
Metabolismo de Combustibles en Hombres y Mujeres, durante y después de un Ejercicio de Larga Duración
Tracy J Horton1, Michael J Pagliassoti1, Karen Hobbs1 y James O Hill1
Center of Human Nutrition, Department of Pediatrics, University of Colorado Health Sciences Center, Denver, Colorado 80262.
Artículo publicado en el journal PubliCE, Volumen 0 del año 1998.
Publicado 24 de marzo de 2004
Resumen
Palabras clave: género, oxidación de combustibles durante el ejercicio, catecolaminas, entrenamiento
INTRODUCCION
El metabolismo de los combustibles durante el ejercicio ha sido bien caracterizado en hombres, pero no en mujeres. Esto es relevante ya que ciertos datos sugieren que durante el ejercicio, hombres y mujeres difieren en la prioridad puesta sobre la utilización de lípidos y carbohidratos. Por ejemplo, se ha reportado que, a la misma intensidad relativa de ejercicio submáximo, las mujeres oxidan relativamente más lípidos y menos carbohidratos que los hombres (1, 25, 31). Concurrentemente, en mujeres se ha observado una menor declinación en el glucógeno muscular en respuesta al ejercicio de resistencia (31, 32). No esta claro si estas diferencias, en base al género en la oxidación corporal total de combustibles, están relacionadas a diferencias en la utilización de sustratos circulantes (1, 9, 31). Con respecto a las respuestas hormonales al ejercicio, hay alguna sugerencia de que puede haber diferencias entre hombres y mujeres (31) y, en particular, que en las mujeres, durante el ejercicio, hay un menor incremento en la concentración de adrenalina (23, 31). Sin embargo, aún no está claro como están mediadas metabolicamente las diferencias en base al género en el patrón de oxidación de combustibles durante el ejercicio.
Un problema con los estudios previos de las diferencias en base al género en el metabolismo durante el ejercicio, es la falta de mediciones de control adecuadas. Para establecer un efecto independiente del ejercicio sobre la oxidación de combustibles, es importante establecer que, bajo condiciones de control, i.e., en reposo y ayuno durante un período similar al del ejercicio, las mujeres no tengan una oxidación de lípidos significativamente mayor. Además, la medición continua de la oxidación de combustibles dentro del período posterior al ejercicio podría establecer si las mujeres compensan la mayor oxidación de grasas durante el ejercicio, por medio de una menor utilización de lípidos que los hombres durante el período de recuperación. Dichas mediciones de la oxidación de combustibles son necesarias para la apropiada interpretación y valoración de las implicaciones de los datos experimentales.
Los estudios sobre la diferencia del metabolismo durante el ejercicio en base al género, requieren también una atención cuidadosa de las cuestiones relacionadas al la selección de los sujetos y al diseño del estudio. Esta puede ser la razón de la falta de diferencias en base al género en el metabolismo durante el ejercicio, observada en ciertas investigaciones (5, 26). Por ejemplo, es importante comprobar el estado menstrual de las mujeres, ya que se sabe que las gonadotrofinas femeninas influencian el metabolismo de los combustibles (12, 16, 21, 27). Además, cuando se realiza un intento por igualar ambos grupos, podría no ser práctico o lógico hacerlo en base a la composición corporal o el consumo máximo de O2 (VO2 máx.), ya que estos dos parámetros naturalmente difieren entre hombres y mujeres. Por ello, puede ser mas apropiado seleccionar e igualar los sujetos en base al estatus de entrenamiento y a la cantidad del mismo, que como se sabe es el principal determinante de la utilización de combustibles durante el ejercicio (3, 8, 13). De este modo, la utilización de esta vía para emparejar los grupos, tiene implicaciones para las condiciones de ejercicio seleccionadas. Elegir la misma intensidad relativa de trabajo para la evaluación del ejercicio podría representar un mismo reto metabólico para hombres y mujeres, más que los mismos individuos ejercitando con la misma carga absoluta. Finalmente, es importante un control del consumo dietario previo al estudio y del rendimiento en el ejercicio para asegurar que los sujetos están en un estado de balance energético y de repleción de combustibles similares. Solo mediante la consideración de estos factores en el diseño de los estudios, puede ser posible establecer claramente alguna diferencia en base al género en el metabolismo durante el ejercicio.
De este modo, la finalidad de este estudio, fue comparar la oxidación de combustibles en hombres y mujeres durante y después del ejercicio y en un día de control. Esto se realizo considerando los factores anteriormente mencionados para el diseño del estudio. Simultáneamente, se caracterizó el ambiente hormonal y de los sustratos circulantes para elucidar posibles mecanismos involucrados en cualquier diferencia observada, en base al género, en la oxidación de combustibles durante el ejercicio. Finalmente, estudiamos sujetos entrenados y desentrenados para determinar si hubo alguna interacción entre el género y el estado de entrenamiento con respecto al combustible para el ejercicio.
METODOS
Sujetos
Para el estudio se reclutaron hombres (n=14) y mujeres (n=14) entrenados y desentrenados (Tabla 1). Los sujetos entrenados eran principalmente ciclistas competitivos, con un triatleta tanto en el grupo de hombres como en el grupo de mujeres. Los sujetos fueron emparejados en base al estado de entrenamiento (volumen e intensidad del entrenamiento, determinada por los registros de actividad reportados por los sujetos). Los sujetos desentrenados participaron en <90 min de ejercicio aeróbico por semana. Todos los sujetos tenían peso normal, no eran diabéticos ni fumadores, y no sufrían de ninguna condición médica o tomaban cualquier medicación crónica conocida que interfiriera con el metabolismo intermedio. Todas las mujeres eran eumenorreicas y no utilizaban anticonceptivos esteroides. El protocolo del estudio fue aprobado por el Comité de Revisión Institucional para la Protección de Sujetos Humanos de la University of Colorado. Todos los sujetos leyeron y firmaron un formulario de consentimiento antes de ser admitidos en el estudio.
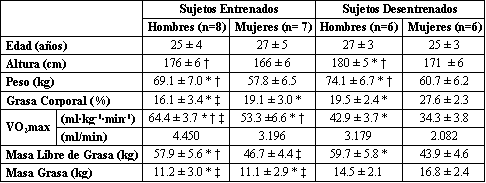
Tabla 1. Características de los sujetos. Los valores son presentados como
medias±DS. Mujeres entrenadas (n=7); se excluyo una mujer entrenada que estaba
en la fase lútea de su ciclo menstrual. VO2 máx., consumo máximo de O2.
* p<0.05 con respecto a mujeres desentrenadas. † p<0.05 con respecto a mujeres
entrenadas. ‡ p<0.05 con respecto a hombres desentrenados.
Evaluaciones Preliminares
VO2 máx.
VO2 máx. se determinó utilizando un test progresivo en ciclo ergómetro (Monark Varberg, Suecia). Los sujetos entrenados pedalearon a una tasa de 90 rpm y los sujetos desentrenados a una tasa de 70 rpm mientras que la resistencia se incrementó gradualmente (comenzando con 1 kp e incrementándose en 0.5 kp cada 1-2 min) hasta el agotamiento. La frecuencia cardiaca se monitoreo continuamente mediante un electrocardiógrafo de 12 derivaciones, a la vez que se midieron la presión sanguínea y el esfuerzo percibido en el minuto final de cada nivel de trabajo. Para asegurar que se alcanzara un esfuerzo máximo, se estableció como requisito que se cumplieran dos de los siguientes criterios: índice de intercambio respiratorio corporal total ≥1.1, una frecuencia cardiaca máxima dentro del 5% de el máximo estimado, y un incremento <2.0 ml•kg-1•min-1 en respuesta a la carga final.
