
¿Qué es la Potenciación Post Activación?
Universidad Católica de Córdoba.
Artículo publicado en el journal PubliCE, Volumen 0 del año 2013.
Publicado 4 de abril de 2013
Resumen
Palabras clave: potenciación muscular, PAP, fosforilación
INTRODUCCION
El músculo esquelético de mamíferos puede verse alterado en su funcionamiento luego de una actividad muscular previa (Rassier et al., 2000).
Así, de este modo, el efecto más ampliamente estudiado sobre la alteración que un músculo o grupo muscular puede tener luego de una actividad previa es la fatiga, la cual es considerada como una respuesta contráctil menor a la esperada u obtenida previamente a dicha actividad. Por lo tanto podemos definir la fatiga como “una disminución en la capacidad de rendimiento neuromuscular afectada por una actividad previa” (MacIntosch et al., 2002; Asmusen, 1979).
La otra posibilidad es obtener una respuesta contráctil acrecentada como resultado de una actividad contráctil previa, a lo cual llamaremos potenciación. De esta manera podemos definir a la potenciación como “una respuesta muscular contráctil incrementada como resultado de una actividad muscular previa” (Abbate et al., 2000).
Este fenómeno fisiológico de la potenciación se puede desencadenar a través de diferentes modalidades o estrategias de activación, y tendrá una denominación diferente en función del método utilizado para provocarla (Abbate et al., 2000; Sale, 2002).
Una de las estrategias de activación es de forma endógena. Esta modalidad se realiza a través de una contracción voluntaria máxima (CVM) o también llamada contracción voluntaria tetánica (Always et al., 1987; Houston et al., 1987).
La fuerza de una contracción muscular se incrementa luego de una contracción voluntaria máxima (Always et al., 1987; Gossen, 2000; Gossen et al., 2001; Houston et al., 1987; Petrella et al., 1989; Vandervoort et al., 1986; Vandervoort et al., 1983). En este caso a la potenciación obtenida mediante este método la llamaremos potenciación post – activación o PPA (O’leary et al., 1997). En la figura 16 podemos ver la potenciación post – activación.
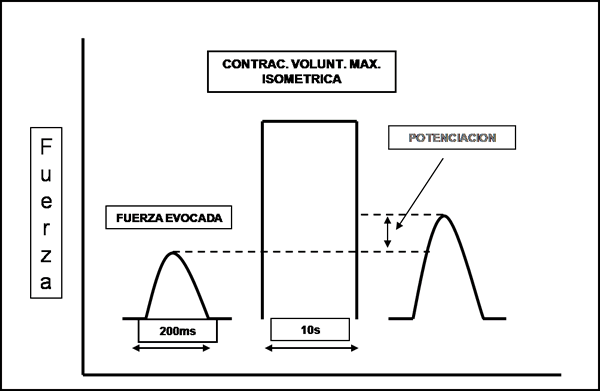
Figura 16. Esquema de una potenciación post-activación luego de una CVMI de 10 s. Modificado de Sale ‘02.
Otra de las formas de activación es mediante una circunstancia exógena o provocada (Hamada et al., 2000; Hamada et al., 2000; MacIntosh et al., 2000). En este caso la estimulación muscular se realiza directamente con un electroestimulador (Boschetti, 2002), en este caso, éste modelo específico de potenciación se denomina potenciación post – tetánica o PPT.
A esta forma exógena de potenciación la podemos llevar a cabo a través de dos formas primarias de estimulación.
Una es la potenciación que se produce luego de una contracción tetánica electroinducida o una estimulación tetánica de alta frecuencia. A este tipo de potenciación también lo llamaremos potenciación post tetánica o PTP. (Por ejemplo: una electroestimulación a 100 Hz) (Always et al., 1987; Behm et al., 1994; MacIntosch et al., 2002; O’leary et al., 1998).
Otra forma exógena de estimulación se produce mediante impulsos eléctricos repetitivos de baja frecuencia. En este caso estaremos hablando de escalera, o también llamado en inglés “Treppe o staircase potentiation”. (Por ejemplo una estimulación repetitiva de baja frecuencia a 10 Hz.) (Klug et al., 1982; MacIntosch et al., 2002; Rassier et al., 2002; Tubman et al., 1996).
Así se ha referido el término potenciación para referirse colectivamente a cualquier condición en la cual la respuesta contráctil muscular está incrementada como consecuencia de una estimulación previa (MacIntosch B. y Rassier, 2002; Rassier et al., 2000).
De esta manera, considerando que ambas potenciación y fatiga resultan de una activación previa, es razonable suponer que estos dos procesos, acontecidos al inicio de la actividad contráctil, pueden coexistir durante y luego de la estimulación. Así la expresión “coexistencia de potenciación y fatiga” es usada para referir una situación en la cual las causas y consecuencias subyacentes de potenciación y fatiga están presentes simultáneamente en un momento dado.
Atendiendo a la bibliografía, existen tres parámetros descriptores de la potenciación (Hamada et al., 2000):
- Fuerza isométrica máxima (FIM).
- El intervalo de tiempo transcurrido desde que comienza a generarse la fuerza hasta que la fuerza isométrica máxima se alcanza (TFIM).
- El intervalo de tiempo que transcurre desde que se alcanza la FIM hasta que, por la relajación muscular que conlleva el cese de la activación, se llega a la mitad del valor de la FIM. A esto se lo denomina tiempo medio de relajación o (TR1/2).
Así, de esta manera, podemos observar que el fenómeno de potenciación produce un aumento en la FIM y una disminución en el TFIM y en el TR1/2. (Always et al., 1987; Hamada et al., 2003; Hamada et al., 2000; Vandenboom et al., 1997). En la figura 17 se pueden observar los diversos parámetros descriptores de la potenciación utilizando la sacudida muscular en estado normal y en estado potencia.
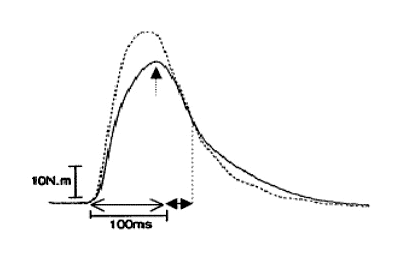
Figura 17. Parámetros descriptores de una sacudida muscular sobre la curva F-T.
