
La Influencia de Diferentes Tiempos de Recuperación Pasiva entre Sprints Máximos Repetidos en la Remoción de Lactato y el Rendimiento
Moisés Diego Germano1,2, Marcio Antonio Gonsalves Sindorf1, Alex Harley Crisp1, Tiago Volpi Braz1, Felipe Alves Brigatto1, Rafael Sakai Zaroni1, Diego Ferrari Cartarozzi2, Luis Gustavo da Cunha2, Alexandre Lopes Evangelista4 y Charles Ricardo Lopes1,3
1Grupo de Investigación en Rendimiento Humano, Universidad Metodista de Piracicaba (UNIMEP), Piracicaba, São Paulo, Brasil
2Universidad Amparense, Amparo, SP, Brasil
3Facultad Adventista de Hortolândia (UNASP), Hortolândia. São Paulo
4Universidad de Nove de Julho, Departamento de Educación, São Paulo, Brasil
Artículo publicado en el journal PubliCE del año 2018.
Publicado 13 de abril de 2018
Resumen
Palabras clave: Recuperación Pasiva, Rendimiento, Entrenamiento
INTRODUCCIÓN
El entrenamiento intervalado de alta intensidad (HIIT) implica sprints máximos repetidos o supra-máximos separados por diferentes períodos de recuperación. El HIIT tiene la capacidad de inducir diversos cambios fisiológicos que resultan en ajustes y adaptaciones neuromusculares, moleculares, metabólicas, cardiorrespiratorias y de rendimiento (3,7,8,13,20). La magnitud de estas respuestas adaptativas depende de la manipulación de las variables agudas que guían el proceso de entrenamiento continuo, como el volumen, el ejercicio, la intensidad y el tiempo de recuperación entre los sprints (7,13).
El trabajo contráctil continuo durante los sprints promueve cambios en la homeostasis bioquímica intramuscular y cardiorrespiratoria con un aumento en la producción de metabolitos musculares. Como resultado, es probable que las perturbaciones fisiológicas sean responsables de la disminución en el rendimiento atlético durante los sprints máximos repetidos (26). Por lo tanto, es razonable especular que los diferentes tiempos de recuperación pueden influir en la magnitud de los trastornos metabólicos, como la resíntesis del suministro de energía, las propiedades sinápticas y la oxidación del lactato post- ejercicio (7).
La capacidad de recuperarse de una actividad y, luego, reproducir el rendimiento de la actividad es muy importante en varias modalidades deportivas, lo que es de considerable interés para los atletas y los técnicos (24). La bibliografía ha demostrado que los períodos cortos de recuperación entre los sprints subsiguientes influyen significativamente en la disminución del rendimiento. Este resultado se debe a una resíntesis incompleta de fosfocreatina (PCr) y a la acumulación de iones de hidrógeno (H+) (2,18). Por otro lado, largos períodos de recuperación se han asociado con el mantenimiento y/o mejora de los parámetros de rendimiento debido a una mayor capacidad para resintetizar los sustratos energéticos principales, así como el mantenimiento del pH intra y extracelular y la función mejorada de los sistemas buffer musculares (13,18,22).
Por lo tanto, el propósito de este estudio fue evaluar los efectos de diferentes tiempos de recuperación pasiva en el comportamiento metabólico y el rendimiento de sprints máximos repetidos en la prueba de Wingate.
MÉTODOS
Sujetos
El número de sujetos se determinó usando los datos de la prueba de Wingate del presente estudio, basados en el tamaño del efecto (d) de 1,04, nivel alfa de 0,05 y una potencia de (1-β) de 0,80 (9). Ocho hombres jóvenes físicamente activos (edad media 22 ± 3,5 años, masa corporal 73 ± 7,3 kg, altura 1,75 ± 0,1 m) involucrados en deportes intermitentes, ciclismo y handball participaron en este estudio. Los sujetos respondieron un cuestionario de historial de salud (REST-Q 76 SPORT) para descartar contraindicaciones para las pruebas antes del comienzo del protocolo experimental. Los criterios de inclusión para la participación fueron: (a) seis meses de experiencia en deportes intermitentes, individuales y/o de equipo; (b) ninguna lesión previa que pueda interferir con el estudio; y (c) actualmente no usar suplementos nutricionales. Todos los sujetos firmaron un formulario de consentimiento informado después de recibir instrucciones con respecto al protocolo experimental. El estudio fue aprobado por el comité de ética de investigación local de la Universidad Metodista de Piracicaba (protocolo 523/2010).
