
Federación Argentina de Triatlón, Buenos Aires, Argentina.
Resumen
INTRODUCCION
El triatlón conformado por las disciplinas de natación, ciclismo y pedestrismo se desarrolla en distintas modalidades y reglas que imponen distintas exigencias físicas.
El primero en llegar a la meta cumpliendo con las reglas del evento es el ganador. Es necesario conocer los mecanismos y razones de la fatiga para entender como y que estrategias nutricionales deben adoptarse para evitar trastornos que puedan afectar seriamente la salud y que permitan desarrollar el máximo potencial físico.
La nutrición es un factor determinante del desarrollo y rendimiento de los seres vivos y la intervención nutricional aguda antes o durante la competencia ofrece oportunidades para mejorar el rendimiento y las claves son (i) identificar los limitantes del rendimiento, (ii) establecer los objetivos nutricionales que puedan modificar positivamente las limitaciones y (iii) desarrollar estrategias nutricionales para alcanzar esos objetivos (Maughan, R.J., 2003).
En un estudio realizado en un Ironman se observaron sustanciales diferencias en el rendimiento que pudieron relacionarse al consumo de carbohidratos e ingesta de líquidos durante el evento. (Downey, B.M., Hopkins, W.G., 2001).
¿Los que más rindieron, lo hicieron gracias a los nutrientes ingeridos o el rendimiento determinó el hábito de consumo?.
La fatiga es un mecanismo de conservación del organismo tendiente a limitar la perturbación inducida en la homeostasis (Hollman, W., Hettinger, Th., 2000). En la actividad física se manifiesta como la disminución reversible de la capacidad funcional en razón de la actividad muscular (Gandevia, S.C., 2001).
¿Cuáles son los factores que limitan la capacidad de desarrollar una actividad como el triatlón y cuáles son las causas de la fatiga?.
Un extenso análisis de los distintos modelos conceptuales que se han postulado como explicación a las limitaciones del trabajo aeróbico y de la fatiga ha sido realizado por Noakes, T.D. (2000) y una de las conclusiones es que diferentes sistemas fisiológicos pueden ser determinantes del rendimiento en distintas condiciones de ejercicio con un regulador central principal; por esta razón se ha postulado el uso de distintos nutrientes que podrían influenciar el rendimiento sea a niveles centrales desde donde se origina la contracción muscular voluntaria hasta los niveles periféricos.
El presente tiene como objetivo analizar las prácticas y productos nutricionales habituales observados en las competencias de triatlón en Argentina y la evidencia científica actual.
Algunas prácticas nutricionales han sido evaluadas científicamente y han dado lugar a recomendaciones específicas para el triatlón (Jeukendrup, A.E., Jentjens, R.L.P.G., Moseley L., 2005), otras prácticas y productos de uso corriente en Argentina son objeto de esta revisión.
FACTORES DETERMINANTES DEL RENDIMIENTO ALTERABLES POR INTERVENCION NUTRICIONAL AGUDA
Fatiga de Origen Central
Uno de los mecanismos propuestos del desarrollo de fatiga durante el ejercicio es el aumento de la captación por el cerebro de triptofano plasmático, cuya concentración en plasma aumenta a expensas del aumento de ácidos grasos libres que compiten por el mismo sitio de fijación a la albúmina y la disminución de aminoácidos ramificados (BCAA) en plasma que compiten por el mismo sistema transportador de ingreso al cerebro; el aumento de captación de triptofano (TRH) produciría un aumento de la síntesis de 5-OH triptamina –serotonina- (Ganvedia, S.C., 2001), mediador químico a nivel central de la aparición de fatiga y control termoregulatorio a nivel del hipotálamo. La ingesta de carbohidratos (CH) reduce la concentración plasmática de ácidos grasos libres y esto aumentaría la fijación de triptofano a la albúmina, razón por la que se postulo el beneficio de los carbohidratos en la aparición de fatiga (Davis, J.L., et al., 1992). La ingestión de carbohidratos durante actividades aeróbicas submáximas mejora el rendimiento previniendo una hipoglucemia (Coggan, A.R., Coyle, E.F., 1987) y otros factores aún no esclarecidos (Ivy, J.L. et al., 1979; Jeukendrup, A. et al., 1997). Existen razones teóricas y datos a favor de la relación entre BCAA, carbohidratos, serotonina y fatiga central, pero la evidencia científica es débil (Davis, J.M., Alderson, N.L., Welsh, R.S., 2000). La alteración de la relación TRH/BCAA por ingesta de aminoácidos podría reducir la aparición de fatiga; efecto que fue detectado en algunos estudios y en otros no se puedo detectar y se postula que el efecto sería directo y no por interferencia con la serotonina (Blomstrand, E., 2006; Newsholme, E.A., Blomstrand, E., 2006).
A nivel central la estimulación de los receptores de la adenosina son responsables de disminución de la motivación, inducción de sueño, supresión de actividad espontánea y fatiga, cuya aparición es retardada por la cafeína al menos en parte por el bloqueo de estos receptores (Davis, J.M., 2003).
Trastornos Gastrointestinales
El sistema gastrointestinal (GI) está sometido en el ejercicio a una reducción de la circulación y aumento de temperatura. Esto asociado a una deshidratación >3% y temperaturas ambientales elevadas someten al sistema GI a isquemia, hipoxia e hipertermia y puede alterar la barrera intestinal y provocar necrosis tisular, endotoxemia, y trastornos circulatorios (Gisolfi, C.V., 2000).
Sin que determinará la imposibilidad de continuar en la carrera, en una carrera de ultarendurance (160 km) se observó que los atletas que padecieron de trastornos gastrointestinales (en mayor proporción nauseas y vómitos que diarrea y espasmos), fueron los que ingirieron en promedio mayores cantidades de carbohidratos (0.15 g.kg-1.km-1 vs. 0.09 g. kg-1.km-1) (Glace, B.W., Murphy, C.A., McHugh, P.H. , 2002). La observación que la tasa de oxidación de carbohidratos exógeno en el calor disminuye y que los reportes de malestares GI en el calor asociados a la ingesta de CH aumentan, sugiere que la absorción intestinal contribuiría a este efecto sin poder descartar una reducción de la captación de glucosa por el músculo por lo que se sugiere adaptar la ingesta de CH en el calor (50-60g/h), en climas fríos (60-70g/h) (Jentjens, R. L. P. G., Wagenmakers, A.J.M., Jeukendrup, A.E., 2001).
Alteración Mental: Confusión, Alucinaciones
La incidencia de alteraciones mentales, desde confusión hasta alucinaciones, en una carrera de ultarendurance (160 km) estuvo asociada a una menor concentración de Na+ en plasma en relación directa a una mayor ingestión de líquidos, (Glace, B.W., Murphy, C.A., McHugh, P.H., 2002). Las observaciones en el rendimiento de 2135 atletas llevaron a la recomendación que la forma de prevenir la hiponatremia asociada al ejercicio en eventos de fondo prolongados es asegurar que los atletas no se excedan en la ingesta de líquidos (Noakes, T.D, et al., 2005).