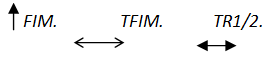
Sea cual fuere el método para inducir la potenciación, se pueden observar tres fases para su determinación:
- Fase 1: Evaluación del comportamiento mecánico del grupo o grupos musculares, en estado no potenciado.
- Fase 2: Aplicación de estímulo desencadenante de la potenciación.
- Fase 3: Evaluación del comportamiento mecánico del grupo muscular en estado potenciado.
Mecanismo responsable de la Potenciación
Antes de comenzar a hablar sobre el mecanismo fisiológico responsable del fenómeno fisiológico de la potenciación vamos ha hacer un breve resumen de la composición de una molécula de proteína.
La molécula se compone estructuralmente de dos hilos de proteínas juntos y enrollados. Ésta está compuesta por seis cadenas polipeptídicas, dos cadenas pesadas, cada una con un peso molecular de 220 kDa. (kiloDalton) y cuatro cadenas ligeras o livianas con pesos moleculares de alrededor de 20 kDa. cada una.
Un extremo de cada una de éstas cadenas pesadas está plegado en una estructura polipéptida globulosa llamada cabeza de miosina. Por tanto, existen dos cabezas libres, situadas una al lado de la otra, en un extremo de la molécula de miosina de doble hélice y la porción larga de la molécula enrollada se denomina cola.
Las cuatro cadenas livianas forman también parte de las cabezas de miosina, dos en cada cabeza. Estas cadenas livianas ayudan a controlar la función de la cabeza durante la contracción muscular.
A su vez cada cadena liviana de miosina se divide en dos tipos las CLM esenciales y las reguladoras. Las CLM esenciales se denominan de éste modo por la solidez estructural que ésta le confiere a la molécula de miosina y se encuentran en una zona más proximal a la cabeza de miosina que la CLM reguladora (CLMr). En la figura 18 se puede observar el esquema de una molécula de miosina.
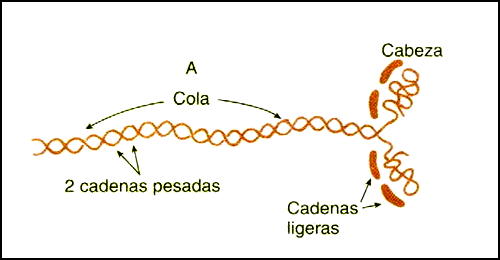
Figura 18. Esquema de una molécula de miosina. Extraído de Tratado de fisiología médica. Guyton ‘99.
Existe considerable evidencia científica de que el fenómeno fisiológico de la potenciación resulta de la fosforilación de las cadenas livianas de miosina reguladoras (FCLR), (Grange et al., 1995; Houston, 1987; Sweeney et al., 1993). Numerosos estudios han mostrado la existencia de una significativa correlación entre la FCLR y la magnitud de la potenciación (Klug et al., 1982; Moore et al., 1984).
Tomando estas observaciones se obtiene una firme evidencia de que el mecanismo principal de la potenciación es la fosforilación de las CLMr.
Todo este proceso comienza con una liberación de Calcio del retículo sarcoplásmico. Este calcio una vez dentro del citoplasma de la célula se une a una proteína llamada calmodulina de esta manera se forma un complejo denominado calcio – calmodulina (Moore R. y Stull, 1984). Este complejo calcio – calmodulina activa una enzima llamada Kinasa de las cadenas livianas de miosina (KCLM) transformándolas de una forma inactiva a una activa (Persechini et al., 1985). Así esta enzima cataliza la incorporación de fosfatos por las cadenas ligeras de miosina provocando de esta manera la fosforilación de las mismas y alterando el estado de los puentes cruzados de acuerdo al estado de fosforilación.
Del mismo modo, una disminución en las concentraciones de calcio dentro del citoplasama, debida a una gran recaptación de calcio producida por el retículo sarcoplásmico, produce una separación entre el calcio y la calmodulina, transformando la KCLM en una forma inactiva. Por otro lado, la magnitud de fosforilación del la CLMr, en última instancia, no dependerá solamente de la activación de la KCLM sino también de la desfosfatasa de las CLM, ésta es una enzima perteneciente a la clase 1 de las proteínas fosfatasas. Su función es remover los grupos fosfatos de la CLM (Klug et al., 1982; Moore R. y Stull, 1984). En la figura 19 podemos observar un esquema del proceso de fosforilación de las CLMr.
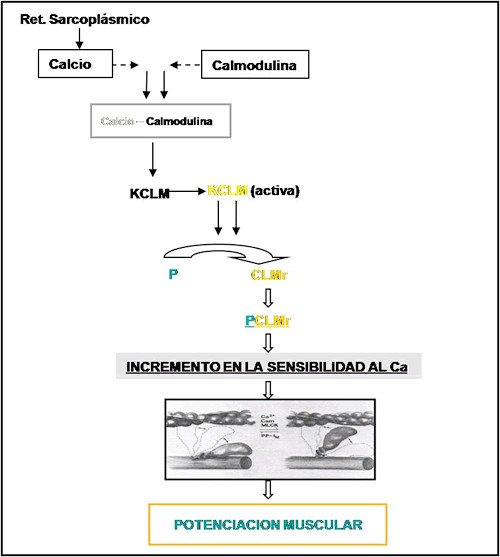
Figura 19. Esquema del proceso de fosforilación de las cadenas livianas de miosina.
Aunque la fosforilación de las CLMr es un proceso relativamente rápido, la reacción contraria, o sea, la desfosforilación requiere varios minutos (Vandenboom, 1995), así, la desfosfatasa de la CLMr actúa a un ritmo relativamente lento, alcanzándose niveles de fosforilación de reposo tras 4-5 min en músculos de mamíferos a 37ºC (Sweeney ,1993).
La fosforilación de las CLR produce un alejamiento de las cabezas de los puentes cruzados separándolas de la columna vertebral del filamento grueso disminuyendo la distancia existente entre el filamento fino y el grueso, aumentando de este modo la velocidad a la cual pueden acercarse a la actina, respondiendo con mayor fuerza a niveles submáximos de calcio sarcoplasmático que un músculo en estado de reposo, con las CLMr desfosforiladas descansando en una disposición ordenada y próximas al eje del filamento de miosina (Sweeney et al., 1993).
