
Tipos de Fibras Musculares y su Relación con el Entrenamiento Deportivo
Artículo publicado en el journal PubliCE, Volumen 0 del año 1993.
Publicado 21 de julio de 2003
INTRODUCCIÓN
Al observar una competencia deportiva podemos apreciar distintos niveles de rendimiento; vemos deportistas que son rápidos y también aquellos que son lentos; están los resistentes y los que no los son. Es indudable que estas diferencias las apreciamos desde un punto de vista “externo”, superficial. Sin embargo, el mayor o menor nivel de rendimiento medido por el cronómetro o cinta métrica responde a factores profundos, “internos”, a valores estructurales y funcionales muy íntimos, los cuales se detectan por observaciones de laboratorio, en algunos de los casos altamente sofisticadas. En este sentido y con la finalidad de obtener alto rendimiento deportivo, además de involucrar altos valores de orden funcional como podrían ser los cardiorrespiratorios la exigencia trasciende a la intimidad de la fibra muscular, su metabolismo y estructura molecular. Estos dos aspectos son vitales para determinar niveles de rendimiento: lento o rápido, resistente o potente.
DIFERENCIAS GENERALES ENTRE LAS FIBRAS MUSCULARES
La diferenciación histológica entre las fibras musculares esquelética no es nada nuevo. Ya hace unos cuarenta años, varios histólogos alemanes efectuaron biopsias para determinar a través de ellas, las distintas posibilidades de rendimiento. Es así, que ya entonces, se habló de dos clases de fibras musculares: las “tónicas” por un lado y las “fáscicas” por el otro (Gunther, P.G.; 1950). Sobre las décadas de los 60, las diferencias se establecieron por el aspecto de las fibras bajo la información del microscopio: rojas u oscuras, blancas o claras.
Las rojas u oscuras eran lentas o poco fatigables, mientras que las blancas eran rápidas, pero poco resistentes. A este respecto, cabe mencionar que sobre la misma década de los 60 se empezó a estudiar la actividad de las enzimas, sus distintas funciones y procesos de adaptación. Se pudo constatar un hecho muy importante: que, entre otras causas, la diferenciación funcional entre las fibras musculares responde al comportamiento de la miosina ATPasa (Brooke y Engle, 1969) con el tiempo, comenzaron a desarrollarse metodologías de investigación más sofisticadas, dado que se aplicaron preparados químicos que alteraban el color básico de la fibra muscular; es por tal motivo, que se decidió denominarlas como “slow twitch fiber” o STF (fibras de contracción lenta) y “fast twitch fiber” o FTF (fibras de contracción rápida). Para facilitar su denominación, también se las designó como fibras I a las de contracción lenta y como II a las veloces (Edstrom y Nystroem, 1969). Otros investigadores decidieron diferenciarlas de acuerdo a su funcionabilidad, tal es así, que a las fibras rápidas se las denominó como glucolíticas anaeróbicas, mientras que a las lentas como oxidativas (Peter, Barnard, Edgerton, Gillespie y Stempel, 1962). A partir de la década pasada, se determinó que si bien la división entre fibras I (STF) y II (FTF) era correcta, era necesario efectuar determinadas subdivisiones, teniendo en cuenta la variedad de rendimiento deportivo por un lado, y las reacciones ante preparados químicos específicos por el otro. Esto se comprobó mediante la preincubación de las fibras musculares con preparados químicos en tres niveles de pH; los mismos permiten la coloración de la ATPasa con la siguiente clasificación en fibras de tipo I, IIa, IIb, IIc, (Billeter, Eximan y Howald, 1981).
La escala del pH ha sido en este caso de 4.35, 4.61 y 10.80. Otras experiencias han diferenciando básicamente cinco grupos de fibras musculares: I, Ib, IIa, IIb, IIc, mediante preincubación con valores de pH 10.3, 4.6 y 4.3. Actualmente se reconoce que el nivel cualitativo del rendimiento deportivo está sustentado principalmente sobre la estructura del “mosaico” de fibras musculares que componen los distintos grupos musculares. Este mosaico lo conforma la supremacía de determinado grupo de fibra muscular; actualmente se reconoce que esto tiene tanta importancia como el VO2máx, el volumen minuto, el tamaño del corazón, la cantidad de hemoglobina, y teniendo en cuenta la fuerza, la hipertrofia de las fibras musculares. Por este motivo cuando se trata del desarrollo de resistencia los procesos de adaptación se desencadenan primero a nivel de las fibras musculares y recién después sobre el aparato cardiovascular (Howald, 1989).
