
La Mioglobina como una Reserva de Oxígeno en el Hombre
Irma Astrand, Per-Olof Astrand, Rune Hedman y Erik Hohwü Christensen
Department of Physiology, Kungl. Gymnastika Centralinstitutet, Stockolm Ö, Suecia.
Artículo publicado en el journal PubliCE, Volumen 0 del año 1960.
Publicado 19 de noviembre de 2009
Resumen
Palabras clave: oxígeno, bicicleta ergométrica, intermitente, VO<sub>2</sub>
INTRODUCCION
En una investigación reciente relacionada al trabajo intenso intermitente [2160 kpm.min-1) (350 W)], fue demostrado que la concentración de ácido láctico permanece baja si el trabajo se alterna con períodos de recuperación cada medio minuto. Si los períodos de trabajo y pausa se incrementan hasta 3 min, la concentración de ácido láctico va a alcanzar valores altos y el tiempo de trabajo total va a estar limitado por el agotamiento (Astrand et al, 1960).
Una explicación posible para el metabolismo del trabajo aeróbico cuando son usados períodos de 0,5 min estuvo basada en la idea de que el oxígeno unido a la miohemoglobina juega un rol importante en el aporte de oxígeno a los músculos que trabajan en la etapa inicial del trabajo. Con el incremento de la duración del período de trabajo, la importancia relativa de esta fracción de oxígeno, disminuye, y si el transporte de oxígeno es insuficiente para la demanda local, los procesos anaeróbicos deben cubrir una cierta fracción del metabolismo de trabajo, y el ácido láctico se acumula en los músculos y la sangre.
El presente trabajo fue planteado para dilucidar el rol de la miohemoglobinca a este respecto. Para obtener resultados más concluyentes, la carga de trabajo se incrementó hasta 2520 kpm.min-1) (411 W). Además, los períodos de trabajo y pausa cambiaron en forma independiente uno del otro, de modo que pudieran ser obtenidos resultados más concluyentes en relación a la importancia de los períodos de recuperación.
METODOS
El trabajo fue realizado por un sujeto físicamente bien entrenado, R. H, en una bicicleta ergométrica. El sujeto y los métodos usados fueron los mismos que los de experimentos previos (Astrand et al., 1960). El trabajo de 2520 kpm.min-1) (411 W) corresponde a una carga de 6 kg con 70 revoluciones de pedal por minuto. El tiempo experimental total fue, de ser posible, de 30 min. Los períodos de trabajo fueron siempre de la misma duración a lo largo de todo el experimento. Los mismos duraron 10, 15, 30 o 60 s.
Los períodos de recuperación duraron desde 20 hasta 240 s (Tabla 1). Debido a la diferente duración de los períodos de recuperación, las cantidades totales de trabajo producido en los diferentes días del experimento, estuvieron entre 6000 y 38000 kpm (58,800 kJ y 372,400 kJ), y la carga de trabajo promedio para los 30 min estuvo entre 200 kpm.min-1 y 1260 kpm.min-1.
RESULTADOS
La Tabla 1 resume los experimentos realizados y presenta los valores máximos para la concentración de ácido láctico sanguíneo, la cantidad total de trabajo realizado en el experimento de 30 min y otros valores calculados de importancia para la discusión.
La cantidad total de 25200 kmp de trabajo es presentada en todas las series, I-IV, ordenados en la tabla de acuerdo a la duración del período de trabajo, y la cantidad de 15120 kpm es encontrada en tres de las secciones. Esto hace posible una comparación directa. Si el período de trabajo dura 10 segundos, 420 kpm van a ser producidos en cada ocasión de trabajo; con una duración mayor, esta cantidad se incrementa y alcanza, en 60 s, 2520 kpm. La demanda de oxígeno calculada para estas diferentes cantidades de trabajo, son presentadas en la Tabla.
Estos valores son, por supuesto, aproximados, pero aun constituyen una buena ilustración de las demandas variables, que son de 0,9 L de O2 para los períodos de 10 s y de 5,6 L para los períodos de 60 s.
Los valores calculados están basados en una eficiencia mecánica de 23,0 por ciento y un coeficiente calórico de oxígeno de 4,85.
