
La Cinética del Consumo de Oxígeno
Publicado 15 de marzo de 2013, 18:57

Todos los que llevamos a cabo alguna función dentro del ámbito del deporte, la actividad física y las ciencias del ejercicio conocemos en mayor o menor profundidad aquellos aspectos relacionados con el concepto de consumo de oxígeno y sobre todo con el de máximo consumo de oxígeno (VO2máx). Desde la introducción del concepto de VO2máx en el campo de la fisiología del ejercicio en la década de 1920 por A. V. Hill y su grupo de investigadores, se han desarrollado una inmensa cantidad de estudios experimentales con el fin de establecer sus determinantes fisiológicos, sus factores limitantes y su contribución al rendimiento en diferentes actividades deportivas y su relación con la aptitud física y la salud. De hecho, la introducción de la “Teoría del Gobernador Central” por el Dr. Timothy Noakes (1997) ha despertado un fuerte debate en la comunidad científica respecto de alguno de los tópicos previamente mencionados.
Sin embargo, un aspecto mucho menos estudiado y discutido del consumo de oxígeno es su cinética (aquí es importante señalar que el término correcto es cinética del consumo de oxígeno y no cinética del oxígeno, ya que se tratan de procesos diferentes). El término cinética puede definirse como “estudio de la velocidad a la que se producen ciertos procesos” (Diccionario de la Real Academia Española) o como “la acción de la fuerza al producir cambios de movimiento (Chamber English Dictionary). Por lo tanto, el estudio de la cinética del VO2 puede considerarse como el estudio de los mecanismos fisiológicos responsables de la respuesta dinámica del VO2 al ejercicio y su subsiguiente recuperación (Jones and Poole, 2005b). El estudio del VO2 y su regulación es importante debido a que el metabolismo oxidativo es la vía principal por la cual el organismo humano genera energía para realizar trabajo en casi todas las actividades cotidianas. Los factores tales como el mayor VO2 alcanzable, el VO2 requerido para realizar ejercicio submáximo (i.e., la economía o eficiencia de ejercicio) y la tasa a la cual el VO2 se eleva durante la transición hacia una actividad con una mayor demanda energética hasta alcanzar el estado estable, influenciarán la tolerancia individual a la actividad física (Jones and Poole, 2005b). Por esta razón, en la presente entrada de blog analizaremos los siguientes aspectos de la cinética del consumo de oxígeno:
- Perspectiva histórica
- Determinación de los modelos matemáticos para su análisis
- Sustento fisiológico/bioquímico
- Cinética pulmonar vs cinética muscular
- Utilización de diferentes parámetros (e.g., constante de tiempo, tiempo
medio de respuesta)
- Cinética “On” vs. Cinética “Off” del Consumo de Oxígeno
- Relación de la cinética con el rendimiento físico y la salud
Perspectiva Histórica
Hace casi unos 100 años, Krogh & Lindhard (1913) graficaron el consumo de oxígeno (VO2) a intervalos temporales específicos luego del comienzo de un ejercicio de moderada intensidad. Estos investigadores hallaron que el VO2 alcanzaba valores casi en estado estable dentro de 1-2 min de trabajo de muy baja intensidad y dentro de los 3 min de ejercicio de moderada intensidad; pero que no alcanzaba el estado estable con ejercicios de muy alta intensidad. A partir de los trabajos de Krog & Lindhard (1913) y de Hill & Lupton (1923), se consideró que el ejercicio sostenido más allá de unos pocos segundos depende del apropiado suministro y de la adecuada utilización del oxígeno (Burnley and Jones, 2007).
Sin embargo,
tal como lo mencionan Burnley & Jones (2007), la utilización de tests incrementales de
ejercicio y de bolsas de Douglas o análisis en cámaras de mezclado (en las
cuales los datos del intercambio pulmonar son promediados en períodos de 30-60
s) ha derivado a considerar
equivocadamente que: (i) luego de un “retraso” en el VO2 al
comienzo de un ejercicio con carga constante (el “déficit de oxígeno”) el
estado estable se alcanza luego de aproximadamente 3 min, y (ii) el VO2
se incrementa linealmente con el incremento de la potencia hasta alcanzar el VO2máx
(Figura 1).
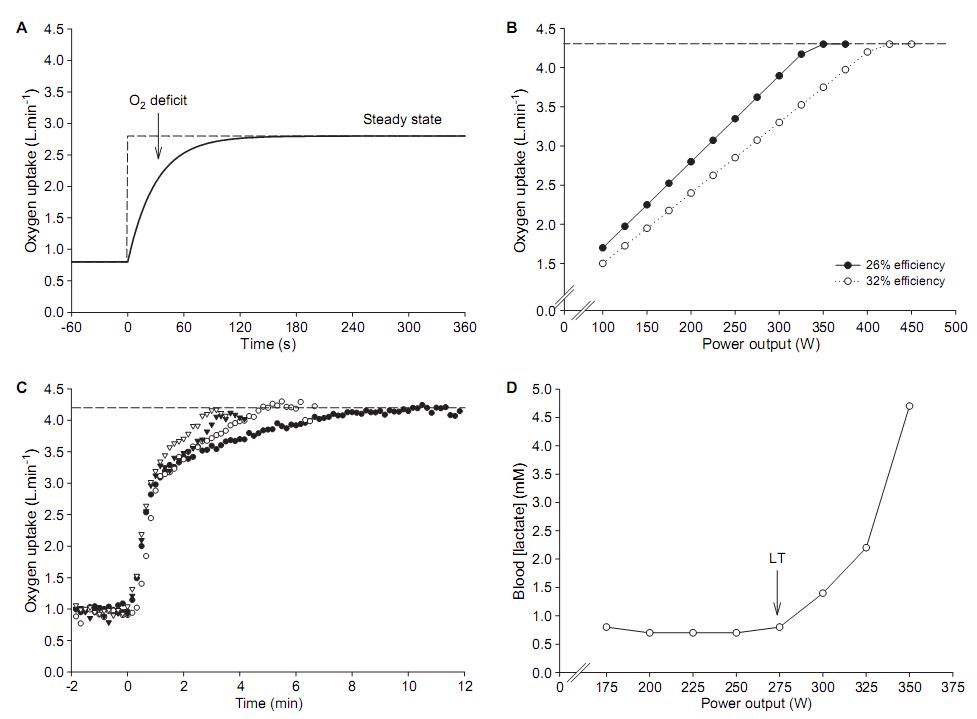
Figura 1. Conceptos tradicionales de las respuestas del consumo de oxígeno y del lactato sanguíneo durante el ejercicio. (A) Ilustración esquemática de la respuesta del VO2 durante una cicloergometría realizada a 200 W. (B) Ilustración esquemática de la respuesta del VO2 a un test incremental. Se muestran dos respuestas, y en cada una se alcanza el mismo VO2máx; sin embargo, la eficiencia mecánica de los dos individuos hipotéticos es diferente, derivando en una diferente relación entre la producción de potencia y el VO2. El individuo con la mayor eficiencia mecánica alcanza el VO2máx a una mayor producción de potencia, lo que significa que posee un mayor alcance (en términos de producción de potencia) para el trabajo aeróbico. (C) La respuesta real del VO2 es consistente cuando se realizan ejercicios de alta intensidad y corta duración (< 15 min). (D) Perfil del lactato sanguíneo durante un test incremental en cicloergómetro. Obsérvese que la concentración de lactato se mantiene a niveles basales hasta los 275 W (el umbral de lactato), pero se eleva posteriormente a este punto.
Las investigaciones de la respuesta del VO2 al comienzo del ejercicio comenzaron a principios de la década del cincuenta. Hohwü Christensen & Högberg (Hohwü Christensen and Högberg, 1950) parecen haber sido los primeros en utilizar el término “déficit de oxígeno” para describir la diferencia entre el VO2 medido y el valor obtenido en estado estable luego del comienzo del ejercicio. Henry et al (Henry, 1951, Henry and Demoor, 1956) describieron la cinética del VO2 al comienzo del ejercicio a través de modelos mono-exponenciales de uno o dos componentes, similares a los utilizados previamente para describir la recuperación post ejercicio, implicando una contribución aláctica y láctica al suministro total de energía. Hacia finales de la década de 1960 y principios de la década de 1970 Karl Wasserman y Brian Whipp, pudieron capitalizar los avances en la tecnología que facilitaron las mediciones respiración por respiración de la ventilación y el intercambio pulmonar de gases. Al mismo tiempo que Wasserman y Whipp en California comenzaron sus estudios acerca de la cinética del VO2, Paolo Carretelli y Pietro Di Prampero en Italia y Leon Farhi en Buffalo (EE.UU.) relacionaban el VO2 con la dinámica del gasto cardíaco al comienzo del ejercicio. Por muchos de sus aportes, todos estos individuos pueden considerarse los pioneros del estudio de la cinética del VO2. En contraste al modelo exponencial de dos componentes simultáneos propuesto para ejercicios de intensidad moderada a alta, Margaria et al (1965), Cerretelli et al (1966), Di Prampero et al (Di Prampero et al., 1970) y Davies et al (Davies et al., 1972), utilizaron una única ecuación exponencial para describir la respuesta incluso a ejercicios de muy alta intensidad. Whipp & Wasserman (1972) describieron el ejercicio de baja intensidad con una única ecuación exponencial pero el modelo de dos componentes utilizado para ejercicio de intensidad moderada a alta implica dos componentes exponenciales diferentes y no simultáneos. Este último modelo para ejercicio de alta intensidad evolucionó en el concepto de Fase III (Figura 2) o “componente lento del VO2” (Whipp, 1994) en el cual la mayor parte de la elevación en el VO2 está asociada con un incremento en la tasa metabólica de los músculos activos (Poole et al., 1991)
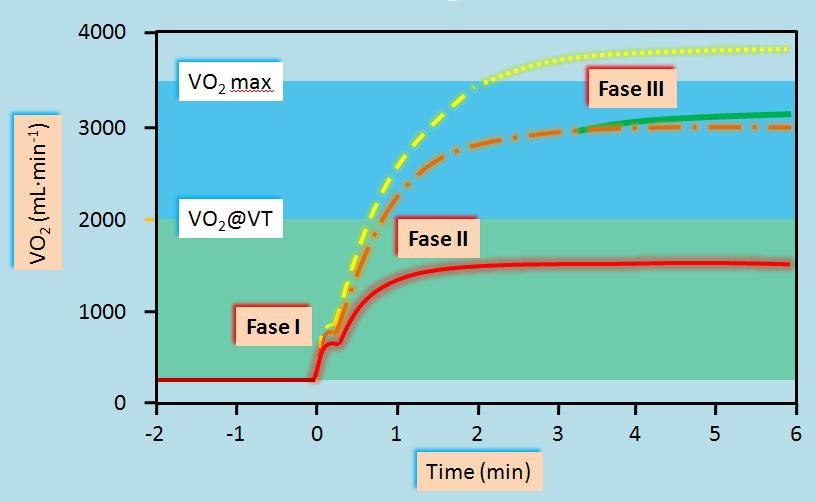
Figura 2. Representación esquemática de la respuesta del VO2 a tres diferentes intensidades de ejercicio. La línea roja inferior indica la respuesta relativamente simple a un ejercicio de baja intensidad por debajo del umbral ventilatorio (VT) durante el cual la Fase I es seguida por un único incremento mono-exponencial (Fase II) hasta que se alcanza el estado estable. La línea anaranjada (puntos y rayas) muestra que para un ejercicio entre el VT y el VO2máx normalmente aparece una Fase III que se desarrollar durante la Fase II de manera que hay una elevación adicional del VO2. La respuesta superior representada por la línea punteada amarilla muestra que no se puede alcanzar un estado estable del VO2 debido a que el nivel requerido se encuentra por encima del VO2máx [Modificado de Hughson (2009)].