Termorregulación
La actividad muscular posee una eficiencia aproximada del 20% en la transformación de la energía química en trabajo muscular y aumenta varias veces la cantidad de calor que el organismo debe disipar para mantener la temperatura corporal. El control de la temperatura está localizado en el hipotálamo y esta involucrado el neurotransmisor serotonina (5-OH Triptofano). Con el aumento de la temperatura ambiental aumenta la participación de la sudoración como mecanismo de enfriamiento y disminuye su efectividad con el aumento de la humedad ambiental. La tolerancia al calor estaría genéticamente determinada.
La hipertermia indujo una reducción de la activación central en ejercicios repetidos de máxima contracción voluntaria (Nybo, L., Nielsen, B., 2001). El aumento de la temperatura corporal produciría una disfunción metabólica determinante de la fatiga aún en presencia de reservas adecuadas de glucógeno limitando el rendimiento (Parkin, J.M. et al., 1999).
Volemia
La reducción de la volemia es el principal factor de reducción del volumen sistólico en el estado deshidratado sea en el calor o frío (González-Alonso, J, Mora-Rodríguez, R., Coyle, E.F., 2000). Especialmente en el calor disminuye la eficiencia del sistema cardiocirculatorio en la termorregulación y se compromete la perfusión de órganos y tejidos por disminución de la presión sanguínea (González-Alonso, J. et al., 1997)
Disociación de Oxihemoglobina
Cuando la hemoglobina oxigenada alcanza los tejidos donde se necesita el oxígeno, este se disocia de la hemoglobina en razón de la diferencia en la presión parcial del mismo entre la sangre y los tejidos y debe difundir pasivamente hasta el interior de la célula y en ella hasta la mitocondria. En el músculo activo el aumento de la temperatura y la acidez de la sangre favorecen la disociación de hemoglobina, lo que se conoce como efecto Bohr.
La “carga de fosfatos” produce un aumento en la concentración plasmática y en los eritrocitos de fosfato y en estos últimos también de 2,3-difosfoglicerato (Bremner, K. et al., 2002); esto favorece la disociación de la hemoglobina pudiendo aumentar el VO2 max. y atenuar la respuesta láctica al ejercicio (Cade, R., et al., 1984).
Reservas de Glucógeno
Numerosos estudios sobre el efecto de las reservas de glucógeno, su manipulación con la dieta y el ejercicio y su efecto sobre el rendimiento se han realizado y revisado y de las conclusiones sobresalientes podemos citar que la estrategia de carga de carbohidratos beneficia a atletas que participen en pruebas de fondo >90 min (Hargreaves, M., Hawley, J.A., Jeukendrup, A.E., 2004; Jeukendrup, A.E., et al., 2005). Rausch, H.G.L, et al. (2005) comprobaron el efecto ergogénico de la carga de carbohidratos y observaron un efecto de la reserva de glucógeno sobre la intensidad de ejercicio elegida para completar la prueba y postulan un mecanismo de regulación central con un sensor muscular periférico a las cantidades de glucógeno.
Reservas de Lípidos
Un efecto de la adaptación a los deportes de fondo es la mayor capacidad de utilizar lípidos como fuente de energía ahorrando glucógeno (Van Looon, L.J.C, 1999). Se puede lograr una adaptación a una mayor utilización de lípidos con una dieta elevada en grasas y favorecer la utilización de estas en el ejercicio ahorrando glucógeno, pero no pudo observarse mejoras en el rendimiento después de 4h de ciclismo a 65% VO2 máx. y 1 hora de prueba de tiempo (Carey, A., 2001) ni en 100 km de ciclismo, mas aún se observó disminuye el rendimiento para realizar un esprint final (Havemann, L. et al., 2006).
Se postula que el consumo de dietas altas en lípidos y su adaptación metabólica muscular tardaría 2 días en producirse (Zderic, T.W., et al., 2004) y que podría beneficiar a las competencias de ultra resistencia a VO2 max. inferiores al 85% (Pendergast, D.R, Leddy, J.J., Venkatraman, J.T., 2000), pero la evidencia actual no comprueba un aumento en el rendimiento ( Jeukendrup, A.E., 2003; Hargreaves, M., Hawley, J.A., Jeukendrup, A.E., 2004; Burke, L.M., Kiens, B., Ivy, J.L., 2004).
Para el trabajo aeróbico es importante que las reservas de lípidos intramiocelulares sean repuestas, después de ejercicios prolongados pueden agotarse (Johnson, N.A., et al., 2003) y su recuperación puede tardar desde 24 hasta 72 horas dependiendo de la dieta (Décombaz, J., et al., 2001; Larson-Meyer, D.E., Bradley R.N., Hunter, G.R., 2002; Van Loon L.J.C., 2003).
Equilibrio Electrolítico
El principal trastorno en el ejercicio se produce con la hiponatremia, las alteraciones relacionadas a la concentración plasmática de K+ son raras y ocurren en pruebas muy prolongadas en el calor, cuando hay una deficiencia de K+ o cuando se han usado agentes diuréticos (Wittbrodt, E.T., 2003).
PRACTICAS Y NUTRIENTES DE UTILIDAD COMPROBADA
Bebidas Deportivas
La bebida deportiva durante la actividad debe proveer a la rápida absorción de agua y carbohidratos. Hipotónica o isotónica con 4 a 6% de carbohidratos y 0,5 a 1,0 g/l de sodio (Na+) especialmente en pruebas de fondo de más de 4 horas, las cantidades de K+ no deberían ser mayores a 200-250mg/l, las cantidades de magnesio (Mg2+) no deberían ser mayores a 75-125mg/l , otros nutrientes, minerales o vitaminas no serían necesarios ni beneficiosos (Striegel, H., Niess, A.M., 2006). Estas bebidas, aún cuando proveen cantidades de carbohidratos inferiores la cantidad total de carbohidratos metabolizados durante el ejercicio, pueden mejorar el rendimiento en actividades prolongadas (Maughan, R.J., Bethell, L.R., Leiper, J.B., 1996).
El objetivo es reponer el agua para poder evitar la hipertermia, evitar la reducción del volumen plasmático, mantener la concentración plasmática de Na+ y aportar los carbohidratos en cantidad que iguale la máxima tasa de oxidación de carbohidratos exógenos. La estrategia de hidratación debe contemplar las necesidades impuestas por la duración de la competencia, la temperatura, las oportunidades de ingerir líquidos, la optimización de la velocidad de vaciamiento gástrico que es el factor limitante en la absorción y evitar los malestares estomacales (Coyle, E.F., 2004; Shirrefs, S.M., 2003), .
Al evaluar la deshidratación con relación a la pérdida de peso corporal hay que considerar que los fluidos en vejiga y tracto gastrointestinal pueden aumentar sin que aporten a la hidratación del organismo. La temperatura de la bebida influencia tanto la velocidad de vaciamiento gástrico como la apetencia de consumo (Boulze, D., Montastruc, P., Cabanac, M., 1983). Bebidas muy frías pueden ocasionar diarrea, la temperatura óptima estaría entre 5º y 15 ºC aunque hay gran variabilidad entre individuos. El aumento de la intensidad del ejercicio disminuye la velocidad de absorción de fluidos (Maughan, R.J., Leiper, J.B., McGaw, B., 1990).