Composición Corporal
La composición corporal fue determinada por medio de la medición de la densidad corporal utilizando el pesaje subacuático, previamente descrito (14). El porcentaje de grasa se estimó a partir de la densidad corporal utilizando la ecuación revisada de Brozek y cols. (4).
Tasa Metabólica de Reposo
La tasa metabólica de reposo (RMR) se midió utilizando calorimetría indirecta mediante un sistema recolección metabólica (modelo 2900, Sensormedics, Yorba Linda, CA). Los sujetos fueron evaluados a la mañana después de 12 h de ayuno. La tasa metabólica se midió durante 15–20 min, después de 30 min de reposo, usando un toldo ventilado. Las concentraciones de gases fueron medidas en el aire existente en el toldo ventilado. El consumo de O2 y la producción de CO2 se utilizaron para calcular la tasa metabólica (35). La RMR fue utilizada para determinar el consumo energético de los sujetos durante el período previo al estudio para el control de la dieta.
Examen Médico y Físico
Los sujetos realizaron un examen médico y físico para confirmar que no había razones médicas para su exclusión del estudio.
Control de la Dieta y el Ejercicio antes del Estudio
Los sujetos fueron alimentados con una dieta controlada durante los 3 días previos a cada día de estudio. Todos los alimentos fueron preparados en la cocina del Centro General de Investigación Clínica (GCRC) en la Universidad de Colorado, y se les pidió a los sujetos que desayunaran en el GCRC, a la vez que se les preparaba otra comida para cuando dejaran el sitio. También se les pidió a los sujetos que consumieran toda la comida que se les suministro y no se permitió ninguna otra comida. La única parte opcional de la dieta fueron dos porciones de comida (de 840 kJ cada una), de las cuales se podían consumir una o ambas si los sujetos sentían hambre. La dieta estaba compuesta de 30% de grasas, 15% de proteínas y 55% de carbohidratos, y la ingesta energética inicial se calculó como 1.6 x RMR y 1.9 x RMR para los sujetos desentrenados y entrenados, respectivamente. Uno de los hombres entrenados y una de las mujeres entrenadas pidieron dos porciones adicionales de comida cada uno. El peso corporal fue medido diariamente antes de que los sujetos consumieran el desayuno para comprobar la estabilidad del mismo. Se les permitió a los sujetos continuar con su rutina de actividades usuales en los dos primeros días de la dieta, mientras que en el último día se abstuvieron de realizar ejercicio.
Días del Estudio
Los sujetos pasaron la noche anterior a cada estudio en el GCRC y fueron evaluados al día siguiente después de 10 h de ayuno. Todos los sujetos completaron 2 días de evaluaciones: un día de ejercicio y un día de control. El orden de las evaluaciones en cada grupo fue aleatorio, y el primer y segundo día del estudio estuvieron separados por un período de 3 a 6 semanas. Las mujeres fueron estudiadas en la fase folicular de su ciclo menstrual (Estudio de Ejercicio: días 7.3±2.8 y 6.8±3.5 en mujeres entrenadas y desentrenadas, respectivamente, donde el día 1 es el primer día de la menstruación; Estudio de Control: días 7.5±3.9 y 6.5±3.3 en mujeres entrenadas y desentrenadas, respectivamente). Para estimar el día deseado para la realización del estudio se utilizó la historia menstrual previa, y tanto la fase como el día del ciclo menstrual fueron confirmados mediante el reporte de la menstruación más reciente y la determinación del estradiol y progesterona séricas en el día del estudio. Se hallo que una de las mujeres entrenadas estuvo en su fase lutea en ambos días del estudio, pero no fue capaz de repetir cualquiera de los dos días. Los análisis de los datos con y sin sus resultados no afectaron los resultados sobre la oxidación de sustratos, pero si tuvieron impacto sobre algunos de los datos sobre las hormonas. Por ello sus resultados fueron excluidos del análisis. En la mañana del estudio, los sujetos fueron despertados a ~05:45 h. Se colocó un catéter flexible (de medida 18–20) en una vena cercana al espacio antecubital de uno de los brazos. Fue colocada una llave de cierre de tres vías para el muestreo y para la infusión de una solución salina normal. Los sujetos fueron entonces transportados en auto (3 minutos) al laboratorio para la evaluación.
Día de Ejercicio
Los sujetos vaciaron completamente sus vejigas y completaron la recolección de orina en las 24 h subsiguientes. En ese momento se recostaron y descansaron por 30 min antes de los 15–20 min de la medición de la RMR. Inmediatamente después, se tomaron las muestras sanguíneas basales. Luego de esto los sujetos pedalearon por 2 h al 40% del VO2 máx. en un ciclo ergómetro (Monark), después de haber realizado 5 min de entrada en calor. Luego de la finalización del ejercicio, los sujetos estuvieron 2 h en reposo en posición semireclinada.
Día de Control
El día de control siguió exactamente el mismo formato que el día de ejercicio, con la excepción de que los sujetos no realizaron ejercicio, pero reposaron en posición semireclinada el mismo tiempo total que durante el día de ejercicio (4 h en total).
Determinaciones Metabólicas
Mediciones del Intercambio de Gases Respiratorios
El intercambio de gases respiratorios se midió durante el período de 4 h de cada evaluación utilizando, el sistema de transferencia metabólico y una mascara Hans Rudolph. Las mediciones se realizaron continuamente por períodos de ~30 min, recalibrando el sistema de transferencia metabólico entre las mediciones. La oxidación de grasas y carbohidratos se calculó a partir del volumen de O2 consumido y el volumen de CO2 espirado, luego de la corrección de la oxidación de proteínas (17). La oxidación de proteínas se estimó dividiendo la excreción de nitrógeno en 24 h en porciones de 2 h. La utilización de este marco de tiempo más que el tiempo del estudio en si mismo, para el cálculo de la oxidación de proteínas, se debió a que: 1) algunos sujetos no pudieron orinar luego del ejercicio y 2) observamos un tiempo de retraso en la excreción de nitrógeno luego del ejercicio como resultado de la oxidación de proteínas. Estimamos una diferencia de menos del ±0.5% en la estimación de gramos de carbohidratos y grasa oxidados cuando utilizamos la recolección de orina en el período de estudio vs. los valores de excreción de nitrógeno estimados a partir de la recolección de orina en 24 h.