Diseño Experimental
El protocolo experimental consistió en 3 sesiones separadas por 1 semana cada una. Todas las sesiones se realizaron en el mismo día y la misma hora de la semana (miércoles a las 10:00 am), y por el mismo investigador para evitar posibles efectos circadianos. El intervalo entre sesiones fue de 168 horas. En este período de tiempo, los sujetos no realizaron ejercicio intenso y mantuvieron sus hábitos alimentarios normales. La primera semana se usó para familiarizar a los sujetos con la prueba de Wingate y las pruebas antropométricas. Durante la segunda semana, los sujetos fueron aleatorizados de modo que la mitad de los sujetos (n=4) se sometieron a dos sprints máximos de 30 segundos con una recuperación pasiva de 2 minutos (RP2') entre los sprints en la prueba de Wingate y la otra mitad (n=4) se sometió a dos sprints máximos de 30 segundos con recuperación pasiva de 6 minutos (RP6') entre los sprints en la prueba de Wingate. Durante la tercera semana, los sujetos realizaron dos sprints máximos más, revirtiendo el orden de las recuperaciones previas. La cinética de remoción del lactato se realizó al final del segundo sprint de cada sesión para determinar las respuestas metabólicas en los siguientes tiempos: pre y post Wingate (0, 2, 5, 10, 15, 20 y 30 min). El diseño del estudio se presenta en la Figura 1.

Figura 1. Diseño Experimental.
Mediciones Antropométricas
La masa corporal de los sujetos se midió utilizando una balanza mecánica (Welmy®) con una precisión de 100 g. Las medidas de altura se obtuvieron usando un estadiómetro (Altura Exata®) con una precisión 0,1 cm. La composición corporal se determinó mediante las medidas de espesor de pliegue cutáneo tomadas con un calibre Cescorf. Se obtuvo una evaluación de pliegues cutáneos de 3 sitios (tríceps, suprailíaco y abdomen) y se utilizó la ecuación desarrollada por Guedes (15) para estimar el porcentaje de grasa corporal. Todos los pliegues cutáneos se midieron por el mismo evaluador.
Prueba Wingate
La prueba anaeróbica de Wingate (WAnT) se realizó utilizando un cicloergómetro (Biotec® 2100, Cefise, Nova Odessa, Brasil). Los sujetos se sentaron en el ergómetro y se hicieron ajustes para garantizar una posición óptima para el ciclismo. Luego, la WAnT se realizó de acuerdo con las recomendaciones para la estandarización (8). La carga fue de 0,75 g·kg-1 por 30 seg. Todos los datos se registraron usando un software (Ergometric 6.0). Antes de la prueba, los sujetos realizaron un calentamiento en el cicloergómetro durante 5 minutos (sin carga). Después de un intervalo de descanso de 5 minutos, se inició la prueba. Se determinó la potencia relativa máxima (máxima alcanzada), la potencia relativa media (valor medio durante 30 segundos) y el índice de fatiga [(potencia relativa máxima - potencia relativa mínima) / potencia relativa máxima] x 100].
Análisis de Lactato
Se recogieron muestras de sangre (25 μl) de las yemas de los dedos en tubos capilares heparinizados y se transfirieron a microtubos que contenían 50 μL de fluoruro de sodio al 1%. La concentración de lactato se analizó mediante un método electro enzimático con un analizador de lactato (YSI 2300 Stat Analyzer, Yellow Springs Instruments, Yellow Springs, OH, EEUU). Las concentraciones de lactato sanguíneo se expresaron en mM.
