
Ritmos Circadianos y Rendimiento Físico: Implicancias para la Adaptación Hormonal y Muscular
Michael R McGuigan3, Teo Weipeng1 y Michael J. Newton2
1School of Exercise, Biomedical and Health Sciences, Edith Cowan University, Joondalup, WA, Australia.
2New Zealand Academy of Sport North Island, Auckland, New Zealand.
3Sport Performance Research Institute New Zealand, Auckland University of Technology, Auckland, New Zealand.
Artículo publicado en el journal PubliCE del año 2011.
Publicado 12 de diciembre de 2014
Resumen
Palabras clave: variación diurna, hormonas esteroides, adaptación neuromuscular
INTRODUCCIÓN
El concepto de ritmos circadianos (CR) en el rendimiento físico de los humanos ha sido extensivamente estudiado (Atkinson and Reilly, 1996; Drust et al., 2005; Redlin and Mrosovsky, 1997; Reilly, 1990). Las actividades físicas relacionadas con la aptitud aeróbica, la aptitud anaeróbica y las destrezas motoras finas y gruesas han mostrado un claro CR (Bessot et al., 2007; Kline et al., 2007; Reilly et al., 2007). Debido a esto, existe un gran interés por dilucidar los mecanismos responsables de esta variación en el rendimiento físico a lo largo del día. En humanos, el marcapasos circadiano principal es el núcleo supraquiasmático (SCN). El SCN, ubicado en el hipotálamo, recibe información directa respecto del ciclo solar desde la retina (Hastings and Herzog, 2004). Con esta información provista a través de la vía retino-hipotalámica, el SCN coordina los ritmos biológicos diarios (i.e., la secreción hormonal, las fluctuaciones en la temperatura, la activación neural) conjuntamente con el tiempo solar y los ciclos de sueño/vigilia (Buijs et al., 2003; Waterhouse et al., 2005). Estas oscilaciones rítmicas de los procesos biológicos gobiernan muchos de nuestros hábitos y acciones, y también influencian las actividades que realizamos durante el día. Muchas funciones fisiológicas asociadas con el rendimiento deportivo también han mostrado seguir un CR específico (Winget et al., 1985). Funciones tales como los niveles de reposo del rendimiento sensomotor, perceptual y cognitivo, y diversas variables neuromusculares, comportamentales, cardiovasculares y metabólicas han mostrado alcanzar su pico en las primeras horas de la tarde, conjuntamente con el pico en el ritmo de temperatura corporal (Cappaert, 1999).
Además de las variaciones diurnas en los sistemas fisiológicos, el momento preferencial del día o la noche para realizar actividades es otro factor psicológico importante que debe considerarse cuando se estudian los CR en relación con el rendimiento físico. Dicho concepto ha sido ampliamente reconocido ya que Keitman (1949) ha documentado que algunos individuos tienen una consistente preferencia por realizar actividades durante el día mientras que otros tienen preferencia por realizar actividades durante la noche. La contrastante preferencia temporal entre los individuos (lo que se conoce comúnmente como “cronotipos”) ha mostrado diferir en diversos ritmos fisiológicos tales como los patrones de sueño/vigilia, los biorritmos (i.e., la temperatura central y los niveles hormonales), la inercia del sueño, los ritmos de ingesta de alimentos y el consumo máximo de oxígeno durante el ejercicio (Baehr et al., 2000; Hill et al., 1988; Kerkhof, 1985). Debido al amplio espectro preferencial para realizar actividades en horas de la mañana o la tarde, se cree que este rasgo refleja la habilidad o inhabilidad subyacente para reaccionar a los diferentes sistemas circadianos (Chelminski et al., 1997). Además, Youngstedt & O’Connor (1999) identificaron otras siete variables, que podrían contribuir al menor rendimiento en horas de la mañana y que podrían ayudar a explicar el CR del rendimiento físico. Entre estas variables se encuentran las diferencias en el estatus nutricional entre la mañana y la tarde, la reducción de la flexibilidad en horas de la mañana, el tiempo insuficiente de recuperación a partir de la inercia del sueño, el tiempo preferido de entrenamiento, las diferencias en la cantidad de horas de recuperación entre sesiones de evaluación, las diferencias individuales en las respuestas fisiológicas y las diferencias en los efectos de la motivación y las expectativas.
Los efectos de la testosterona (T) sobre la fuerza y las adaptaciones musculares al ejercicio están bien documentadas (Bhasin et al., 2001; Sinha-Hikim et al., 2002). Una de las principales funciones de la T es mantener el anabolismo, promoviendo la síntesis proteica dentro del sistema muscular (Ferrando et al., 1998). Evidencia provista por Kvorning et al (2006) respalda la hipótesis de que la supresión de la T endógena atenúa las adaptaciones al entrenamiento de la fuerza en hombres saludables, mostrando así la importancia de esta hormona para provocar adaptaciones musculares. En circunstancias normales, el perfil circadiano de la T exhibe un máximo en las primeras horas de la mañana antes de declinar lentamente a medida que progresa el día (Guignard et al., 1980). En contraste, el cortisol (C) es un glucocorticoide que con frecuencia se utiliza como marcador tanto del estrés fisiológico como psicológico. La elevación prolongada del C ha mostrado provocar un efecto inhibitorio sobre el sistema neuromuscular. Tafet et al (2001) reportaron una correlación negativa entre el rendimiento físico y niveles sostenidamente altos de C en saliva. El ritmo circadiano del C es similar al de la T, exhibiendo un pico en horas de la mañana antes de declinar lentamente a lo largo del día y elevándose nuevamente en las primeras horas de sueño (Guignard, et al., 1980). Así como ambas hormonas esteroides y el rendimiento físico exhiben un CR característico y debido a que estos factores tienen fuertes implicancias para la adaptación al ejercicio, es posible que exista una relación entre ambas variables, a pesar de tener un patrón de CR inverso (Hayes et al., 2010).