Determinación de los Niveles Circulantes de Sustratos y Hormonas
Las muestras de sangre se extrajeron de una vena del antebrazo, por ello, las mediciones de sustratos y hormonas circulantes representan concentraciones arteriales, así como también el drenaje venoso del lecho muscular local. Aproximadamente un ml de sangre total se adiciono a un tubo pre pesado que contenía 3 ml ácido perclórico congelado (8%). Luego el tubo fue pesado y centrifugado para separar el sobrenadando. Para la determinación de las concentraciones de catecolaminas plasmáticas toda la sangre (2.5 ml) fue adicionada a 40 μg de solución preservativa (3.6 mg de EGTA + 2.4 mg de glutatión en agua destilada). Las muestras fueron inmediatamente puestas en hielo y centrifugada. Se permitió que se coagularan aproximadamente 7 ml de sangre total, y después de la centrifugación se le separo el suero. Todas las muestras de plasma, suero y sobrenadando fueron guardadas a –70 ºC hasta los análisis. Las catecolaminas fueron determinadas en duplicado mediante análisis radioenzimatico [coeficientes de variación intra-análisis (CV) = 16 y 8% para la noradrenalina y la adrenalina, respectivamente] (24). El método de radioinmunoensayo fue utilizado para determinar la insulina sérica (Kabi Pharmacia, Piscataway, NJ); cortisol, progesterona, estradiol y testosterona (Diagnostic products, Los Angeles, CA) y globulina ligadora de las hormonas sexuales (SHBG, Wien Laboratories, Saccasunna, NJ). Las muestras fueron analizadas en duplicado, con un coeficiente de variación intra-análisis de 10, 6.7, 8.8, 7.5, 9.1, y 7.0%, respectivamente. La glucosa sérica se midió enzimáticamente, solamente una vez, utilizando el método de la hexoquinasa en un analizador de glucosa automático (Rohe COBAS Mira Plus, coeficiente de variación intra-análisis = 0.7%). Los análisis enzimáticos también fueron utilizados para la determinación de glicerol sérico (Boehringer Mannheim Diagnostics, Indianapolis, IN) y los ácidos grasos libres (AGL, Wako Chemical, Richmond, VA), solamente una vez (coeficiente de variación intra-análisis= 7.8 y 1.2%, respectivamente). El ácido β-hidroxibutírico (β-HBA) y el lactato sanguíneo (Sigma Diagnostics, St. Louis, MO) fueron analizados en duplicado con un coeficiente de variación intra-análisis de 10.8 y 4.2%, respectivamente. Todos los análisis de las muestras de ambos días del estudio de cada sujeto fueron realizados simultáneamente.
Análisis de los Datos
Los datos fueron comparados por sexo y estado de entrenamiento, en cada día de estudio, determinando también cualquier interacción entre los factores de agrupamiento. Ya que solo se observaron efectos significativos del ejercicio sobre la oxidación de combustibles entre géneros, todos los datos fueron presentados utilizando una separación en base al género. Para la oxidación de combustibles y el gasto energético (GE), cada día del estudio fue dividido en dos partes que comprendían las primeras dos horas de mediciones (período de ciclismo, en el día de ejercicio) y las segundas dos horas de mediciones (período de recuperación en el día de ejercicio). La oxidación de combustibles se expresó como la oxidación absoluta en el período de 2 h y como la oxidación relativa (en porcentaje): (kJ de oxidación de combustible/total de kJ oxidados) x 100. El último modo de expresión tiene en cuenta los diferentes GE entre individuos y grupos, lo cual podría automáticamente resultar en diferentes tasas de oxidación absoluta de combustibles.
Inicialmente se utilizo el análisis de varianza ANOVA univariado para determinar diferencias entre los grupos en las variables para el GE y la oxidación de combustibles en períodos de tiempo comparables, en cada uno de los días del estudio. Donde se observaron diferencias significativas, estas fueron confirmadas por medio del análisis multivariado que incluyo al porcentaje de grasa y al VO2 máx., como covarianzas. Estas dos variables fueron incluidas en el modelo estadístico, ya que son consideradas factores de confusión en los resultados del análisis univariado. Sin embargo todos los datos son presentados como la media sin ajustar con las estadísticas del análisis univariado. Para los sustratos y las hormonas, se utilizo el ANOVA para medidas repetidas, usando el género como factor de agrupamiento y cada punto en el tiempo en que se tomaron muestras como medidas repetidas. Cualquier interacción entre el género y el tiempo fue determinada para establecer si el patrón de cambio en cada parámetro difería entre los géneros. Cuando se hallaron interacciones significativas entre el tiempo y el género, se utilizo el test t para datos no apareados, para determinar si también hubo puntos del tiempo específicos donde ocurrieran diferencias en base al género. Las características de los grupos se compararon utilizando ANOVA y donde fuera apropiado, se realizaron comparaciones post hoc utilizando el test de Fisher. Las diferencias en los parámetros dentro del grupo (entre períodos de mediciones y entre días) fue comparada utilizando el test t para datos apareados. Se establecieron diferencias significativas con una p<0.05.
RESULTADOS
Características Físicas
No hubo diferencias entre los géneros en la edad y la masa grasa. Sin embargo, los hombres fueron significativamente mas pesados, mas altos y tenían mayor masa libre de grasa que las mujeres (p<0.05). El VO máx. fue mayor en los hombres que en mujeres en el margen de la significancia estadística (p<0.06), mientras que el porcentaje de grasa fue significativamente mayor en mujeres (p<0.05). Los niveles de actividad fueron similares en mujeres y hombres entrenados (ciclismo: 135±41 y 132±54 millas/semana, respectivamente, otras actividades aeróbicas: 3 y 2.5 h/semana respectivamente; entrenamiento de la fuerza: 2 y 2.5 h/semana, respectivamente). La ingesta dietaria y el cambio en el peso a lo largo de los 3 días en cada uno de los estudios son mostrados en la Tabla 2. La ingesta energética y de nutrientes fue similar en los 3 días previos a los días de control y de ejercicio, y el peso se mantuvo estable tanto en hombres como en mujeres antes de cada uno de los días del estudio.

Tabla 2. Ingesta dietaria y cambios en el peso corporal antes de cada uno de
los días del estudio. Los valores son presentados como medias±DS. CHO,
carbohidratos. El cambio en el peso corporal se estimo a partir del peso
corporal medido en ayunas en la mañana del primer día de control de la dieta -
el peso corporal medido en ayunas en la mañana del día del estudio.
Gasto Energético
La RMR relativa a la masa libre de grasa o masa magra fue similar en hombres y en mujeres (Figura 1). No hubo diferencias entre la RMR medida en el día de ciclismo y la medida en el día de control (Tabla 3: 5.38±0.21 y 5.33±0.13 kJ/min respectivamente para los hombres; y 4.37±0.08 y 4.41±0.12 kJ/min, respectivamente para las mujeres). El gasto energético de los 120 min de ejercicio en bicicleta fue significativamente mayor en los hombres que en las mujeres (Tabla 4). Esto estuvo relacionado a la mayor tasa de trabajo absoluta realizada por los hombres (338±16 y 219±14 kpm/min para los hombres y las mujeres, respectivamente, p<0.05). Sin embargo, la intensidad relativa de ejercicio fue similar entre ambos grupos (frecuencia cardiaca promedio: 121±4 y 125±4 latidos/min para los hombres y las mujeres, respectivamente, intensidad= 42.1±0.9 y 42.3±0.8% del VO2 máx., para los hombres y las mujeres respectivamente). Durante la recuperación, el gasto energético se elevó significativamente por encima del valor del día de control en los hombres (p<0.001) pero no en las mujeres.
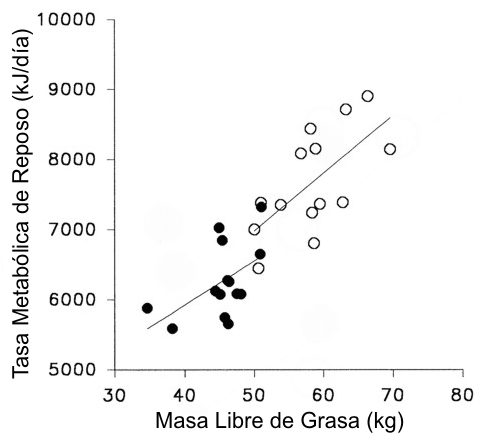
Figura 1. Relación
entre la masa libre de grasa y la tasa metabólica de reposo en hombres (○) y
mujeres (●). La tasa metabólica de reposo medida en los días de ciclismo y de
control fue promediada. r=0.67 para hombres y 0.52 para mujeres.