Análisis Estadísticos
La normalidad y la homogeneidad de las varianzas de datos se confirmaron mediante la prueba de Shapiro-Wilk y la prueba de Levene, respectivamente. Se utilizó un ANOVA (2 x 8) de medidas repetidas con factores de tiempo de pausa (RP2' y RP6') y condición pre- y post-Wingate (0, 2, 5, 10, 15, 20 y 30 min) para verificar las diferencias significativas para las variables de lactato. Se realizó una comparación porcentual de los valores máximos de lactato (Lacmáx) al final de las dos pausas. El t test se usó para determinar diferencias significativas en el rendimiento de la prueba de Wingate (Potenciamáx, Potenciamedia, Potenciamínima e Índice de Fatiga a Watts·kg-1). Se empleó un análisis de Cohen (22) para medir el tamaño del efecto (d). Los valores de referencia para individuos físicamente activos son: <0,35 trivial; 0,35 – 0,80 pequeño; 0,80 – 1,50 moderado y >1,50 grande. Se utilizó una significación estadística (α) del 5% para todas las pruebas estadísticas, utilizando el software SPSS versión 18.0.
RESULTADOS
No se observaron diferencias significativas en la comparación de Potenciamáx (Watts·kg-1) intra y entre las recuperaciones (P=0,05) (1° sprint de RP2'= 9,63 ± 0,92 vs. 2° sprint de RP2' = 8,93 ± 1,11 - d 0,76 - Δ% = 7,2; 1º sprint de RP6'= 9,66 ± 0,94 vs. 2º sprint de RP6' = 9,08 ± 1,06 - d 0,61 - Δ% = 6,0). Además, no hubo diferencias significativas en el índice de fatiga (%) entre la comparación intra y entre recuperaciones (1º sprint de RP2'= 41,5 ± 13,7 vs. 2° sprint de RP2' = 47,7 ± 9,51 - d 0,46 - Δ% = 15,4; 1º sprint de RP6'= 36,7 ± 3,57 vs. 2° sprint de RP6' = 41,3 ± 8,12 - d 1,28 - Δ% = 12,5).
Por otro lado, se observó una diferencia significativa en la comparación intra-recuperaciones de Potenciamedia (Watts·kg-1) (P=0,01) (1° sprint de RP2'= 7,25 ± 0,67 vs. 2° sprint de RP2' = 5,99 ± 0,67 - d 1,88 - Δ% = 17,3; 1º sprint de RP6'= 7,36 ± 0,7 vs. 2° sprint de RP6' = 6,66 ± 0,83 - d 1,08 - Δ% = 10,3). Además, se observó una diferencia significativa en la Potenciamedia (Watts·kg-1) al comparar recuperaciones (P=0,01) (RP2'= 2° sprint = 5,99 ± 0,67 vs. 2° sprint de RP6' = 6,66 ± 0,83 - d 0,91) (Figura 2a). En la Potenciamínima (Watts·kg-1) se observó una diferencia significativa en la comparación intra-recuperaciones (P=0,01) (1° sprint de RP2'= 5,58 ± 1,21 vs. 2° sprint de RP2' = 4,62 ± 0,84 - d 0,79 Δ% = 17,2). La Tabla 1 muestra todos los datos, y la Figura 2 ilustra las diferencias estadísticas en el rendimiento (Figura 2b).
La Figura 3 muestra las concentraciones de lactato en sangre (mM·L-1) después de las sesiones de HIIT con diferentes tiempos de recuperación pasiva durante 30 minutos de recuperación. No se observaron cambios significativos en las concentraciones entre recuperaciones después del HIIT (P=0,05). Sin embargo, ambas recuperaciones mostraron cambios significativos en las concentraciones sanguíneas de lactato en todo momento en relación con sus respectivos valores de reposo (P=0,01). Además, no se observaron cambios significativos en el valor total (Δ mM·L-1) de lactato en sangre entre las recuperaciones (RP2' 86,9 ± 10,8 vs. RP6' 86,0 ± 17,9) (P=0,05) y el valor máximo (RP2' = 17,01 vs. RP6' = 17,88).