Modelos Matemáticos Utilizados para el
Análisis de la Cinética del VO2
En general se acuerda que la cinética del VO2 presenta un modelo trifásico que caracterizan diferentes respuestas fisiológicas. Básicamente, la cinética del VO2 se analiza a partir de una Fase I también denominada “componente cardiodinámico” de la cinética del VO2, una Fase II y finalmente una Fase III que representa el estado estable. En los dominios de intensidad pesada (“Heavy”, entre el UL y la Potencia Crítica [CP]) y severa, se requiere un tiempo mayor para alcanzar el estado estable y los valores de VO2 observados suelen ser mayores que los que se predecirían a partir de una carga particular mediante extrapolación de la respuesta al ejercicio moderado. Este incremento del VO2 más allá de los valores predichos se ha denominado “componente lento” de la cinética del VO2. Es importante aclarar que los dominios de intensidad mencionados aquí son los que se han utilizado con mayor frecuencia en la investigación sobre la cinética del VO2, no obstante en diversos estudios también se utilizan otras clasificaciones (e.g., el dominio de intensidad pesada y severa se menciona como un único dominio de intensidad, y en algunos casos se adiciona el dominio de intensidad extrema, en cuyo caso no es posible alcanzar el estado estable). En la Tabla 1 se muestran los diferentes dominios de intensidad con sus respectivas características (obsérvese que en la Figura 1 los dominios de intensidad han sido clasificados con algunos nombres diferentes, sin embargo se trata de los mismos dominios).
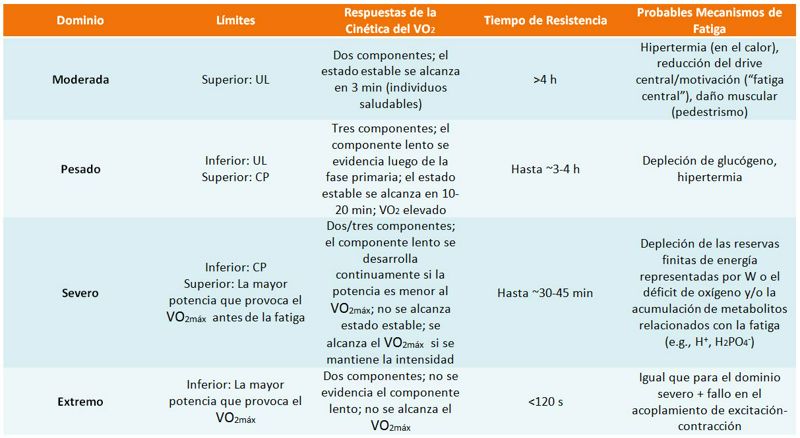
Tabla 1. Dominios de intensidad del ejercicio. UL = umbral de
lactato; CP = Potencia crítica.
La Fase I de la cinética del VO2 se produce en los primeros 10-25 s, y se cree que este incremento inicial en el VO2 se debe principalmente al incremento en el gasto cardíaco (Wasserman et al., 1974) y secundariamente a los cambios en el contenido mixto de O2 venoso (Casaburi et al., 1989b) y a las reservas de gases pulmonares (Barstow and Mole, 1987). La Fase II sigue a la Fase I, y en esta fase se produce un rápido incremento exponencial hasta un nivel de VO2 en estado estable para ejercicios de moderada intensidad por debajo del umbral de lactato (UL) (Barstow, 1994, Casaburi et al., 1989a, Whipp and Wasserman, 1972). La Fase II se denomina “componente primario” o “componente principal” de la cinética del VO2. La Fase III representa un nuevo nivel de VO2 en estado estable. Con cargas de trabajo por encima del UL, el VO2 en función del tiempo (VO2t) no sigue la misma respuesta temporal observada con cargas por debajo del UL, sino que se produce un continuo incremento en el VO2 hasta alcanzar un mayor valor que el estimado a partir de los valores iniciales de VO2. Este lento incremento en el VO2 durante ejercicios con carga constante ha sido denominado componente lento de la cinética del consumo de oxígeno y se cree que comienza luego de unos 100-200 s después de iniciado el ejercicio (Barstow and Mole, 1991).
Un aspecto esencial en la caracterización de la respuesta dinámica del VO2 con el ejercicio es, por lo tanto, establecer en una primera instancia el dominio de intensidad sobre la que se analizará la respuesta cinética. Debido a que la cinética del VO2 es altamente dependiente de la intensidad (Barstow, 1994, Grassi, 2003, Jones and Poole, 2005a, Linnarsson, 1974, Whipp and Mahler, 1980, Whipp et al., 2005), solo es monoexponencial con cargas de trabajo moderadas. A mayores cargas de trabajo, el componente lento de la cinética del VO2, de comienzo retrasado, se superpone a la respuesta exponencial fundamental (Figura 3), causando que el VO2 se incremente a valores mayores que los estimados a partir de la respuesta en el dominio de intensidad moderada (i.e., debajo del UL). Durante un ejercicio de alta intensidad (i.e., entre el UL y lo que ha sido denominado potencia crítica [CP]) (Monod and Scherrer, 1965, Moritani et al., 1981), el VO2 alcanza un estado estable retrasado, solo luego de que el exceso del VO2 ha alcanzado un valor asintótico. El ejercicio de muy alta intensidad (entre la CP y el VO2máx) está caracterizado por una respuesta del VO2 que continua incrementándose inexorablemente a través del test hasta, o hacia, el VO2máx (de manera tal que no puede alcanzarse un estado estable para el VO2).
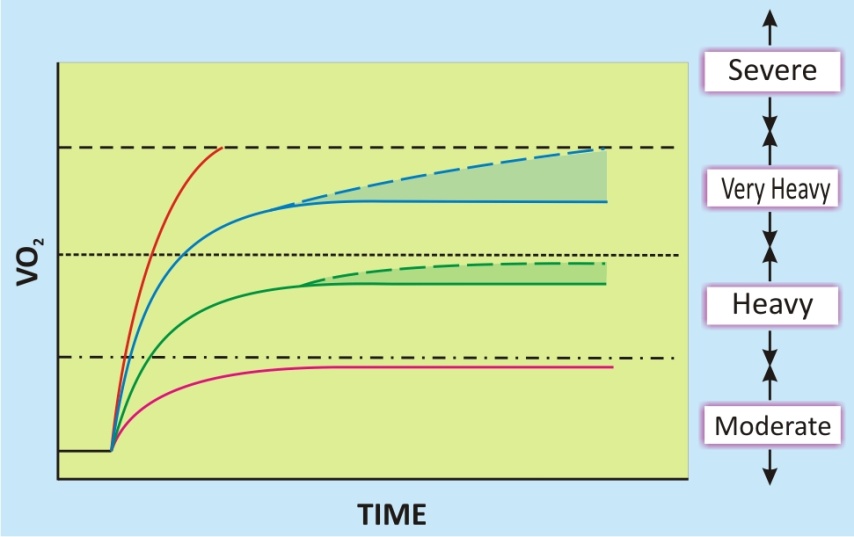
Figura 3. Representación esquemática de la respuesta del consume de oxígeno (VO2) durante un ejercicio con carga constante a diferentes intensidades. (- - -) consumo máximo de oxígeno; (·········) potencia crítica; (− · − · −) umbral de lactato. Las zonas sombreadas muestran el “componente lento” de la cinética del VO2
En general se acuerda que el análisis de la cinética del VO2 en el dominio de intensidad moderada (por debajo del UL) puede caracterizarse a partir de un modelo monoexponencial cuya expresión matemática es:
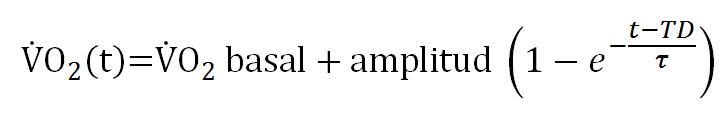
Donde VO2(t) es el VO2 a cualquier tiempo t, VO2 basal es el VO2 antes del inicio del ejercicio, la amplitud es el “estado estable” hacia el cual se proyecta el VO2, TD es el tiempo de retraso que precede al incremento en el VO2 muscular y τ es la constante de tiempo que describe la tasa a la cual el VO2 se incrementa hacia el estado estable. Es importante señalar que la utilización de un modelo monoexponencial excluye la fase cardiodinámica o Fase I de la cinética del VO2, debido a que se ha sugerido que incluir la Fase I en el análisis de la cinética del VO2 puede obscurecer las conclusiones de este análisis. Aún se debate cual es el enfoque más apropiado para dar cuenta de la Fase I y el debate está dividido entre aquellos que ajustan el término exponencial de la Fase I para determinar la transición Fase I-Fase II y aquellos que eliminan los primeros 20 s de datos luego del comienzo del ejercicio para así aislar la Fase II. La constante de tiempo (τ) es una medida del tiempo necesario para que el VO2 alcance el 63% de su amplitud final: Una vez transcurridas dos constantes de tiempo el VO2 habrá alcanzado el 86% de su amplitud final (i.e., 0.63 + 063 × [1.0 – 0.63] = 0.86). Una vez transcurridas tres constantes de tiempo, la respuesta del VO2 habrá alcanzado aproximadamente el 95% de la amplitud final y luego de transcurridas cuatro constantes temporales, el VO2 habrá alcanzado más del 98% de su amplitud final y la respuesta estará esencialmente completa.