Por lo tanto, el objetivo de la presente revisión es señalar los mecanismos subyacentes al CR del ejercicio, y los efectos que este tiene sobre el rendimiento de ejercicio. Además, en esta revisión se examinará la relación entre el CR y las hormonas esteroides, T y C, y el CR del rendimiento físico.
EVIDENCIA DEL CR EN EL RENDIMIENTO FÍSICO
Existen diversos factores subyacentes que pueden contribuir al CR del rendimiento físico. Estos factores pueden tener su origen en los cambios que se producen tanto a nivel interno (fisiológico) como externo (ambiental) a lo largo del día. Identificar una única causa de la fluctuación en el rendimiento es extremadamente difícil ya que la fluctuación en el rendimiento puede verse afectada por diferentes sistemas fisiológicos que actúan al mismo tiempo. Reilly & Waterhouse (2009) identificaron tres factores que podrían ser determinantes del CR en el rendimiento deportivo. El primer factor determinante considera las influencias externas (ambientales) que comúnmente son incontrolables. Estos factores incluyen, pero no están limitados a, la temperatura ambiente y la estimulación física o psicológica del ambiente. El segundo factor determinante incluye las influencias internas (fisiológicas) que tienen su origen en el ambiente interno del individuo (i.e., el ritmo biológico del individuo y su capacidad para adaptarse a este ritmo). Por último, también deben considerarse las influencias del estilo de vida (psico-biológicas) que afectan el horario preferencial en el que se realizan las actividades, los patrones de sueño y la capacidad para manejar la inercia del sueño.
Con frecuencia es difícil controlar los factores que afectan el rendimiento físico, y en algunos casos, es una tarea casi imposible. Los factores ambientales, ya sean positivos o negativos, son inevitables en cualquier competencia o sesión de ejercicios y comúnmente son un aspecto incontrolable. Sin embargo, los mecanismos endógenos, pueden manipularse a través del entrenamiento y, en estos casos, el conocimiento de los mecanismos del CR puede ser de utilidad para provocar adaptaciones positivas. Como se mencionara previamente, el CR del rendimiento físico, con puntos bajos por las mañanas y picos por las tardes, son hallazgos comunes en tests de laboratorio y campo (para un resumen ver Tabla 1). Tradicionalmente, la temperatura corporal central se ha utilizado como el principal indicador del CR en los procesos biológicos y del rendimiento físico. Un incremento en la temperatura corporal puede derivar en un incremento en la utilización de carbohidratos, por sobre las grasas, como fuente combustible; y también podría facilitar el mecanismo de formación de puentes cruzados de actina y miosina dentro de la unidad musculo-esquelética (Starkie et al., 1999). Por esta razón, se ha postulado que el pico de rendimiento se produce alrededor de las primeras horas del atardecer, ya que coincide con el pico en la temperatura corporal (Cappaert, 1999). En un intento por demostrar los efectos del incremento en la temperatura corporal sobre el rendimiento de ejercicio, en un reciente estudio llevado a cabo por Taylor et al. (2011) se observó que, mediante la prolongación de la entrada en calor durante sesiones de evaluación realizadas por las mañanas, se producía una atenuación de la reducción de la potencia y la fuerza en saltos con contramovimiento. Al adicionar 20 minutos de entrada en calor activa a una entrada en calor estandarizada, estos investigadores fueron capaces de provocar un incremento en la temperatura corporal comparable al observado durante las sesiones de evaluación realizadas por las tardes. Por lo tanto, se concluyó que el incremento en la temperatura corporal fue el responsable del incremento en la producción de potencia y en otras variables asociadas con el salto. Los resultados del estudio de Taylor et al (2001) coinciden parcialmente con los de un estudio previo llevado a cabo por Atkinson et al (2005), quienes examinaron la influencia de la entrada en calor sobre el rendimiento en una prueba de ciclismo contra reloj. Los resultados de este estudio confirmaron que la entrada en calor mejoró el rendimiento en las pruebas contra reloj llevadas a cabo en dos horarios diferentes del día, pero el tiempo medio de ciclismo fue mayor en la prueba de las 07:30 hs que en el de la prueba de las 17:30 hs; incluso luego de la entrada en calor. La temperatura intra-aural fue mayor en todas las pruebas llevadas a cabo a las 17:30 hs, ya sea que los ciclistas realizaran o no la entrada en calor. Estos resultados sugieren que puede observarse un rendimiento físico superior durante las horas de la tarde, aun cuando los sujetos realizaron una entrada en calor vigorosa de 25 minutos en las sesiones de la mañana. Los resultados del estudio de Souissi et al. (2007) también confirman un efecto de la hora del día respecto de la contribución aeróbica al ejercicio de alta intensidad. En este estudio se comparó la potencia pico, la potencia media, el trabajo total realizado y el consumo de oxígeno entre dos sesiones de evaluación llevadas a cabo por la mañana y la tarde, utilizando el test de Wingate. Se halló que la contribución aeróbica fue mayor en la sesión de la tarde conjuntamente con una incrementada temperatura corporal. Además, la pérdida de potencia fue mayor en la sesión de la mañana que en la de la tarde. A partir de estos experimentos, parece claro que existe una relación entre el CR de la temperatura corporal y las capacidades de rendimiento físico, resaltando así la importancia de realizar entradas en calor activas durante las sesiones de entrenamiento matutinas o en climas fríos, para mejorar el rendimiento físico.