Desde este punto de vista, éste mecanismo modulador tiende a aumentar la economía, ya que cuando la CLMr se encuentra fosforilada la activación requiere un menor incremento en la concentración de calcio sarcoplasmático, por lo tanto una menor concentración de calcio deberá ser recapturado por éste, lo cual constituirá un menor coste energético y una relajación más rápida.
Por otro lado, un músculo recientemente activado, con las CLM fosforiladas respondería más rápidamente y con mayor cantidad de fuerza, a concentraciones submáximas de calcio dada la incrementa sensibilidad al mismo (Young, 1998; O’Leary et al., 1997; Fowles 2003).
Del mismo modo, otro de los efectos producidos por la fosforilación de las CLMr sería la producción de una mayor cantidad de puentes cruzados activos. Esto también podría explicar en parte el aumento de la fuerza y de la velocidad de desarrollo de la fuerza en el músculo esquelético de mamíferos. (O’leary et al., 1997; Sweeney et al., 1986; Vandenboom et al., 1995).
Así mismo, estudios con músculos atrofiados aportan argumentos que sostienen a la fosforilación de las cadenas livianas de miosina (FCLM) como el principal mecanismo responsable de la potenciación (Tubman et al., 1996; Tubman, 1997). Estas investigaciones muestran una disminución de la FCLM y la disminución de la potenciación en el músculo gastronemio de ratas atrofiado. La asociación de estos hallazgos proporciona evidencia indirecta de que la FCLM es el mecanismo más importante de la potenciación.
Por otro lado, la evidencia de que la FCLM es el mecanismo principal de la potenciación no excluye la participación de otros mecanismos. De esta manera el calcio puede influir directamente sobre la potenciación dado el importante papel que éste tiene en el proceso de FCLM (Always et al., 1987).
Por su parte (Rassier et al., 1999) muestra que la potenciación será mayor, para un nivel dado de fosforilación de las CLM, cuanto más incrementada esté la concentración de calcio.
A estos resultados se suman los datos aportados por Persechini et al., 1985 quienes encontraron que la potenciación es debida a la FCLM a pesar de ser sensible a la concentración de calcio citoplasmático. Por todo esto es que se deduce que el calcio tiene un papel modulador de la magnitud de la potenciación (Hamada et al., 2000; Rassier et al., 1997).
A su vez Vandenboom et al., 1997 proponen que la FCLM aumenta la sensibilidad de los puentes cruzados al calcio, mejorando de esta manera la velocidad de desarrollo de la fuerza de sujetos en estado potenciado.
Otros autores proponen que el fundamento fisiológico de la potenciación se sustenta en un efecto de carácter neural. Verkhoshansky explica las causas de dicho incremento sobre la base de la teoría del “efecto de retardo de la actividad muscular”. Para este autor, cualquier estímulo previo a una acción motriz, sea momentáneo o no, genera una “huella en el sistema nervioso” que permite mantener los niveles de fuerza obtenidos durante un cierto tiempo y aun mejorarlos.
El concepto que surge del método por contrastes, según Verkhoshansky (2000), indica que la aplicación de una actividad previa influye en el funcionamiento posterior del músculo esquelético. Sin dudas, lo más cuestionable, del método es la comprobación de la existencia real de una “huella neuromuscular” que explique en su totalidad al fenómeno.
Según Gullich y Schmidtbleicher (1995) la potenciación muscular se produce por el desencadenamiento del reflejo H (Reflejo de Hoffman), el cual puede ser evaluado a través de electromiografía midiendo la amplitud de la onda H. Este reflejo produce la acumulación de potenciales de acción en la placa neuromuscular, lo cual desencadena el reclutamiento de una mayor cantidad de unidades motoras, luego de la contracción voluntaria máxima (CVM).
Estos autores plantean que no es posible reclutar todas las unidades motoras con acciones motoras voluntarias, sino que quedan unidades motoras de reserva (sin activar). De esta manera numerosas sinapsis se proyectan sobre las alfa motoneuronas. El potencial de acción es transmitido solo si la cantidad de neurotransmisores es suficiente junto con una adecuada acumulación de calcio en las células pre y post sinapsis, lo cual produce una mayor sensibilidad del receptor post sináptico y esta mayor sensibilidad en las motoneuronas sería en última instancia lo que permitiría que ante un estímulo determinado se reclute una mayor cantidad de unidades motoras.
Desde esta perspectiva las unidades motoras de reserva que se activan como consecuencia de la potenciación son las de mayor tamaño (correspondientes a las fibras FT), o sea que el patrón de reclutamiento coincide con el principio del tamaño o de la talla descripto por Henneman en el año 1965, por lo tanto, las unidades motoras y las fibras musculares extras que se activarán como resultado de la potenciación, siempre serán las que posean el umbral exitatorio más elevado, o sea, las fibras musculares FT.
Cuando se aplica un estímulo desencadenante de potenciación, éste estímulo en el mismo momento también provoca fatiga.
Existe una interacción o coexistencia entre potenciación y fatiga luego de la aplicación del estímulo desencadenante de la potenciación (Rankin et al., 1988; Rassier et al., 2000).
O’Leary, (1997) muestra una disminución del 15% de la FIM durante la contracción tetánica de 7 segundos.
Ante esto Alway (1987) plantea que la potenciación y la fatiga acontecen en ubicaciones diferentes dentro de los procesos de excitación – contracción.
Por su parte Sale (2002) propone que luego del estímulo desencadenante de la PAP comienza a coexistir la potenciación y la fatiga, pero la fatiga se disipa más rápidamente que la potenciación o dicho de otro modo la fatiga se recupera antes de que la potenciación decaiga excediendo de este modo el rendimiento contráctil obtenido previamente (Ver figura 20).