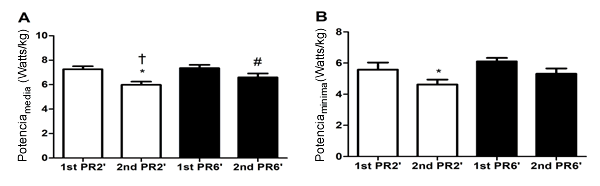
Figura 2. Valores de Media y Desviación Estándar de Potenciamedia y Potenciamínima (Watts·kg-1) de los Dos Sprints Máximos con Diferentes Tiempos de Recuperación Pasiva. *Diferencia significativa entre el 1° sprint vs. el 2° sprint de RP2' (P=0,01). # = Diferencia significativa entre el 1° sprint vs. el 2° sprint de RP6' (P=0,01). † = Diferencia significativa entre el 2° sprint de RP2' vs. el 2° sprint de RP6' (P=0,01)
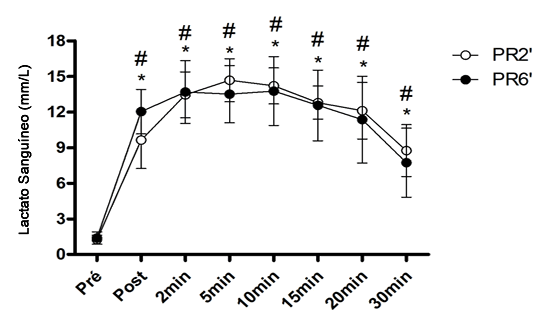
Figura 3. Media y Desviación Estándar de las Concentraciones de Lactato Sanguíneo (mM·L-1). No se observaron cambios significativos entre las dos recuperaciones pasivas (P=0,05). *Diferencia significativa en relación con el pre RP2'. # = Diferencia significativa en relación con el pre RP6'
DISCUSIÓN
El propósito del presente estudio fue verificar de forma aguda las influencias de los diferentes tiempos de recuperación pasiva en el HIIT sobre los parámetros metabólicos y de rendimiento. La manipulación de la recuperación entre sprints parece ser una clave variable con la capacidad de interferir en la magnitud de las respuestas y ajustes neuromusculares, metabólicos, moleculares y de rendimiento (3,7,8,13,20). La mayoría de los estudios (11,17-19,25) investigaron la influencia de la recuperación mediante sprints a corto plazo (3 a 10 segundos) y comparando recuperaciones pasivas y activas.
La bibliografía (10,16) ha descrito algunos de los principales aspectos de la recuperación muscular intracelular en el rendimiento de sprints repetidos, como la resíntesis de fosfocreatina (PCr), la remoción de iones de hidrógeno (H+), la restitución del gradiente de potasio (K+), la regulación de fosfato inorgánico (Pi) y la oxidación de lactato. En este sentido, varios estudios (9,11,13,18,22,25) han demostrado que, en la comparación entre recuperaciones pasivas y activas, las recuperaciones pasivas presentan una mayor capacidad en el mantenimiento del rendimiento debido a la mayor restauración metabólica intracelular. Sin embargo, pocos estudios (13,18,22) han analizado el impacto de los diferentes tiempos de recuperación pasiva durante los sprints repetidos en los cambios metabólicos y de rendimiento.