La velocidad de vaciamiento gástrico depende del volumen, de la osmolaridad y en el caso de proteínas e hidrolizados de proteínas, principalmente de la densidad energética (Maughan, R.J., Leiper, J.B., Vist, G.E., 2004)
No se sabe la razón, pero las mujeres son más propensas a padecer de hiponatremia, la ingestión de 1 l de fluido/hora durante un evento de 4 horas es suficiente para generar una sobrecarga de agua en mujeres y se recomienda una ingesta de 680 mg Na+/h para disminuir el riesgo de hiponatremia (Twerenbold, R., et al., 2003).
La oxidación de carbohidratos aportados por la bebida no fue diferente en el frío del calor, siendo aproximadamente de 0,7 g/min para intensidad de 80% VO2 max. en cicloergómetro y menor para una solución 2% CH respecto de 6 % y 12 % las que no resultaron diferentes (Galloway, S.D.R., et al., 2001). El mantenimiento de la hiperglucemia durante ejercicios prolongados, 120 min en cicloergómetro al 70 % del VO2 máx., aumento la oxidación de carbohidratos (1.33-2.78 g/min) sin producir un ahorro en la utilización de glucógeno muscular (MacLaren, D. P. M., 1999).
En una prueba de 1h en cicloergómetro al 83% del VO2 máx. con provisión de solución de CH 6 %, de los 84 g ingeridos solo 22 g aparecieron en sangre, redujo débilmente la gluconeogenesis, aumento la oxidación de CH y no afectó el metabolismo muscular ni el rendimiento (McConell, G.K., et al., 2000)
El vaciamiento gástrico y la absorción de fluídos es similar con agua, o tres soluciones al 6% CHO de 197, 295, 414 mosmol/kgH2O (17-18 meq Na+, 3.2 meq K+) durante 85 min a 63% VO2 máx. en cicloergómetro (Gisolfi, C.V., et al., 1998)
La hiponatremia inducida por el ejercicio puede ocurrir en actividades prolongadas de más de 4 horas de duración, en actividades de menor duración se han reportado dos casos; se debe principalmente al exceso en el consumo de fluidos y se recomiendan para prevenirlo beber según necesidad (ad libitum) o siguiendo las recomendaciones de la USATF para estimar la tasa de sudoración y evitar el sobreconsumo de fluídos. No hay evidencias de que el consumo de Na+ o bebidas hipotónicas con electrolitos prevengan la aparición de hiponatremia en atletas que beben en exceso (Hew-Butler, T., et al., 2005).
Carbogeles
Los geles de carbohidratos son una fuente densa de carbohidratos (65-75%) fáciles de consumir, transportar y digerir. Son prácticos cuando es dificultoso transportar u obtener bebidas deportivas y útiles en competencias que requieran la provisión de carbohidratos (>90min). Es necesario consumirlos con una ingesta adecuada de líquidos para evitar malestares gastrointestinales y reducción del vaciamiento gástrico que disminuirá la absorción de agua. Un gel de 50 g tendrá aproximadamente 35 g de carbohidratos, y deberá consumirse con 500 ml de agua para lograr una concentración de carbohidratos aprox. de 7%. Un carbogel de bajo índice glicémico (GI: 50) constituido por miel y otro de alto (GI: 100) de dextrosa consumidos de a 15 g con 250 ml de agua cada 16 km y 250 ml de agua adicionales cada 3,2 km mejoraron el rendimiento en una prueba por tiempo de 64 km de ciclismo, sin apreciarse diferencias entre los dos tipos de geles (Earnest, C.P., et al. 2004). En general tienen sodio para reponer las pérdidas por sudoración y algunas otras sustancias como cafeína, triacilglicéridos de ácidos grasos de cadena media (MCT), vitaminas y extractos vegetales.
Carga de Carbohidratos
La supercompesación de glucógeno puede ser mantenida durante 3 días sin actividad y dieta moderada de carbohidratos (Goforth, Jr., H.W, et al., 1997)
La acumulación de glucógeno es mayor y más prolongada en protocolos de carga que comienzan con una depleción que en los que consisten en una disminución de la intensidad de ejercicio; actividades que se realizan a una intensidad correspondiente al 65% del VO2 máx. por 20 min no afectan la supercompesación de glucógeno (Goforth, Jr., H.W., et al, 2003).
Interesante es la observación que en condiciones de experimentación similares a las de las competencias actuales en una prueba de tiempo de ciclismo de 100 km, la carga de carbohidratos influenció minimamente el rendimiento cuando los atletas recibieron fluidos con carbohidratos durante la actividad (Burke, L.M., et al., 2000).
Según aclara Tarnopolsky, M.A. (2003), en varios estudios se dedujo que las mujeres no supercompesaban las reservas de glucógeno cuando los carbohidratos en la dieta se aumentaban con relación a la ingesta habitual (ej. 55%→75%) (Tarnopolsky, M.A., et al., 2001), la supercompensación se produce con dietas de carbohidratos de 8 g.kg-1.d-1 en relación la masa magra y en general para las mujeres 10 g.kg-1.d-1 con relación al peso corporal, lo que obliga a consumir a las mujeres calorías en exceso.
Una carga de carbohidratos de 24 horas después de 150 segundos de ciclismo a una intensidad correspondiente al 130% VO2 máx. y un esprint máximo de 30 seg, con carbohidratos de alto IG (80% de los carbohidratos en forma fluida de maltodextrina y 20% en forma de pasta, pan y arroz) aumentó las reservas de glucógeno en todas los tipos de fibras musculares en un 84% (Fairchild, T.J., el al., 2002).
Una dieta rica en carbohidratos (82%) 24 horas antes de una actividad física no influenció la concentración de lactato plasmática ni la concentración máxima de lactato plasmática en estado estable ni la potencia desarrollada, solo el consumo de oxígeno fue mayor y estadísticamente significativo (Frank, I., Boutellier, U., Knöpfli-Lenzin, C., 2004). La adición de proteínas a los carbohidratos ingeridos después del ejercicio no aumentó ni disminuyó la velocidad de síntesis de glucógeno a pesar de haber inducido una mayor insulinemia (Jentjens, R.L.P.G., et al., 2001) ni aumentó ni disminuyó la concentración de glucógeno en músculo (Carrithers, J.A., et al., 2000). Un resultado distinto manifiesta Ivy, J.L. (2004) al mantener que la eficiencia de la acumulación de glucógeno puede ser aumentada combinando proteínas y carbohidratos en relación 8:2 y que además reduce la frecuencia de ingestiones necesarias. Al elegir las proteínas a consumir hay evidencias que con las proteínas de suero de leche se logran mayores niveles de glucógeno en hígado (Morifuji, M., Sakai, K. Sugiura, K., 2005) y en músculo (Morifuji, M., comunicación personal).