La Figura 1 y 2 ilustra los cambios en la concentración de ácido láctico sanguíneo durante el trabajo con una cantidad total de 25200 y 15120 kpm, respectivamente. La relación entre tiempo de trabajo y recuperación es siempre 1 a 2 en la Figura 1, y 1 a 4 en la Figura 2. Cuando los períodos de trabajo cortos de 10 s fueron seguidos por pausas de 20 s, la concentración de ácido láctico fue de aproximadamente 20 mg por 100 mL (2,2 mM), mientras que en el experimento con pausas de 40 s, fue de aproximadamente 15 mg cada 100 mL (1,6 mM); esto es, en ambos casos, muy cerca del valor de reposo normal de aproximadamente 10 mg cada 100 mL (1,1 mM). Si los períodos de trabajo se prolongan hasta 30 s y las pausas hasta 60 s, uno encuentra un incremento significativo en la concentración de ácido láctico sanguíneo; después de aproximadamente 9 min, se alcanza un valor máximo de 70 mg cada 100 mL (7,7 mM) (ver Figura 1).
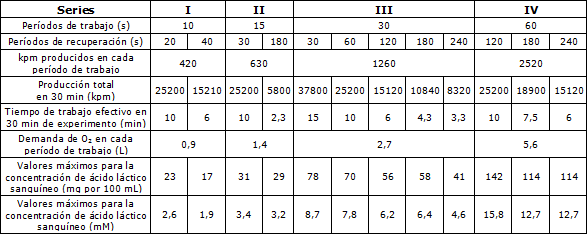
Tabla 1. Concentración pico de ácido láctico sanguíneo en los experimentos con trabajo intermitente de diferentes duraciones de los períodos de trabajo y pausa. La producción de trabajo, el tiempo de trabajo efectivo y la demanda de oxígeno, han sido calculadas y tabuladas.
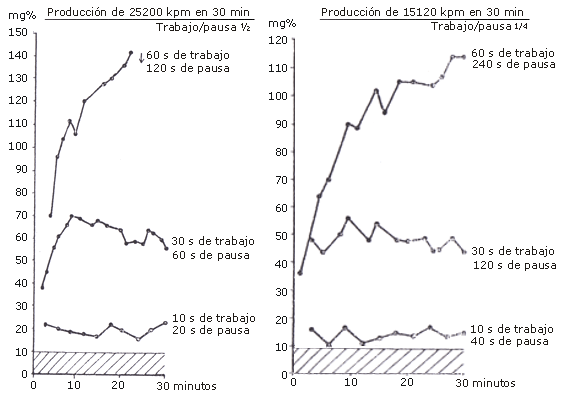
Figura 1 y 2. Concentración de ácido láctico sanguíneo a una producción de trabajo total de 1) 25200 kpm.min-1 y 2) 15120 1260 kpm.min-1 durante un tiempo experimental de 30 min. El trabajo es realizado con una carga de 2520 kpm.min-1. Los períodos de trabajo duraron 10, 30 y 60 s y los períodos de recuperación correspondientes: 1) 20, 60 y 120 s y 2) 40, 120, y 240 s.
Después de esto, hay una pequeña reducción y los valores permanecen aproximadamente en 60 mg cada 100 mL (6,6 mM) hasta el final del experimento. Con pausas más prolongadas, de 120 s, y consecuentemente, una menor cantidad de trabajo realizado (Figura 2), también se alcanza un valor máximo después de aproximadamente 9 min, ahora a 56 mg cada 100 mL (6,2 mM), y después los valores se estabilizan entre 40 (4,4 mM) y 50 mg cada 100 mL (5,5 mM). De este modo, aun con períodos de trabajo de 30 s, es alcanzado un cierto equilibrio entre la producción y la eliminación de ácido láctico.
Sin embargo, con períodos de trabajo de 60 s, el correspondiente equilibrio, nunca es alcanzado; la concentración de ácido láctico se incrementa hasta el final del experimento. Con pausas relativamente cortas de 120 s (Figura 1), la concentración de ácido láctico sanguíneo alcanza un valor máximo de 142 mg cada 100 mL (15,6 mM) después de 22 min. El experimento fue luego interrumpido, debido a que el sujeto no pudo continuar. Con pausas de 240 s (Figura 2), la tarea pudo ser completada por 30 min y la concentración de ácido láctico sanguíneo alcanzó un valor de 114 mg cada 100 mL (12,5 mM) al final.