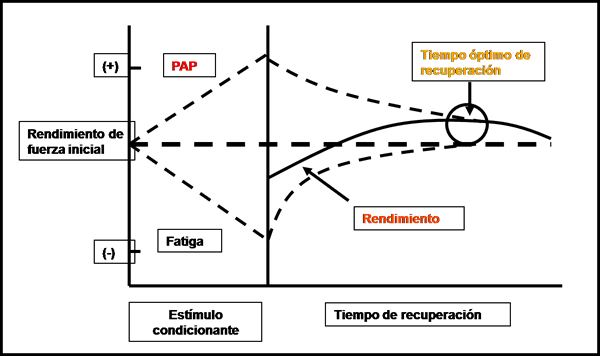
Figura 20. “Competencia” entre pap y fatiga. Modificado de Sale ‘02.
Al respecto Rassier y col (2001) demuestran que sujetos entrenados pueden hacer que la potenciación aumente y la fatiga disminuya en paralelo aún más, dentro de ésta coexistencia de potenciación y fatiga.
Algunos Estudios que encontraron Potenciación
Verhoshansky (2000) cita en su libro “Superentrenamiento” a Murray (1959) como el primero en hallar un incremento directo y estadísticamente significativo en la velocidad de un movimiento sin carga (salto vertical) después de haber ejecutado el mismo movimiento con peso adicional.
Este autor encontró las siguientes observaciones luego de hacer sentadillas con barra.
- La fuerza máxima aumenta significativamente durante el primer minuto, en torno al 25% del nivel inicial.
- Después de 4’ a 5’ la fuerza sigue creciendo hasta un 65% con respecto a la fuerza inicial.
Hasta aquí este fenómeno del retardo de la actividad muscular provocaría un aumento en los niveles de fuerza máxima posteriores al ejercicio aunque cabe aclarar que es muy probable que haya existido algún tipo de error de traducción o tipeo en la trascripción de los datos mostrados por este prestigioso entrenador ya que nos parecen demasiado elevados los porcentajes de mejora luego de la realización del ejercicio de sentadillas. Veamos que sucede con respecto al tiempo en alcanzar dicha fuerza máxima. Analizando esfuerzos isométricos máximos en el tiempo después de hacer sentadillas, observamos que el tiempo requerido para alcanzar la fuerza máxima se redujo un 2.6% y un 4.6% después de 3’ y 4’ respectivamente. Luego aumentaría hasta exceder el límite inicial (figura 1.20).
Por último, Verhoshansky destaca la importancia del tipo de ejercicio que influye en la mejora del trabajo subsiguiente. Por ejemplo la altura del salto vertical varía durante el período de trabajo posterior a sentadillas con barra.
Otro estudio realizado por Verhoshansky muestra un aumento en el salto en largo por encima de los niveles iniciales, después de la sentadilla de un 6.8% a los 3’ – 4’ posteriores.
De la misma manera se presenta un aumento del salto vertical en un 8% a los 8’ – 10’ respectivamente luego del ejercicio de sentadillas.
Al respecto Duthie et al., 2002, concluyen que el entrenamiento a través del método de contrastes produce mayores mejoras en la producción de potencia que el método tradicional. Cabe aclarar que si bien este autor utiliza el nombre de método de entrenamiento de contrastes, no evalúa los resultados de un método de entrenamiento, o sea que no realiza un estudio de carácter longitudinal, sino que evalúa la respuesta muscular en un momento dado, por lo tanto su estudio es de carácter transversal. A su vez, el mismo autor, muestra que individuos con altos niveles de fuerza obtienen mejores resultados con entrenamiento de contrastes que individuos con bajos niveles de fuerza.
Por su parte, Young et al., 1998, estudiaron el efecto que, sobre el test de contramovimiento con sobrecarga, tiene la realización previa (4’ de pausa) de una serie de media sentadilla utilizando 5 RMs. y comprobaron que se podían conseguir mejoras estadísticamente significativas (p > a 0.05) de un 2.8%.
Young emplea una carga de 5 RM en sentadillas basándose en estudios de Verhoshansky (1986), quien asegura que una carga de tal magnitud provoca una estimulación del sistema nervioso central facilitando un mayor esfuerzo explosivo en ejercicios subsiguientes con cargas livianas.
A su vez, Radcliffe y Radcliffe (1996) consiguen efectos similares al estudiar diferentes modelos de calentamientos, con y sin sobrecargas, antes de realizar un test de salto de longitud a pies juntos. Los autores vieron que al calentar haciendo 4 series de 4 repeticiones de arrancada se conseguían 3.9 cm más que haciendo cualquier otro tipo de calentamiento.
Por su parte Gullich y Schmidtbleicher (1995) realizaron una investigación, ésta se dividía en dos partes y tenía dos objetivos principales:
- Examinar el rendimiento de la fuerza explosiva previo y posterior a una contracción voluntaria máxima (CVM).
- Determinar cuanto de la potenciación podía ser considerada como la consecuencia de un efecto general y cuanto se le podía atribuir a una respuesta neural, a través de evaluaciones de activación neural previo y posterior a una CVM.
Para llevar a cabo la primera parte del presente estudio, se utilizaron 36 atletas que competían en diferentes actividades que requerían fuerza explosiva, de los cuales 22 eran atletas de nivel regional y 14 de nivel nacional o internacional. Con respecto a la primera parte del estudio realizado por Gullich y Schmidtbleicher se midieron saltos con contramovimiento (CMJ) y sin contramovimiento (SJ) antes e inmediatamente después de 5 contracciones voluntarias máximas. Los CMJ y SJ posteriores a la contracción isométrica fueron significativamente aumentados un 2.6% y 3.2% respectivamente comparados con los saltos previos a la contracción isométrica.
Con respecto a la segunda parte del trabajo de investigación, se utilizaron 7 estudiantes de educación física los cuales no tenían experiencias de fuerza y 10 atletas con experiencia en el entrenamiento de la fuerza explosiva; en este caso se procedió a evaluar la fuerza explosiva del músculo sóleo y del músculo gastronemio lateral a través de un dinamómetro electrónico, y se examinó la respuesta del reflejo H (reflejo de Hoffman) mediante electromiografía antes y después de un contracción voluntaria máxima (CVM). Los resultados sugieren que:
- Entre 0 y 1 minuto posterior a la contracción voluntaria máxima, se produce un efecto de depresión post tetánica, o sea que el rendimiento de la potencia en ese intervalo es menor al obtenido en la evaluación previa.
- Entre los 4 y los 11 minutos se produce una potenciación estadísticamente significativa (p<0.05).