Según el grupo muscular, existirá predominio de uno de los tipos básicos de fibra muscular por sobre el otro. Por ejemplo el músculo soleo, posee prevalentemente, fibras del grupo I; relativamente poco fatigables, pero también con escasa capacidad de hipertrofia. En general, ocurre los mismos con los músculos antigravitacionales de la espalda (Saltin, Hemeriksson, Nygaard y Andersen, 1977); en cambio, los músculos extensores del codo tienen prevalencia de las fibras II.
Aunque las fibras musculares de un determinado grupo se encuentran mezcladas con otras, de todas maneras poseen las mismas características o reacciones (Buchtal y Schmalbruch, 1980). Las fibras musculares del tipo II poseen una estructura nerviosa de axón o cilindro eje mas grueso, con alta frecuencia de impulsos en la unidad de tiempo, e incluso se reclutan a mayor velocidad que las fibras lentas I: 35 ms para las primeras y 75 ms para las últimas (Gollnick, Armstrong, Saubert, Pihel y Saltin, 1972).
DIFERENCIAS DE ÍNDOLE METABÓLICO Y ESTRUCTURALES ENTRE LAS FIBRAS MUSCULARES
Entre las diferencias más notorias entre ambos tipos de fibras musculares se destaca los reservorios de glucógeno, los cuales predominan en las fibras II con respecto a las I. Este último grupo, en cambio tiene mayor densidad mitocondrial que las II; ello es obvio, desde el momento que son menos fatigables y con predominancia de los procesos oxidativos, las fibras II tienen una correlación lineal con el grado de fatigabilidad (r=0.86, p<0.01): esto determina entonces que el grado o punto de aparición de la fatiga del músculo esta en intima correlación con la superficie de las fibras II (Thortensson y Karlsson, 1976). Cambia el panorama cuándo mayor es la superficie de las fibras I dentro del paquete muscular. En este caso disminuye la aparición de fatiga, se incrementa la duración del trabajo, aunque presenten menor velocidad de contracción y desarrollo de fuerza en relación a las fibras II. Uno de los hechos más importantes que se presentan para justificar dicha diferencia funcional es la actividad del Mg2+, estimulador de la ATPasa. La actividad de esta enzima no es similar para ambos tipos de fibras musculares (Thortensson, Sjodin, Tesch y Karlsson, 1977): las del grupo II tienen mayor actividad, con una más rápida hidrólisis en relación alas del grupo I (Barmard, 1970; Peter, 1972; Kugelberg, 1973; Schantz 1986).
En realidad las enzimas citoplasmáticas muestran una mayor actividad en las fibras II. La relación del Mg2+ (estimulador de la ATPasa) es casi tres veces superior en las fibras II con respecto a las I; aunque no hay mucha diferencia en la actividad de la CPK (1.3/1).
Es indudable, que la mayor actividad de las fibras II, responde a mayores contenidos de determinados substratos, lo que se resume en la Tabla 1.
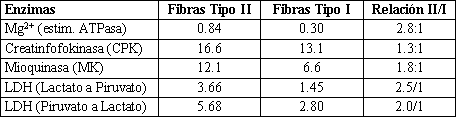
Tabla 1. Valores en moles x 10 gramos de proteínas (Según Thorstensson,
Sjodin, Tesch y Karlsson, 1977)
DIFERENCIA DE ÍNDOLE MUSCULAR ENTRE LAS FIBRAS MUSCULARES
En investigaciones más recientes se han podido constatar diferencias fundamentales desde el punto de vista molecular entre ambos tipos de fibra, inclusive dentro del mismo grupo, la subdivisión surge, no sólo como producto de preincubaciones químicas a distintos niveles de pH, sino también por la distinta estructura molecular.
Las diferencias en este último aspecto, se constatan específicamente a nivel del sarcómero, particularmente en los filamentos gruesos de miosina. No se han constatado deferencias entre las fibras I y II en relación a los filamentos de actina (Billeter, 1981).
Teniendo en cuenta los distintos niveles de rendimiento en el deporte, dicha diferenciación surge, entre otros factores, debido a los distintos encadenamientos de las cadenas peptídicas de la miosina.