Fat Loading
El objeto de la carga de lípidos es aumentar las reservas de lípidos intramiocelulares y la oxidación de ácidos grasos durante el ejercicio. Las dietas ricas en lípidos no influyen sobre el VO2 max., y no son efectivas en individuos no entrenados. La carga de lípidos, para que sea efectiva en el mejoramiento de la capacidad de resistencia aeróbica, tiene que realizarse por varios días con una dieta rica en lípidos (65%) con entrenamiento intensivo simultaneo, una carga de carbohidratos previa a la competencia no modifica la adaptación al metabolismo de los lípidos. Los atletas que pueden sacar provecho de esta práctica son aquellos que deban competir por más de 6 horas o días a una intensidad del 50-60% VO2 max. (Knechtle, B., 2005).
Cafeína
La cafeína es una sustancia presente en las bebidas cola, bebidas energéticas, café, té, mate y en algunos carbogeles y otros suplementos. Desde el año 2004 dejo de estar dentro de las sustancias prohibidas (en realidad no estaba prohibida sino limitada) por la World Anti-Doping Agency (WADA, 2004).
Debido a la alta incidencia de trastornos GI en los deportes de resistencia se investigó si una bebida con carbohidratos, electrolitos y cafeína podría alterar la respuesta gastrointestinal al ejercicio con relación a otra sin cafeína o agua, no pudiendo observarse diferencias en 90 min de ciclismo a 70% de la potencia máxima (Wmax). (Van Nieuwenhoven, M. A., Brummer, R.-J. M., Brouns, F., 2000)
Jacobson et al. (2001) no pudieron comprobar que la coingestión de cafeína (7 mg/kg peso corporal) con carbohidratos o lípidos pudiera mejorar la utilización de sustratos en una prueba en cicloergómetro a 70% VO2 máx. por 120 min, ni el rendimiento en una prueba posterior de tiempo a 85 % VO2 max (Jacobson, T.L., et al, 2001). En otro estudio no se pudo observar un aumento de la disponibilidad de ácidos grasos, pero si una mejora en el rendimiento con cafeína 3,2mg/kg PC que no aumento con cafeína 4,2mg/kg PC ingeridos antes y durante prueba de tiempo en cicloergómetro, en este mismo estudio se observó que durante el ejercicio el efecto diurético de la caféina es minimo e improbable que pueda perjudicar la hidratación (Kovacs, E.M.R., Stegen, J.H.C.H., Brouns, F., 1998). La cafeína no produce un efecto ahorrador de glucógeno en atletas con alto contenido inicial de glucógeno y disminuiría el umbral de liberación de ß-endorfinas y cortisol (Didier, L., et al., 2000). Uno de los campos donde más atención se ha prestado a las sustancias ergogénicas es en el militar y según conclusiones presentadas en el Canadian Military Journal, donde aprueban y recomiendan el uso de cafeína, esta mejora el rendimiento cognoscitivo frente a privación de sueño y el tiempo hasta agotamiento en individuos ejercitando a 80%W máx., observándose efecto mayor y más duradero en individuos no habituados. (McLellan, T.M., et al., 2003; Douglas, B., McLellan, T.M., 2002), El mecanismo de acción no esta completamente dilucidado, sería mixto disminuyendo la percepción de fatiga a nivel central donde bloquearía receptores a la adenosina (Davis, M.J., et al., 2003), alterando el proceso sensorial desde el músculo (Plaskett, C. J.,Cafarelli, E., 2001) incrementando la excitabilidad y habilidad de activar voluntariamente unidades motoras (Kalmar, J. M., Cafarelli, E., 1999) por acción directa sobre el músculo esquelético (Tarnopolsky, M., Cupido, C., 2000; Jackman, M., 1996), .
PRACTICAS Y NUTRIENTES DE UTILIDAD DUDOSA O INUTILES
Bebidas Oxigenadas
Son publicitadas como favorecedoras del rendimiento físico y recuperación. La inspiración de cantidades adicionales de oxigeno durante el ejercicio aumenta el oxígeno arterial, disminuye la ventilación pulmonar (VP), frecuencia cardiaca (FC) submáxima y los niveles sanguíneos de lactato ([La-]s). Se investigó en con un diseño doble ciego cruzado (Willmert, N., et al., 2002) si el consumo de estas bebidas influenciaban la VP, FC y [La-]s en reposo, ejercicio submáximo y máximo no pudiendo comprobarse este efecto ni aumento en el tiempo hasta agotamiento ni en un segundo test máximo después del primero. Se cuestiona que 500 ml de bebida aportarían 24 ml de O2 que solo alcanzarían para cubrir 1% de la demanda de 1 minuto de actividad máxima y que si el O2 se absorbiera a nivel gastrointestinal, el oxígeno extra lo consumiría el epitelio gástrico y que la absorción se realizaría al sistema venoso que transporta la sangre para su reoxigenación en los pulmones donde se saturaría de O2 a los niveles normales (Willmert, N., et al., 2002).
Proteínas
Las necesidades de proteínas de los atletas están aumentadas en relación a la población inactiva. De los 20 aminoácidos que conforman las proteínas, 8 son esenciales y deben provenir de la dieta. Ahora bien, es necesario ingerir proteínas durante el ejercicio? Sorprendentemente estudios realizados durante el Tour de France mostraron un balance nitrogenado cuando la cantidad de calorías ingeridas fue adecuada y las proteínas aprox. 1,4g/kg peso corporal./día. A diferencia de lo que sucede con carbohidratos y agua no se ha demostrado que la deficiencia de proteínas en la ingesta aguda afecte el rendimiento en pruebas de fondo (Tarnopolsky, M.A., et al., 2005) . Aún después de 6h de ejercicio extenuante cuando durante la actividad se consumieron carbohidratos, la oxidación de proteínas no fue diferente que en reposo o durante la recuperación; la adición de proteínas mejoró el balance neto de proteínas en el reposo y en la recuperación pos ejercicio (Koopman, R., et al., 2004) Para el músculo en actividad la síntesis de proteínas no es prioritaria y se encuentra reducida (Atherton, P.J., Rennie, M., 2006; Rose, A.J., et al., 2005),
Aminoácidos Ramificados
Los aminoácidos ramificados (l-Leucina, l-Isoleucina y l-Valina) son esenciales, no pueden ser sintetizados por el organismo y deben ser aportados por la dieta. Forman parte del músculo y también se encuentran libres en el plasma. Por dos razones se ha propuesto que serían beneficiosos en la actividad física: cuando la disponibilidad de carbohidratos disminuye como sucede en la actividad física los aminoácidos ramificados son captados y metabolizados por el músculo (Jackman, L.M., et al., 1997), disminuye su concentración plasmática (Suryawan, A., et al., 1998); la disminución de la concentración plasmática de BCAA fue relacionada con la fatiga de origen central por su efecto competitivo con el l-Ttriptofano en el acceso a al cerebro o por su acción directa (Newsholme, E.A., Blomstrand, E., 2006).