Si bien la temperatura corporal aun es aceptada como el indicador circadiano principal del rendimiento físico, reciente evidencia ha desafiado la visión tradición de la relación entre la temperatura corporal y el rendimiento de ejercicio. Estudios recientes que han investigado los efectos de las diferentes horas del día sobre el rendimiento neuromuscular han revelado un CR característico en variables fisiológicas independientes de los cambios en la temperatura. Por ejemplo, Martin et al. (1999) quienes investigaron el efecto del CR sobre la activación neural y las propiedades contráctiles del músculo aductor del pulgar en humanos. Sus hallazgos indicaron que la fuerza producida durante una contracción voluntaria máxima (MVC) fue mayor en horas de la tarde que en horas de la mañana. Debido a que el incremento en la fuerza de una MVC y de contracciones inducidas eléctricamente fue similar, se sugirió que la modulación de los mecanismos contráctiles periféricos fue la responsable de las fluctuaciones diurnas en la fuerza. Estos investigadores también propusieron que el incremento en la producción de fuerza durante la tarde pudo deberse a una incrementada liberación de calcio desde el retículo sarcoplasmático, al incremento de la sensibilidad de las proteínas contráctiles al calcio y a la alteración de la actividad de la miosina ATPasa.
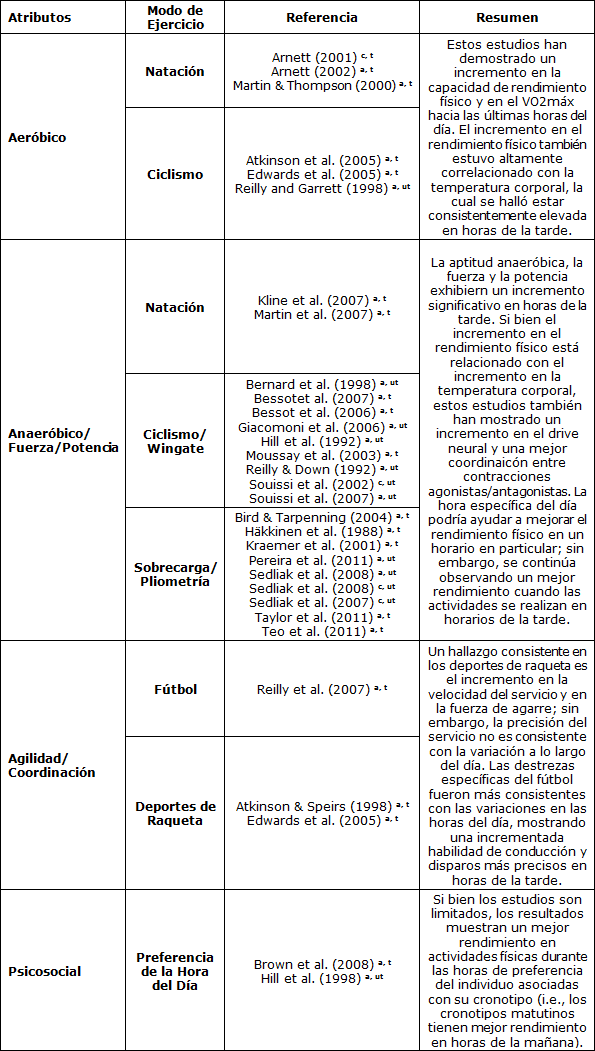
Tabla 1. Efectos del ritmo circadiano sobre el rendimiento físico. (a) Agudo = menos de 4 semanas, (C) crónico = más de 8 semanas; (t) sujetos entrenados = más de 12 meses de experiencia en el entrenamiento, (ut) sujetos desentrenados = menos de 12 meses de experiencia en el entrenamiento.
Un estudio llevado a cabo por Guette et al. (2005) también reportó similares hallazgos, respaldando así la evidencia de que los cambios a nivel muscular pueden ser responsables de las fluctuaciones diurnas en la fuerza. Sus resultados demostraron un efecto significativo de la hora del día sobre el torque de una MVC de los extensores de la rodilla de la pierna dominante y no dominante, hallándose los mayores valores a las 18:00 hs. Estos investigadores sugirieron que debido a que el proceso de formación de puentes cruzados de actina y miosina está mayormente afectado por la concentración de fosfato inorgánico, el cual presenta un CR. La presencia de un CR en el rendimiento muscular también podría explicarse, al menos parcialmente, por las variaciones diurnas en el fosfato inorgánico intracelular.