- El momento en el cual se produce la potenciación muestra una gran variación interindividual 8.7 ± 3.6min.
- El momento de la potenciación en el sóleo (S) fue similar al del gastronemio lateral (GL) (r = 0.84).
- La potenciación en el sóleo fue menor y consecuentemente la distancia pico a pico de la onda del reflejo H (PH) fue menor en el sóleo (PH-S = 1.32 +/- 0.21 – 32%) que en el Gastronemio lateral (PH-GL = 1.20 +/- 0.20 – 20%).
- Los Atletas entrenados en actividades que requieren de fuerza explosiva mostraron valores de potenciación significativamente más altos en el GL que los estudiantes de educación física (PH-Atletas 1.42 +/- 0.17 - 42%; PH-Estudiantes 1.11+/-0.25 – 11%) (p<0.05).
- Los efectos de la potenciación tenían un efecto más duradero en atleta de alto rendimiento que en estudiantes de educación física (Tiempo de PH- Atletas 8.1 ± 3.6min.; Tiempo de PH- Estudiantes 5.9 ± 3.8min.) (p<0.05).
- Se muestra una correlación de 0.89 entre el momento en el cual se produce un aumento del reflejo H y el momento en el cual se produce el aumento de la potenciación.
Respecto a esto Hodgson y et al., (2005) proponen que si bien el reflejo H puede ser uno de los desencadenantes de la potenciación muscular, pero sería demasiado apresurado concluir que el reflejo H es el principal responsable de la misma dada la escasa cantidad de información que existe al respecto.
Por último trataremos las principales variables que intervienen en la potenciación post – activación.
Variables Intervinientes en la Magnitud de la PAP
Son muchas las variables que intervienen en la potenciación post – activación. De todas ellas solo trataremos las que mayor influencia tienen sobre dicho fenómeno. Por último mencionaremos algunas variables que pueden influir en la PAP.
En primer lugar vamos a tomar al estímulo desencadenante de la potenciación como una posible variable interviniente de la potenciación muscular. En este sentido, Requena, (2005) comparó la cantidad de potenciación producida en el press en banca lanzado utilizando como formas de estimulación la estimulación eléctrica percutánea y la contracción voluntaria máxima isométrica, concluyendo que no existen diferencias estadísticamente significativas entre ambos tipos de estimulación.
Otra de las variables más importantes a tener en cuenta, cuando hablamos de la capacidad de producir PAP, es la intensidad del ejercicio, a este respecto, si bien la mayoría de los autores consideran que se necesitan altas cargas, en relación a un porcentaje de la repetición máxima para producir PAP, Baker, (2003), en un estudio realizado con 16 jugadores de rugby de nivel profesional, encontró que se producía PAP con una carga del 65% de la RM, a este respecto, el autor considera que una carga alta, como por ejemplo el 90% de la RM, es demasiado para los miembros superiores.
En relación a la intensidad de la carga, Rahimi, (2007) investigó los efectos de 2 series de 4 repeticiones al 60%, 70% y 85% de 1RM, sobre el tiempo de sprint en una carrera de 40mts. Los resultados del estudio de Rahimi, indican que la carrera mejoró un 1.9%, 2.77% y 2.98% respectivamente.
En un estudio recientemente publicado por Lowery, (2012), tomó 13 sujetos con experiencia en el entrenamiento de fuerza, utilizó sentadillas como ejercicio estimulador en 3 niveles de intensidades, 5 rep. al 56% de 1RM, 4 rep. al 70% de 1RM y , 3 rep. al 93% de 1RM (cabe aclarar que estas series y repeticiones representaban la misma carga de trabajo) y como forma de evaluar la potenciación producida, se midió la saltabilidad sobre una plataforma de salto previo al ejercicio estimulador, inmediatamente después, y a los 2, 4, 6, 8 y 12 minutos post estimulación.
Los resultados de este estudio, muestran que, a baja intensidad (56% RM), no se apreciaron diferencias entre la evaluación inicial, y ninguna de las evaluaciones posteriores a la estimulación con sentadillas. A moderada intensidad (70% RM), los resultados post estimulación, mostraron potenciación a los 4 minutos, volviendo a los niveles basales en la próxima evaluación a los 8 minutos y por último, a alta intensidad (93% RM), los resultados post estimulación, mostraron potenciación a los 4 minutos y mantuvieron esta potenciación en la próxima evaluación a los 8 min., para volver a niveles basales a los 12 minutos.
Los autores, concluyen en este estudio, que la estimulación con alta intensidad produce potenciación muscular durante un período más prolongado de tiempo que a moderada intensidad.
Por otro lado, otro aspecto a tener en cuenta es el tiempo que tarda en manifestarse la PAP. En un estudio realizado en nuestro laboratorio, y cuyos datos no han sido publicados, se evaluó a 11 levantadores olímpicos de nivel nacional, 14 jugadores de básquetbol juveniles y 25 jugadores de fútbol de 4ta y 5ta división, encontramos el momento donde se encontró la mayor cantidad de potenciación, osciló entre los 2 y los 4 minutos.
Por su parte, Baudry (2004) estudió la cantidad de potenciación producida por diferentes tipos de contracción, este autor evaluó cuatro mujeres y cinco hombres utilizando como forma de estimulación diferentes tipos de contracción muscular (isométrica, concéntrica y excéntrica) no encontrando diferencia estadísticamente significativa entre ellas y concluyendo que el tipo de contracción muscular no afecta la cantidad de potenciación producida.
El principal factor que influye sobre la PAP parece ser el tipo de fibra muscular. Diversos estudios en humanos comprueban como los grupos musculares con los menores tiempos de contracción y mayor proporción de fibras rápidas (fibras de tipo II) muestran la mayor PAP (Hamada et al., 2003; Hamada et al., 2000; O’leary et al., 1997; Chiu L.Z.F. et al.,2004) debido a las diferencias en las propiedades contráctiles de los diferentes tipos de fibras (Vandervoort et al.,1983).
La kinasa de las cadenas livianas de miosina es tres veces mayor en las fibras musculares de tipo II frente a las de tipo I (Russell et al., 1984). Esta es la razón por la cual las fibras de tipo II tienen una mayor capacidad para fosforilar las cadenas livianas, produciendo de este modo mayor cantidad de potenciación (Sweeney et al., 1990).