En el estado hipohidratado en el ejercicio se compromete la termorregulación, el mecanismo hipotalámico de la sensación de fatiga en hipertermia está regulado en parte por serotonina, como los BCAA compiten con el precursor de la serotonina por atravesar la barrera hematoencefálica se estudió que influencia pudieran tener la suplementación con BCAA en estado hipohidratado no pudiendo comprobarse alteraciones en el rendimiento, ni en la percepción de fatiga ni en las funciones cognitivas (Cheuvront, S.N., et al., 2004) ni en condiciones de depleción de glucógeno en el calor (Watson, P., Shirreffs, S.M., Maughan, R.J., 2004) . Los BCAA son reguladores estimuladores de la síntesis proteica en músculo (Liu, Z., et al., 2002; Bolster, D.R., et al., 2004), pero no se pudo observar que afecte el metabolismo durante el ejercicio sino en la recuperación (Blomstrand, E., Saltin, B., 2001) y su utilidad en la ingestión previa al ejercicio se vio en la recuperación disminuyendo los dolores tardíos asociados al ejercicio (DOMS) y la fatiga (Yoshiharu, S., et al., 2006). El efecto directo de la suplementación de BCAA sobre el rendimiento en pruebas de fondo ha dado resultados inconsistentes (Yoshiharu, S., et al., 2004) o no pudo evidenciar un efecto ergogénico (Madsen, K. et al., 1996)
MCT (Triacilglicéridos de Cadena Media)
Los triacilgliceridos de ácidos grasos de cadena media a diferencia de los de cadena larga no retardan el vaciamiento gástrico, son rápidamente hidrolizados en el lumen intestinal y absorbidos en el torrente sanguíneo (Bach, A.C., Ingenbleek, Y., Frey, A., 1996), no son reesterificados y penetran en la mitocondria sin depender del transporte con carnitina (carnitina:acilcarnitina translocasa, CT); por esta razón se los investigó suponiendo que podrían ser metabolizados ahorrando glucógeno e independientemente de la inhibición de la oxidación de ácidos grasos que se observa con el aumento plasmático de hidratos de carbono (Horowitz, J.F, Klein, S., 2000). La suplementación con cantidades tolerables (25 g) de MCT no produjo efecto ahorrador del glucógeno ni aumento del rendimiento en pruebas de 30 min de alta intensidad (Horowitz, J.F., et al., 2000), ni submáximas en prueba de tiempo de 100 km de ciclismo (Angus, D.J., 2000).
Minerales
Magnesio
El ejercicio y en particular en los de fondo de larga duración aumentan la demanda y/o excreción de magnesio. Se ha comprobado que los deportistas de fondo padecen con mayor frecuencia que las personas inactivas deficiencias de magnesio a pesar de ingerir mayores cantidades (Saur, P., et al., 2002). La hipomagnesemia que se manifiesta en plasma durante el ejercicio prolongado debido principalmente a la redistribución del magnesio a compartimentos, desde eritrocitos al plasma y al tejido muscular activo y a los adipocitos, no es de importancia desde el punto de vista de la salud para personas con niveles normales de magnesio y es solo transitoria recuperándose dentro de las 12-72 h (Warburton, D.E.R., et al., 2002).
La suplementación con magnesio durante algunas semanas podría mejorar el metabolismo, el rendimiento, la actividad neuromuscular, fuerza o regeneración pero su efectividad es aún tema en discusión. No se ha podido comprobar que mejoras en estos parámetros no se hayan debido a una deficiencia latente de magnesio. Una sobre dosificación por administración oral cuando la función renal es normal es improbable. Aún a dosis terapéuticas se puede producir diarrea (Saur, P., 2004).
Según resumen Saur, P. et al. (2002) la ingesta diaria de magnesio recomendada es en: Alemania (250-300mg), Australia (270-320mg), Canada (250-300 mg), Francia (350mg), Italia (300-400mg), Nueva Zelandia (300-400mg), Polonia (350-400mg), USA (300-400mg), y en Argentina (400mg) (Código Alimentario Argentino).
Potasio
Difundida es la creencia que durante o después de la actividad física las pérdidas de potasio son cuantiosas y que su reposición mejora el rendimiento y recuperación, razón por la cual siempre alguien recomienda comer bananas porque contienen mucho potasio (1 banana mediana de 100 g: K+ 328mg; 1 naranja de 160 g: K+ 312mg (USDA)). En una prueba de ciclismo realizada a una intensidad correspondiente al 50 % de VO2 max. por 2 horas a 45ºC cada día por cuatro días y con una ingesta diaria de 25mEq K+ no se produjo una reducción significativa del K+ corporal (Costill, D.L., Cote, R., Fink, W.J., 1982). Las perturbaciones y fatiga se deben principalmente a una redistribución de las [K+] desde el espacio intracelular hacia el extracelular, en el restablecimiento del equilibrio que depende del estado nutricional y del entrenamiento del individuo (Sejersted, O.M., Sjøgaard, G., 2000). Las pérdidas de potasio y otros minerales por el ejercicio son sobrestimadas y no debería adicionarse a las bebidas durante el ejercicio o no superar los 200-250mg/l (Striegel, H., Niess, A.M., 2006) siendo las pérdidas por sudor en el ejercicio al 60% VO2 max. a 34ºC de 200mg K+/l (Shirreffs, S. M., Maughan, R.J., 1997).
Calcio
Aproximadamente 99% del calcio del organismo se encuentra en el tejido óseo. El calcio está involucrado en una gran variedad de procesos fisiológicos asociados con el metabolismo energético y la contracción muscular. La [Ca2+] plasmática varía muy poco por el equilibrio establecido con el tejido óseo. Por esta razón el calcio no es considerado un suplemento ergogénico potencial (Williams, M.H., 2005).
Hierro
Las deficiencias de hierro con o sin anemia disminuyen el rendimiento aeróbico (Zhu, I.H, Haas, J.D., 1997; Beard, J., Tobin, B., 2000). La suplementación con hierro no modificará la condición en forma aguda, generalmente ocasiona nauseas y dolores epigástricos 30-60 minutos después de la ingesta y si no es la adecuada puede ocasionar hemocromatosis, daños hepáticos severos, diabetes y afectar la función cardiaca (Beutler, E., Hoffbrand, A.V., Cook, J.D., 2003).
Carnitina
La L-carnitina es un aminoácido sintetizado en el organismo en hígado y riñones. Durante el ejercicio la limitación en la captación de ácidos grasos del plasma para su oxidación se verificaría a nivel intracelular en el ingreso de los ácidos grasos a la mitocondria para lo que deben esterificarse con L-carnitina (Roepstorff, C., et al., 2004) , y se postula que la disponibilidad de l-Carnitina libre para esterificarse con los ácidos grasos libres en la membrana externa de la mitocondria catalizado por CPT1 es limitante para la β-oxidación en el músculo con elevada cantidad de glucógeno durante el ejercicio (Roepstorff, C., et al., 2005). Por su participación en el metabolismo de los ácidos grasos y de los carbohidratos se busco favorecer la oxidación de ac. grasos ahorrando carbohidratos durante la actividad física por medio de la suplementación con L-carnitina; de la ingestión de L-carnitina se absorbe en el intestino delgado aprox. 5-15% , el pico de concentración en sangre se observa a las 3 horas y a pesar de administración prolongada (> 4 semanas) en el músculo el aumento de [L-carnitina]TOTAL es mínima. Su uso en intervención aguda no mejoró el rendimiento, y los resultados en utilización crónica son controversiales observándose en algunos reducción de la acumulación de lactato, aumento del VO2 max. y de la utilización de ácidos grasos (Brass, E.P., 2000), más bien su uso crónico sería beneficioso para personas mayores y no entrenados para evitar daños asociados al estrés inducido por el ejercicio (Volek, J.S., et al., 2002) y protección del músculo frente a la hipoxia (Kraemer, W.J., et al., 2003).