Los resultados de estos estudios sugieren que existe una variedad de mecanismos involucrados en el CR del rendimiento físico. Estos mecanismos subyacentes comúnmente presentan su propio perfil circadiano característico; sin embargo, casi todos estos muestran un pico en hacia la última parte del día. Es importante señalar que, debido al hecho de que el CR del rendimiento deportivo es afectado por mecanismos multifactoriales, la dilucidación de estos mecanismos es extremadamente difícil.
IMPACTO DEL EJERCICIO SOBRE LA RESPUESTA CIRCADIANA DEL CORTISOL Y LA TESTOSTERONA
Sorprendentemente, existe muy poca evidencia en la literatura que sugiera que el ejercicio, especialmente el entrenamiento de la fuerza, pueda tener algún tipo de influencia sobre el perfil circadiano de la T y del C. Diversas investigaciones han sugerido que la realización del entrenamiento de la fuerza (TST) a una hora específica del día podría resultar en modificaciones de los niveles hormonales de reposo; sin embargo, poco se sabe acerca de los efectos de las adaptaciones a corto y largo plazo sobre el CR de estas hormonas. Un estudio llevado a cabo por Häkkinen et al. (1988) mostró que los cambios en los niveles séricos de T y C debido al entrenamiento a corto plazo podrán ser más indicativos del estrés fisiológico en respuesta al TST que un cambio en el CR. Estos investigadores estudiaron las respuestas adaptativas diarias de los sistemas neuromuscular y endócrino a una semana de entrenamiento intensivo de la fuerza, realizando dos sesiones diarias de entrenamiento con ocho levantadores de pesas de elite. Durante el período de 1 semana, en las sesiones realizadas por la tarde se observó un incremento en los niveles séricos de testosterona libre y testosterona total. Las concentraciones de T total y T libre exhibieron elevaciones en las mediciones de la mañana y la tarde, pero luego de solo un día de reposo, ambos marcadores retornaron a los niveles pre-entrenamiento. Las sesiones de entrenamiento realizadas por las tardes también resultaron en cambios similares en las concentraciones séricas de C y somatotrofina, pero las concentraciones matutinas de C no se vieron significativamente alteradas a lo largo del período de entrenamiento. Los resultados obtenidos por Häkkinen y colaboradores fueron similares a los obtenidos posteriormente por Kraemer et al. (2001). Estos investigadores estudiaron los efectos de un programa de entrenamiento con sobrecarga de alta intensidad sobre el CR de la T en saliva e hipotetizaron que una sesión aguda de entrenamiento en las mañanas no sería suficiente como para alterar el patrón circadiano de la T en saliva. Se recolectaron muestras de saliva de diez hombres entrenados en la fuerza antes, durante y después del entrenamiento y a cada hora luego del entrenamiento, para determinar el patrón circadiano de la T en saliva. Los resultados de este estudio concordaron con la hipótesis establecida por los investigadores, ya que no se observó una modificación significativa del patrón circadiano de la T en saliva. Los resultados de estos estudios sugieren que la influencia de protocolos de entrenamiento a corto plazo es insuficiente como para alterar el perfil circadiano de la T y el C. En vista de estos hallazgos, Sedliak et al. (2007) propuso que se probablemente se requiera de un período de entrenamiento de mayor duración (varias semanas) para observar aluna modificación en los niveles hormonales de reposo. Sedliak et al. (2007) hallaron que un programa de entrenamiento con sobrecarga de 10 semanas de duración modificó las concentraciones séricas y el CR de la testosterona y el cortisol, así como también provocó mejoras en la fuerza isométrica máxima de los extensores de la rodilla. Los resultados de este estudio indicaron que luego de un TST de 10 semanas, las concentraciones séricas de T y C registradas entre las 07:00 y las 20:30 hs declinaron en ambos días de evaluación; sin embargo, la declinación en la concentración sérica de la T no fue significativa.
Debido a la falta de evidencia substancial acerca de los efectos del ejercicio sobre el perfil circadiano de la T y la C, aún queda por determinar si el ejercicio puede, de hecho, ejercer una influencia lo suficientemente fuerte como para alterar el CR de estas hormonas. A partir de la poca evidencia disponible, el ejercicio agudo y el entrenamiento a corto plazo no parece ofrecer el prospecto de alterar el perfil circadiano de la T y el C para inducir algún tipo de adaptación positiva a nivel muscular y en la fuerza. Sin embargo, el TST a largo plazo podría llegar a influenciar, hasta cierto punto, la respuesta hormonal y mejorar las capacidades de rendimiento físico en el momento específico del entrenamiento. No obstante, se requieren más estudios para investigar los efectos de diferentes estímulos de entrenamiento (i.e., tipo de ejercicio, intensidad y relación trabajo/pausa) sobre el CR de la T y el C y también para determinar si un TST de mayor duración (> 10 semanas) podría inducir cambios más permanentes en la T y el C.