Las mismas se dividen entre cadenas pesadas y cadenas livianas. Las cadenas pesadas son de tres tipos:
- Cadena Pesada Rápida A (HCA, Heavy Chain A)
- Cadena Pesa Rápida B (HCB, Heavy Chain B)
- Cadena Pesada Lenta (HCS, Heavy Chain Slow)
Por su parte, las cadenas livianas de la miosina se identifican mediante la siguiente clasificación:
- Cadena Liviana Rápida 1 (LCF1, Light Chain Fast 1)
- Cadena Liviana Rápida 2 (LCF2, Light Chain Fast 2)
- Cadena Liviana Rápida 3 (LCF3, Light Chain Fast 3)
- Cadena Liviana Lenta 1 (LCS1, Light Chain Slow 1)
- Cadena Liviana Lenta 2 (LCS2, Light Chain Slow 2)
Existen diferencias en los pesos moleculares entre las cadenas pesadas y las livianas; las primeras tienen un peso molecular de 200000, mientras que las cadenas livianas varían entre 14600 y 23900. Las cadenas livianas se encuentran ubicadas en la cabeza de la miosina, lo cual es de gran importancia debido a que, precisamente en este lugar se encuentra ubicada la ATPasa, responsable en determinar el deslizamiento de la miosina con respecto a la actina. Las fibras musculares IIa y IIb tienen las mismas tres cadenas livianas rápidas (LCF1, LCF2, LCF3); esto induce a pensar que la sutil diferencia entre ambas puede estar en las dos cadenas pesadas, las cuales, obviamente son rápidas. Concretamente las deferencias se dan a través de distintas secuencias de aminoácidos primarios, las cuales se evidencian en diferentes niveles de la ATPasa durante su coloración histoquímica.
Las diferencias estructurales entre las fibras I con respecto a las II son muy evidentes, no sólo del punto de vista cualitativo sino también cuantitativo. Las fibras de contracción rápida II no poseen cadenas pesadas lentas, sólo rápidas, lo mismo ocurre en relación a las cadenas livianas. La cadena pesada de la fibra I, en cambio, es lenta; las cadenas livianas poseen, no solamente cadenas livianas lentas, sino también rápidas.
El número de las mismas varía, y ello determinaría su distinta funcionabilidad (Billeter, 1981). Se han podido encontrar variadas combinaciones de cadenas livianas lentas y rápidas; en algunos casos se encontraron solamente dos cadenas livianas lentas y dos rápidas. Hay que destacar, nuevamente, que esta variada estructura molecular determina distintos tipos de fibras I (Schanz, 1986).
En las actuales divisiones de fibras musculares también se hace mención a las IIc. ¿Cuales son sus características? Las mismas no son únicamente de contracción rápida, sino además son resistentes debido a que poseen elevada densidad mitocondrial (Howald, 1989). Desde un pinto de vista molecular, se encontró en estas fibras una amplia combinación d e características propias de las fibras IIa, IIB y I. En las fibras IIc, la miosina posee dos cadenas pesadas, una lenta y otra rápida: HCF + HCS. Además de ello, existen tres cadenas livianas rápidas y dos cadenas livianas lentas. Estas características estructurales hacen de las fibras IIc las más versátiles desde el punto de vista funcional, en relación a las restantes. Sin embargo, entre las distintas biopsias efectuadas solo se han encontrado a las mismas en una proporción del 2% con respecto a las restantes (Billeter, 1981). La explicación que se da este fenómeno, es que las fibras IIc podría constituir el paso intermedio entre las fibras I y II (en ambas direcciones y como producto de un proceso de entrenamiento).
CAMBIOS ENTRE LOS DISTINTOS TIPOS DE FIBRA
Hay que reconocer en este aspecto que la fibra muscular posee un optimo grado de plasticidad superior a los que décadas pasadas se sostuvo (Nett, 1955). Es indudable que, para entonces, se consideraron únicamente cambios funcionales sumamente globales, los que se podrían apreciar únicamente por un mejor rendimiento deportivo. Para que se constaten reales variaciones en estos cambios, es necesario realizar extensos estudios longitudinales que abarquen desde meses hasta, quizás, años de entrenamiento muy específicos. De todos modos, con un entrenamiento sistemático que abarque algunas semanas, se constatan íntimos cambios estructurales como también metabólicos.