Vitaminas
Las graves deficiencias de vitaminas pueden ocasionar serios trastornos a la salud como el escorbuto, beri-beri, pelagra etc. Las deficiencias de vitaminas pueden disminuir el rendimiento físico. Se considera que la actividad física aumenta las necesidades de ciertas vitaminas como la B1, B2 y B6 y que el consumo por los atletas de dietas balanceadas que cubran las necesidades aumentadas de calorías también cubren las de vitaminas (Manore, M.M., 2000). El tema objeto de este artículo es si la suplementación con vitaminas puede influenciar el rendimiento físico y se puede decir que no hay evidencias se que puedan tener efecto ergogénico (Weight, L.M., Myburgh, K.H., Noakes, T.D., 1988) más bien el exceso puede disminuir el rendimiento aeróbico como la niacina o ser fatales coma el de vitamina A u ocasionar trastornos hepáticos y otros aunque se trate de vitaminas hidrosolubles (Williams, M.H., 2004) que según la creencia popular son eliminadas sin perjuicio al organismo. La suplementación que aparece como no riesgosa y con beneficios en la actividad física, no en el rendimiento sino en la protección al daño celular ocasionado por radicales libres son las vitaminas E y C (Williams, J.E., 2000; Zimmermann, M.B., 2003; Clarkson, P.M., Thompson, H.S., 2000) y en un estudio sobre un modelo animal se le atribuyó al consumo de altas dosis de vitamina E un efecto negativo en la función muscular deprimiendo la contractilidad del músculo a bajas frecuencias de estimulación (Coombes, J.F., et al., 2001). Para mayor y detallada información acerca de requerimientos nutricionales de vitaminas ver “Human Vitamin and Mineral Requirements, Report of a joint FAO/WHO expert consultation Bangkok, Thailand, 1998” ().
ATP
La suplementación nutricional con 250mg/día de ATP (trifosfato de adenosina) por 14 días no tuvo efecto sobre [ATP] plasmática ni sanguínea ni sobre ningún parámetro de la evaluación anaeróbica de Wingate y fuerza muscular (1RM y tres series de repeticiones hasta la fatiga al 70% de 1RM), (Jordan, A.N., et al., 2004). La administración oral crónica de ATP o adenosina modifica el recambio extracelular de purinas (Kichennin K., Seman, M., 2000) y parámetros fisiológicos bajo el control de receptores purínicos como la resistencia vascular periférica y la resistencia pulmonar (Kichennin, K, et al., 2000).
Ginseng
Bajo este nombre encontramos especies vegetales diferentes permitidas en los suplementos dietarios argentinos: Panax quinquefolium (nombre común: Ginseng americano), Panax ginseng (n.c.: Ginseng coreano, G. asiático, G. chino), Panax japonico (n.c.: Ginseng chikutsu japonés), Panax notoginseng (n.c.: Ginseng himalayo) y Eleuterococcus senticosus, (n.c.: Ginseng siberiano, Ciwuja). Estudios acerca del efecto ergogénico de P. Ginseng (Allen, J.D., et al., 1998) P. quinquefolium (Hsu, C.C., et al., 2005), E. senticosus (Goulet, E.D.B., Dionne, I.J., 2004) no reportaron ningún beneficio sobre el rendimiento aeróbico. Se comprobó en Panax ginseng un efecto protector de los daños oxidativos en las fibras musculares asociados al ejercicio agudo (Voces, J., et al., 2004). El uso agudo de Ginseng (P. ginseng, P. quinquefolium, E. senticosus) no mejora el rendimiento aeróbico y en usos de más de ocho semanas los resultados son controversiales observándose que podría beneficiar más a los individuos no entrenados y los de edad avanzada (>40 a) (Bucci, L., 2000). Medicinalmente P. ginseng está indicado como adaptógeno, preventivo y restaurador de las capacidades mentales y físicas, en casos de debilidad, agotamiento, cansancio, dificultad de concentración y convalecencia, efectos que no se observan de forma aguda; relativamente raro y con el uso crónico de altas dosis puede producir insomnio, nerviosidad, diarreas (WHO, 1999).
Glicerol
El glicerol es un metabolito natural del organismo. Lo encontramos ampliamente distribuido en los alimentos, principalmente como triacilglicerido y rara vez como tal. El glicerol puede ser sintetizado en el organismo en el hígado y el riñón y también transformarse en glucosa en la gluconeogénesis. Por esta razón se investigo su uso para prevenir la hipoglucemia y fatiga en pruebas de fondo sin que pudiera observarse un efecto positivo (Miller, J.M., et al., 1983). La ingestión de glicerol con agua (aprox. 1-1,5 g/kg corporal y 1,5-2 l agua) aumentan la concentración en sangre de glicerol en 15 minutos y se alcanza la máxima concentración en 75 minutos. Esto provoca un aumento de la osmolaridad y la capacidad de retener agua que también se manifiesta en los espacios intra- e intercelulares. El estado de hiperhidratación que se logra mejoraría la termoregulación en climas cálidos y húmedos y la hipervolemia favorecería el rendimiento cardíaco y así el rendimiento en ejercicios submáximos en climas cálidos y/o húmedos siendo en últimos estudios solo atribuido a la hipervolemia (Latzka, W.A., et al., 1997; Kavouras, S.A., et al., 2006). Los resultados y opiniones sobre estos beneficios son ambiguos. En el Australian Institut of Sport (AIS, 2006) y Mannhart, C. (2003) del Sportwissenschaftliches Institut (BASPO, Schweiz) encuentran que podría ser útil en competencias en clima cálido húmedo con difícil acceso a provisión de agua; Coyle, E.F (2004) no lo encuentra beneficioso y Jeukendrup A.E. et al. (2005) consideran que los triatletas que pudieran resultar seriamente deshidratados durante el ejercicio podrían beneficiarse pero que deben considerar los potenciales efectos adversos. Malestares gastrointestinales y en razón de que el glicerol penetra lentamente la barrera hemato-encefálica y el globo ocular, se pueden experimentar mareos, dolor de cabeza y visión borrosa.