En el presente estudio, no se encontraron diferencias significativas en la Potenciamáx (P=0,05), que representa los primeros segundos del sprint, y, en consecuencia, el suministro energético a través de la PCr, dejando en claro que ambos tiempos de recuperación fueron suficientes en su resíntesis. Este hallazgo está de acuerdo con los resultados obtenidos en otros estudios (5,10,14,16). Por otro lado, ambas recuperaciones (RP2' y RP6') mostraron una disminución significativa en la Potenciamedia entre el 2° sprint y el 1° sprint (P=0,01), con un efecto grande d 1,88 en RP2' y un efecto moderado d 1,08 en RP6'. Estos valores de d corroboran la comparación entre recuperaciones (P<0,01) en la Potenciamedia, que presentan una disminución más pronunciada entre el 2° sprint de RP2' en relación con el 2° sprint de RP6' con un efecto moderado de (d 0,91), posiblemente debido al menor tiempo de recuperación que genera mayores alteraciones metabólicas y una posterior disminución en el rendimiento (9,11,13,18,21-23,25).
La Potenciamedia representa el momento de mayor contribución de la glucólisis anaeróbica, debido al gran reclutamiento de fibras musculares inherentes a los sprints y, en consecuencia, a una mayor demanda de energía. Por lo tanto, el presente estudio evaluó el comportamiento metabólico del HIIT con diferentes recuperaciones pasivas después de la sesión de ejercicio (período de recuperación, cinética de remoción del lactato sanguíneo). Los resultados mostraron cambios significativos en las comparaciones intra-recuperación entre los valores iniciales y durante el período de recuperación post-sesión (P=0,01). Además, ambos tiempos de recuperación tuvieron altos picos de concentración de lactato en sangre (RP2' = 17,01 mM·L-1 y RP6' = 17,88 mM·L-1) y valor de lactato total (Δ) (RP2' = 86,9 ± 10,8 mM·L-1 y RP6' = 86,0 ± 17,9 mM·L-1). Sin embargo, no se encontraron cambios significativos entre las pausas (P=0,05). Las concentraciones de lactato sanguíneo en ambos tiempos de recuperación indican una gran contribución de la vía glucolítica en la producción de energía, debido al gran trabajo de la maquinaria contráctil en los sprints repetidos. A partir de esto, es posible explicar la disminución del rendimiento en ambos tiempos de recuperación, y las grandes concentraciones de lactato se acompañan de grandes concentraciones de H+, directamente responsables de la disminución del pH intracelular y los niveles de fuerza y rendimiento (5). Aunque los metabolitos inherentes a la recuperación muscular intracelular no se han medido, varios estudios (5,7,10,13,16,18,22) están de acuerdo con los resultados del presente estudio.
La bibliografía (7,11-13,18,22,25) ha sugerido que la recuperación pasiva responde al mantenimiento del rendimiento. Sin embargo, en el presente estudio ambos tiempos de recuperación no fueron suficientes para mantener la Potenciamedia, lo que sugiere que no existe una relación de causa y efecto entre el aumento de lactato en sangre y el rendimiento disminuido. Estos resultados corroboran con el estudio de Bogdanis et al. (5) que no hay una relación de causa y efecto entre el lactato y el rendimiento con diferentes recuperaciones pasivas durante los sprints máximos de 30 segundos.
Toubekis et al. (26) compararon diferentes tiempos (45 seg y 120 seg) y tipos de recuperaciones (activa x pasiva) en sprints máximos de corta duración (8 x 25 m) en parámetros metabólicos y rendimiento en natación. Los resultados mostraron que la recuperación pasiva promovió un mayor mantenimiento del rendimiento a lo largo de los sprints, mientras que las dos pausas activas presentaron un aumento significativo en el tiempo para completar los sprints. Sin embargo, en la comparación entre las dos recuperaciones pasivas, los autores no observaron diferencias. En el comportamiento metabólico, se evidenciaron las recuperaciones pasivas, aunque mostraron una mayor concentración de lactato durante los sprints repetidos en la natación, se mostró un mayor mantenimiento del rendimiento de los sprints repetidos, lo que podría explicar los diferentes hallazgos en su estudio que utilizó sprints repetidos que duraron hasta 15 segundos en la natación; mientras que el presente estudio utilizó un ejercicio de bicicleta durante 30 segundos, así como las diferencias metodológicas en la manipulación de las sesiones de ejercicio.