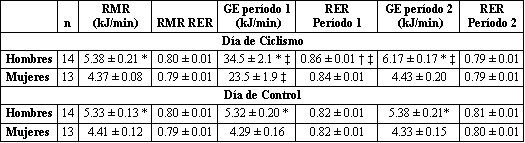
Tabla 3. RMR, gasto energético del ejercicio, y RMR no proteico. Los valores
son presentados como medias±DS. RMR, tasa metabólica de reposo; RER, índice de
intercambio respiratorio; GE, gasto energético. Período 1: tasa de gasto
energético durante las primeras 2 h de medición (comprende el período entero de
ejercicio en ciclo ergómetro en el día de ciclismo); Período 2: tasa de gasto
energético durante las segundas 2 h de mediciones (comprende el período de
recuperación post-ejercicio en el día de ciclismo). Significativamente diferente
con respecto a las mujeres, * p<0.001; † p<0.05. ‡ Significativamente diferente
con respecto al día control, p<0.001.
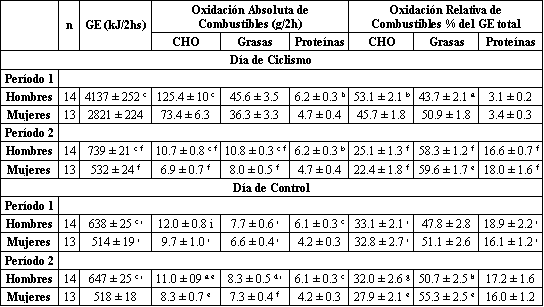
Tabla 4. Gasto energético y oxidación de combustibles. Los valores son
presentados como medias±DS. Ver la tabla 3 para la explicación de los períodos 1
y 2. Significativamente diferente con respecto a las mujeres: a p<0.02, b
p<0.01; c p<0.001. Significativamente diferente con respecto al mismo día del
período 1: d p<0.05; e p<0.01; f p<0.001. Significativamente diferente con
respecto al día de ciclismo del mismo período: g p<0.05, h p<0.01; i p<0.001.
Oxidación de Combustibles
El índice de intercambio respiratorio de reposo no fue diferente entre los dos grupos entre los días de estudio (día de ciclismo: 0.80±0.01 y 0.79±0.01 para hombres y mujeres, respectivamente; día de control: 0.80±0.01 y 0.79±0.01 para hombres y mujeres, respectivamente). Durante el ejercicio, la oxidación absoluta de carbohidratos y proteínas fue significativamente mayor en los hombres que en las mujeres (Tabla 4; p<0.001 y p<0.01, respectivamente), pero no hubo diferencias en base al género en la oxidación absoluta de las grasas. Cuando los datos de la oxidación fueron expresados como un porcentaje del gasto energético, la contribución de la oxidación de grasas al gasto energético del ejercicio fue significativamente mayor en mujeres que en los hombres (p<0.02), mientras que la contribución de la oxidación de carbohidratos al gasto energético total en el ejercicio fue significativamente mayor en los hombres (p<0.01; Tabla 4). No se observaron diferencias en base al género en la contribución relativa de la oxidación de grasas y carbohidratos al gasto energético total durante el período de recuperación o en el día de control.
Como se esperaba, la oxidación absoluta de combustibles fue mayor durante el período 1 del día de ciclismo (ejercicio) que durante el período 2 (recuperación) y que durante el período 1 del día de control (Tabla 4). Durante el período posterior al ejercicio la contribución de las grasas al gasto energético total se incremento significativamente en relación al período de ejercicio en hombres (p<0.001) y en mujeres (p<0.01), mientras que la contribución de carbohidratos disminuyó significativamente en ambos grupos (p<0.001). En el día de control, hubo también un incremento en la contribución de las grasas al gasto energético total desde el período 1 al período 2, pero este incremento solo fue significativo para las mujeres (p<0.01). Consecuentemente, el incremento en la oxidación relativa de grasas en hombres durante la recuperación en el día de ejercicio fue mayor que la oxidación relativa de grasas durante el mismo período de tiempo en el día de control (p<0.01).
Niveles de Sustratos Circulantes
Los niveles de substrato circulante en los días de ejercicio y de control son presentados en la Figura 2. Para ambos grupos, en el día de ciclismo, la glucosa, AGL, glicerol, y lactato se incrementaron con el ejercicio y disminuyeron después del ejercicio (p<0.0001), mientras que el β-HBA se incremento principalmente en el período post-ejercicio (P<0.0001). Aunque los niveles de lactato se incrementaron con el ejercicio, la elevación fue relativamente pequeña (cambio máximo de ~0.2 mmol/l para ambos grupos), y fue improbable que tuviera algún impacto sobre la producción no metabólica de CO2. Durante el ejercicio el incremento en los AGL fue significativamente mayor en mujeres que en hombres (p<0.004). Inversamente, la glucosa y el β-HBA fueron mayores en los hombres durante el período post-ejercicio (p<0.005 y p<0.003, respectivamente). En el día de control el pequeño incremento en los AGL, glicerol, lactato y β-HBA fue también significativo con el tiempo (p<0.001). En este día, los sustratos se incrementaron de forma similar en ambos grupos. Los niveles de lactato fueron significativamente más altos en hombres que en mujeres en el día de control (p<0.02) aunque las diferencias fueron pequeñas en magnitud (~0.2 mmol/l).
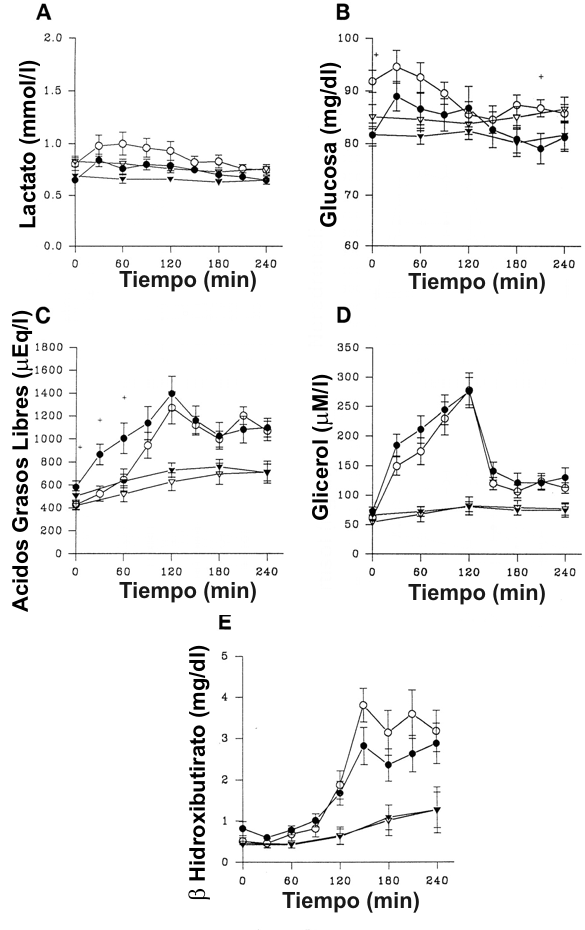
Figura 2. Niveles de sustratos circulantes en hombres (símbolos abiertos) y
mujeres (símbolos rellenos) en los días de ejercicio (círculos) y de control
(triángulos). A: Para el lactato, hubo un efecto significativo del tiempo
(p<0.0001) sobre los días de ciclismo y de control; y un efecto significativo
sobre el género (p<0.02) en el día de control. B: Para la glucosa hubo un efecto
significativo del tiempo (p<0.0001) y un efecto de interacción significativa del
género por el tiempo (p<0.005); solamente para el día de ciclismo, +p<0.05 (t-test
para datos no apareados). C: Para los ácidos grasos libres hubo un efecto
significativo del tiempo (p<0.0001) sobre los días de ciclismo y de control y un
efecto de interacción significativa del género por el tiempo (p<0.0004) en el
día de ciclismo; +P<0.05 (t-test para datos no apareados) D: Para el glicerol
hubo un efecto significativo del tiempo sobre los días de ciclismo y de control
(p<0.0001 y p<0.001, respectivamente). E: Para el β-hidroxibiutirato hubo un
efecto significativo del tiempo (p<0.0001) sobre los días de ciclismo y de
control, y un efecto de interacción significativa (p<0.003) en el día de
ciclismo).