DISCUSION
Los presentes resultados confirman los primeros hallazgos de Astrand et al. (1960). Los mismos demuestran de forma concluyente que existe una diferencia significativa durante el trabajo intermitente en la respuesta del hombre a los períodos de trabajo cortos y relativamente largos, aun si la cantidad de trabajo en un cierto tiempo es la misma. Además, los resultados responden la pregunta de si la duración del período de trabajo o de la pausa constituye el factor decisivo para el aumento de la concentración de ácido láctico sanguíneo. En la pasada serie de experimentos, tal como fue señalado arriba, la duración de los períodos de trabajo y pausa fue siempre igual en el mismo experimento, lo cual hizo difícil hacer interpretaciones concluyentes a este respecto. A partir de los resultados presentados en la Tabla 1, se puede apreciar que el factor más decisivo es la duración del período de trabajo (comparación entre las series I-IV). A partir de los resultados en la serie III, se aprecia bien que incluso la duración de la pausa tiene una cierta significancia. En estas series, el período de trabajo fue de 30 s y las pausas en los diferentes días experimentales estuvieron entre 30 y 240 s. Con una pausa de 30 s, hubo 30 ocasiones de trabajo durante la media hora, con una duración de pausa de 240 s, hubo solo 7 ocasiones de trabajo.
Menos períodos de trabajo, naturalmente disminuyen las posibilidades de producir ácido láctico, y pausas más prolongadas proporcionan mayores posibilidades de eliminación del mismo. Esto explica la disminución en la concentración de ácido láctico sanguíneo desde 78 mg cada 100 mL (8,6 mM), el cual fue el valor máximo con pausas de 30 s, hasta 41 mg cada 100 mL (4,5 mM) con pausas de 240 s. Por consiguiente, la duración de las pausas tiene una importancia secundaria en comparación a la duración de los períodos de trabajo. Esto también está ilustrado por los resultados proporcionados en la Figura 1 y 2.
Es de gran importancia determinar si estos resultados coinciden o no con la hipótesis mencionada arriba que plantea que la reacción en los períodos de trabajo cortos (10 y 15 s) y largos (30 y 60 s) puede ser explicada por el rol de la miohemoglobina como una reserva de oxígeno.
Se sabe que el transporte de oxígeno a través de la sangre hacia los músculos, se incrementa con la duración del período de trabajo, y que puede tomar varios minutos hasta que ocurra la equiparación entre los requerimientos y el aporte de oxígeno. En la serie de experimentos realizados con 10 s de trabajo y 20 s de recuperación, la captación de oxígeno fue determinada durante el período de trabajo, y correspondió a 2,8 L.min-1. Durante los 10 s de trabajo, el verdadero consumo de oxígeno fue 1/6 de esto, o 0,47 L. El consumo de oxígeno durante el trabajo fue 10 veces mayor que el valor correspondiente al reposo de 0,043 L. El consumo de oxígeno durante los últimos 30 s de un período de trabajo de 60 s con pausas de 120 s arrojó un valor máximo de 4,08 L de O2. min-1, o 0,68 L de O2 para 10 s. En el último caso, el consumo de oxígeno fue 16 veces mayor que el valor de reposo. Para que sea adecuada a una carga de 2520 kpm.min-1, se requiere un aporte de oxígeno de 5,6 L.min-1 o 0,9 L cada 10 s, de acuerdo a la Tabla 1. Durante el experimento con trabajo por 10 s y pausas de 20 s, fue calculado un déficit de 0,90 – 0,47 = 0,43 L de O2. Con trabajo por 60 s y pausas por 120 s, el consumo de oxígeno durante todo el período de trabajo fue como máximo, 3,26 L.min-1 (durante el primer medio minuto 2,43 L.min-1 y durante la segunda mitad 4,08). Esto significa un déficit de 5,60-3,26=2,34 L de O2. La razón por la que el consumo de oxígeno durante el primer medio minuto de un período de trabajo de 1 min es relativamente menor que durante los períodos de 10 s (2,43 y 2,80 L.min-1, respectivamente) es que durante las pausas cortas de 20 s, la circulación y la respiración nunca disminuyen en forma significativa antes de que el trabajo comience nuevamente. Si las pausas son prolongadas hasta 2 min, el tiempo para el ajuste se incrementa significativamente.
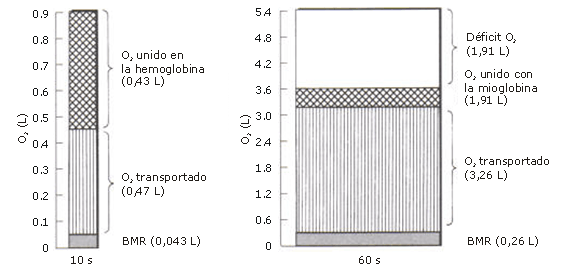
Figura 3. Demanda de oxígeno para 10 y 60 s de trabajo con una carga de 2520 kpm.min-1. Se procura ilustrar la fracción de O2 que: a) está unida a la miohemoglobina, b) es transportada por la sangre y c) el déficit de oxígeno.