Para el ejercicio en los dominios “pesado” y “severo” de la intensidad, se requiere un término exponencial adicional para ajustar satisfactoriamente la respuesta del VO2 luego de haberse completado la Fase I.
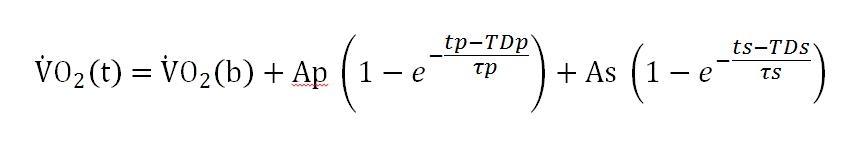
Donde Ap y As son las amplitudes de los componentes primario y lento, respectivamente; TDp y TDs son los tiempos de retraso independientes antes del inicio de los componentes primario y lento, respectivamente; y τp y τs son las constantes de tiempo para los componentes primarios y lento, respectivamente.
Para dar cuenta del modelo trifásico de la cinética del consumo de oxígeno (i.e., Fase cardiodinámica + Componente Primario + Componente Lento), Stirling et al., (2005) indican la utilización del siguiente modelo:
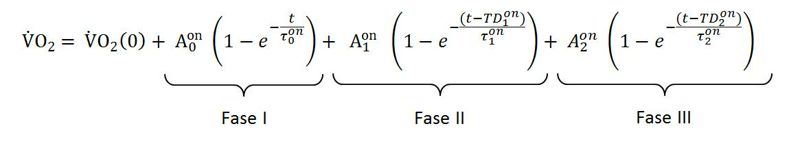
En este punto es importante señalar que la mayoría de los estudios acerca de la cinética del VO2 analizan el componente primario de la cinética, descartando el componente cardiodinámico. Por lo cual, en general se utiliza un modelo monoexponencial.
Es importante señalar que la cinética del VO2 cambia en relación con la duración y la intensidad del ejercicio. Por este motivo, Stirlig et al. (2005) propusieron un nuevo modelo para valorar la cinética del VO2. En este modelo el VO2(v,t) representa la serie temporal para la cinética del consumo de oxígeno que cambia tanto con el tiempo (t) como con la intensidad del ejercicio (v). Sin entrar en demasiados detalles sobre este modelo, la cinética del VO2 puede modelarse de la siguiente manera:
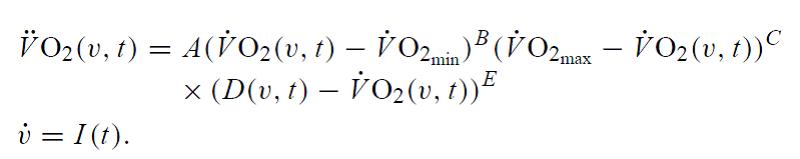
Sustento Fisiológico/Bioquímico
¿Cuáles son los factores asociados con la cinética del VO2? Para examinar esto veamos un ejemplo. Luego de que suena el disparo que da comienzo a una carrera, un atleta debe acelerar desde una posición estática hasta el ritmo de carrera requerido dentro de unos pocos segundos. Dependiendo de la distancia (y por ende de la duración) de la carrera y del estatus de entrenamiento del atleta, la tasa de recambio de ATP en los músculos activos puede incrementarse unas 100 veces en los primeros segundos. Para evitar una reducción catastrófica en la concentración muscular de ATP y el agotamiento casi inmediato, el ATP es resintetizado a través de la aceleración de la reacción de la creatina quinasa, con una consecuente reducción de la concentración de fosfocreatina y de la tasa glucolítica, resultando en un incremento en la concentración de H+ y de lactato. Simultáneamente, los cambios en el potencial de fosforilación en los músculos (e.g., reducción del ATP, incremento del ADP y Pi) son comunicados a la mitocondria, estimulando un incremento en la tasa de fosforilación oxidativa (y por lo tanto en el VO2). El “drive” para esto último depende de la diferencia entre el VO2 instantáneo y el VO2 requerido (asumiendo que la tasa de recambio de ATP en estado estable puede ser cubierta por el suministro de ATP a través de la vía oxidativa y esto no ocurrirá durante un ejercicio donde los requerimientos para el estado estable de VO2 excedan el VO2máx del sujeto). La consecuencia necesaria de esto es el incremento exponencial característico en el VO2 desde los valores pre ejercicio hacia el VO2 requerido en estado estable (Whipp and Rossiter, 2005). La magnitud del “déficit de O2” y por lo tanto la magnitud de la contribución a nivel de fosforilación de sustratos y las perturbaciones en la homeostasis muscular, es función de la amplitud de la respuesta del VO2 (VO2 en estado estable menos el VO2 previo al comienzo del ejercicio), y de la constante de tiempo del VO2 (un parámetro que describe la tasa a la cual el VO2 se incrementa hacia el estado estable) (Whipp and Rossiter, 2005)
En la actualidad se reconoce que un más rápida cinética del VO2 [observada por ejemplo luego del entrenamiento (Cerretelli et al., 1979)] está asociada con un menor déficit de O2, una menor alteración de la homeostasis celular y con consecuencias positivas para la tolerancia al ejercicio y la fatiga muscular. El interés en el estudio de la cinética del VO2 deriva del hecho de que dicha investigación permite obtener información valiosa acerca de los mecanismos básicos del control metabólico durante la contracción muscular. En este sentido, si bien el estado estable metabólico provee un escenario estable para la recolección de datos; las respuestas transitorias proveen la mejor información acerca de los sistemas de control (Grassi, 2006b). La cuestión de los factores limitantes o regulatorios de la cinética del VO2 ha sido cuestión de debate y controversia por muchos años. Las dos corrientes entre las que se divide este debate son:
(i) Aquellos que indican que el ajuste finito de la cinética del VO2 con el incremento de la carga se atribuye principalmente a un retraso en la activación metabólica o a una lentitud intrínseca del metabolismo oxidativo intracelular para ajustarse a los nuevos requerimientos; lo cual se ha denominado como Hipótesis de la Inercia Oxidativa o “Limitación Metabólica” (di Prampero and Margaria, 1968, Grassi, 2005, Margaria et al., 1965)
(ii) Aquellos que indican que uno de los principales factores limitantes es la cinética finita del transporte de O2 hacia las fibras musculares; lo cual se ha denominado Hipótesis de la Disponibilidad de O2 o “Limitación del Transporte de O2” (Hughson et al., 2001, Tschakovsky and Hughson, 1999)
En este sentido, aquellos resultados experimentales que indiquen una incrementada PO2 microvascular o un incremento en la oxigenación durante la fase inicial de la transición reposo-ejercicio indicaría un más rápido ajuste del suministro de O2 vs el del VO2; proveyendo evidencia indirecta contra la hipótesis de la “limitación del transporte de O2”. A la inversa, una reducción en la PO2 microvascular u oxigenación, sugeriría un más rápido ajuste del VO2 vs. el suministro de O2, proveyendo evidencia indirecta a favor de la hipótesis de la “limitación del transporte de O2”. Sin embargo, los resultados experimentales obtenidos en diversos estudios (Grassi et al., 2003, Behnke et al., 2002) sugieren que en los segundos posteriores al incremento en la carga de trabajo la extracción de O2 no exhibe cambios (o se reduce muy ligeramente), lo cual refleja un ajustado acople entre el suministro de O2 y el VO2. Asimismo, los resultados de los estudios citados sugieren que el rápido y pronunciado incremento en el suministro de O2 durante la transición permite al VO2 incrementarse incluso en ausencia de cambios en la extracción de O2. Solo luego de este retraso inicial, el incremento en la extracción de O2 a nivel muscular contribuirá, conjuntamente con el suministro de O2, a incrementar adicionalmente el VO2.
Entre los factores limitantes/reguladores de la cinética del VO2 se han propuesto los siguientes:
- Convección
y Difusión del O2
- Actividad
de la Piruvato Deshidrogenasa (PDH)
- Inhibición
de la Respiración Mitocondrial por Óxido Nítrico (NO)
- Hidrólisis
de la PCr como “Buffer Energético”
En condiciones “normales” (e.g., normoxia, suministro normal de O2, ausencia de condiciones patológicas), el suministro convectivo y difusivo de O2 hacia las fibras musculares esqueléticas no parece representar un importante determinante de la cinética de ajuste de la fosforilación oxidativa luego del incremento en las demandas metabólicas. Por lo tanto, en condiciones “normales” los factores limitantes de la cinética del VO2 parecen ubicarse principalmente dentro de las fibras musculares (Grassi, 2006a). Por otra parte, el rol de la PDH como factor limitante de la cinética del VO2 no ha sido confirmado experimentalmente. En relación con el NO, entre la gran cantidad de funciones que este realiza, parece inhibir completamente el VO2 en la cadena de transporte de electrones; específicamente a nivel de la citocromo oxidasa c (Brown, 2000). A través de sus efectos combinados de vasodilatación e inhibición del VO2, el NO podría servir como parte de un mecanismo de retroalimentación cuyo objetivo sería incrementar el suministro de O2 y reducir la dependencia en la extracción de O2 para cubrir el incrementado VO2 muscular. A través de este mecanismo, el NO podría actuar en el mantenimiento de altos niveles de PO2 intramiocito durante el ejercicio (Kindig et al., 2001). En este sentido, el rol de la inhibición mitocondrial por NO requiere de mayor investigación. Por lo tanto, al parecer, los determinantes principales de la cinética del VO2 muscular están asociados con el interjuego entre los diferentes mecanismos de provisión de energía al comienzo del ejercicio. Al actuar como buffer energético de alta capacitancia, la hidrólisis de la PCr y la glucólisis rápida podrían retrasar o atenuar el incremento en la [ADP] dentro de la célula luego del rápido incremento en la demanda de ATP, amortiguando así la más rápida activación de la fosforilación oxidativa.