En un estudio se compararon los efectos cardiovasculares, termoregulatorios y sobre el rendimiento aeróbico de la hiperhidratación inducida por agua y por una solución que proveía 1,2 g de glicerol por kg de peso corporal cada 26ml de solución en una prueba de ciclismo de 2h a una intensidad del 66% de VO2 max a una temperatura de 25 ºC seguido de un test incremental hasta agotamiento. Se observó que la hiperhidratación previa al ejercicio no fue significativamente diferente entre tratamientos, ni tampoco fueron diferentes la tasa de sudoración, la temperatura rectal, la frecuencia cardiaca ni la percepción del esfuerzo y que ninguno de los tratamientos indujo efectos indeseables y se concluyó que la hiperhidratación inducida con glicerol comparada con la inducida con agua disminuye la producción de orina, no mejora las funciones cardiovasculares y termoregulatorias y no mejora la percepción del esfuerzo en atletas entrenados consumiendo 500ml/h de líquido durante el ejercicio en las condiciones del experimento (Goulet, E.D. et al., 2006)
Bebidas Energizantes
Las bebidas energéticas fueron incorporadas al CAA como suplementos dietarios por disposición ANMAT 6611/00 y redefinidas disminuyendo la cantidad máxima de cafeína permitida por disposición ANMAT 3634/05 actualmente en vigencia efectiva.
¿Qué contienen?. Son bebidas no alcohólicas que pueden contener ingredientes como taurina (max. 400mg/100ml), glucuronolactona (max. 250mg/100ml), cafeína (20mg/100ml), inositol (20mg/100ml), acompañados de hidratos de carbono, de vitaminas o minerales y otros ingredientes autorizados. Se consideran “Altos en energía” si el contenido energético aportado principalmente por hidratos de carbono es igual o superior a 45kcal/100ml. La cantidad de cafeína permitida es la misma que para otras bebidas con cafeína como las colas.
Es importante remarcar que en caso de consumir bebidas energéticas fabricadas en otros países estas pueden contener mayores cantidades de cafeína.
El Comité Científico sobre Alimentos (SCF) de la Comisión Europea emitió opinión sobre la seguridad y efectos sobre el ejercicio de las bebidas energéticas con cafeína, taurina y glucuronolactona (SCF, 2003). Las conclusiones principales fueron que sobre la base de la evidencia científica no se puede establecer un nivel máximo seguro de consumo para taurina y glucuronolactona ni prohibir su consumo, y con relación al ejercicio concluye que es poco probable que la glucuronolactona pueda tener interacción con cafeína, taurina, alcohol o el efecto del ejercicio, y que un efecto aditivo diurético se observa entre taurina y cafeína que podría resultar, asociados a la perdida de fluidos por sudoración, en una deshidratación temprana (SCF, 2003).
Otros países de la CEE como Dinamarca y Francia no autorizan su comercialización. La Agencia Francesa de Seguridad Sanitaria de los Alimentos considera que la inocuidad no está probada, de la glucuronolactona por su toxicidad renal y de la taurina por los efectos indeseables sobre el comportamiento nervioso y posibles alteraciones de la glándula tiroides (AFSSA, 2002) y en una reevaluación concluyen que no está probada la inocuidad de las bebidas energéticas con cafeína, taurina y glucuronolactona (AFSSA, 2006).
Para evaluar la efectividad sobre el rendimiento físico (fuerza, velocidad y potencia) el tiempo de reacción, la memoria reciente y estado de ánimo se realizó un estudio doble ciego con control placebo en atletas no encontrándose diferencias significativas entre los grupos estudiados concluyendo los autores que al momento es inapropiado recomendar como ayuda ergogénica el consumo de bebidas energéticas y que son necesarios más estudios acerca de su efectividad y toxicidad (Carvajal-Sancho, A., Moncada-Jimenez, J., 2005).
APLICACIONES
Una de las características de los seres biológicos es la variabilidad y el hombre es parte de ella. Cada atleta debe encontrar la práctica nutricional qué mejor le siente y como regla de oro no debe buscarla durante una competencia.
Se pueden mencionar algunas generalidades y considerar la intensidad, la duración y la temperatura en el día de la prueba.
Días Previos
Reducción en la intensidad y duración del entrenamiento y la dieta basada en un aporte calórico de carbohidratos de aproximadamente 65% del total de calorías ingeridas (Tarnopolsky, M.A., et al., 2005)
Carga de Carbohidratos
La carga de carbohidratos puede mejorar el rendimiento en pruebas de más de 90 min de duración por lo que en la modalidad Sprint no sería adecuado utilizarla (Jeukendrup, A.E., Jentjens, R.L.P.G., Moseley L., 2005); el aumento de peso asociado al aumento de las reservas de glucógeno se produce porque cada 1 g de glucógeno combina aproximadamente 3 g de agua y en una masa total de 800 g de glucógeno corporal se fijan 2,2 kg de agua y puede ser perjudicial para pruebas con limitaciones de peso o en las que el aumento de peso pueda reducir el rendimiento (Hollman, W., Hettinger, Th., 2000). Hay descriptas distintas prácticas y métodos para lograr la supercompensación y carga de carbohidratos. El principio general consiste en cambiar la dieta mixta equilibrada por una dieta con una alta proporción de carbohidratos.
El método clásico descrito por Karlsson, J. y Saltin, B. (1971) consiste en tres fases: primero una depleción de glucógeno con ejercicio aeróbico de fondo y reducción de la ingestión de calorías en los 7 a 4 días previos a la competencia, segundo dos a tres días en los que la dieta debe ser pobre en carbohidratos y rica en proteínas y grasas manteniendo un alto volumen de entrenamiento y en la última fase aumentar la proporción de carbohidratos a un 70% de las calorías de la dieta y reducción de la intensidad y volumen de entrenamiento lográndose una concentración de 34 g de glucógeno por kg de músculo contra 17 g de glucógeno por kg de músculo que se logran con una dieta normal y equilibrada.
El método modificado consiste en una reducción de la intensidad de entrenamiento en la semana previa a la competencia y una dieta con 60-70% de las calorías aportadas por carbohidratos (Sherman, W. et al., 1981).
El método rápido descrito por Fairchild, T.J. et al. (2002) consiste en realizar en ejercicio anaeróbico supramáximo para una rápida reducción del glucógeno muscular seguido de la ingestión de aproximadamente 10 g de carbohidratos por kg de peso corporal en 24 horas, lográndose aumentar el glucógeno desde 109 mmol a 198 mmol por kg de músculo.
No se puede decir que una es mejor que otra. Cada atleta debe encontrar la más conveniente.
Previo a la Competencia
La última ingesta debe hacerse no más allá de 2 horas antes de la competencia y se deben evitar fibras y grasas que retrasaran el vaciamiento gástrico, si en los momentos previos a la competencia (1/2 hora) se sufre de hambre se puede ingerir una pequeña cantidad (50g) de carbohidratos de fácil digestibilidad y de bajo índice glicémico (Jeukendrup, A.E., Jentjens, R.L.P.G., Moseley L., 2005).
A la competencia debe llegarse en estado euhidratado. Minutos antes de la competencia beber 300 500 ml de líquido.
Durante la Competencia
Las posibilidades de ingerir o beber alimentos se dan durante las transiciones, durante el ciclismo sin la provisión externa de agua o alimentos y durante el pedestrismo con alimentos y bebidas acarreados o provistos por los organizadores del evento.
Importante es informarse acerca de la distribución de los puestos de provisión y qué bebidas y alimentos serán puestos a disposición de los atletas para planificar la estrategia nutricional.
