
Metabolismo y Rol de las Grasas durante el Ejercicio (Parte II)
Instituto del Profesorado en Educación Física. Córdoba, Argentina.
Artículo publicado en el journal PubliCE, Volumen 0 del año 2002.
Publicado 20 de octubre de 2002
En el artículo publicado con anterioridad (Desarrollo de la celularidad adiposa), se describió como se producía el desarrollo de las células grasas durante la vida en cuanto a su número y tamaño y como repercutía el balance energético positivo y negativo sobre los cambios en estas células.
En el presente artículo, se describe el camino que siguen las grasas una vez que son absorbidas y su utilización durante el ejercicio; así como las diferentes propiedades que presentan las grasas y carbohidratos para contribuir durante el ejercicio físico satisfaciendo las demandas energéticas de la actividad.
Las grasas conjuntamente con los carbohidratos, constituyen los nutrientes de mayor importancia durante la realización de ejercicios de baja intensidad (se hace referencia a ejercicios cuyas intensidades de trabajo sean menores al 80% del VO2 máx.). En relación a las proteínas, estas solo adquieren relevancia como sustrato energético en situaciones especiales como por ejemplo la inanición, el ayuno prolongado, etc.
En relación a las grasas y carbohidratos, consideremos algunas propiedades físicas y químicas de estos, que los diferencian: (Ver Figura 1)

Fig. 1.
- En primer lugar, las grasas aportan mas
del doble de energía por gramo que los carbohidratos. Mientras las primeras
aportan 9 Kcal , los carbohidratos solo 4 Kcal/gr.
- Ambos nutrientes se almacenan. Los
Hidratos de Carbono lo hacen como glucógeno en hígado y músculo; y por cada
gramo de éste almacenado, se asocian conjuntamente aproximadamente 2.7 gr de
agua. En cambio, las grasas se almacenan en forma de triglicéridos en tejido
adiposo y músculo, pero lo hacen casi en forma anhidra, es decir con escaso
volumen de agua. Esto hace de las grasas un combustible mas eficiente por
unidad de peso. Si todas las grasas de nuestro organismo fueran sustituidas
por una cantidad equienergética de carbohidratos, nuestro peso, sería
entonces aproximadamente el doble, debido a la gran cantidad de agua que se
almacenaría.
- Otra de las propiedades que diferencian a
las grasas de los carbohidratos, es el aporte de ATP. Las grasas son capaces
de aportar más ATP que una molécula de glucosa. Por ejemplo, una molécula de
glucosa aporta aproximadamente 38 ATP cuando es degradada por vía oxidativa,
mientras que una molécula de ácido Esteárico (representando a las grasas) es
capaz de producir 147 ATP.
- Sin embargo, en relación a la cantidad de
oxígeno consumido para producir la ”misma cantidad de ATP”, los ácidos
grasos utilizan más oxígeno que los hidratos de carbono; alrededor de 6 mol
de oxígeno para estos últimos versus 26 mol de oxígeno para el ácido
Esteárico.
- En relación a las reservas energéticas en
Kcal de ambos nutrientes, estas son bastante diferentes. Los carbohidratos
representan en energía acumulada aproximadamente 2000 Kcal para un hombre de
80 Kg, y estas, están distribuidas a nivel plasmático, hepático y muscular
(Ver Figura 2). Las grasas, en cambio, constituyen una reserva energética
acumulada de alrededor de 110.000 a 135.000 Kcal, distribuidas a nivel
plasmático (ácidos grasos libres y triglicéridos), tejido adiposo y
triglicéridos intramusculares (Ver Figura 2) Con estas reservas calóricas,
se podrían satisfacer en ejercicio no mas de 95 minutos de carrera, si el
aporte es realizado por los hidratos de carbono. Sin embargo, el tiempo de
carrera puede extenderse hasta aproximadamente 119 horas, si la energía es
provista por las grasas.
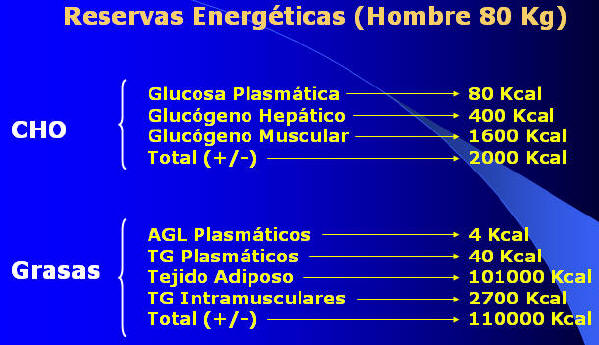
Fig. 2.