Estos cambios se producen desde el tipo de fibras II al I, pero no a la inversa (Jolesz, Sreter, 1981), sin que estos cambios enzimáticos o moleculares impliquen una transformación histoquímica del tipo de fibra I o II. Las transformaciones se pueden resumir de la siguiente forma (teniendo en cuenta la aplicación de estímulos lentos):
- la actividad enzimática oxidativa y la densidad capilar pueden incrementarse con una semana de trabajo;
- con dos semanas de entrenamiento se producen cambios en la propiedad contráctil isométrica;
- en la tercera un descenso de la actividad de la ATPasa;
- entre la tercera y sexta semana existen modificaciones a nivel molecular con respecto a la estructura de la miosina y la troponina (Salmons, Henriksson, 1982).
Esto en lo que se refiere a esfuerzos desarrollados en forma relativamente “lenta”, a la manera de la metodología aeróbica, pero no corriendo, sino aplicando estímulos eléctricos de baja frecuencia (10 impulos/seg) y durante un periodo relativamente prologado. ¿Ocurre lo mismo en el camino inverso? ¿Pueden las fibras I adoptar las características del grupo II?. Los hechos y comprobaciones científicas están demostrando que no. Aparentemente, en un primer momento, existe una transformación; las fibras I se hacen “mas veloces”, pero no bien cesa el estímulo, estas fibras retornan a sus características genéticas originales (Salmons, Henriksson, 1981). Esto lo comprendemos desde el momento en que las actividades de la vida diaria de relación constituyen, básicamente, estímulos de bajo umbral de excitación, las cuales están en consonancia con las fibras de contracción lenta: las fibras I están diariamente en su verdadero medio ambiente. Observaciones como estas nos inducen a pensar que los deportistas especialistas en velocidad podrían mejorar sensiblemente su potencia y capacidad aeróbica mediante entrenamientos sistemáticos efectuados a la manera de un fondista, por el contrario, el camino inverso parece improbable.
Cambios Metabólicos Causados por el Entrenamiento
Las modificaciones que pueden tener algunos tipos de fibras, es producto de aquellos cambios que sufren como consecuencia de sistemáticos estímulos específicos a los cuales se las someten. Es por este motivo que, estudios histoquímicos y electromicroscópicos han podido constatar interesantes transformaciones en las fibras como producto del entrenamiento sistemático. Además, es interesante que una misma clase de estímulo puede producir diferentes resultados, según el grupo de fibra muscular de que se trate; en otros casos, en cambio, ambos tipos de fibras tienen cambios similares, pero con estímulos de distintas características. El volumen mitocondrial se incrementa en gran magnitud como producto de entrenamientos sistemáticos, a la manera de los fondistas. Holloszy, 1971, encontró un incremento del 70% del contenido proteico mitocondrial por gramo de músculo, como también un incremento del 100% de la capacidad oxidativa, en coballos entrenados con esfuerzos prolongados. El incremento del volumen mitocondrial se da tanto en e las fibras I como también en las II; sin embargo los más llamativo de ello es que estoa cambios se dan en mayor magnitud en las fibras II con respecto a las I. Además se produce una elevada adaptación a nivel enzimático, con incremento de su actividad. Esfuerzos aeróbicos de baja intensidad modifican la actividad enzimática de las fibras I y el mismo resultado se comprueba en las fibras musculares rápidas II, pero en este último caso, mediante la combinación de trabajos anaeróbicos (Henriksson, Reitmann, 1976). Las enzimas que incrementan su actividad son las oxidativas tales como succinato deshidrogenasa (SDH), succinato oxidasa (SO) y la citocromo oxidasa (Holloszy, 1971; Howald, 1989). Esto podría apoyar la hipótesis que la resistencia y/o potencia aeróbica, no solamente se puede incrementar mediante prolongados esfuerzos de duración, sino también mediante el entrenamiento fraccionado rápido (aeróbico/anaeróbico); usando esta metodología, se tienen muy especialmente en cuenta las fibras musculares II, de contracción rápida y sin desmedro de la velocidad como en el caso anterior. Por otra parte, los trabajos de fuerza muscular activan las enzimas citoplasmáticas tales como la mioquinasa, CPK, ATPasa y PFK. Las fibras musculares de contracción rápida son considerablemente más susceptibles a la hipertrofia que las STI, con mayor síntesis proteica a nivel ribosomal.