Con trabajo por 10 s y pausas por 2 s, debemos asumir condiciones prácticamente aeróbicas en los músculos que trabajan. Si este no fuera el caso, los 60 períodos de trabajo deberían haber provocado una acumulación sucesiva de ácido láctico como consecuencia, en la comparación de la Figura 1.
Pensamos que como conclusión se puede plantear que 0,43 L de O2 estuvieron disponibles en los músculos que trabajan al inicio de cada nuevo período de trabajo, naturalmente, incluso en los períodos de 60 s. Cuantitativamente, esto significa que un aporte de oxígeno para períodos de 10 s es garantizado por esa cantidad, que ya está en los músculos y por la cantidad que puede ser transportada por la sangre durante el trabajo en si mismo. Para un trabajo de 60 s, se genera un déficit de 1,91 L. Esto debe ser cubierto por procesos anaeróbicos, lo cual resulta en un incremento de la concentración de ácido láctico en la sangre.
La Figura 3 ilustra esquemáticamente la importancia relativa de la cantidad de oxígeno que ha sido postulado que existe en los músculos en los períodos de trabajo de 10 y 60 s, respectivamente.
Naturalmente, el aporte de oxígeno a los músculos se vuelve más pequeño en una ocasión aislada de trabajo, y el factor anaeróbico es cuantitativamente de gran importancia. Sin embargo, si el tiempo de trabajo es suficientemente corto, la cantidad de oxígeno unido a la miohemoglobina debería ser capaz de jugar un rol decisivo para el metabolismo muscular, aun en esos casos. Una consecuencia, debe ser la re-evaluación del llamado efecto Simonsen. De acuerdo a Müller y Hettinger (1957), este efecto es importante durante los primeros 10 s de trabajo y resulta en un consumo de oxígeno extra como resultado de las condiciones anaeróbicas en los músculos.
En base a los resultados presentados aquí, la cantidad de oxígeno que está disponible al inicio del ejercicio no puede, por supuesto, ser determinada. Si se intenta calcular la cantidad de oxígeno unido a la mioglobina, uno inmediatamente confronta con una serie de más o menos factores. No se está familiarizado con el tamaño de la masa muscular activa, la concentración de miohemoglobina de los músculos o el grado de reducción de la oximiohemoglobina. Si uno usa el valor proporcionado en la literatura para la miohemoglobina, por ejemplo, por Biörck (1949, p. 131), uno encuentra que cada gramo de músculo puede unirse a aproximadamente 10 mm3 de O2. Si uno supone que hay 20 kg de músculos activos para el sujeto en el trabajo mencionado aquí, uno arriba a un valor de 200 mL de O2. Todavía hay una deficiencia de aproximadamente 230 mL, de acuerdo a lo planteado arriba. Los valores proporcionados por Biöck son en este caso, muy bajos, ya que está generalmente aceptado que la cantidad de de miohemoglobina se incrementa con el entrenamiento, y los valores proporcionados arriba no fueron derivados de individuos especialmente bien entrenados.
Es bastante evidente, a partir de las investigaciones de Scholander, Irving y Grinnell (1942), realizadas en focas que buceaban, que la miohemoglobina puede constituir un factor importante para el aporte de oxígeno a la musculatura. De acuerdo a nuestra concepción, los resultados experimentales presentados aquí, indican que la miohemoglobina tiene una función importante como reserva de oxígeno aun en el hombre. El lector es referido a Biöck (1949, p. 42), en relación a referencias a favor y en contra de tal concepción basada en hallazgos anteriores.
Antes de que una respuesta definitiva pueda ser dada a la pregunta del rol cuantitativo, que juega la miohemoglobina a este respecto, son requeridas más investigaciones acerca de la concentración de miohemoglobina en individuos entrenados.
Esperamos que los resultados experimentales relacionados aquí, ayuden a crear interés en esta área de investigación.
Referencias
1. Astrand I., P.-O Astrand, E. H. Christensen and R. Hedman (1960). Intermitent muscular work. Acta physiol. Scand., 48, 448-453
2. Scholander P. F., L. Irving and S. W. Grinnell (1942). Aerobic and anaerobic changes in muscles during diving. J. Biol. Chem.; 142, 431-440