29. McCully, K.K., B.P. Boden, M. Tuchler, M.R. Fountain, and B. Chance (1989). Wrist flexor muscles of elite rowers measured with magnetic resonance spectroscopy. J. Appl. Physiol. 67:926932
30. Metzger, J.M., and R.H. Fitts (1987). Role of intracellular pH in muscle fatigue. J. Appl. Physiol. 62:13921397
31. Montpetit, R.R (1990). Applied physiology of squash. Sports Med. 10:3141
32. Parolin, M.L., A. Chesley, M.P. Matsos, L.L. Spriet, N.L. Jones, and G.J.F. Heigenhauser (1999). Regulation of skeletal muscle glycogen phosphorylase and PDH during maximal intermittent exercise. Am. J. Physiol. 277:E890E900
33. Pate, E., M. Bhimani, K. Franks-Skiba, and R. Cooke (1995). Reduced effect of pH on skinned rabbit psoas muscle mechanics at high temperatures: Implications for fatigue. J. Physiol. 486:689694
34. Putman, C.T., N.L. Jones, L.C. Lands, T.M. Bragg, M.G. Hollidge-Horvat, and G.J.F. Heigenhauser (1995). Skeletal muscle pyruvate dehydrogenase activity during maximal exercise in humans. Am. J. Physiol. 269:E458E468
35. Pyne, D.B., T. Boston, D.T. Martin, and A. Logan (2000). Evaluation of the Lactate Pro blood lactate analyser. Eur. J. Appl. Physiol. 82:112116
36. Robertson, R.J., J.E. Falkel, A.L. Drash, A.M. Swank, K.F. Metz, S.A. Spungen, and J.R. Leboeuf (1986). Effect of blood pH on peripheral and central signals of perceived exertion. Med. Sci. Sports Exerc. 18:114122
37. Robinson, J.M., M.H. Stone, R.L. Johnson, C.M. Penland, B.J. Warren, and D.L. Lewis (1995). Effects of different weight training exercise/rest intervals on strength, power and high intensity exercise endurance. J. Strength Cond. Res. 9:216221
38. Roussel, M., D. Bendahan, J.P. Mattei, Y. Le Fur, and P.J. Cozzone (2000). 31P magnetic resonance spectroscopy study of phosphocreatine recovery kinetics in skeletal muscle: The issue of intersubject variability. Biochim. Biophys. Acta. 1457:1826
39. Sahlin, K., R.C. Harris, and E. Hultman (1979). Resynthesis of creatine phosphate in human muscle after exercise in relation to intramuscular pH and availability of oxygen. Scand. J. Clin. Lab. Invest. 39:551558
40. Sahlin, K., R.C. Harris, B. Nylind, and E. Hultman (1976). Lactate content and pH in muscle obtained after dynamic exercise. Pflüg. Arch. 367:143149
41. Sahlin, K., and J.M. Ren (1989). Relationship of contraction capacity to metabolic changes during recovery from a fatiguing contraction. J. Appl. Physiol. 67:648654
42. Sahlin, K., M. Tonkonogi, and K. Soderlund (1998). Energy supply and muscle fatigue in humans. Acta Physiol. Scand. 162:261266
43. Smith, J.C., A.C. Fry, L.W. Weiss, Y. Li, and S.J. Kinzey (2001). The effects of high-intensity exercise on a 10-second sprint cycle test. J. Strength Cond. Res. 15:344348
44. Spriet, L.L., M.I. Lindinger, R.S. McKelvie, G.J.F., Heigenhauser, and N.L. Jones (1989). Muscle glycogenolysis and H+ concentration during maximal intermittent cycling. J. Appl. Physiol. 66:813
45. Stone, M.H., K. Sanborn, L.L. Smith, H.S. O'Bryant, T. Hoke, A.C. Utter, R.L. Johnson, R. Boros, J. Hruby, K.C. Pierce, M.E. Stone, and B. Garner (1999). Effects of in-season (5 weeks) creatine and pyruvate supplementation on anaerobic performance and body composition in American football players. Int. J. Sport Nutr. 9:146165
46. Swank, A., and R.J. Robertson (1989). Effect of induced alkalosis on perception of exertion during intermittent exercise. J. Appl. Physiol. 67:18621867
47. Walter, G., K. VanDenBorne, K.K. McCully, and J.S. Leigh (1997). Noninvasive measurement of phosphocreatine recovery kinetics in single human muscles. Am. J. Physiol. 272:C525C534
48. Wiseman, R.W., T.W. Beck, and P.B. Chase (1996). Effect of intracellular pH on force development depends on temperature in intact skeletal muscle from mouse. Am. J. Physiol. 271:C878C886
49. Wootton, S., and C. Williams (1983). The influence of recovery duration on repeated maximal sprints. In: Biochemistry of Exercise, International Series in Sports Science XIII. H.G. Knuttgen, J.A. Vogel, and J. Poortmans, eds. Champaign, IL: Human Kinetics. pp. 269273