Germano et al. (13) analizaron las influencias de forma aguda en diferentes tiempos de recuperación (2 min y 8 min) y diferentes tipos de recuperación (activa x pasiva), analizando los parámetros cardiorrespiratorios, metabólicos y el rendimiento en el HIIT. Las sesiones de entrenamiento se formaron por 5 sprints máximos en la cinta caminadora hasta el tiempo límite (Tlim) con diferentes modelos de recuperación. Los resultados mostraron que la recuperación pasiva larga (8 min) fue responsable de mantener el rendimiento durante los 5 sprints repetidos, mientras que la recuperación pasiva corta (2 min) y las dos recuperaciones activas (2 min y 8 min) indujeron una disminución significativa en el rendimiento. Estos resultados son consistentes con los hallazgos actuales, aunque ambos estudios utilizaron recuperaciones, encontraron una disminución significativa en la Potenciamedia, la recuperación de 2 minutos mostró una mayor disminución de la magnitud en comparación con la recuperación de 6 minutos, posiblemente porque un tiempo de recuperación pasiva más largo resulta en una mejor restauración intracelular, resíntesis de los sustratos energéticos principales y menor saturación de oxihemoglobina.
Por otro lado, en el mismo estudio de Germano et al. (13), la comparación no mostró diferencias entre las dos recuperaciones pasivas. La explicación de tales hallazgos puede basarse en el pico de valores de lactato sanguíneo que fue 11,5 ± 3,4 mM·L-1 para la recuperación pasiva corta y 10,7 ± 2,0 mM·L-1 para la recuperación pasiva larga, lo que indica que los sprints repetidos hasta el Tlim proporcionan una mayor fosforilación oxidativa, mientras que, el presente estudio manipuló sprints repetidos de 30 segundos que requirieron menos contribución aeróbica. Además, no se encontraron diferencias significativas en las concentraciones y la remoción del lactato entre ninguna de las recuperaciones, lo que demuestra que no hubo relación de causa y efecto entre el lactato sanguíneo y el rendimiento. Este hallazgo también concuerda con el presente estudio.
Bret et al. (6) investigaron la capacidad de remoción de lactato y la relación con el rendimiento en corredores de élite con diferentes especialidades, lo que requiere características metabólicas específicas. Treinta y cuatro corredores de alto nivel (velocistas y distancia media) se dividieron en dos grupos según sus especialidades (100 a 400 m y 800 a 1500 m). Todos los sujetos realizaron 1 minuto de carrera a 25,2 km·h-1 seguido de una recuperación pasiva de 90 minutos para analizar la curva de remoción del lactato. Los corredores de distancia media exhibieron una mejor capacidad de remoción de lactato en comparación con los corredores de distancia corta. Los corredores que presentaron el mayor rendimiento fueron los que tuvieron la mayor capacidad de remoción de lactato. Se puede especular que las respuestas metabólicas son diferentes para atletas de élite, sujetos físicamente activos y posiblemente para individuos sedentarios. Estas diferencias pueden estar relacionadas con el contenido y la eficacia de las proteínas implicadas en la regulación del pH y la remoción de lactato.
Los resultados metabólicos obtenidos en el presente estudio se refieren a sujetos físicamente activos, y posiblemente a un menor contenido de proteínas responsables de la remoción de lactato y metabolitos. La Potenciamínima mostró una disminución significativa solo en RP2' del 2° sprint comparado con su respectivo 1° sprint (P=0,05 - d 0,80 moderado). Este resultado puede explicarse debido a un menor tiempo de recuperación que induce una menor magnitud de la restauración metabólica, lo que implica una menor utilización de la vía aeróbica en el suministro de energía (alcanzando y manteniendo un alto porcentaje de VO2máx). Esto es diferente de la RP6', que al tener un tiempo de recuperación más largo, probablemente fue capaz de inducir una mayor resíntesis de energía y mantenimiento de un alto porcentaje de VO2máx.