De acuerdo a ello cambia la relación FTF/STF en relación a su superficie y en beneficio de las fibras rápidas (FTF). Esto se ha comprobado en levantadores de pesas como también en lanzadores y saltadores (Edstrom y Ekblom, 1972; Pince, 1976).
En conclusión la fibra muscular posee un elevado grado de adaptación, constituyéndose en excelente receptor para métodos específicos de entrenamiento. Sin embargo, también puede ocurrir lo contrario: estos mismos métodos de entrenamiento pueden desadaptar la fibra muscular en razón de no cumplir con los objetivos de la especialidad deportiva. De esta forma, no solamente se modificara su metabolismo, sino que también se altera su estructura molecular. Esto nos lleva a concluir que la preponderancia metodológica del entrenamiento debería estar estructurada, prevalentemente, sobre el mecanismo energético que caracteriza a la especialidad deportiva. Cuánto más no alejamos de este principio tanto mayor será la desadaptación celular, lo que afectará el rendimiento.
Referencias
1. Berenard, R.J., Edgerton, V.R., Peter, J.B (1970). Effect of exercise on skeletal muscle: biochemical and histological properties. J Appl Physiol; 28
2. Billeter, R., Heitzman, C.H., Howald, H (1981). Analysis of myosin light and heavy chain types in single human sekeletal muscle fibers. Eur J Biochem; 116
3. Brooke, M.H., Engel, W.K (1969). The histographic analisys of human muscles biopsies with regard to fiber types I. Adult males and females. Neuorology; 19
4. Buchtal, F. Schmalbruch, H (1980). Motor unit of mammalian muscle. Physiol Rev; 60
5. Edstrom, L. Nystrom, B (1969). Histochemical types and size of fibers in normal human muscles. Acta Neurologica Scandinavica; 45
6. Gollnick, P., Armstrong, R., Saubert, C., Piehl, K., Saltin, B (1972). Enzyme activity and fiber composition in skeletal muscle of untrained and trained men. J Appl Physiol; 33
7. Gunter, P.G (1950). Uber das Vorkommen von tetanischen Fasern in der quergestreiften. Skelettmuskulatur des Menschen
8. Holloszy, J.O., Oscay, L.B., Mole, P.A., Don, I.J (1971). Biomechanical adaptation to endurance exercise in sekeletal muscle. Muscle metabolism during exercise. Penum Press, New York- London
9. Howald, H (1989). Veranderunden, der Meskelfasern durch Training. Leistunsport 2
10. Jolesz, F., Sreter, F.A (1981). Development innervation and activity pattern induced changes in skeletal muscle. Annual Rev Physiol; 43
11. Kugelberg, E (1973). Hisotchemical composition contraction speed and fatigability of rat soleus motor unit. J Neurol Sci; 20
12. Nett, T (1960). Der Lauf, Bartels & Wernitz.
13. Peter, J.B., Bernard, R.J., Edgerton, V.R., Gillespie, C.A., Stempel, K.E (1972). Metabolic profiles of three fibers types skeletal muscles in guinea pigs and rabbits. Biochemestry; 11
14. Prince, F.P., Hikida, R.S., Hagerman, F.C (1976). Muscle fiber types in power lifters, distance runners and untrained subjects. Pflugers Archives, 363
15. Salmons, S. Henriksson, J (1981). The adaptative response of skeletal muscle to increased use. Muscle Nerve; 4
16. Saltin, B., Henriksson, J. Nygaard, E, Anderson, P (1977). Fiber type and metabolic potentials of skeletal muscles in sedentary man and endurance runners. Annals of the New York Academy of Science; 301
17. Schantz, P.G (1986). Influence of physical Training on phenotypic expression of slow and fast isoform of miofibrillar proteins. Acta Physiol Scand; Vol 128, Suppl 558
18. Thortensson, A., Karlsson, J (1976). Fatiganility and fiber composition of human skeletal muscle. Acta Physiol Scand; 98
19. Thortensson, A., Sjodin, B., Tesch, P., Karlsson, J (1977). Actomyosin ATPasa, CPK, and LDH in human fast and slow twitch fibers. . Acta Physiol Scand; 99