Hormonas Circulantes
Las figuras 3 y 4 muestran los cambios hormonales durante ambos días del estudio. En el día de ciclismo, todos los niveles de hormonas circulantes cambiaron significativamente con el tiempo (p<0.0001), generalmente incrementándose con el ejercicio y disminuyendo después del ejercicio. Lo mas notable fue un incremento significativamente más grande en la adrenalina (p<0.0009) y la noradrenalina (p<0.007) durante el ejercicio en los hombres que en las mujeres. Los niveles de insulina post-ejercicio fueron significativamente mayores en los hombres que en las mujeres (p<0.0009), pero la magnitud absoluta de esta diferencia fue pequeña (1 μU/ml). Aunque los niveles de cortisol fueron mayores en los hombres que en las mujeres en el día de ciclismo, esta diferencia no alcanzo significancia estadística (p=0.08)
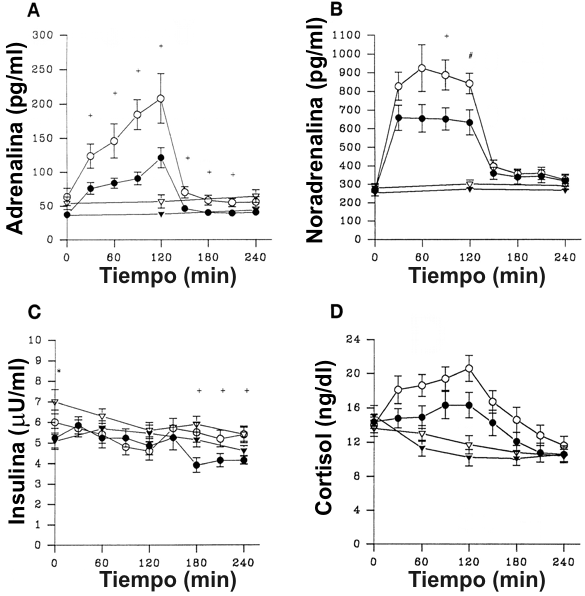
Figura 3. Niveles de
hormonas circulantes en hombres (símbolos abiertos) y mujeres (símbolos
rellenos) en los días de ejercicio (círculos) y de control (triángulos). A: Para
la adrenalina hubo un efecto significativo del tiempo (p<0.0001) y del género
(p<0.003), y una interacción significativa del género por el tiempo (p<0.007)
solamente en el día de ciclismo; +P<0.05 (t-test para datos no apareados). B:
Para la noradrenalina hubo un efecto significativo del tiempo (p<0.0001) y una
interacción significativa del género por el tiempo (p<0.0009) solamente en el
día de ciclismo, +P<0.05, #P = 0.06 (t-test para datos no apareados) C: Para la
insulina hubo un efecto significativo del tiempo (p<0.0001 y p<0.004,
respectivamente) y una interacción significativa del género por el tiempo
(p<0.0009 y p<0.01, respectivamente) en los días de ciclismo y de control; +
p<0.05 en el día de ciclismo, * p<0.05 en el día de control (t-test para datos
no apareados) D: Para el cortisol hubo un efecto significativo del tiempo
(p<0.0001) en los días de ciclismo y de control.
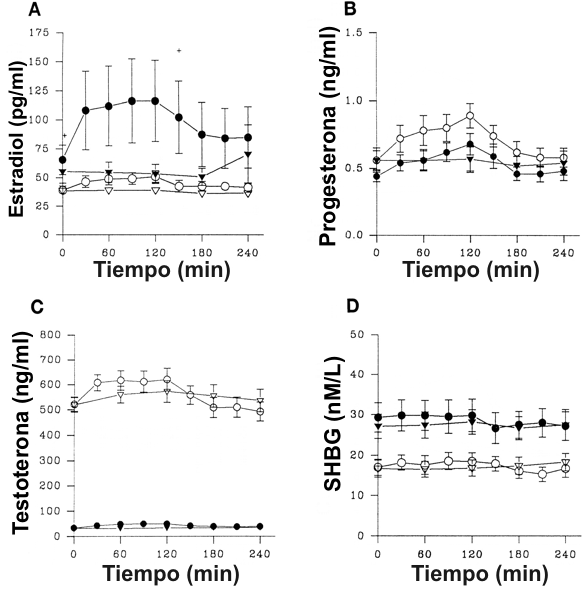
Figura 4. Niveles de
hormonas gonadotróficas en hombres (símbolos abiertos) y mujeres (símbolos
rellenos) en los días de ejercicio (círculos) y de control (triángulos). A: Para
el estradiol hubo un efecto significativo del tiempo (p<0.0001) y un efecto de
interacción significativo del género por el tiempo (p<0.04) solamente en el día
de ciclismo; +p<0.05 (t-test para datos no apareados). B: Para la progesterona
hubo un efecto significativo del tiempo (p<0.0001 y p<0.007 respectivamente) en
los días de ciclismo y de control. C: Para la testosterona hubo un efecto
significativo del tiempo (p<0.0001) y un efecto de interacción significativo del
género por el tiempo (p<0.0001) solamente en el día de ciclismo y un efecto
significativo del género en los días de ciclismo y de control (p<0.0001). D:
Para la globulina ligadora de las hormonas sexuales (SGBH) hubo un efecto
significativo del tiempo (p<0.0001) solamente en el día de ciclismo y un efecto
significativo del género (p<0.009 y p<0.01 respectivamente) en los días de
ciclismo y de control.
Con respecto a las hormonas gonadotróficas, en los días de control y de ejercicio, los niveles de SHBG fueron significativamente mayores en mujeres que en hombres (p<0.01 y p<0.009, respectivamente), mientras que los niveles de testosterona fueron significativamente mayores en los hombres (p<0.0001). El incremento en el estradiol durante el ejercicio fue mayor en las mujeres que en los hombres (p<0.04). La testosterona se incrementó con el ejercicio, más en los hombres que en las mujeres (p<0.0001), pero los niveles no fueron significativamente diferentes con respecto a los valores en el día de control en cualquiera de los grupos.