Consideremos el recorrido que siguen las grasas una vez que son absorbidas a nivel intestinal y como pasan a formar parte del tejido de reserva o combustibles a nivel muscular para sustentar el ejercicio:
Una vez que los lípidos han sido degradados a través del proceso de la digestión, pasan a los vasos linfáticos en forma de Quilomicrones y luego de allí a la circulación general (Ver Figura 3). El hígado, toma parte de los quilomicrones y junto a los lípidos que en este órgano se sintetizan, forma las Lipoproteínas, que son estructuras mas pequeñas que los quilomicrones pero de composición similar. Las lipoproteínas fabricadas por el hígado pueden ser de diferentes tipos:
- VLDL (very low density lipoprotein o lipoproteínas de muy baja densidad), que transportan gran cantidad de triglicéridos y menor cantidad de fosfolípidos y colesterol.
- IDL (intermediate density lipoprotein o lipoproteínas de densidad intermedia), que son VLDL, pero con un contenido menor de triglicéridos, por lo que aumenta el contenido de fosfolípidos y colesterol.
- LDL (low density lipoprotein o lipoproteína de baja densidad), que son IDL, pero estas solo transportan un gran contenido de colesterol y fosfolípidos, sin presencia de triglicéridos.
- HDL (high density lipoprotein o lipoproteína de alta densidad), también llamado colesterol bueno. Estas transportan aproximadamente el 50% de proteínas y el porcentaje restante de colesterol y fosfolípidos.
La función básica de las lipoproteínas es transportar los componentes lipídicos por la sangre. En general, VLDL transporta los triglicéridos formados en el hígado hacia el tejido adiposo. El resto de las lipoproteínas, transportan fosfolípidos y colesterol del hígado a los tejidos y viceversa.
Una vez que los lípidos circulan por sangre, se estimula una lipoproteína denominada LPLa-m (Lipoproteína lipasa adiposa muscular), presente en la membrana de los vasos sanguíneos, que permite que los ácidos grasos hidrolizados desde los triglicéridos circulantes formen parte del tejido adiposo de reserva o al interior muscular nuevamente como triglicéridos.
Al realizar actividad física, por ejemplo (o en otras situaciones que se detallarán posteriormente), se estimula otra lipoproteína, denominada lipasa hormono-sensible (LPL hs), presente en la membrana de los adipocitos, que facilita el desdoblamiento de los triglicéridos en ácidos grasos y glicerol. Estos ácidos grasos liberados, son transportados por acción de la LPLm al músculo donde serán utilizados como fuente energética preponderante durante la actividad física de baja intensidad. El glicerol restante, se transporta al hígado y se utiliza como precursor neoglucogénico.
En el músculo, los ácidos grasos son activados por una enzima llamada AcilCOa sintetasa, que facilita la unión de ácidos grasos con Coa formando AcilCOa. Esta ultima, atraviesa la membrana de la mitocondria facilitada por la acción de un transportador denominado Carnitina. Posteriormente, en el interior mitocondrial el Acil Coa es degradado por medio de la llamada beta-oxidación, dando origen a AcetiCOa, el cual ingresa al ciclo de Krebs y Cadena respiratoria para culminar su ruptura. (Ver Figura 3)
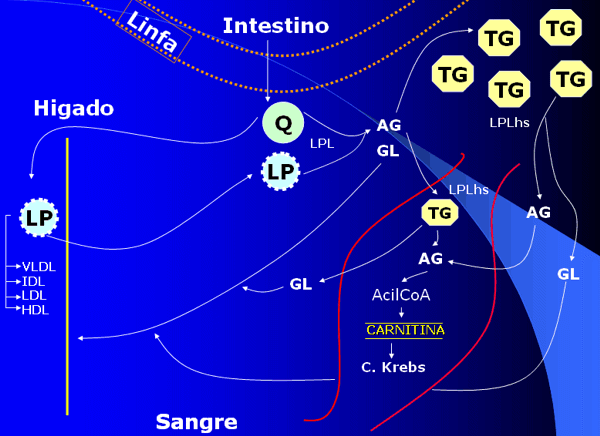
Fig. 3.
En reposo, mas del 50% de los requerimientos metabólicos es derivado de la oxidación de las grasas. Esto podría indicar que en este estado, existe una alta concentración de ácidos grasos plasmáticos. Sin embargo, esto no ocurre, ya que solo el 30% de los ácidos grasos provenientes de la lipólisis aparecen en el plasma. El 70% restante, es reesterificado, es decir es utilizado para la nueva formación de triglicéridos.
En ejercicio, en cambio, si este es de baja intensidad, se produce un aumento en la concentración de ácidos grasos plasmáticos por la lipólisis producida en el tejido adiposo y por el aumento del flujo sanguíneo a dicho tejido. Esto genera una mayor oferta de ácidos grasos disponibles para la oxidación y por tanto el porcentaje de reesterificación disminuye, y hasta incluso puede desaparecer.
La lipólisis originada durante el ejercicio, ya sea en el tejido adiposo o de los triglicéridos intramusculares se encuentra controlada por la acción de la LPLhs, que se activa por la estimulación de receptores beta-adrenérgicos, debido al incremento e los niveles de catecolaminas circulantes.
Los cambios en el uso de nutrientes durante el ejercicio de baja intensidad, pueden ser explicados, al menos en parte, por los cambios manifestados en las concentraciones hormonales. Esta variación en la secreción de hormonas determinará cuan preponderante es el uso de un nutriente sobre otro.