Limitaciones de este Estudio
El presente estudio puede haber presentado una limitación por no haber medido las respuestas cardiorrespiratorias asociadas con las respuestas metabólicas y de rendimiento. Sería interesante verificar la influencia de diferentes tiempos de recuperación pasiva en el alcance y mantenimiento de los altos porcentajes de VO2máx (1,4,22).
CONCLUSIÓN
Nuestros resultados sugieren que los dos modelos de recuperación pasiva no fueron suficientes para mantener la Potenciamedia en los sprints repetidos de la prueba de Wingate. Sin embargo, la RP6' tuvo una menor magnitud de disminución en comparación con una RP2'. Creemos que estos resultados posiblemente se deben a la gran contribución glucolítica en el suministro de energía durante los sprints repetidos, que desencadenaron grandes trastornos metabólicos, con una mayor magnitud en la RP2'. Aunque no se han observado cambios significativos en la remoción del lactato sanguíneo en ambas recuperaciones, tales alteraciones metabólicas pueden confirmarse por las grandes concentraciones de lactato observadas en las dos recuperaciones. Por lo tanto, para mantener el rendimiento en sprints repetidos con una característica glucolítica, se sugiere el uso de una recuperación pasiva larga. Sin embargo, se necesitan más estudios para determinar el potencial de adaptación frente a diferentes tiempos y tipos de recuperación entre los esfuerzos máximos en la población general.
AGRADECIMIENTOS
Agradecemos a los sujetos que dedicaron una gran cantidad de tiempo para participar en este estudio. Los autores no tienen conflictos de interés.
Dirección de correo: Charles Ricardo Lopes, FACIS -UNIMEP - Campus Taquaral Rodovia do Açúcar, Km 156, s/n, Piracicaba -SP, Brazil, Email: chrlopes@unimep.br
Referencias
1. Astrand I, Astrand PO, Christensen EH, Hedman R. (1960). Intermittent muscular work. Acta Physiol Scand. 1960;48:448-453.
2. Bangsbo J, Aagaard T, Olsen M, Klens B, Turcotte LP, Richter EA. (1995). Lactate and H+ uptake in inactive muscles during intense exercise in man. J Physiol. 1995;488:219-229.
3. Billat VL. (2001). Interval training for performance: A scientific and empirical practice: Special recommendations for middle-and long-distance running. Part I: Aerobic interval training. Sports Med. 2001;31(1):13-31.
4. Billat VL, Slawinski J, Bocquet V, Chasaing P, Demarle A, Lafitte L, Koralsztein JP. (2000). Intermittent runs at the velocity associated with maximal oxygen uptake enables subjects to remain at maximal oxygen uptake for a longer time than intense but submaximal runs. Eur J Appl Physiol. 2000;81:188-196.
5. Bogdanis GC, Nevill ME, Boobis LH, Lakomy HK. (1996). (1996). Contribution of phosphocreatine and aerobic metabolism to energy supply during repeated sprint exercise. J Appl Physiol. 1996;80(03): 876-884.
6. Bret C, Messonier L, Nouck Nouck JM, Freund H, Dufour AB, Lacour JR. (2003). Differences in lactate exchange and removal abilities in athletes specialized in different track running events (100 to 1500 m). Int J Sports Med. 2003;24(2):108-113.
7. Buchheit M, Laursen PB. (2013). High-intensity interval training, solutions to the programming puzzle. Part I: Cardiopulmonary emphasis. Sports Med. 2013;43(5):313-338.
8. Burgomaster KA, Cermak NM, Phillips SM, Benton CR, Bonen A, Gibala MJ. (2007). Divergent response of metabolite transport proteins in human skeletal muscle after sprint interval training and detraining. Am J Physiol Regul Integr Comp Physiol. 2007;292(5):1970-1976.
9. Coso JD, Hamouti N, Jimenez RA, Rodrigues RM. (2009). Respiratory compensation and blood pH regulation during variable intensity exercise in trained versus untrained subjects. Eur J Appl Physiol. 2009;107:83-93.