IMPACTO DE LA RESPUESTA CIRCADIANA DE LA TESTOSTERONA Y EL CORTISOL SOBRE LAS ADAPTACIONES MUSCULARES Y DE LA FUERZA
En el caso de que un TST no fuera capaz de ejercer algún tipo de influencia sobre el perfil circadiano de la T y el C ¿existiría una relación causal entre el CR de la T y el C y las adaptaciones musculares y de la fuerza? Los estudios que han tratado de examinar la relación entre las fluctuaciones diurnas de la T y el C y las adaptaciones neuromusculares hasta el momento han mostrado resultados contrastantes. Sale et al. (2008), proveyeron evidencia de un efecto negativo del incremento de la concentración de C sobre la función neuromuscular por las mañanas. Estos investigadores compararon la eficiencia del sistema neuromuscular por las mañanas y las tardes al estimular la corteza motora mediante estimulación magnética transcranial y hallaron que los potenciales motores evocados, valorados mediante registros electromiográficos, fueron menores en horas de la mañana lo que fue atribuido a un incremento en los niveles de C. Para confirmar sus hallazgos, los investigadores le administraron a los sujetos hidrocortisona por vía oral, realizando el mismo protocolo en horas de la mañana y de la tarde. Luego de la administración de hidrocortisona por vía oral, se hallaron similares reducciones en el potencial motor evocado. Si bien estos hallazgos demuestran una reducción en la eficiencia neuromuscular, luego de la administración de hidrocortisona, también es muy posible que los neurotransmiores (i.e., GABA, dopamina y serotonina) involucrados en la regulación ascendente de las redes inhibitorias dentro del CNS puedan haber influenciado los potenciales motores evocados durante las sesiones de la mañana ya que estos neurotransmisores también exhiben un CR (Monti and Jantos, 2008; Murillo-Rodriguez et al., 2009). Un estudio llevado a cabo por Bird & Tarpenning (2004) demostró que el CR de los perfiles hormonales puede tener cierto rol en la creación de un ambiente más favorable para el anabolismo, optimizando así las adaptaciones musculares y de fuerza asociadas con el entrenamiento con sobrecarga. Estos investigadores observaron que la realización de ejercicios con sobrecarga de alta intensidad en horas de la tarde alteraba positivamente el perfil del cortisol y el índice testosterona/cortisol (T/C). La concentración de C pre-ejercicio también fue más baja en horas de la tarde, incrementando así el índice T/C y sugiriendo una reducción en el ambiente catabólico que sería beneficiosa para provocar adaptaciones al entrenamiento. Sin embargo, debido al hecho de que en el estudio los sujetos solo fueron evaluados en dos días separados, no es posible determinar si los cambios en el índice T/C fueron permanentes o transitorios. Además, los sujetos utilizados en este estudio fueron individuos que tenían, al menos, 12 meses de experiencia previa en entrenamientos con sobrecarga. Por lo tanto es posible que la alteración positiva en el perfil hormonal estuviera inevitablemente influenciada por el estatus de entrenamiento de los sujetos y la preferencia por un horario de entrenamiento en comparación con un individuo desentrenado (Lusa Cadore et al., 2009).
En contraste, en un reciente estudio llevado a cabo pro Teo et al. (2011) se halló que las variaciones diurnas en la T y la C no mostraron una fuerte relación con el CR de la producción de fuerza y potencia. Utilizando veinte sujetos, con al menos 12 meses de experiencia en el entrenamiento con sobrecarga, se programaron 4 sesiones diarias (08:00, 12:00, 16:00 y 20:00 hs) en cuatro días diferentes para medir la fuerza isométrica, la fuerza dinámica y la potencia. Antes de cada sesión de entrenamiento y en otros tres horarios del día, se recolectaron muestras de saliva para el análisis de las concentraciones de T y C. Si bien ambas respuestas hormonales (T y C) y el rendimiento físico exhibieron un patrón circadiano, no se hallaron correlaciones entre las variables. Estos hallazgos fueron respaldados por los resultados del estudio de West et al. (2010) quienes tampoco observaron relación alguna entre los incrementos de la fuerza y los cambios en las hormonas anabólicas endógenas (hormona del crecimiento, IGF-1 y T). En este estudio se dividieron a los participantes en dos grupos; un grupo realizó flexiones de codo aisladas para mantener las concentraciones hormonales basales y el otro grupo realizó el miso ejercicio de flexiones de codo pero a esto le siguió un entrenamiento de sobrecarga de alto volumen para el tren inferior para provocar un gran incremento en las hormonas endógenas. Luego de 15 semanas de entrenamiento, los incrementos en el área de sección cruzada muscular de las fibras tipo I y II fueron similares en ambos grupos, por lo que los autores concluyeron que el incremento en las hormonas endógenas no fueron responsables de las adaptaciones en la fuerza y el incremento en la hipertrofia.
Similarmente, la noción de que las fluctuaciones diurnas de los perfiles hormonales pueden influenciar los cambios diarios en el rendimiento neuromuscular es dudosa. Las diferencias diarias entre los picos y los nadires de las hormonas anabólicas y catabólicas son naturalmente pequeñas y no es probable que ejerzan un efecto significativo sobre el sistema neuromuscular. Cualquier cambio observable a nivel neuromuscular asociado con cambios en las hormonas anabólicas endógenas, podría estar en realidad influenciado por los protocolos de entrenamiento a largo plazo que probablemente resultaran en la elevación de las hormonas anabólicas a lo largo del día (Ahtiainen et al., 2003). En vista de la contrastante evidencia provista, se sugiere precaución a la hora de interpretar las fluctuaciones diurnas en los perfiles hormonales asociados con el rendimiento físico ya que solo existe limitada evidencia hasta el momento. Aún debe determinarse si existe en efecto una asociación que vincule el CR de la T y el C con las adaptaciones musculares y de la fuerza.