Para comprender de qué modo los mecanismos de provisión de energía el comienzo del ejercicio desempeñan un rol central en la cinética del VO2 y la posibilidad de su aceleración con el entrenamiento es importante introducir el concepto de estabilidad metabólica absoluta y estabilidad metabólica relativa. En principio una buena estabilidad metabólica durante la transición reposo-ejercicio en los músculos esqueléticos significa una menor reducción en la [PCr] y en el potencial citosólico de fosforilación, así como también un menor incremento en [Pi], [ADPLIBRE], [AMPLIBRE] e [IMPLIBRE] para un incremento dado en el consumo de oxígeno/intensidad del ejercicio (Zoladz et al., 2006). La estabilidad metabólica absoluta, por ejemplo, de la concentración de ADP libre se refiere a los cambios absolutos en [ADPLIBRE] (en µM), mientras que la estabilidad relativa (en %, o expresada en múltiplos de los valores de reposo) se refiere a los cambios relativos en [ADPLIBRE]. Estos dos tipos de estabilidad metabólica no son equivalentes entre sí. La distinción entre estos dos tipos de estabilidad metabólica es muy importante ya que, la estabilidad metabólica absoluta (especialmente en términos de [PCr]) es relevante para el τp de la cinética pulmonar del VO2; mientras que la estabilidad relativa (especialmente en términos de [ADPLIBRE]) se refiere al orden de reacción (pendiente de la relación VO2/[ADPLIBRE]) fenomenológica (esto es, observada in vivo como consecuencia no solo de la dependencia mecanicista VO2/[ADPLIBRE] sino también como consecuencia de la activación directa de la fosforilación oxidativa). Por lo tanto, la estabilidad relativa está asociada a los mecanismos de regulación de la fosforilación oxidativa y a las adaptaciones inducidas por el entrenamiento en este proceso (Korzeniewski and Zoladz, 2003).
Diversos estudios (Hagberg et al., 1975, Weltman and Katch, 1976, Whipp and Wasserman, 1972) han mostrado que los individuos entrenados exhiben una cinética del VO2 mucho más rápida que los individuos no entrenados. En un estudio llevado a cabo por Norris & Petersen (1998) se observó una aceleración de la cinética del VO2 en atletas previamente entrenados. En dicho estudio, se observó una reducción de 5 s en la τp de la pVO2 luego de las primeras 4 semanas de entrenamiento (5 sesiones x 40-55 min), mientras que se observó una reducción adicional de 3 s luego de las segundas 4 semanas de entrenamiento. Otros estudios en este campo (Phillips et al., 1995) han mostrado que el/los mecanismo/s responsable/s del acortamiento de la cinética del VO2 se encuentra/n activado/s en las primeras etapas (i.e., < 4 semanas) del entrenamiento de la resistencia. En base a lo establecido hasta aquí, parece claro que los mecanismos responsables de la regulación/aceleración de la cinética del VO2, al menos en las primeras etapas del entrenamiento, se encuentran dentro de la fibra muscular. En este sentido, es aparente que la aceleración de la cinética del VO2 en las primeras etapas del entrenamiento puede estar asociada con la mejora simultánea de la estabilidad absoluta y relativa de la [ADPLIBRE] y de la relación [PCr]/[Cr] con una [ADPLIBRE] de reposo constante (Zoladz et al., 2006) y esto podría alcanzarse a través del incremento en el número/volumen mitocondrial o a través de otro mecanismo regulatorio tal como la regulación simultánea/activación paralela de la producción y utilización de ATP.
Está bien documentado que las adaptaciones metabólicas el entrenamiento de la resistencia incluyen el incremento en las enzimas mitocondriales implicadas en la oxidación de carbohidratos y ácidos grasos (Holloszy, 1967, Holloszy et al., 1970) así como también el incremento en el tamaño y número de mitocondrias (Holloszy and Booth, 1976, Hoppeler et al., 1973); y que el contenido mitocondrial puede incrementarse un 30-100% en aproximadamente 4-6 semanas de entrenamiento (Henriksson and Reitman, 1977). Sin embargo, en diversos estudios experimentales (Constable et al., 1987, Green et al., 2000, Green et al., 1991) se ha observado que el entrenamiento/acondicionamiento no provocó cambios en la [ADPLIBRE] de reposo (una de las condiciones establecidas para la aceleración de la cinética del VO2). Asimismo, parece que un entrenamiento de corto plazo (de unos pocos días a un mes de duración) que derive en un incremento significativo en la estabilidad metabólica, pero no en cambios detectables en la cantidad y actividad de las proteínas mitocondriales, tampoco afecta la [ADPLIBRE] (Green et al., 2000, Phillips et al., 1995). Por otra parte, si el incremento inducido por el entrenamiento en el volumen/actividad mitocondrial estuviera asociado con una reducción de la [ADPLIBRE], no daría cuenta de una mejora significativa en la estabilidad relativa para la [ADPLIBRE] y la [PCr]/[Cr] y además la cantidad de mitocondrias no se incrementa significativamente en las etapas tempranas del entrenamiento y por lo tanto no puede ser responsable del incremento temprano en la estabilidad metabólica. Estas limitaciones no se aplican al incremento inducido por el entrenamiento en la activación paralela de la demanda y suministro de ATP, lo cual puede mejorar la estabilidad absoluta y relativa en la [ADPLIBRE] y la [PCr]/[Cr], lo cual podría acelerar la cinética del VO2 sin provocar cambios en la [ADPLIBRE] de reposo, y es probable que este proceso tenga lugar en las primeras etapas del entrenamiento (Korzeniewski and Zoladz, 2003, Korzeniewski and Zoladz, 2004). Este mecanismo no requiere la síntesis de cantidades significativas de proteínas (regulación a nivel genético), que por otra parte requeriría el incremento en el número de mitocondrias.
Consumo de Oxígeno Pulmonar vs. Muscular
Para investigar con precisión la bioenergética muscular y el control metabólico durante el ejercicio, es vital cuantificar en forma precisa la tasa de “turnover” energético dentro del músculo activo. La medición directa de los eventos metabólicos dentro del músculo (e.g., mediante biopsia muscular) provee un una excelente indicación de la energética muscular. Sin embargo, este método solo provee una “instantánea” tomada en un punto temporal dado. La utilización de técnicas tales como la Resonancia Magnética con Fósforo 31 (31P-MRS) ha permitido capturar dichos eventos (Rossiter et al., 1999) y ha mostrado que los cambios en la concentración de fosfocreatina ([PCr]) provee una imagen especular del consumo de oxígeno muscular (mVO2) (Grassi et al., 1996). Sin embargo, dichos estudios son costosos y solo pueden realizarse mediciones en grupos musculares aislados (Jones et al., 2008). La medición de la cinética pulmonar del VO2 (pVO2) es una alternativa a los métodos mencionados previamente, proveyendo una estimación de la mVO2 (Whipp and Ward, 1992).
Mediante la utilización de modelos computarizados, Barstow et al., (1990) demostraron que la constante de tiempo (τ) de la Fase II de la pVO2 podría proveer una aproximación a la τ de la mVO2. Grassi et al., (1996) confirmaron esta aproximación teórica demostrando que la pVO2 es esencialmente una “imagen especular” de la reducción exponencial en la [PCr] durante el mismo período de tiempo, mostrando un tiempo medio similar para la pVO2 (25.5 s) y la mVO2 (27.9 s). En la Figura 4 puede apreciarse este fenómeno.
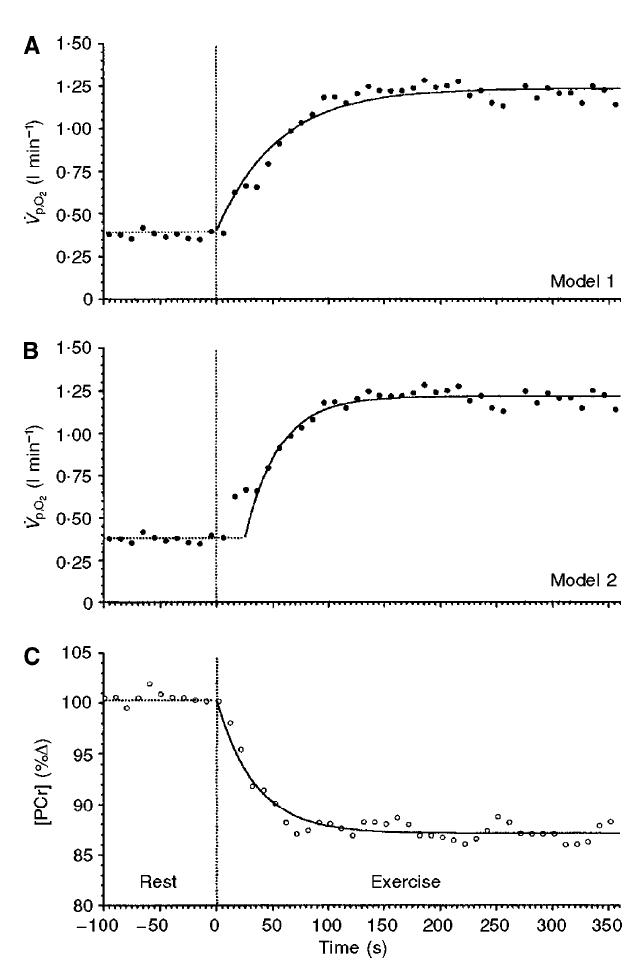
Figura 4. Respuestas simultáneas del consumo de oxígeno y de la concentración de fosfocreatina (PCr) en respuesta al ejercicio de extensiones de rodilla de moderada intensidad (Rossiter et al., 1999). (A) La respuesta del VO2 fue modelada sin un retraso temporal (Modelo 1). (B) La misma respuesta fue modelada con un retraso temporal (Modelo 2). En (B) se observa claramente el componente cardiodinámico de la cinética del VO2. Además, la respuesta de la cinética del VO2 utilizando el Modelo 2 (Fase II) es idéntica a la cinética de utilización de la PCr al comienzo del ejercicio (Panel C).