10. Coso JD, Hamouti N, Jimenez RA, Rodrigues RM. (2010). Restoration of blood pH between repeated bouts of high-intensity exercise: Effects of various active-recovery protocols. Eur J Appl Physiol. 2010;108:523-532.
11. Dupont G, Berthoin S. (2004). Time spent at a high percentage of VO2max for short intermittent runs: Active versus passive recovery. Can J Appl Physiol. 2004;29:3-16.
12. Dupont G, Blonde N, Berthoin S. (2003). Performance for short intermittent runs: Active recovery vs. passive recovery. Eur J Appl Physiol. 2003;89(6):548-554.
13. Germano MD, Sindorf MAG, Da Silva CE, Evangelista AL, Bocalini DS, Lopes CR. (2015). High intensity interval training: Cardiorespiratory adaptations, metabolic and performance. Inter J Sports Sci. 2015;5(6):240-247.
14. Glaister M. (2005). Multiple sprint work: Physiological responses, mechanisms of fatigue and the influence of aerobic fitness. Sports Med. 2005;35(9):757-777.
15. Guedes DP. (1985). Study of body fat through the measurement of body density and skinfold thickness in university students. Federal University of Santa Maria, Santa Maria.
16. Iaia FM, Thomassen M, Kolding H, Gunnarsson T, Wendell J, Rostgaard T, et al. (2008). Reduced volume but increased training intensity elevates muscle Na+-K+ pump alpha1-subunit and NHE1 expression as well as short-term work capacity in humans. Am J Physiol Regul Integr Comp Physiol. 2008;294(3):966-974.
17. Juel C, Klarskov C, Nielsen JJ, Krustrup P, Mohr M, Bangsbo J. (2004). Effect of high-intensity intermittent training on lactate and H+ release from human skeletal muscle. Am J Physiol Endocrinol Metab. 2004;286(2):245-251.
18. Krustrup P, Soderlund K, Mohr M, Bangsbo J. (2004). The slow component of oxygen uptake during intense, sub-maximal exercise in man is associated with additional fiber recruitment. Pflugers Arch. 2004;447(6):855-866.
19. Lopes CR. (2010). Lactate removal kinetics in the definition of pauses for high intensity interval training. State University of Campinas, Campinas.
20. Mandroukas A, Heller J, Metaxas TI, Sendelides T, Riganas C, Vamvakoudis E, Chistoulas K, Stefanidis P, Karagiannis V, Kyparos A, Mandroukas K. (2011). Cardiorespiratory and metabolic alterations during exercise and passive recovery after three modes of exercise. J Strength Cond Res. 2011;25(6):1664-1672.
21. McKenna MJ, Hargreaves M. (2008). Resolving fatigue mechanisms determining exercise performance: Integrative physiology at its finest! J Appl Physiol. 2008;104(1):286-287.
22. Millet GP, Candau R, Fattori P, Bignet F, Varray A. VO2 (2003). Responses to different intermittent runs at velocities associated with VO2max. Can J Appl Physiol. 2003;28:410-423.
23. Rhea MR. (2004). Determining the magnitude of treatment effects in strength training research through the use of the effect size. J Strength Cond Res. 2004;18(4):918-920.
24. Siri WE. (1993). Body composition from fluid spaces and density: Analysis of methods. Nutrition. 1961;9(5):480-491; Discussion, 92.
25. Thevenet D, Tardieu-Berger M, Berthoin S, Prioux J. (2007). Influence of recovery mode (passive vs. active) on time spent at maximal oxygen uptake during an intermittent session in young and endurance-trained athletes. Eur J Appl Physiol. 2007;99(2):133-142.
26. Toubekis AG, Douda HT, Tokmakidis SP. (2005). Influence of different rest intervals during active or passive recovery on repeated sprint swimming performance. Eur J Appl Physiol. 2005;93(5-6):694-700.