CONCLUSIONES
Esta revisión ha presentado evidencia de que los ritmos biológicos y psicológicos ejercen un efecto principal sobre el rendimiento físico. Si bien la mejora del rendimiento con frecuencia se observa en horas de la tarde, se debe considerar el cronotipo de los individuos. La utilización de una hora específica del día para realizar el entrenamiento parece ser un método efectivo para mejorar el rendimiento físico en un punto particular del día. La utilización de una entrada en calor activa, especialmente en horas de la mañana o en un ambiente frío, es importante para elevar la temperatura corporal previamente a una competencia o entrenamiento. Con respecto a las relaciones entre el CR de las hormonas y el rendimiento físico aún debe determinarse si existe una relación causal. La interpretación de los datos asociados con la modificación del perfil circadiano de las hormonas debe realizarse con cuidado ya que estos cambios podrían ser solo transitorios y podrían estar influenciados por el nivel de experiencia de entrenamiento de los individuos.
Puntos Clave
- Se puede observar un CR característico en el rendimiento físico.
- El CR del rendimiento físico está fuertemente asociado con el CR en la temperatura central corporal.
- Tanto la T como el C exhiben un claro CR; sin embargo, la evidencia actual no muestra una clara relación con las adaptaciones a nivel neuromuscular.
- El entrenamiento con sobrecarga es capaz de provocar cambios en las variables del rendimiento físico en un momento particular del día, pero esto no se debe al perfil circadiano de la T y el C.
Referencias
1. Ahtiainen, J.P., Pakarinen, A., Alen, M., Kraemer, W.J. and Hakkinen, K (2003). Muscle hypertrophy, hormonal adaptations and strength development during strength training in strength-trained and untrained men. European Journal of Applied Physiology 889, 555-563
2. Arnett, M.G (2001). The effect of a morning and afternoon practice schedule on morning and afternoon swim performance. Journal of Strength and Conditioning Research 115, 127-131
3. Arnett, M.G (2002). Effects of prolonged and reduced warm-ups on diurnal variation in body temperature and swim performance. Journal of Strength and Conditioning Research 116, 256-261
4. Atkinson, G. and Reilly, T (1996). Circadian variation in sports performance. Sports Medicine 221, 292-312
5. Atkinson, G. and Speirs, L (1998). Diurnal variation in tennis service. Perceptual and Motor Skills 886, 1335-1338
6. Atkinson, G., Todd, C., Reilly, T. and Waterhouse, J (2005). Diurnal variation in cycling performance: influence of warm-up. Journal of Sport Sciences 223, 321-329
7. Baehr, E.K., Revelle, W. and Eastman, C.I (2000). Individual differences in the phase and amplitude of the human circadian temperature rhythm: with an emphasis on morningness-eveningness. Journal of Sleep Research 99, 117-127
8. Bernard, T., Giacomoni, M., Gavarry, O., Seymat, M. and Falgairette, G (1998). Time-of-day effects in maximal anaerobic leg exercise. European Journal of Applied Physiology and Occupational Physiology 777, 133-138
9. Bessot, N., Moussay, S., Clarys, J. P., Gauthier, A., Sesboüé, B. and Davenne, D (2007). The influence of circadian rhythm on muscle activity and efficient force production during cycling at different pedal rates. Journal of Electromyography and Kinesiology 117, 176-183
10. Bessot, N., Nicolas, A., Moussay, S., Gauthier, A., Sesboüé, B. and Davenne, D (2006). The effect of pedal rate and time of day on the time to exhaustion from high-intensity exercise. Chronobiology International 223, 1009-1024
11. Bird, S.P. and Tarpenning, K.M (2004). Influence of circadian time structure on acute hormonal responses to a single bout of heavy-resistance exercise in weight-trained men. Chronobiology International 221, 131-146
12. Brown, F.M., Neft, E.E. and LaJambe, C.M (2008). Collegiate rowing crew performance varies by morningness-eveningness. Journal of Strength and Conditioning Research 222, 1894-1900
13. Buijs, R.M., van Eden, C.G., Goncharuk, V.D. and Kalsbeek, A (2003). The biological clock tunes the organs of the body: timing by hormones and the autonomic nervous system. Journal of Endocrinology 1117, 17-26
14. Cappaert, T.A (1999). Review: Time of day effect on athletic performance: an update. Journal of Strength and Conditioning Research 113, 412-421
15. Chelminski, I., Ferraro, F.R., Petros, T. and Plaud, J.J (1997). Horne and ostberg questionnaire: A score distribution in a large sample of young adults. Personal Individual Difference 223, 647-652
16. Drust, B., Waterhouse, J., Atkinson, G., Edwards, B. and Reilly, T (2005). Circadian rhythm in sports performance- An update. Chronobiology International 222, 21-44