DISCUSION
Este estudio demostró diferencias significativas en base al género en la oxidación de combustibles y la respuesta hormonal al ejercicio. Con la misma intensidad relativa de ejercicio (de suave a moderada), la oxidación de grasas contribuyó más al gasto energético total en las mujeres que en los hombres. Consecuentemente, la contribución de la oxidación de carbohidratos al gasto energético del ejercicio fue menor en las mujeres que en los hombres. La observación mas notable, sin embargo, fue el hallazgo de niveles significativamente mayores de adrenalina y noradrenalina en los hombres que en las mujeres durante el ejercicio. No puede ser directamente establecido si estas hormonas desempeñan un rol en las diferencias observadas en base al género, en la oxidación de los combustibles. Los presentes datos confirman observaciones anteriores de una mayor oxidación relativa de grasas y una menor oxidación relativa y/o absoluta de carbohidratos en mujeres en comparación con hombres (1, 25, 31, 32). Esto también extiende observaciones anteriores estableciendo que estas diferencias en base al género, están limitadas a la condición del ejercicio en si misma. Nosotros no hemos observado diferencias entre hombres y mujeres en la contribución de lípidos y carbohidratos al gasto energético previo al ejercicio, luego del ejercicio o en el día de control. Los datos fueron analizados teniendo en cuenta las diferencias en el VO2 máx. y el porcentaje de grasa entre hombres y mujeres. De este modo, parece que hay diferencias sexuales específicas en el establecimiento de una prioridad de las grasas y los carbohidratos, como combustibles, para la oxidación durante el ejercicio. Además, esto fue independiente del estado de entrenamiento, tanto en hombres como en mujeres.
Aunque la magnitud de las diferencias observadas en el presente estudio fue pequeña, otros investigadores han observado mayores diferencias en base al género entre la contribución de la oxidación de grasas y la contribución de la oxidación de carbohidratos al gasto energético durante el ejercicio (25, 31, 32). Estos estudios utilizaron una intensidad de ejercicio algo mayor (65 – 85% del VO2 máx.) que la utilizada en el presente estudio (40% VO2 máx.). De este modo, la magnitud de las diferencias sexuales entre la oxidación de grasas y carbohidratos puede estar relacionada a la intensidad del ejercicio realizado. Es posible que a intensidades suaves de ejercicio, las diferencias en base al género en la oxidación de grasas puedan ser menores, ya que ambos grupos pueden fácilmente utilizar lípidos como fuentes de energía. Inversamente, parece probable que a intensidades de ejercicio muy altas, donde la utilización de carbohidratos es predominante y se incrementa el metabolismo anaeróbico, las diferencias sexuales en la utilización de combustibles podrían no ocurrir.
En el presente estudio transversal, los sujetos entrenados no tuvieron una oxidación de lípidos significativamente mayor que los sujetos desentrenados. Esto contrasta con los estudios de entrenamiento longitudinales, en los cuales se observaron mayores tasas de oxidación de lípidos después del entrenamiento (15). Esta discrepancia puede ser explicada por las diferencias en el diseño del estudio (transversal vs. longitudinal) y en el uso de diferentes regímenes de ejercicio (intensidades de ejercicio relativas vs. absolutas). Además, la intensidad de ejercicio utilizada en el presente estudio fue relativamente suave a moderada para todos los sujetos y pudo favorecer una utilización semejante de lípidos tanto en los sujetos desentrenados como en los entrenados.
La implicación de diferencias sexuales en la oxidación de combustibles durante el ejercicio aeróbico se relaciona al control de las reservas de combustibles, tanto de lípidos como de carbohidratos (glucógeno). Con respecto a la grasa corporal total, se podría intuir que las mujeres oxidan más lípidos que los hombres durante el ejercicio, dado el hecho de que las mujeres normalmente mantienen mayores niveles de grasa corporal que los hombres. Sin embargo, la magnitud de las diferencias sexuales en la oxidación relativa de grasas observada durante el ejercicio fue pequeña y puede fácilmente compensarse por medio de diferencias en el gasto energético post-ejercicio y/o la ingesta de alimentos. Con respecto a las reservas de carbohidratos, la menor oxidación de carbohidratos durante el ejercicio en mujeres ha mostrado estar relacionada a una menor depleción de glucógeno (31, 32). De este modo, el incremento de las reservas de glucógeno para mejorar el rendimiento puede ser menos efectivo en mujeres que en hombres (32).
La respuesta de las catecolaminas, que durante el ejercicio fue significativamente mayor en los hombres que en las mujeres sugiere un rol de estas hormonas en las diferencias sexuales en la oxidación de combustibles. Observaciones previas han sugerido una mayor respuesta al ejercicio de la adrenalina (23, 31), pero no de la noradrenalina (9, 31), en los hombres en comparación con mujeres. Uno de los principales efectos de la adrenalina es la estimulación de la lipólisis (10, 20). Sin embargo, en el presente estudio no observamos diferencias basadas en el género en los niveles circulantes de glicerol, una estimación de la lipólisis corporal total. Esto también se ha observado previamente (9, 31), aunque los reportes son inconsistentes (1, 31). Los autores reconocen que los cambios en los niveles circulantes de glicerol no reflejen solamente la liberación de glicerol desde las reservas de triacilglicéridos; sin embargo, estos dos parámetros parecen estar altamente correlacionados (2, 29). De esta manera, si se asume que un cambio en el nivel de glicerol es una estimación razonable de la lipólisis, los datos del glicerol en este estudio sugieren una mayor sensibilidad a la acción lipolítica de una o ambas catecolaminas en las mujeres. Esto es consistente con datos anteriores sobre la respuesta a la hipoglucemia en hombres y mujeres (6). Sin embargo, bajo condiciones de reposo no se observaron diferencias sexuales en los niveles de glicerol durante la infusión con adrenalina (34). Esto sugiere que puede haber una interacción entre las catecolaminas y el estrés (i.e. ejercicio y/o hipoglucemia) que afecta la sensibilidad lipolítica en la mujeres en relación a los hombres.
La adrenalina puede también estimular la glucogenolisis, particularmente en el músculo (7). Por ello, el mayor nivel de catecolaminas durante el ejercicio en los hombres pudo haber resultado en una mayor movilización y, potencialmente, utilización del glucógeno muscular. En efecto, se ha observado previamente una mayor declinación en las reservas de glucógeno, después del ejercicio en hombres en comparación con mujeres (31). Sin embargo, en el presente estudio, no hemos medido la glucogenólisis. Se podría postular que una mayor sensibilidad en las mujeres en comparación con los hombres, a la acción lipolítica de las catecolaminas, en conjunto con un efecto glucogenolítico similar, podría favorecer la movilización de lípidos sobre la de carbohidratos a un nivel de catecolaminas dado.
La principal diferencia sexual observada en las concentraciones de sustratos circulantes durante el ejercicio fue el nivel de AGL mayor en las mujeres que en los hombres. Esto ha sido reportado por otros autores que utilizaron ejercicio suave (1), mientras que con ejercicios de intensidad moderada no se han observado diferencias en los AGL circulantes (9, 31). Sin embargo, los niveles de sustratos circulantes no reflejan la utilización de combustibles, pero representan un balance entre la producción y la utilización. Esto es relevante, ya que nosotros observamos diferencias sexuales en la concentración de AGL circulantes, pero no en los niveles de glicerol circulante. De este modo, si la lipólisis estuvo estimulada con la misma amplitud en ambos grupos, los menores niveles de AGL en los hombres en comparación con las mujeres pudieron haberse debido a una mayor oxidación de AGL derivados del tejido adiposo que las mujeres. Esto podría ser solamente confirmado por mediciones del producción y remoción de sustrato. Los AGL derivados del tejido adiposo pueden, por ello, no haber contribuido a la oxidación relativa de las grasas en las mujeres. Durante el ejercicio, otras fuentes de lípidos pudieron haber contribuido a la mayor oxidación relativa de grasas en las mujeres durante el ejercicio, incluyendo AGL derivados de triacilglicéridos intramusculares (31) y, en una menor extensión, lipoproteínas de muy baja densidad circulantes (33).