Rossiter et al., (1999) proporcionaron evidencia adicional utilizando 31P-MRS para demostrar que la medición simultánea, no solo de la pVO2, sino también de la degradación de PCr durante extensiones de rodilla de intensidad moderada poseen características similares (Figura 4). Este estudio además mostró que existe un tiempo de retraso entre los eventos que ocurren en el músculo y aquellos que se miden a nivel pulmonar (i.e., Fase I) y además que existe una simetría entre el incremento en la pVO2 y la declinación de la [PCr] luego de la remoción de la Fase I (Rossiter et al., 1999). La comparación de mediciones directas (Grassi et al., 1996, Koga et al., 2005) e indirectas (Rossiter et al., 1999) indica que una medición cuidadosa y un análisis preciso de la constante de tiempo τ de la Fase II de la pVO2 parece reflejar estrechamente (dentro de un 10%) la observada a nivel muscular (Barstow et al., 1990, Grassi et al., 1996, Poole et al., 1992). Por lo tanto, el vínculo aparente entre los eventos que ocurren en el músculo (i.e., mVO2) y aquellos medidos a nivel pulmonar (i.e., pVO2) resaltan la importancia de la medición e interpretación de la cinética pVO2 como un método no invasivo de la energética muscular.
Utilización de Diferentes Parámetros
Si bien la completa caracterización de la cinética del VO2 implica la utilización de diversos parámetros del retraso temporal y de las amplitudes de la respuesta para las fases relevantes (Barstow et al., 1996), los parámetros más relevantes para describir la tasa de incremento en el VO2 son la constante de tiempo del componente primario (τp) o el tiempo medio de la respuesta total (t½). La τp representa el tiempo para alcanzar [1 · 1/e] × 100% = 63% de la respuesta final del VO2 durante la Fase II de la transición reposo-trabajo. Por otra parte, el t½ indica el tiempo necesario para alcanzar el 50% de la respuesta final del VO2 durante la transición reposo-trabajo. Cuando se analizan los datos de la literatura respecto de la cinética del VO2 se deben tener en cuenta los diferentes significados de los parámetros cinéticos medidos en los músculos activos (Bangsbo et al., 2000, Grassi et al., 2005) y en los pulmones (i.e., pVO2); aunque como hemos mencionado previamente, el valor de τp que caracteriza el componente primario de la cinética pulmonar del VO2 refleja estrechamente la cinética del VO2 determinada en los músculos activos. La τp puede variar significativamente en humanos saludables, entre 20 y 60 segundos (Whipp et al., 2002). Generalmente en los humanos la τp está inversamente correlacionada con el consumo máximo de oxígeno (Whipp et al., 2002). Los menores valores de τp, ~ 10 s, han sido reportados en individuos bien entrenados (Barstow and Mole, 1991, Koppo et al., 2004, Zoladz et al., 2005). Por otra parte, los mayores valores de τp, ~ 70 s, han sido observados en pacientes con insuficiencia cardio-pulmonar (Sietsema, 1992, Sietsema et al., 1994). Si bien una más rápida cinética del VO2 en condiciones fisiológicas normales está asociada con una mayor capacidad física y una menor fosforilación a nivel de sustrato; recientemente se ha postulado que una mayor cinética del VO2 al inicio del ejercicio no está necesariamente asociada con una mejora en la función muscular (Korzeniewski and Zoladz, 2006).
Cinética “On” vs. Cinética “Off” del Consumo de Oxígeno
Hasta el momento hemos enfocado nuestra atención hacia lo que ocurre con la cinética del VO2 durante la transición entre el reposo y el ejercicio, lo que se denomina “VO2 on-kinetics”. Sin embargo, así como existe una cinética del VO2 para la transición reposo-ejercicio, también existe una cinética del VO2 para la transición ejercicio-reposo o cinética de la recuperación (VO2 off-kinetics) (Figura 5).
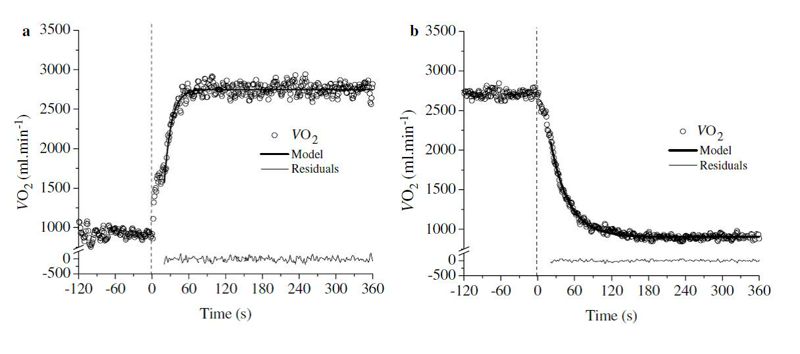
Figura 5. Modelos característicos de la cinética “on” (a) y de la cinética “off” del VO2 en el dominio de intensidad moderada. En ambos casos la cinética del VO2 se ha caracterizado utilizando un monoexponencial.
La caracterización de la cinética “off” del VO2, al igual que la cinética “on”, posee una amplitud, un tiempo de retraso y una constante temporal (τoff). Las consideraciones actuales acerca de la cinética pulmonar transitoria “on” y “off” del VO2 (i.e., Fase II) durante ejercicios de intensidad moderada (i.e., debajo del UL) sugieren que tanto la fase “on” como la fase “off” están caracterizadas por una respuesta monoexponencial y que ambas fases son simétricas (Linnarsson, 1974, Ozyener et al., 2001, Paterson and Whipp, 1991). Sin embargo, por encima del UL, la respuesta del VO2 es más compleja (Linnarsson, 1974, Hughson and Morrissey, 1982, Barstow and Mole, 1991, Paterson and Whipp, 1991). Tal como hemos establecido previamente, la cinética “on” del VO2 en los dominios supra umbrales no puede ser caracterizada por un modelo monoexponencial y al parecer tampoco existe una simetría “on-off” cuando la intensidad es mayor al UL (Linnarsson, 1974, Cerretelli et al., 1977, Paterson and Whipp, 1991, Gerbino et al., 1996, Langsetmo and Poole, 1999, Ozyener et al., 2001). La Figura 6 muestra la asimetría entre las cinéticas “on” y “off” del VO2.
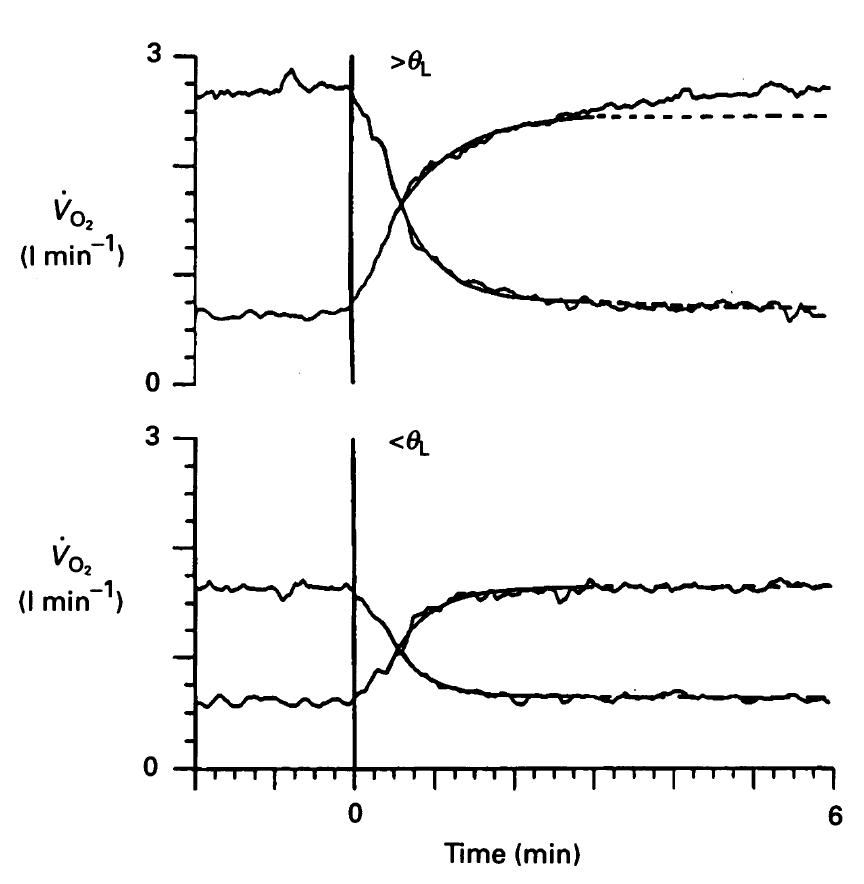
Figura 6. Respuestas “on” y “off” del VO2 a series de 6 min de trabajo a carga constante por debajo del UL (110 W, panel inferior) y por encima del UL (210 W, panel superior). Las respuestas son promedios de cuatro tests. Como puede observarse en el panel superior, en el dominio de intensidad supra umbral la respuesta dinámica del VO2 no exhibe una simetría entre las fases “on” y “off” (θL = Umbral de lactato).
Interesantemente, existe evidencia que indica además, que la misma asimetría dinámica que se produce en la cinética del VO2 también se produce en la cinética de la [PCr] e incluso que la cinética de la [PCr] puede exhibir un componente lento (Rossiter et al., 2002) (Figura 7). Esto podría tener consecuencias no solo para el rendimiento en deportes de resistencia sino también en deportes de prestación intermitente.
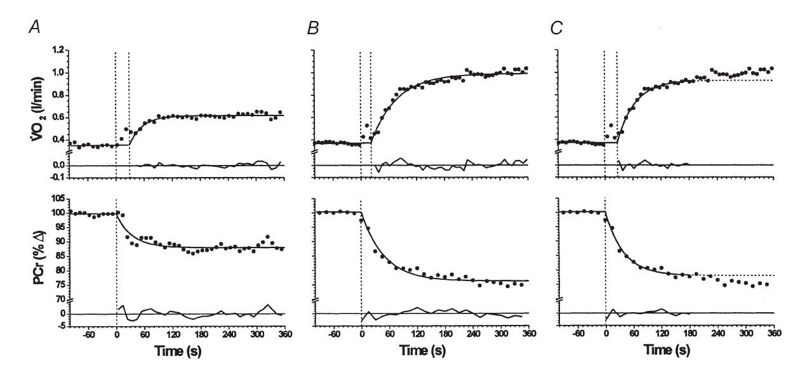
Figura 7. Respuestas del VO2 y la [PCr] mostrando los modelos y residuales. (A). Ejercicio de intensidad moderada. (B) Ejercicio de alta intensidad para el cual toda la respuesta fue modelada con un modelo monoexponencial. (C) Ejercicio de alta intensidad utilizando un modelo monoexponencial pero limitado solo a la fase fundamental.