17. Edwards, B., Lindsay K. and Waterhouse, J (2005). Effect of time of day on the accuracy and consistency of the badminton serve. Ergonomics 448, 1488-1498
18. Edwards, B.J., Edwards, W., Waterhouse, J., Atkinson, G. and Reilly, T (2005). Can cycling performance in an early morning, laboratory-based cycle time-trial be improved by morning exercise the day before? . International Journal of Sports Medicine 226, 651-656
19. Ferrando, A.A., Tipton, K.D., Doyle, D., Phillips, S.M., Cortiella, J. and Wolfe, R.R (1998). Testosterone injection stimulates net protein synthesis but not tissue amino acid transport. American Journal of Physiology2275, E864-871
20. Giacomoni, M., Billaut, F., and Falgairette, G (2006). Effects of the time of day on repeated all-out cycle performance and short-term recovery patterns. International Journal of Sports Medicine 27, 468-474
21. Guette, M., Gondin, J. and Martin, A (2005). Time-of-day effect on the torque and neuromuscular properties of dominant and non-dominant quadriceps femoris. Chronobiology International 222, 541-558
22. Guignard, M.M., Pesquies, P.C., Serrurier, B.D., Merino, D.B. and Reinberg, A.E (1980). Circadian rhythms in plasma levels of cortisol, dehydroepiandrosterone, delta 4-androstenedione, testosterone and dihydrotestosterone of healthy young men. Acta Endocrinologica 994, 536-545
23. Häkkinen, K., Pakarinen, A., Alén, M., Kauhanen, H. and Komi, P.V (1988). Daily hormonal and neuromuscular responses to intensive strength training in 1 week. International Journal of Sports Medicine 99, 422-428
24. Hastings, M.H. and Herzog, E.D (2004). Clock genes, oscillators, and cellular networks in the suprachiasmatic nuclei. Journal of Biological Rhythms 119, 400-413
25. Hayes, L.D., Bickerstaff, G.F. and Baker, J.S (2010). Interactions of Cortisol, Testosterone, and Resistance Training: Influence of Circadian Rhythms. Chronobiology International 227, 675-705
26. Hill, D.W., Borden, D.O., Darnaby, K.M., Hendricks, D.N. and Hill, C.M (1992). Effect of time of day on aerobic and anaerobic responses to high-intensity exercise. Canadian Journal of Sports Science 117, 316-319
27. Hill, D.W., Cureton, K.J., Collins, M.A. and Grisham, S.C (1988). Diurnal variations in responses to exercise of "morning types" and "evening types". Journal of Sports Medicine and Physical Fitness 228, 213-219
28. Kerkhof, G.A (1985). Inter-individual differences in the human circadian system: a review. Biological Psychology220, 83-112
29. Kleitman, N (1949). Biological rhythms and cycles. American Physiological Society 229, 1-30
30. Kline, C.E., Durstine, J.L., Davis, J.M., Moore, T.A., Devlin, T.M., Zielinski, M.R. and Youngstedt, S.D (2007). Circadian variation in swim performance. Journal of Applied Physiology 1102, 641-649
31. Kraemer, W.J., Loebel, C.C., Volek, J.S., Ratamess, N.A., Newton, R.U., Gotshalk, L.A., Duncan, N.D., Mazzetti, S.A., Gomez, A.L., Rubin, M.R., Nindl, B.C. and Hakkinen, K (2001). The effects of heavy resistance training on the circadian rhythm of salivary testosterone in men. European Journal of Applied Physiology 884, 13-18
32. Kvorning, T., Andersen, M., Brixen, K. and Madsen, K (2006). Suppression of endogenous testosterone production attenuates the response to strength training: a randomized, placebo-controlled, and blinded intervention study. American Journal of Physiology. Endocrinology and Metabolism 2291, E1325-1332
33. Lusa Cadore, E., Lhullier, F.L., Arias Brentano, M., Marczwski Da Silva, E., Bueno Ambrosini, M., Spinelli, R., Ferrari Silva, R. and Martins Kruel, L. F (2009). Salivary hormonal responses to resistance exercise in trained and untrained middle-aged men. Journal of Sports Medicine and Physical Fitness 449, 301-307
34. Martin, A., Carpentier, A., Guissard, N., van Hoecke, J. and Duchateau, J (1999). Effect of time of day on force variation in a human muscle. Muscle Nerve 222, 1380-1387
35. Martin, L., Nevill, A.M. and Thompson, K.G (2007). Diurnal variation in swim performance remains, irrespective of training once or twice daily. International Journal of Sports Physiology and Performance 22, 192-200
36. Martin, L. and Thompson, K (2000). Reproducibility of diurnal variation in sub-maximal swimming. International Journal of Sports Medicine 221, 387-392
37. Monti, J.M. and Jantos, H (2008). The roles of dopamine and serotonin, and of their receptors, in regulating sleep and waking. Progress in Brain Research 1172, 625-646
38. Moussay, S., Bessot, N., Gauthier, A., Larue, J., Sesboüe, B. and Davenne, D (2003). Diurnal variations in cycling kinematics. Chronobiology International 220, 879-892
39. Murillo-Rodriguez, E., Arias-Carrion, O., Sanguino-Rodriguez, K., Gonzalez-Arias, M. and Haro, R (2009). Mechanisms of sleep-wake cycle modulation. CNS and Neurological Disorders. Drug Targets 88, 245-253