Al comienzo del ejercicio, los hidratos de carbono constituyen el nutriente que cubre las mayores demandas energéticas durante la actividad. Esto ocurre, ya que las primeras hormonas que aumentan en el plasma son las catecolaminas, es decir adrenalina y noradrenalina, posteriormente se produce un incremento en los niveles de glucagón, por lo que se facilita el efecto glucogenolítico (Ver Figura 4). De este modo los carbohidratos aportan la mayor cantidad de energía intraesfuerzo y en menor proporción se realiza un aporte calórico a partir de la oxidación de ácidos grasos.
Luego de 15-20 minutos de actividad, los niveles de hormona del crecimiento (STH) comienzan a elevarse en plasma(Ver Figura 4). Esta hormona promueve una disminución en la utilización de hidratos de carbono y un incremento en la utilización de lípidos. Esto genera alrededor de los 30 minutos el llamado cross-over o cruce de nutrientes (Ver Figura 5). Este punto, implica una disminución en porcentaje en el uso de los carbohidratos como fuente energética y un incremento en la utilización de los lípidos para satisfacer las demandas calóricas de la actividad; por lo que ahora la relación carbohidratos-grasas se invierta en relación a lo que se manifestaba al inicio del ejercicio.
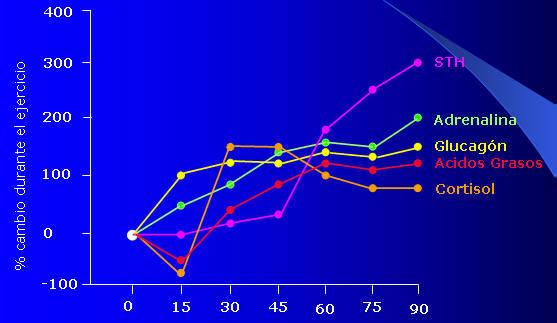
Fig. 4.
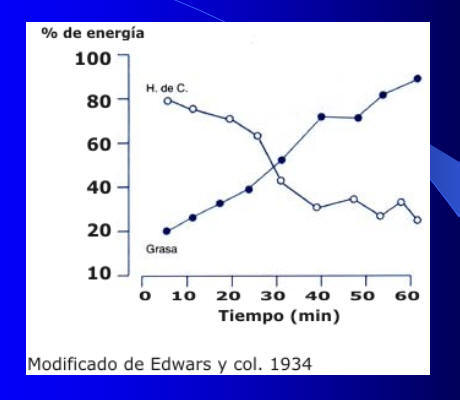
Fig. 5.
El cross-over, no es un punto inflexible, sino que puede modificarse en el tiempo según el nivel de entrenamiento. Es decir que puede generarse, como adaptación una mayor eficiencia en el tiempo y en el tipo de nutriente utilizado durante la actividad. Contrariamente, el sedentarismo, provocará un retardo temporal en el cambio del uso de nutrientes más allá de los 30 minutos; por lo que las demandas energéticas se realizarán en mayor medida movilizando las reservas de carbohidratos y en menor medida las de lípidos.
Finalmente, se incrementan los niveles de Cortisol plasmáticos, por lo que se potencian aún mas los efectos lipolíticos de catecolaminas, glucagón y STH. De este modo, si la actividad física continúa a la misma intensidad y si las reservas de carbohidratos son suficientes, las grasas seguirán siendo el nutriente oxidado de mayor predilección para sustentar la actividad.
Referencias
1. Blanco A (1996). Química Biológica. Ed. El Ateneo
2. Clarkson Priscilla (1990). Pérdida de Peso, Suplementos y Sustancias Químicas: Ganando la Guerra en Contra de la Grasa. Health and Fitness Journal. Vol. 2, numero 4, Pág. 18-26
3. Coyle Edward (1998). Oxidación de las Grasas Durante el Ejercicio: Rol de la Lipólisis, Disponibilidad de Ácidos Grasos Libres, Y Flujo Glucolítico. Proceedings Biosystem
4. Guyton A; Hall J (1997). Tratado de Fisiología Médica. Ed. McGrawn-Hill Interamericana
5. Malina Robert, Bouchard Claude (1991). Growth, Maduration, and Physical Activity. Ed. Human Kinetics
6. McArdle W; Katch M; Katch V (1995). Fisiología del Ejercicio. Ed. Alianza Deporte
7. Menshikov V; Volkov N (1990). Bioquímica. Ed. Vneshtorgizdat
8. Murray Robert; Mayes Peter; Granner Daryl; Rodwell Victor (2001). Bioquímica de Harper. Ed. El Manual Moderno
9. Pujol P (1991). Nutrición, Salud Y Rendimiento Deportivo. Ed. Espaxs (Publicaciones Médicas)
10. Wilmore Jack; Costill David (1998). Fisiología del Esfuerzo y del Deporte. Ed. Paidotribo