En la presente investigación, los niveles circulantes de glucosa fueron consistentemente menores en mujeres que en hombres, tanto en respuesta al ayuno de la misma duración en el día de control como durante el ejercicio y la recuperación posterior al mismo. Otros investigadores han reportado que, con ejercicio de intensidad moderada (65% del VO2 máx.), la glucosa sanguínea durante el mismo es menor en los hombres que en las mujeres (31). Esto nuevamente implica que la intensidad del ejercicio es un determinante importante de las diferencias sexuales en el metabolismo de los combustibles durante el ejercicio. En el presente estudio es aparente que las mujeres fueron más capaces que los hombres de mantener los niveles de glucosa sanguínea (aunque a un menor nivel total) durante el ejercicio. En el único reporte que midió la cinética de la glucosa en hombres y mujeres (1 h de ciclismo al 50% del VO2 máx.), la utilización de glucosa durante el ejercicio no fue diferente entre los géneros (23). Sin embargo, es difícil realizar inferencias a partir de este estudio realizado con trazadores isotópicos, relacionadas a la presente investigación ya que el anterior estudio no observó diferencias sexuales en la oxidación corporal total de combustibles, probablemente debido al pequeño numero de sujetos estudiados (n=4/género). En el presente estudio, no puede determinarse si los mayores niveles de catecolaminas durante el ejercicio en los hombres resultaron en diferencias en la producción de glucosa. De esta manera, las potenciales diferencias sexuales en la regulación de la glucosa sanguínea y su utilización durante el ejercicio requieren investigaciones adicionales.
Actualmente se desconoce si las diferencias en la utilización de grasas y carbohidratos en hombres y mujeres puede ser explicada por diferencias en el transporte de sustratos, acción hormonal, o la capacidad para la utilización de sustratos. Se puede hipotetizar que la capacidad para oxidar carbohidratos es mayor en los hombres que en las mujeres; y por ello, por defecto, las mujeres deben usar mas lípidos. Esta hipótesis esta respaldada por datos que demuestran una mayor tasa de fosfofructoquinasa/3-hidroxiacilCoA dehidrogenasa en los hombres en comparación con las mujeres, sugiriendo un mayor potencial enzimático para la glucólisis en los hombres (11). Se necesitan futuros trabajos para establecer como los factores circulantes (hormonas y sustratos) y las características musculares dictan las preferencias relacionadas al sexo por los carbohidratos vs. los lípidos.
De manera similar investigaciones anteriores, el β-HBA plasmático se incremento significativamente en hombres y mujeres, después del ejercicio (18). Además los niveles de β-HBA fueron significativamente mayores en los hombres que en las mujeres, después del ejercicio. Los elevados niveles de AGL, después del ejercicio proveen una fuente para la producción de cetónas (22). Sin embargo, el mayor nivel de β-HBA en los hombres no estuvo asociado con una mayor concentración de AGL en el período posterior al ejercicio. La acumulación de cetónas puede derivar en errores en la estimación de la oxidación de combustibles luego del ejercicio (30). Sin embargo, la magnitud de este error es probablemente pequeña (30). La contribución de un mayor nivel de cetónas en los hombres que las en mujeres a la oxidación de combustibles luego del ejercicio no puede ser estimada, ya que no se sabe si la mayor acumulación de β-HBA en los hombres se debió a una mayor producción o una menor remoción. Tampoco medimos el acetoacetato, la otra cetona principal. Sin embargo, es improbable que las diferencias sexuales en los niveles de cetonas pudieran afectar significativamente las estimaciones de la oxidación de combustibles, de modo que la conclusión de ausencia de diferencias sexuales en la oxidación de combustibles post-ejercicio pudiera ser cambiada.
Las otras hormonas medidas en el presente estudio mostraron una diferencia menos dramática entre hombres y mujeres, que la de las catecolaminas. Nosotros observamos un nivel de cortisol mayor no significativo en los hombres en el día de ejercicio durante el ciclismo y la recuperación. Dicha diferencia no fue aparente durante el día de control. Esto sugiere que el cortisol podría tener algún rol, aunque no principal, en la determinación de las diferencias sexuales en la oxidación de combustibles. Se observo un incremento en todas las hormonas gonadotróficas durante el ejercicio. Esto ha sido previamente reportado por otros investigadores (19, 36). Esta elevación en el estradiol, progesterona, y testosterona durante el ejercicio parece deberse a una elevación el las hormonas libres, ya que los niveles de SHGB no cambiaron. Se ha sugerido que una disminución en el clearance o “lavado” hepático de estas hormonas esteroides como resultado de una disminución en el flujo sanguíneo al hígado es responsable del incremento en los niveles circulantes inducido por el ejercicio (28). Los niveles de estradiol fueron muy variables en las mujeres el día de ciclismo. Esto fue debido a que los sujetos fueron estudiados en diferentes puntos de la fase folicular, y se sabe que el estradiol se incrementa a partir de la parte temprana a la parte tardía de esta fase. Sin embargo, es posible que el mayor incremento en el estradiol durante el ejercicio en las mujeres tuviera algún efecto sobre el metabolismo de los combustibles. Por ejemplo, se ha reportado que los estrógenos disminuyen la utilización de glucosa, pero pueden incrementar la disponibilidad de lípidos a través de la estimulación de la lipólisis (12, 27). Se necesitan estudios adicionales para confirmar un rol, directo o indirecto, de las hormonas gonadotróficas y especialmente del estradiol en el metabolismo de los combustibles durante el ejercicio.
En resumen, hemos observado diferencias sexuales significativas en el patrón de oxidación de combustibles tanto en mujeres como en hombres desentrenados y entrenados, agrupados por nivel de actividad física. Las mujeres derivaron mayor energía durante el ejercicio a partir de la oxidación de las grasas y menos a partir de la oxidación de los carbohidratos que los hombres. No hubo diferencias sexuales en la oxidación relativa de combustibles bajo condiciones de reposo antes del ejercicio, después del ejercicio o en un día de control. Las diferencias entre hombres y mujeres en la oxidación de combustibles durante el ejercicio estuvieron asociadas a diferencias sexuales significativas en la respuesta de las catecolaminas al ejercicio. Los hombres tuvieron un incremento mayor tanto en los niveles de adrenalina como de noradrenalina durante el ejercicio. Se espera que futuros estudios puedan proveer una idea acerca de las fuentes de grasas y carbohidratos que contribuyen a las diferencias sexuales observadas en la oxidación de combustibles y también de los mecanismos hormonales y/o metabólicos involucrados en la determinación de un patrón específico, en relación al sexo, en la oxidación de combustibles. Finalmente, podría especularse que, debido a su rol en la reproducción, las mujeres tienen una mayor prioridad por la conservación de los carbohidratos que los hombres bajo condiciones de una demanda de energía incrementada.
Agradecimientos
Los autores quieren agradecer a todos los sujetos que se ofrecieron para el estudio y apreciar su tiempo y cooperación. Los autores también agradecen al Laboratorio Central del GCRC y especialmente a Teddi Wiest-Kent por la asistencia en los aspectos de análisis del estudio.
Notas al Pie
Esta investigación fue respaldada por las Becas DK-42549 y DK-48520 del National Institute of Diabetes and Digestive and Kidney y la Beca 5-01-RR-00051 del National Institutes of Health Division of Research Resources.
Dirección para el pedido de reimpresiones
T. J. Horton, Center for Human Nutrition, Box C225, University of Colorado Health Sciences Center, 4200 East 9th Ave., Denver, CO 80262.