Relación de la Cinética del VO2 con el Rendimiento Físico y la Salud
Una experiencia común para los atletas competitivos y recreacionales es que es posible mantener un ritmo rápido y a la vez confortable por un período considerable de tiempo. Sin embargo, si el ritmo se incrementa solo “un poco”, entonces el esfuerzo percibido se incrementa considerablemente, y la duración tolerable de ejercicio se reduce dramáticamente. Este efecto se ve exacerbado en los individuos que sufren de diversas condiciones patológicas. Para estos individuos, dicha experiencia puede ocurrir incluso a bajas intensidades de ejercicio, dificultando no solo su capacidad de ejercicio sino la realización de actividades cotidianas. Durante el ejercicio, la transición entre “confortable” y “no tan confortable” parece corresponderse con el límite entre los dominios de intensidad “pesada” y “severa” (Tabla 1); y su fundamento fisiológico y matemático está incluido en la relación potencia-duración (Jones et al., 2010, Monod and Scherrer, 1965, Moritani et al., 1981).
Los individuos con una mayor tolerancia al ejercicio suelen exhibir una más rápida fase primaria de la cinética del VO2 (i.e., una menor τ), y una potencia crítica (CP) situada a un mayor porcentaje de un VO2máx similarmente alto (Cerretelli et al., 1979, Jones, 2006). Este escenario minimiza el déficit de O2 al inicio del ejercicio, extiende el rango de cargas de trabajo a las cuales se puede estabilizar el componente lento, y provee un mayor rango para su desarrollo (i.e., un alto VO2máx). En contraste, los individuos desentrenados (Koppo et al., 2004, Neder et al., 2000), y particularmente aquellos que sufren de diversas condiciones respiratorias, cardiovasculares o enfermedades musculares (Mezzani et al., 2010, Neder et al., 2000, Nery et al., 1982, Sietsema et al., 1986) exhiben una cinética del VO2 más lenta e incurren en un mayor déficit de O2 al comienzo del ejercicio. Esto provoca el incremento en la tasa de utilización de glucógeno y la CP se produce a una carga de trabajo mucho menor, y el componente lento del VO2 aparece antes y solo se desarrolla hasta un VO2máx relativamente bajo. Murgatroyd et al. (2011) han propuesto que la cinética del VO2 está directamente relacionada con los parámetros de la relación potencia-duración y proveen datos de correlación para testear las siguientes hipótesis: (i) la constante temporal del componente primario de la cinética del VO2 está inversamente relacionado a la CP; y (ii) la amplitud del componente lento del VO2 está positivamente correlacionado con la capacidad anaeróbica.
Se ha sugerido que la producción de potencia al umbral de lactato está fuertemente correlacionada con el rendimiento en eventos con una duración entre 30 y 60 min. Por lo tanto, un candidato obvio para establecer la máxima potencia que puede mantenerse en este tiempo es el umbral de lactato. Sin embargo, los atletas son capaces de completar una maratón a velocidades similares o incluso mayores al umbral de lactato (Jones, 2006). Similarmente, los corredores de 10 km y de media maratón pueden competir a velocidades considerablemente mayores al umbral de lactato. No obstante, teóricamente la mayor potencia sostenible está representada por la potencia crítica y este parámetro ha sido sugerido como el límite entre los dominios de intensidad “pesado” y “severo” (Tabla 1). La potencia crítica es equivalente a la producción de potencia al máximo estado estable de lactato. En base a esto surge la siguiente pregunta ¿Qué papel desempeña la cinética del VO2 para establecer la relación potencia-duración? Al parecer la clave para responder a esta cuestión reside en la existencia del componente lento del VO2.
El rendimiento para un corredor, por ejemplo, puede representarse por la relación entre su potencia personal (velocidad) y el tiempo hasta el agotamiento (tiempo límite) (Billat et al., 1999). Existen velocidades particulares que delinean los dominios de la intensidad que están determinados por las respuestas del consumo de oxígeno (VO2) y del lactato sanguíneo en el tiempo (Whipp, 1994) y pueden ser utilizados para definir la fase lenta de la cinética del VO2 (el componente lento del VO2) que solo aparece durante el ejercicio intenso. El consumo de oxígeno se incrementa en forma mono-exponencial y se estabiliza al aproximadamente 80% en corredores de maratón de alto nivel por al menos una hora y media de ejercicio continuo. Luego de este tiempo, es posible que el consumo de oxígeno se incremente debido a cuestiones termorregulatorias, y este incremento es denominado “drift de VO2” (desviación del VO2). Esta intensidad de ejercicio corresponde a la velocidad que puede mantenerse durante una maratón y es aproximadamente igual a la velocidad asociada con el VO2máx determinada durante un test progresivo (vVO2máx) (Billat and Koralsztein, 1996). A una mayor intensidad se produce el estado estable de lactato (Brooks, 1985), cuando la tasa de aparición de lactato en sangre iguala a la tasa de desaparición. El VO2 se estabiliza luego de 3 min a aproximadamente el 85% del VO2máx. Esto corresponde a la mayor velocidad que un atleta puede sostener por una hora (85% de la vVO2máx para un atleta de resistencia bien entrenado). A una mayor intensidad, aproximadamente al 90% de la vVO2máx, la tasa de aparición de lactato en sangre excede la tasa de desaparición y la concentración de lactato en sangre se incrementa. Luego de este primer incremento monoexponencial en el VO2, se produce un segundo incremento (luego de aproximadamente tres minutos) que se define como el componente lento del VO2. El VO2 alcanza un estado estable retrasado que es mayor que el requerimiento de VO2 estimado para la relación entre el VO2 y una tasa de trabajo moderada. Por ejemplo, en este caso, un atleta puede correr al 90% de la vVO2máx y alcanzar y estabilizar el 95% del VO2máx al sexto minuto de ejercicio (el tiempo hasta el agotamiento a esta velocidad es de aproximadamente 10-15 min) (Billat, 2000). Esto se corresponde con la llamada potencia crítica. El tiempo límite a la velocidad crítica se reduce a menos de 30 minutos debido a la rápida depleción de glucógeno (Jenkins and Quigley, 1991, Housh et al., 1989). La velocidad crítica es la velocidad más alta por debajo del nivel máximo (VO2máx) al cual el consumo de oxígeno puede alcanzar un estado estable. La amplitud del componente lento representa aproximadamente el 10% del componente primario y depende de la intensidad absoluta de ejercicio (Billat, 2000). El valor del componente lento del VO2 puede alcanzar 500 mL·min-1 y en general es considerado significativo cuando su valor es mayor a los 200 mL·min-1. El componente lento puede identificarse tal como lo describieran inicialmente Whipp & Wasserman (1972), calculando la diferencia entre los valores del VO2 obtenidos entre el 6to y 3er minuto (Whipp and Wasserman, 1972) o, si el ejercicio se realiza hasta el agotamiento, entre el tercer y el último minuto (Billat et al., 1998). La aparición de este componente lento del VO2 se debe principalmente al reclutamiento de las fibras tipo II con la fatiga (Poole et al., 1991). Se ha demostrado que las fibras tipo II tienen un índice de fosfato-oxígeno que es un 18% menor que en las fibras tipo I, probablemente debido a la mayor dependencia del shuttle de α-glicerofosfato que del shuttle de malato-aspartato (Willis and Jackman, 1994). Por lo tanto se requiere más oxígeno para producir el mismo nivel de resíntesis de ATP y mantener una potencia dada. El otro 15% se debe a un incremento en el trabajo cardíaco y ventilatorio (Billat, 2000).
Por lo tanto, la tolerancia al ejercicio podría estar directa o indirectamente determinada por la cinética del VO2 en un amplio rango de duraciones de ejercicio (i.e., todas las tareas realizadas a producciones de potencia por encima del UL). En este contexto, la proximidad de la producción de potencia con el UL determina si se produce o no el componente lento del VO2. Si la producción de potencia es mayor a la del UL, entonces la proximidad de la producción de potencia a la CP determina el comportamiento del componente lento del VO2 (i.e., si este se estabiliza o no). Si la producción de potencia se encuentra por encima de la CP, entonces el VO2máx interactúa con la trayectoria del componente lento del VO2 y la magnitud de la capacidad anaeróbica determinará la duración tolerable de la tarea. Las correlaciones establecidas entre el umbral de lactato, la CP/MLSS y el VO2máx con el rendimiento de resistencia podrían verse como una consecuencia directa de la influencia de estos parámetros sobre el comportamiento de la respuesta del VO2.
Por último, cabría preguntarse si la cinética del VO2 posee alguna relación con el rendimiento físico en deportes de prestación intermitente. Como hemos señalado previamente, la cinética “off” del VO2 representa la fase de recuperación del ejercicio. Por lo tanto, se podría presuponer que una más rápida cinética de recuperación del VO2 entre repeticiones de ejercicios de alta intensidad, contribuiría a una mejor recuperación y por lo tanto a un mayor rendimiento en este tipo de actividades. En este sentido, Dupont et al. (2010) han observado que una más rápida cinética del VO2 durante la recuperación, indicada por un menor valor de τoff, se correlacionó significativamente con el rendimiento en sprints repetidos. Sin embargo, Buccheit et al. (2012a, 2012b) no han hallado correlaciones significativas entre los parámetros de la cinética “off” del VO2 y el rendimiento en sprints repetidos. En este sentido, la investigación en este campo está garantizada.
Sebastián Del Rosso
REFERENCIAS
BANGSBO, J., KRUSTRUP, P., GONZALEZ-ALONSO, J., BOUSHEL, R. & SALTIN, B. 2000. Muscle oxygen kinetics at onset of intense dynamic exercise in humans. Am J Physiol Regul Integr Comp Physiol, 279, R899-906.
BARSTOW, T. J. 1994. Characterization of VO2 kinetics during heavy exercise. Med Sci Sports Exerc, 26, 1327-34.
BARSTOW, T. J., JONES, A. M., NGUYEN, P. H. & CASABURI, R. 1996. Influence of muscle fiber type and pedal frequency on oxygen uptake kinetics of heavy exercise. J Appl Physiol, 81, 1642-50.
BARSTOW, T. J., LAMARRA, N. & WHIPP, B. J. 1990. Modulation of muscle and pulmonary O2 uptakes by circulatory dynamics during exercise. J Appl Physiol, 68, 979-89.