Desde este punto de vista se destaca el trabajo publicado por Hamada (2000) el cual realiza su investigación con 20 sujetos universitarios medianamente entrenados a los cuales le realiza biopsia muscular sobre el vasto lateral con el objetivo de determinar cuales son los 4 sujetos con mayor cantidad de fibras musculares rápidas o sea de tipo II y cuales son los que poseen mayor cantidad de fibras lentas (tipo I) y menor cantidad de fibras explosivas. Posteriormente comprobó que el grupo que obtuvo mayores niveles de potenciación era el que tenía mayores niveles de fibras rápidas de tipo II como así también y una mayor cantidad de área de superficie ocupada por éstas (p < 0.001).
Otro trabajo muy importante con respecto a la composición muscular en cuanto a diferentes tipos de fibra y potenciación es el de Vandervoort (1983), este autor comparó la cantidad de potenciación producida por dos músculos con composición fibrilar diferente como lo son el sóleo, el cual está compuesto predominantemente por fibras lentas de tipo I y el gastronemio el cual está compuesto principalmente por fibras rápidas de tipo II encontrando mayores niveles de potenciación en el músculo gastronemio (p < 0.05).
Por otro lado, la fatiga es otra de las variables que puede influir en la potenciación muscular. En este sentido Rassier y MacIntosch (2002) sugieren que la disminución de la sensibilidad al calcio puede ser debida a una menor afinidad del calcio a la troponina o a una menor cantidad de fuerza producida por cada puente cruzado; concomitantemente la disminución de la sensibilidad al calcio sería en parte la causal de la fatiga acontecida en los primeros minutos de recuperación luego de realizar ejercicios de intensidad elevada.
Gullich y Schmidtbleicher (1995) aportan un dato muy importante relacionado con las variables que intervienen el la potenciación y está relacionado con el nivel de fatiga previa o el tipo de entrenamiento realizado el día anterior a la ejecución de ejercicios que produzcan potenciación. Éstos autores encontraron que los sujetos que habían realizado una alta carga de entrenamiento anaeróbico láctico el día anterior, habían obtenido resultados negativos o inferiores (una reducción promedio del 3.1%) al comparar la evaluación previa y posterior a una contracción voluntaria máxima.
Otra variable que puede influir sobre los niveles de potenciación alcanzados es el grado de entrenamiento de los sujetos. Al respecto Chiu L.Z.F. (2003) muestra como individuos entrenados en deportes que requieren capacidad de fuera explosiva desarrollan mayores niveles de potenciación que individuos recreacionales, o sea individuos que practican deporte o actividad física de modo recreativo y no profesional o competitivo.
A este respecto, Baker (1993), encontró dentro de su muestra de 16 jugadores de Rugby profesional, que los sujetos con mayores niveles de fuerza generaban mayores niveles de PAP, comparado con los sujetos con menores niveles de fuerza, 6,2% y 0,8% de promedio respectivamente.
Por su parte Hamada (2000) muestra como la cantidad de Potenciación se ve incrementada tanto en entrenamiento de resistencia como entrenamiento de fuerza. Según este autor el entrenamiento de resistencia provocaría, por un lado, una mayor cantidad de fosforilación de las CLMr en las fibras lentas y por otro lado provocaría una mayor resistencia a la fatiga lo cual permitiría la prevalencia de la potenciación en la competencia que ésta realiza con la fatiga. Del mismo modo, los atletas entrenados en la capacidad de fuerza también producen mayores niveles de potenciación muscular y este autor propone que esta diferencia se produce por las adaptaciones típicas del entrenamiento de fuerza de lograr una mayor hipertrofia en las fibras de tipo II con respecto a las de tipo I y por lo tanto una proporción mayor del área muscular estaría ocupada por las fibras rápidas de tipo II.
Del mismo modo, Sale (2002) también propone que sujetos sedentarios producen mayores niveles de potenciación luego de someterse a un período de entrenamiento de la fuerza dado que este tipo de entrenamiento produce una hipertrofia selectiva de fibras rápidas. A continuación se muestra una gráfica con la cantidad de potenciación producida por individuos no antes y después de un periodo de entrenamiento de fuerza explosiva.

Figura 21. Cantidad de potenciación producida por un sujeto sedentario y luego de un período de entrenamiento de fuerza. Modificado de Sale ‘02.
Otra característica que modifica la magnitud de la potenciación es la edad de los sujetos (Vandervoort, 2002). Petrella (1989) en un estudio realizado sobre 12 hombres jóvenes y 11 hombres adultos mayores comprueba que los sujetos de edad avanzada poseen menores valores de PAP que los de menor edad (p < 0.001). Estos autores proponen que la menor PAP en la senectud se debe principalmente a la menor proporción de fibras de tipo II que tiene lugar en esta etapa de la vida. En la figura 22 podemos observar la comparación de valores de potenciación entre sujetos jóvenes y ancianos.

Figura 22. Comparación entre la cantidad de potenciación producida por suetos jovenes y acnianos. Modificado de Petrella ‘89.
Otra variable a considerar es la temperatura muscular. Varios autores consideran a la temperatura muscular como un factor generador de variación en los resultados de la PAP. En este sentido Gossen (2001) muestra el efecto que la temperatura ejerce sobre la potenciación de los músculos flexores dorsales de pie de 10 hombres jóvenes. Este autor comparó la potenciación producida a bajas temperaturas musculares (10ºC), en condiciones de control (21ºC) y a elevadas temperaturas (45ºC), encontrando que a mayor temperatura, mayor potenciación con valores de PPT de 40%, 47% y 52% respectivamente.
Por último consideraremos al género como un posible factor influyente sobre la PAP. Al respecto O’leary, (1998) muestran que el valor de la fuerza isométrica máxima de los flexores dorsales del pie se incrementó un 42% en hombres y un 45% en las mujeres luego de 5” de tetania muscular respecto del valor no potenciado; concluyendo en este estudio que la PAP es similar entre hombres y mujeres; no encontrando de esta manera diferencia significativa entre géneros. Esta ausencia de diferencia en la cantidad de potenciación que puede deberse a que ambos géneros de alguna manera poseen una ventaja con respecto al otro, así, el género femenino se beneficia en poseer una mayor resistencia a la fatiga lo cual haría, de algún modo prevalecer la potenciación. Por otro lado, el género masculino posee la particularidad de poseer un mayor porcentaje de fibras rápidas de tipo II que el género femenino (Simoneau, 1985), lo cual le confiere una mayor posibilidad de generar potenciación muscular.