Referencias
1. Blatchford, F. K., R. G. Knowlton, and D. A. Schneider (1985). Plasma FFA responses to prolonged walking in untrained men and women. Eur. J. Appl. Physiol. 53: 343-347
2. Bortz, W. M., P. Paul, A. C. Haff, and W. L. Holmes (1972). Glycerol turnover and oxidation in man. J. Clin. Invest. 51: 1537-1546
3. Brooks, G. A., and J. Mercier (1994). Balance of carbohydrate and lipid utilization during exercise: the "crossover" concept. J. Appl. Physiol. 76: 2253-2261
4. Brozek, J., F. Grande, J. T. Anderson, and A. Keys (1963). Densitometric analysis of body composition: revision of some quantitative assumptions. Ann. NY Acad. Sci. 110: 113-140
5. Costill, D. L., W. J. Fink, L. H. Getchell, J. L. Ivy, and F. A. Witzmann (1979). Lipid metabolism in skeletal muscle of endurance-trained males and females. J. Appl. Physiol. 47: 787-791
6. Davis, S. N., A. D. Cherrington, R. E. Goldstein, J. Jacobs, and L. Price (1993). Effects of insulin on the counterregulatory response to equivalent hypoglycemia in normal females. Am. J. Physiol. 265 (Endocrinol. Metab. 28): E680-E689
7. Febbraio, M. A., D. L. Lambert, R. L. Starkie, J. Prietto, and M. Hargreaves (1998). Effect of epinephrine on muscle glycogenolysis during exercise in trained men. J. Appl. Physiol. 84: 465-470
8. Freidlander, A. L., G. A. Casazza, M. A. Horning, M. A. Huie, and G. A. Brooks (1997). Training-induced alterations in glucose flux in men. J. Appl. Physiol. 82: 1360-1369
9. Freidmann, B., and W. Kindermann (1989). Energy metabolism and regulatory hormones in women and men during endurance exercise. Eur. J. Appl. Physiol. 59: 1-9
10. Galster, A. D., W. E. Clutter, P. E. Cryer, J. A. Collins, and D. M. Bier (1981). Epinephrine plasma thresholds for lipolytic effects in man. J. Clin. Invest. 67: 1729-1738
11. Green, H. J., I. G. Fraser, and D. A. Ranney (1984). Male and female differences in enzyme activities of energy metabolism in vastus lateralis muscle. J. Neurol. Sci. 65: 323-331
12. Hansen, F. M., N. Fahmy, and J. H. Nielson (1980). The influence of sexual hormones on lipogenesis and lipolysis in rat fat cells. Acta Endocrinol. 95: 566-570
13. Holloszy, J. O., and E. F. Coyle (1984). Adaptations of skeletal muscle to endurance exercise and their metabolic consequences. J. Appl. Physiol. 56: 831-838
14. Horton, T. J., H. J. Drougas, T. A. Sharp, L. A. Martinez, G. W. Reed, and J. O. Hill (1994). Energy balance in endurance-trained female cyclists and untrained controls. J. Appl. Physiol. 76: 1937-1945
15. Hurley, B. F., P. M. Nemeth, W. H. Martin, J. M. Hagberg, G. P. Dalsky, and J. O. Holloszy (1986). Muscle triglyceride utilization during exercise: effect of training. J. Appl. Physiol. 60: 562-567
16. Jensen, M. D., M. L. Martin, P. E. Cryer, and L. R. Roust (1994). Effects of estrogen on free fatty acid metabolism in humans. Am. J. Physiol. 266 (Endocrinol. Metab. 29): E914-E920
17. Jequier, E., K. Acheson, and Y. Schutz (1987). Assessment of energy expenditure and fuel utilization in man. Annu. Rev. Nutr. 7: 187-208
18. Johnson, R. H., J. L. Walton, H. A. Krebs, and D. H. Williamson (1969). Metabolic fuels during and after severe exercise in athletes and non-athletes. Lancet 30: 452-455
19. Jurkowski, J. E., N. L. Jones, W. C. Walker, E. V. Younglai, and J. R. Sutton (1978). Ovarian hormonal responses to exercise. J. Appl. Physiol. 44: 109-114
20. Kurpad, A. V., K. Khan, A. G. Calder, and M. Elia (1994). Muscle and whole body metabolism after norepinephrine. Am. J. Physiol. 266 (Endocrinol. Metab. 29): E877-E884
21. Matute, M. L., and R. K. Kalkoff (1973). Sex steroid influence on hepatic gluconeogenesis and glycogen formation. Endocrinology 93: 762-768
22. McGarry, J. D., and D. W. Foster (1980). Regulation of hepatic fatty acid oxidation and ketone body production. Annu. Rev. Biochem. 49: 395-420
23. Mendenhall, L. A., S. Sial, and A. R. Coggan (1996). Gender differences in substrate metabolism during moderate intensity cycling (Abstract). Med. Sci. Sports Exerc. 27: S213
24. Passon, P. G., and J. D. Peuler (1973). A simplified radiometric assay for plasma norepinephrine and epinephrine. Anal. Biochem. 51: 618-631
25. Phillips, S. M., S. A. Atkinson, M. A. Tarnopolsky, and J. D. MacDougall (1993). Gender differences in leucine kinetics and nitrogen balance in endurance athletes. J. Appl. Physiol. 75: 2134-2141
26. Powers, S. K., W. Riley, and E. T. Howley (1980). Comparison of fat metabolism between trained men and women during prolonged aerobic work. Res. Q. Exerc. Sport 51: 427-431
27. Ruby, B. C., R. A. Robergs, D. L. Waters, M. Burge, C. Mermier, and L. Stolarczyk (1997). Effects of estradiol on substrate turnover during exercise in amenorrheic females. Med. Sci. Sports Exerc. 29: 1160-1169
28. Shangold, M. M (1984). Exercise and the adult female: hormonal and endocrine effects. In: Exercise and Sports Science Reviews, edited by R. L. Terjung. New York: Heath, p. 53-79
29. Shaw, W. A. S., T. B. Issekutz, and B. Issekutz, Jr (1975). Interrelationship of FFA and glycerol turnovers in resting and exercising dogs. J. Appl. Physiol. 39: 30-36
30. Simonson, D. C., and R. A. DeFronzo (1990). Indirect calorimetry: methodological and interpretive problems. Am. J. Physiol. 258 (Endocrinol. Metab. 21): E399-E412
31. Tarnopolsky, L. J., J. D. MacDougall, S. A. Atkinson, M. A. Tarnopolsky, and J. R (1990). Sutton. Gender differences in substrate for endurance exercise. J. Appl. Physiol. 68: 302-308
32. Tarnopolsky, M. A., S. A. Atkinson, S. M. Phillips, and J. D. MacDougall (1995). Carbohydrate loading and metabolism during exercise in men and women. J. Appl. Physiol. 78: 1360-1368
33. Terjung, R. L., and H. Kaciuba-Uscilko (1986). Lipid metabolism during exercise: influence of training. Diabetes Metab. Rev. 2: 35-51
34. Webber, J., and I. A. McDonald (1993). A comparison of the cardiovascular and metabolic effects of incremental versus continuous dose adrenaline infusions in men and women. Int. J. Obes. 17: 37-43
35. Weir, D. J. B (1949). New methods for calculating metabolic rate with special reference to protein metabolism. J. Physiol. (Lond.) 109: 1-9
36. Wells, C. L (1991). Women, Sport, and Performance. A Physiological Perspective. Champaign, IL: Human Kinetics