BARSTOW, T. J. & MOLE, P. A. 1987. Simulation of pulmonary O2 uptake during exercise transients in humans. J Appl Physiol, 63, 2253-61.
BARSTOW, T. J. & MOLE, P. A. 1991. Linear and nonlinear characteristics of oxygen uptake kinetics during heavy exercise. J Appl Physiol, 71, 2099-106.
BEHNKE, B. J., KINDIG, C. A., MUSCH, T. I., SEXTON, W. L. & POOLE, D. C. 2002. Effects of prior contractions on muscle microvascular oxygen pressure at onset of subsequent contractions. J Physiol, 539, 927-34.
BILLAT, L. V. & KORALSZTEIN, J. P. 1996. Significance of the velocity at VO2max and time to exhaustion at this velocity. Sports Med, 22, 90-108.
BILLAT, L. V., KORALSZTEIN, J. P. & MORTON, R. H. 1999. Time in human endurance models. From empirical models to physiological models. Sports Med, 27, 359-79.
BILLAT, V. L. 2000. VO2 slow component and performance in endurance sports. Br J Sports Med, 34, 83-5.
BILLAT, V. L., RICHARD, R., BINSSE, V. M., KORALSZTEIN, J. P. & HAOUZI, P. 1998. The V(O2) slow component for severe exercise depends on type of exercise and is not correlated with time to fatigue. J Appl Physiol, 85, 2118-24.
BROOKS, G. A. 1985. Anaerobic threshold: review of the concept and directions for future research. Med Sci Sports Exerc, 17, 22-34.
BROWN, G. C. 2000. Nitric oxide as a competitive inhibitor of oxygen consumption in the mitochondrial respiratory chain. Acta Physiol Scand, 168, 667-74.
BUCHHEIT, M., ABBISS, C. R., PEIFFER, J. J. & LAURSEN, P. B. 2012a. Performance and physiological responses during a sprint interval training session: relationships with muscle oxygenation and pulmonary oxygen uptake kinetics. Eur J Appl Physiol, 112, 767-79.
BUCHHEIT, M., HADER, K. & MENDEZ-VILLANUEVA, A. 2012b. Tolerance to high-intensity intermittent running exercise: do oxygen uptake kinetics really matter? Front Physiol, 3, 406.
BURNLEY, M. & JONES, A. M. 2007. Oxygen uptake kinetics as a determinant of sports performance. Eur J Sport Sci, 7, 63-79.
CASABURI, R., BARSTOW, T. J., ROBINSON, T. & WASSERMAN, K. 1989a. Influence of work rate on ventilatory and gas exchange kinetics. J Appl Physiol, 67, 547-55.
CASABURI, R., DALY, J., HANSEN, J. E. & EFFROS, R. M. 1989b. Abrupt changes in mixed venous blood gas composition after the onset of exercise. J Appl Physiol, 67, 1106-12.
CERRETELLI, P., PENDERGAST, D., PAGANELLI, W. C. & RENNIE, D. W. 1979. Effects of specific muscle training on VO2 on-response and early blood lactate. J Appl Physiol, 47, 761-9.
CERRETELLI, P., SHINDELL, D., PENDERGAST, D. P., DI PRAMPERO, P. E. & RENNIE, D. W. 1977. Oxygen uptake transients at the onset and offset of arm and leg work. Respir Physiol, 30, 81-97.
CERRETELLI, P., SIKAND, R. & FARHI, L. E. 1966. Readjustments in cardiac output and gas exchange during onset of exercise and recovery. J Appl Physiol, 21, 1345-50.
CONSTABLE, S. H., FAVIER, R. J., MCLANE, J. A., FELL, R. D., CHEN, M. & HOLLOSZY, J. O. 1987. Energy metabolism in contracting rat skeletal muscle: adaptation to exercise training. Am J Physiol, 253, C316-22.
DAVIES, C. T., DI PRAMPERO, P. E. & CERRETELLI, P. 1972. Kinetics of cardiac output and respiratory gas exchange during exercise and recovery. J Appl Physiol, 32, 618-25.
DI PRAMPERO, P. E., DAVIES, C. T., CERRETELLI, P. & MARGARIA, R. 1970. An analysis of O2 debt contracted in submaximal exercise. J Appl Physiol, 29, 547-51.
DI PRAMPERO, P. E. & MARGARIA, R. 1968. Relationship between O2 consumption, high energy phosphates and the kinetics of the O2 debt in exercise. Pflügers Archiv, 304, 11-19.
DUPONT, G., MCCALL, A., PRIEUR, F., MILLET, G. P. & BERTHOIN, S. 2010. Faster oxygen uptake kinetics during recovery is related to better repeated sprinting ability. Eur J Appl Physiol, 110, 627-34.
GERBINO, A., WARD, S. A. & WHIPP, B. J. 1996. Effects of prior exercise on pulmonary gas-exchange kinetics during high-intensity exercise in humans. J Appl Physiol, 80, 99-107.
GRASSI, B. 2003. Oxygen uptake kinetics: old and recent lessons from experiments on isolated muscle in situ. Eur J Appl Physiol, 90, 242-9.
GRASSI, B. 2005. Delayed metabolic activation of oxidative phosphorylation in skeletal muscle at exercise onset. Med Sci Sports Exerc, 37, 1567-73.
GRASSI, B. 2006a. O2 uptake kinetics in skeletal muscle: why is it so slow? And what does it mean? Adv Exp Med Biol, 578, 17-22.
GRASSI, B. 2006b. Oxygen uptake kinetics: Why are they so slow? And what do they tell us? J Physiol Pharmacol, 57 Suppl 10, 53-65.
GRASSI, B., HOGAN, M. C., KELLEY, K. M., HOWLETT, R. A. & GLADDEN, L. B. 2005. Effects of nitric oxide synthase inhibition by L-NAME on oxygen uptake kinetics in isolated canine muscle in situ. J Physiol, 568, 1021-33.
GRASSI, B., POGLIAGHI, S., RAMPICHINI, S., QUARESIMA, V., FERRARI, M., MARCONI, C. & CERRETELLI, P. 2003. Muscle oxygenation and pulmonary gas exchange kinetics during cycling exercise on-transitions in humans. J Appl Physiol, 95, 149-58.
GRASSI, B., POOLE, D. C., RICHARDSON, R. S., KNIGHT, D. R., ERICKSON, B. K. & WAGNER, P. D. 1996. Muscle O2 uptake kinetics in humans: implications for metabolic control. J Appl Physiol, 80, 988-98.
GREEN, H., TUPLING, R., ROY, B., O'TOOLE, D., BURNETT, M. & GRANT, S. 2000. Adaptations in skeletal muscle exercise metabolism to a sustained session of heavy intermittent exercise. Am J Physiol Endocrinol Metab, 278, E118-26.
GREEN, H. J., JONES, S., BALL-BURNETT, M. E., SMITH, D., LIVESEY, J. & FARRANCE, B. W. 1991. Early muscular and metabolic adaptations to prolonged exercise training in humans. J Appl Physiol, 70, 2032-8.
HAGBERG, J. M., NAGLE, F. J. & CARLSON, J. L. 1975. Transient oxygen uptake (VO2) responses at the onset of exercise. Federation Proc, 34, 443.
HENRIKSSON, J. & REITMAN, J. S. 1977. Time course of changes in human skeletal muscle succinate dehydrogenase and cytochrome oxidase activities and maximal oxygen uptake with physical activity and inactivity. Acta Physiol Scand, 99, 91-7.
HENRY, F. M. 1951. Aerobic oxygen consumption and alactic debt in muscular work. J Appl Physiol, 3, 427-38.
HENRY, F. M. & DEMOOR, J. C. 1956. Lactic and alactic oxygen consumption in moderate exercise of graded intensity. J Appl Physiol, 8, 608-14.
HOHWÜ CHRISTENSEN, E. & HÖGBERG, P. 1950. Steady-state, O2-deficit and O2-debt at severe work. Arbeitsphysiologie, 14, 251-254.
HOLLOSZY, J. O. 1967. Biochemical adaptations in muscle. Effects of exercise on mitochondrial oxygen uptake and respiratory enzyme activity in skeletal muscle. J Biol Chem, 242, 2278-82.
HOLLOSZY, J. O. & BOOTH, F. W. 1976. Biochemical adaptations to endurance exercise in muscle. Annu Rev Physiol, 38, 273-91.
HOLLOSZY, J. O., OSCAI, L. B., DON, I. J. & MOLE, P. A. 1970. Mitochondrial citric acid cycle and related enzymes: adaptive response to exercise. Biochem Biophys Res Commun, 40, 1368-73.
HOPPELER, H., LUTHI, P., CLAASSEN, H., WEIBEL, E. R. & HOWALD, H. 1973. The ultrastructure of the normal human skeletal muscle. A morphometric analysis on untrained men, women and well-trained orienteers. Pflugers Arch, 344, 217-32.
HOUSH, D. J., HOUSH, T. J. & BAUGE, S. M. 1989. The accuracy of the critical power test for predicting time to exhaustion during cycle ergometry. Ergonomics, 32, 997-1004.
HUGHSON, R. L. 2009. Oxygen uptake kinetics: historical perspective and future directions. Appl Physiol Nutr Metab, 34, 840-50.
HUGHSON, R. L. & MORRISSEY, M. 1982. Delayed kinetics of respiratory gas exchange in the transition from prior exercise. J Appl Physiol, 52, 921-9.
HUGHSON, R. L., TSCHAKOVSKY, M. E. & HOUSTON, M. E. 2001. Regulation of oxygen consumption at the onset of exercise. Exerc Sport Sci Rev, 29, 129-33.
JENKINS, D. G. & QUIGLEY, B. M. 1991. The y-intercept of the critical power function as a measure of anaerobic work capacity. Ergonomics, 34, 13-22.
JONES, A. M. 2006. The physiology for the world record holder of the women's marathon. Int J Sports Sci Coaching, 1, 101-116.
JONES, A. M. & POOLE, D. C. 2005a. Oxygen uptake dynamics: from muscle to mouth--an introduction to the symposium. Med Sci Sports Exerc, 37, 1542-50.
JONES, A. M., VANHATALO, A., BURNLEY, M., MORTON, R. H. & POOLE, D. C. 2010. Critical power: implications for determination of V O2max and exercise tolerance. Med Sci Sports Exerc, 42, 1876-90.