Referencias
1. Abbate,F., A.J. Sargeant, VerdijkP.W.L. y A.deHaan (2000). Effects of hight-frecuency initial pulses and posttetanic potentiation on power output of skeletal muscle. J. Appl. Physiol. 88:35-40
2. Always S.E., Hughston R.L., Green H. J., Patla, and Frank J.S (1987). Twitch potentiation after fatiguing exercise in man. Eur.J.Physiol.56:461-466
3. Asmussen E (1979). Muscle fatigue. Medical Science and Sports Review, nº11, pp.: 313-321
4. Baudry S. and Duchateau J (2004). Postactivation potentiation in human muscle is not related to the type of maximal conditioning contraction. MuscleNerve 30:328-36
5. Baker, Daniel (2003). Acute Effect of Alternating Heavy and Light Resistances on Power Output During Upper-Body Complex power Training. J Strenght Cond Res. 17(3), 493-497
6. Behm David G. and Sale Digby G (1994). Voluntary and evoked muscle contractile characteristics unactive men and women. . Can. J. Appl. Physiol.19:253-265
7. Boschetti Gianpaolo (2002). ¿Qué es la electroestimulación? . Edit.: Paidotribo
8. Bustos A. F., Cappa D. F., y Bonfanti P. C Influencia de diferentes tipos de fuerza máxima sobre el fenómeno de potenciación muscular. Datos sin publicar
9. Chiu Loren Z. F., Fry Andrew C., Schilling Brian K, Joy Jonson E and WeissLarry W Neuromuscular fatigue and potentiation following two successive hight intensity resistance exercise sessions. Datos en trámites de publicación
10. Chiu Loren Z. F., Fry Andrew C., Schilling Brian K, Joy Jonson E. and Weiss Larry W., and Smith Stacey L (2003). Postactivation potentiation response in athletic and recreationally trained individuals. J. Strenght Cond. Res. 17:671-677
11. Duthie Grant M., Young Warren Band Aitken David A (2002). The acute effects or heavy loads on jump squat performance. An evaluation of the complex and contrast method sof power development. J. Strenght and Cond. Res. 16(4), 530-538
12. Fox, E. L (1973). A simple, accurate technique for predicting maximal aerobic power. J Appl Physiol. 35, 914-916
13. Gossen Rodrich E. Sale Digby G (2000). Effects of postactivation potentiation on dynamic knee extensión performance. Eur. J. Physiol. 83:524-530
14. Gossen Rodrich E., Kerry Allingham, and Sale Digby G (2001). Effects of temperatura on post-tetanic potentiation in human dorsiflexors muscles. Can. J. Phisiol. Pharmacol.79:49-58
15. Grange R. W., Vandenboom R. and Houston M. E (1995). Myosin phosphorylation auguments force-displacement and force-velocity relationships of mouse fast muscle. Am. J. Physiol. 269:713-724
16. Gullich A. and Schmidtbleicher (1995). Short-termpotentiation of power performance induced by maximal voluntary contraccions. In:Book of Abstracts-XVth Congress of the international Society of Biomechanics. Hakkinen K, Keskinen K. L., Komi P. and Mero A., ed. Jyvaskyla. ISB, 348-349
17. Gullich A. Schmidtbleicher D (1996). MCV-induced short-term potentiation of explosive force. NSA. 11(4), 67-81
18. Hamada Taku, Sale Digby G., Mac Dougall Duncan J., and Tarnopolsky Mark A (2003). Interaction of fiber type, potentiation and fatigue un human knee extensor muscles. Acta Physiol.Scand.178:165-173
19. Hamada Taku, Sale Digby G., Mac Dougall Duncan J., and Tarnopolsky Mark A (2000). Postactivation potentiation, fiber type and twitch contraction time in human knee extensors muscles. J. Appl. Physiol. 88:2131-2137
20. Hamada Taku, Sale Digby G. and Mac Dougall Duncan J (2000). Postactivation potentiation in endurance trained male athletes. Med. Sci. Sport Exerc. 32:403-411
21. Heyward, V.H (1998). Advanced fitness assessment & exercise prescription. Champaign, IL, Human Kinetics
22. Hodgson M., Docherty D. and Robins D (2005). Post – activation potentiation underlying physiology and implications for motor performance. Sports med35 (7) 585-595
23. Houston M. E., Lingley M. D., Stuart D. S. and Grange R.W (1987). Myosin light chain phosphorylation in intact human muscle. Fed. Eur. Bioc.Sci. 219:469-471. 1987
24. Klug Gary A., Boterman Barry R, and Stull James T (1982). The effect of low frecuency stimulation of myosin light chain phosphorylation in skeletal muscle. J. Biol. Chem. 257:4688-4690
25. Lowery Ryan P., Nevine M. Duncan, Jeremy P LOeneke, Eric M. Sikorski, Marshall A. Naimo, Lee E. Brown, Floyd G Willson, and Jacob M Wlson (2012). The effects of potentiating Stimuli Intensity under varying rest periods on vertical jump performance and power. J. Strenght Cond. Res. 26(12)/3320-3325
26. Mac Intosch Brian R. and Janine C. Willis (2000). Force-frecuency relationship and potentiation in mammalian skeletalmuscle. J. Appl. Physiol., 88:2088-2096
27. Mac Intosch Brian R. and Shirley N. Bryan (2002). Potentiation of shortening during repeated isotonic titanic contractions in mammalian skeletal muscle. Eur. J. Physiol. 443:804-812
28. Mac Intosch Brian R. and Rassier Dilson E (2002). What is fatigue? . J. Appl. Physiol. 27:42-55
29. Moore Rusell L. and Stull James T (1984). Myosin light chain phosphorylation in fast and slow skeletal muscles in situ. Am. J. Physiol. 247:462-471
30. O’leary Deborah D., Karen Hope, and SaleDigby G (1998). Influence of gender on post-tetanic potentiation in human dorsiflexors. Can. J. Phisiol. Pharmacol.76:772-779
31. O’leary DeborahD., HopeKaren and SaleDigby G (1998). Posttetanic potentiation of human dorsiflexors. J. Appl.Physiol., 83:2131-2138