40. Pereira, R., Machado, M., Ribeiro, W., Russo, A.K., de Paula, A. and Lazo-Osorio, R.A (2011). Variation of Explosive Force at Different Times of Day. Biology of Sport 228, 3-9
41. Redlin, U. and Mrosovsky, N (1997). Exercise and human circadian rhythms: what we know and what we need to know. Chronobiology International 114, 221-229
42. Reilly, T (1990). Human circadian rhythms and exercise. Critical Reviews in Biomedical Engineering 118, 165-180
43. Reilly, T., Atkinson, G., Edwards, B., Waterhouse, J., Farrelly, K. and Fairhurst, E (2007). Diurnal variation in temperature, mental and physical performance, and tasks specifically related to football (soccer). Chronobiology International 224, 507-519
44. Reilly, T. and Down, A (1992). Investigation of circadian rhythms in anaerobic power and capacity of the legs. Journal of Sports Medicine and Physical Fitness 332, 343-347
45. Reilly, T. and Garrett, R (1998). Investigation of diurnal variation in sustained exercise performance. Ergonomics 441, 1085-1094
46. Reilly, T. and Waterhouse, J (2009). Sports performance: is there evidence that the body clock plays a role?. European Journal of Applied Physiology 1106, 321-332
47. Sale, M.V., Ridding, M.C. and Nordstrom, M.A (2008). Cortisol inhibits neuroplasticity induction in human motor cortex. Journal of Neuroscience 228, 8285-8293
48. Sedliak, M., Finni, T., Cheng, S., Haikarainen, T. and Häkkinen, K (2008). Diurnal variation in maximal and submaximal strength, power and neural activation of leg extensors in men: multiple sampling across two consecutive days. International Journal of Sports Medicine 229, 217-224
49. Sedliak, M., Finni, T., Cheng, S., Kraemer, W.J. and Häkkinen, K (2007). Effect of time-of-day-specific strength training on serum hormone concentrations and isometric strength in men. Chronobiology International224, 1159-1177
50. Sedliak, M., Finni, T., Peltonen, J. and Häkkinen, K (2008). Effect of time-of-day-specific strength training on maximum strength and EMG activity of the leg extensors in men. Journal of Sport Sciences 226, 1005-1014
51. Sinha-Hikim, I., Artaza, J., Woodhouse, L., Gonzalez-Cadavid, N., Singh, A.B., Lee, M.I., Storer, T.W., Casaburi, R., Shen, R. and Bhasin, S (2002). Testosterone-induced increase in muscle size in healthy young men is associated with muscle fiber hypertrophy. American Journal of Physiology. Endocrinology and Metabolism2283, E154-164
52. Souissi, N., Bessot, N., Chamari, K., Gauthier, A., Sesboüé, B. and Davenne, D (2007). Effect of time of day on aerobic contribution to the 30-s Wingate test performance. Chronobiology International 224, 739-748
53. Souissi, N., Gauthier, A., Sesboüé, B., Larue, J. and Davenne, D (2002). Effects of regular training at the same time of day on diurnal fluctuations in muscular performance. Journal of Sport Sciences 20, 929-937
54. Starkie, R.L., Hargreaves, M., Lambert, D.L., Proietto, J. and Febbraio, M.A (1999). Effect of temperature on muscle metabolism during submaximal exercise in humans. Experimental Physiology 884, 775-784
55. Tafet, G.E., Idoyaga-Vargas, V.P., Abulafia, D.P., Calandria, J.M., Roffman, S.S., Chiovetta, A. and Shinitzky, M (2001). Correlation between cortisol level and serotonin uptake in patients with chronic stress and depression. Cognitive, Affective and Behavioural Neuroscience 11, 388-393
56. Taylor, K., Cronin, J.B., Gill, N., Chapman, D.W. and Sheppard, J.M (2011). Warm-Up Affects Diurnal Variation in Power Output. International Journal of Sports Medicine 332, 185-189
57. Teo, W., McGuigan, M.R., and Newton, M.J (2011). The effects of circadian rhythmicity of salivary cortisol and testosterone on maximal isometric force, maximal dynamic force and power output. Journal of Strength and Conditioning Research 225, 1538-1545
58. Waterhouse, J., Drust, B., Weinert, D., Edwards, B., Gregson, W., Atkinson, G., Kao, S., Aizawa, S. and Reilly, T (2005). The circadian rhythm of core temperature: origin and some implications for exercise performance. Chronobiology International 222, 207-225
59. West, D. W.D., Burd, N.A., Tang, J.E., Moore, D.R., Staples, A.W., Holwerda, A.M., Baker, S.K. and Phillips, S.M (2010). Elevations in ostensibly anabolic hormones with resistance exercise enhance neither training-induced muscle hypertrophy nor strength of the elbow flexors. Journal of Applied Physiology 1108, 60-67
60. Winget, C.M., DeRoshia, C.W. and Holley, D.C (1985). Circadian rhythms and athletic performance. Medicine and Science in Sports and Exercise 117, 498-516
61. Youngstedt, S.D. and O'Connor, P.J (1999). The influence of air travel on athletic performance. Sports Medicine228, 197-207