JONES, A. M., WILKERSON, D. P., DIMENNA, F., FULFORD, J. & POOLE, D. C. 2008. Muscle metabolic responses to exercise above and below the "critical power" assessed using 31P-MRS. Am J Physiol Regul Integr Comp Physiol, 294, R585-93.
JONES, M. A. & POOLE, D. C. 2005b. Oxygen Uptake Kinetics in Sports, Exercise and Medicine, London, Routledge
KINDIG, C. A., MCDONOUGH, P., ERICKSON, H. H. & POOLE, D. C. 2001. Effect of L-NAME on oxygen uptake kinetics during heavy-intensity exercise in the horse. J Appl Physiol, 91, 891-6.
KOGA, S., POOLE, D. C., SHIOJIRI, T., KONDO, N., FUKUBA, Y., MIURA, A. & BARSTOW, T. J. 2005. Comparison of oxygen uptake kinetics during knee extension and cycle exercise. Am J Physiol Regul Integr Comp Physiol, 288, R212-20.
KOPPO, K., BOUCKAERT, J. & JONES, A. M. 2004. Effects of training status and exercise intensity on phase II VO2 kinetics. Med Sci Sports Exerc, 36, 225-32.
KORZENIEWSKI, B. & ZOLADZ, J. A. 2003. Training-induced adaptation of oxidative phosphorylation in skeletal muscles. Biochem J, 374, 37-40.
KORZENIEWSKI, B. & ZOLADZ, J. A. 2004. Factors determining the oxygen consumption rate (VO2) on-kinetics in skeletal muscles. Biochem J, 379, 703-10.
KORZENIEWSKI, B. & ZOLADZ, J. A. 2006. Biochemical background of the VO2 on-kinetics in skeletal muscles. J Physiol Sci, 56, 1-12.
KROGH, A. & LINDHARD, J. 1913. The regulation of respiration and circulation during the initial stages of muscular work. J Physiol, 47, 112-36.
LANGSETMO, I. & POOLE, D. C. 1999. V(O2) recovery kinetics in the horse following moderate, heavy, and severe exercise. J Appl Physiol, 86, 1170-7.
LINNARSSON, D. 1974. Dynamics of pulmonary gas exchange and heart rate changes at start and end of exercise. Acta Physiol Scand Suppl, 415, 1-68.
MARGARIA, R., MANGLLI, F., CUTTICA, F. & CERRETELLI, P. 1965. The kinetics of the oxygen consumption at the onset of muscular exercise in man. Ergonomics, 8, 49-54.
MEZZANI, A., CORRA, U., GIORDANO, A., COLOMBO, S., PSAROUDAKI, M. & GIANNUZZI, P. 2010. Upper intensity limit for prolonged aerobic exercise in chronic heart failure. Med Sci Sports Exerc, 42, 633-9.
MONOD, H. & SCHERRER, J. 1965. The work capacity of a synergic muscular group. Ergonomics, 8, 329-338.
MORITANI, T., NAGATA, A., DEVRIES, H. A. & MURO, M. 1981. Critical power as a measure of physical work capacity and anaerobic threshold. Ergonomics, 24, 339-50.
MURGATROYD, S. R., FERGUSON, C., WARD, S. A., WHIPP, B. J. & ROSSITER, H. B. 2011. Pulmonary O2 uptake kinetics as a determinant of high-intensity exercise tolerance in humans. J Appl Physiol, 110, 1598-606.
NEDER, J. A., JONES, P. W., NERY, L. E. & WHIPP, B. J. 2000. Determinants of the exercise endurance capacity in patients with chronic obstructive pulmonary disease. The power-duration relationship. Am J Respir Crit Care Med, 162, 497-504.
NERY, L. E., WASSERMAN, K., ANDREWS, J. D., HUNTSMAN, D. J., HANSEN, J. E. & WHIPP, B. J. 1982. Ventilatory and gas exchange kinetics during exercise in chronic airways obstruction. J Appl Physiol, 53, 1594-602.
NOAKES, T. D. 1997. 1996 J.B. Wolffe Memorial Lecture. Challenging beliefs: ex Africa semper aliquid novi. Med Sci Sports Exerc, 29, 571-90.
NORRIS, S. R. & PETERSEN, S. R. 1998. Effects of endurance training on transient oxygen uptake responses in cyclists. J Sports Sci, 16, 733-8.
OZYENER, F., ROSSITER, H. B., WARD, S. A. & WHIPP, B. J. 2001. Influence of exercise intensity on the on- and off-transient kinetics of pulmonary oxygen uptake in humans. J Physiol, 533, 891-902.
PATERSON, D. H. & WHIPP, B. J. 1991. Asymmetries of oxygen uptake transients at the on- and offset of heavy exercise in humans. J Physiol, 443, 575-86.
PHILLIPS, S. M., GREEN, H. J., MACDONALD, M. J. & HUGHSON, R. L. 1995. Progressive effect of endurance training on VO2 kinetics at the onset of submaximal exercise. J Appl Physiol, 79, 1914-20.
POOLE, D. C., GAESSER, G. A., HOGAN, M. C., KNIGHT, D. R. & WAGNER, P. D. 1992. Pulmonary and leg VO2 during submaximal exercise: implications for muscular efficiency. J Appl Physiol, 72, 805-10.
POOLE, D. C., SCHAFFARTZIK, W., KNIGHT, D. R., DERION, T., KENNEDY, B., GUY, H. J., PREDILETTO, R. & WAGNER, P. D. 1991. Contribution of exercising legs to the slow component of oxygen uptake kinetics in humans. J Appl Physiol, 71, 1245-60.
ROSSITER, H. B., WARD, S. A., DOYLE, V. L., HOWE, F. A., GRIFFITHS, J. R. & WHIPP, B. J. 1999. Inferences from pulmonary O2 uptake with respect to intramuscular [phosphocreatine] kinetics during moderate exercise in humans. J Physiol, 518 ( Pt 3), 921-32.
ROSSITER, H. B., WARD, S. A., KOWALCHUK, J. M., HOWE, F. A., GRIFFITHS, J. R. & WHIPP, B. J. 2002. Dynamic asymmetry of phosphocreatine concentration and O(2) uptake between the on- and off-transients of moderate- and high-intensity exercise in humans. J Physiol, 541, 991-1002.
SIETSEMA, K. E. 1992. Oxygen uptake kinetics in response to exercise in patients with pulmonary vascular disease. Am Rev Respir Dis, 145, 1052-7.
SIETSEMA, K. E., BEN-DOV, I., ZHANG, Y. Y., SULLIVAN, C. & WASSERMAN, K. 1994. Dynamics of oxygen uptake for submaximal exercise and recovery in patients with chronic heart failure. Chest, 105, 1693-700.
SIETSEMA, K. E., COOPER, D. M., PERLOFF, J. K., ROSOVE, M. H., CHILD, J. S., CANOBBIO, M. M., WHIPP, B. J. & WASSERMAN, K. 1986. Dynamics of oxygen uptake during exercise in adults with cyanotic congenital heart disease. Circulation, 73, 1137-44.
STIRLING, J. R., ZAKYNTHINAKI, M. S. & SALTIN, B. 2005. A model of oxygen uptake kinetics in response to exercise: including a means of calculating oxygen demand/deficit/debt. Bull Math Biol, 67, 989-1015.
TSCHAKOVSKY, M. E. & HUGHSON, R. L. 1999. Interaction of factors determining oxygen uptake at the onset of exercise. J Appl Physiol, 86, 1101-13.
WASSERMAN, K., WHIPP, B. J. & CASTAGNA, J. 1974. Cardiodynamic hyperpnea: hyperpnea secondary to cardiac output increase. J Appl Physiol, 36, 457-64.
WELTMAN, A. & KATCH, V. 1976. Min-by-min respiratory exchange and oxygen uptake kinetics during steady-state exercise in subjects of high and low max VO2. Res Q, 47, 490-8.
WHIPP, B. J. 1994. The slow component of O2 uptake kinetics during heavy exercise. Med Sci Sports Exerc, 26, 1319-26.
WHIPP, B. J. & MAHLER, M. 1980. Dynamics of gas exchange during exercise. In: WEST, J. B. (ed.) Pulmonary Gas Exchange. New York: Academic Press.
WHIPP, B. J. & ROSSITER, H. B. 2005. The oxygen upteke kinetics. Physiological inferences from the parameters. In: JONES, A. M. & POOLE, D. C. (eds.) Oxygen Uptake Kinetics in Sports, Exercise and Medicine. London, New York: Routledge.
WHIPP, B. J., ROSSITER, H. B. & WARD, S. A. 2002. Exertional oxygen uptake kinetics: a stamen of stamina? Biochem Soc Trans, 30, 237-47.
WHIPP, B. J. & WARD, S. A. 1992. Pulmonary gas exchange dynamics and the tolerance to muscular exercise: effects of fitness and training. Ann Physiol Anthropol, 11, 207-14.
WHIPP, B. J., WARD, S. A. & ROSSITER, H. B. 2005. Pulmonary O2 uptake during exercise: conflating muscular and cardiovascular responses. Med Sci Sports Exerc, 37, 1574-85.
WHIPP, B. J. & WASSERMAN, K. 1972. Oxygen uptake kinetics for various intensities of constant-load work. J Appl Physiol, 33, 351-6.
WILLIS, W. T. & JACKMAN, M. R. 1994. Mitochondrial function during heavy exercise. Med Sci Sports Exerc, 26, 1347-53.
ZOLADZ, J. A., KORZENIEWSKI, B. & GRASSI, B. 2006. Training-induced acceleration of oxygen uptake kinetics in skeletal muscle: the underlying mechanisms. J Physiol Pharmacol, 57 Suppl 10, 67-84.
ZOLADZ, J. A., SZKUTNIK, Z., DUDA, K., MAJERCZAK, J. & KORZENIEWSKI, B. 2005. Preexercise metabolic alkalosis induced via bicarbonate ingestion accelerates Vo2 kinetics at the onset of a high-power-output exercise in humans. J Appl Physiol, 98, 895-904.

La Cinética del Consumo de Oxígeno por Sebastián Del Rosso se encuentra bajo una Licencia Creative Commons Atribución-NoComercial-SinDerivadas 3.0 Unported.
Basada en una obra en http://g-se.com/es/evaluacion-deportiva/blog/la-cinetica-del-consumo-de-oxigeno.