32. Persechini Antony, Stull James T., and Cooke Roger (1985). The effect of myosin phosphorylation on the contractile properties of skinned rabbit skeletal muscle fibers. J. Biol. Chem. 260:7951-7954
33. Petrella R.J., Cunningham D.A., Vandervoort A.A., and Paterson D.H (1989). Comparison of twitch potentiation in the gastrocnemius of Young and elderly men. Eur. J. Physiol.58:395-399
34. Radcliffe J.C., and Radcliffe L (1996). Effects of different warm-up protocols on peak power output during a single response jump task (abstract). Med.Sci.Sports Exerc.28(5):S189
35. Rankin LucindaL., Enoka Roger M., Volz Kathryn A., and Stuart Douglas G (1998). Coexistence of twitch potentiation and tetanic forcé decline in rath in dlimb muscle. J.Appl.Physiol.65:2687-2695
36. Rassier Dilson E. and Mac Intosch Brian R (2000). Coexistence of potentiation and fatigue in skeletal muscle. J.Med.Biol.Res 33
37. Rassier Dilson E. and Herzog Walter (2002). Effects of potentiation the length-dependent twitch potentiation in skeletal muscle. J. Appl. Physiol. 92:1293-1299
38. Rassier Dilson E., Tubman, L.A. and Mac Intosch Brian R (1997). Inhibition of Ca release in rat atrophied gastrocnemius muscle. Experimental Physiology 82:665-676
39. Rassier Dilson E., Tubman, L.A. and Mac Intosch Brian R (1997). Length-dependent potentiation and myosin light chain phosphorylation in rat gastrocnemius muscle. Am. J. Physiol. 273:198-204
40. Rassier Dilson E. and Mac Intosch Brian R (2002). Sarcomere length-dependence of activity-dependent twitch potentiation in mouse skeletal muscle. BMC Physiology 2-19
41. Rassier Dilson E., Tubman, L.A. and MacIntosch Brian R (1999). Staircase un mammalian muscle without light chain phosphorylation. Braz J. Med. Biol. Res. 32:121-129
42. Rassier Dilson E (2000). The effects of length on fatigue and twitch potentiation in human skeletal muscle. J. C. Physiol. B008-0283
43. Rassier Dilson E. and Walter Herzog (2001). The effects of Training and twitch potentiation in human skeletal muscle. E. J. vol.1 issue3
44. Requena B., Zabala M., Ribas J., Ereline J., Paasuke M., Gonzalez-Badillo JJ (2005). Effect of post-tetanic potentiation of pectoralis and tríceps brachii muscles on bench press performance. J. Strenght Cond. Res. Aug; 19(3):622-7
45. Robbins Daniel W. and David Docherty (2005). Effects of Loading on enhancement of power performance over three consecutive trials. J Strenght Cond Res. Aug; 19(4), 898-902
46. Russell L., Moore and James T. Stull (1984). Myosin light chain phosphorylation in fast and slow skeletal muscles in situ. Am.J.Physiol.247 (CellPhysiol.16):462-471
47. Sale Digby G (2002). Postactivation potentiation: Role in human performance. Excerc. Sport Sci. Rev. 30:138-143
48. Sweeney H.L. and Stull James T (1990). Alteration of cross-bridge kinetics by myosin light chain phosphorylation in rabbit skeletal muscle: Implications for regulation of actin-myosin interactions. Proc. Natl. Acad. Sci. 87414-418
49. Sweeney H.L., Bowman Bonita F., and Stull James T (1993). Myosin light chain phosphorylation in vertebrates striated muscle: regulation and function. Am. J. Physiol. 264:1085-1095
50. Sweeney H. L. and Stull James T (1986). Phosphorylation of myosin in permeabilized mammalian cardiac and skeletal muscle cells. Am. J. Physiol. 250:657-660
51. Tubman, L.A., Rassier Dilson E. and Brian R. MacIntosch (1996). Absence of myosin light chain phosphorylation and twitch potentiation in atrophied skeletal muscle. Can.J.Phisiol.Pharmacol.74:723-728
52. Tubman L.A., Dilson E. Rassier and MacIntosch Brian R (1997). Attenuation of myosin light chain phosphorylation and posttetanic potentiation in atrophied skeletal muscle. Eur. J. Physiol. 434:848-851
53. Tubman, L.A., MacIntosh, B.R.; Maki W.A (1996). Myosin light chain phosphorylation and posttetanic potentiation in fatigued skeletal muscle. Pflugers Arch.,431:882-7
54. Vandenboom R., Xeni J., Bestic N. M., and Houston M.E (1997). Increased force development rates of fatigued mouse skeletal muscle are graded to myosin light chain phosphate content. Am. J. Physiol. 272:1980-1984
55. Vandenboom R., Grange R.W., and Houston M.E (1995). Myosin phosphorylation enhances rate force development in fast-twitch skeletal muscle. Am. J. Physiol.268:596-603
56. Vandervoort A. A., and Mc Comas A.J (1983). A Comparison of the contractile properties of the human gastrocnemius and soleus muscles. Eur. J. Physiol. 51:435-440
57. Vandervoort A.A (2002). Aging of the human neuromuscular system. Muscle and Nerve review 25:17-25
58. Vandervoort A.A., and McComas A.J (1986). Contractile changes in opposing muscles of the human ankle joint with aging. J. Appl. Physiol. 61:361-367
59. Vandervoort A. A., Quilan J., and McComas A. J (1983). Twitch potentiation after voluntary contraction. Exp. Neurol. 81:141-152
60. Verkhoshansky Y (1986). Speed-strength preparation and development of strength endurance of athletes in various specializations. Sov. Sports Rev. 21(3):120-124
61. Verkhoshansky Yuri, Siff Mel C (2000). Superentrenamiento. Ed.:Paidotribo
62. Young Warren B., Jenner Andrew, and Kerrin Griffiths (1998). Acute Enhancement of Power Performance From Heavy Load Squats. J. Strenght and Cond. Res. 12:82-84