En la modalidad Sprint puede utilizarse solo agua en las otras modalidades una bebida con 4 a 6% de carbohidratos es conveniente para adecuar una ingesta de carbohidratos de 50-60g/h en el calor y de 60-70g/h en el frío porque se ha demostrado que la tasa de oxidación de carbohidratos ingeridos es menor en ambientes cálidos (aprox. 10%) que en el frío (Jentjens, R.L., Wagenmakers, A.J., Jeukendrup, A.E., 2002). La temperatura conveniente de las bebidas es de aprox. 14ºC, llevar las caramañolas con el líquido muy fío o con hielo y mantenerlas en el parque cerrado fuera del sol. En lo posible ingerir de 200-300 ml de líquido cada 20 min comenzando inmediatamente después de la natación con una cantidad de líquido adecuada a la duración de esta etapa.
Si durante el pedestrismo se ofrecen bebidas deportivas (4-6% carbohidratos) se puede prescindir del uso de carbogeles u otra fuente de carbohidratos; si solo se ofrece agua o en algunos puestos agua y otros bebidas deportivas, habrá que consumir aproximadamente 15 g de carbohidratos (1/2 carbogel de 40 g) con 250 ml de agua en pequeñas porciones para lograr la concentración óptima de carbohidratos en el estómago. Para pruebas de más de 4 horas de duración debe considerarse la ingestión de bebidas con sodio que aporten a una ingesta 680 mg de Na+/h para prevenir una hiponatremia.
Agradecimientos
Agradezco a la Federación Argentina de Triatlón y a su entrenador Prof. Enrique Bollana el apoyo y oportunidades otorgados.
REFERENCIAS
1. AFSSA (2003). D-glucuronolactone, de diverses vitamines et de caféine (à une dose supérieure à celle actuellement admise dans les boissons) dans une boisson dite «énergétique». Afssa Saisine n° 2002-SA-0260
2. AFSS (2006). D-glucuronolactone, inositol, vitamines B2, B3, B5, B6 et B12. Afssa – Saisine n° 2006-SA-0236
3. Allen J.D., McLung, J., Nelson, A.G, Welsch, M (1998). Ginseng supplementation does not enhance healthy young adults peak aerobic exercise performance. J Am Coll Nutr; 17(5): 462466
4. Angus, D.J., Hargreaves, M., Dancey, J., Febbraio, M.A (2000). Effect of carbohydrate or carbohydrate plus medium-chain triglyceride ingestion on cycling time trial performance. J Appl Physiol; 88: 113-119
5. (2000). Antioxidants: what role do they play in physical activity and health?. J Clin Nutr; 72(suppl): 637S-46S
6. Atherton, P.J., Rennie, M (2006). Protein synthesis a low priority for exercising muscle. J. Physiol. published online Apr 6; DOI: 10.1113/jphysiol.2006.110247. Acceso 27.04.06 en http://jp.physoc.org
7. Australian Institut of Sport (2006). AIS sport supplement fact sheet: glycerol. Acceso 09.06.2006 en http://www.ais.org.au/nutrition/documents/SupGlycerol06.pdf
8. Bach, A.C., Ingenbleek, Y., Frey, A (2006). The usefulness od dietary medium-chain triglycerides in body weight control: fact or fancy?. J. Lipid Research; 37: 708-726 (Review)
9. Beard, J., Tobin, B (2000). Iron status and exercise. Am J Clin Nutr; 72(suppl): 594S5977S
10. Bell, D.G., McLellan, T.M (2002). Exercise endurance 1, 3, and 6 h after caffeine ingestion in caffeine users and nonusers. J Appl Physiol; 93: 1227-1234
11. Beutler, E., Hoffbrand, A.V., Cook, J.D (2003). Iron deficiency and overload. Hematology; 40-61
12. Blomstrand, E (2006). A Role for branched-chain amino acids in reducing central fatigue. J Nutr; 136: 544S547S
13. Blomstrand, E., Saltin, B (2001). BCAA intake affects protein metabolism in muscle after but not during exercise in humans. Am J Physiol Endocrinol Metab; 281: E365-E374
14. Bolster, D.R., Vary, T.C., Kimball, S.R., Jefferson, L.S (2004). Leucine Regulates Translation initiation in rat skeletal muscle via enhanced eIF4G phosphorylation. J Nutr; 134: 1704-1710
15. Boulze, D., Montastruc, P., Cabanac, M (1983). Water intake, pleasure and water temperature in humans. Physiol Behav; 30(1): 97-102. (Abstract)
16. Brass, E.P (2000). Supplemental carnitine and exercise. Am J Clin Nutr;72(suppl): 618S-23S
17. Bremner, K., Bubb, W.A., Kemp, G.E., Trenell, M.I., Thompson, C.H (2002). The effect of phosphate loading on erythrocyte 2,3-bisphosphoglycerate levels. Clin Chim Acta; 323(1-2): 111-4. (Abstract)
18. Bucci, L (2000). Selected herbals and human exercise performance. Am J Clin Nutr; 72(suppl): 624S36S
19. Burke, L.M., Hawley, J.A., Schabort, E.J.,St Clair Gibson, A., Mujika, I., Noakes, T.D (2000). Carbohydrate loading failed to improve 100-km cycling performance in a placebo-controlled trial. J Appl Physiol; 88: 1284-1290
20. Burke, L.M., Kiens, B., Ivy, J.L (2004). Carbohydrates and fat for training and recovery. J Sports Sci; 22: 1530
21. Cade R., Conte M., Zauner C., Mars D., Peterson J., Lunne D., Hommen N., Packer D (1984). Effects of phosphate loading on 2,3-diphosphoglycerate and maximal oxygen uptake. Med Sci Sports Exerc; 16(3): 263-8. (Abstract)
22. Carey, A.L., Staudacher, H.M., Cummings, N.K., Stepto, N.K, Nikolopoulos, V., Burke, L.M., Hawley, J.A (2001). Effects of fat adaptation and carbohydrate restoration on prolonged endurance exercise. J Appl Physiol ; 91: 115122
23. Carrithers, J.A., Williamson, D.L, Gallagher, P.M., Godard, M.P., Schulze, K.E., Trappe, S.W (2000). Effects of postexercise carbohydrateprotein feedings on muscle glycogen restoration. J Appl Physiol; 88: 1976-1982
24. Casa, J.D (2006). USATF Self-testing program for optimal hydration. Acceso 25.06.2006 en www.usatf.org/groups/Coaches/library/hydration/ USATFSelfTestingProgramForOptimalHydration.pdf
25. Carvajal-Sancho A., Moncada-Jimenez J (2005). The acute effect of an energy drink on the physical and cognitive performance of male athletes. Kinesiologia Slovenica; 11: 5-16
26. Cheuvront, S.N., Carter III, R., Kolka, M.A., Lieberman, H.R., Kellogg, M.D., Sawka, M.N (2004). Branched-chain amino acid supplementation and human performance when hypohydrated in the heat. J Appl Physiol; 97: 1275-1282
27. Coggan, A.R., Coyle, E.F (1987). Reversal of fatigue during prolonged exercise by carbohydrate infudion or ingestion. J Appl Physiol; 63(6): 2388-2395
28. Coombes, J.S., Powers. S.K., Rowell, B., Hamilton, K.L., Dodd, S.L., Shanely, A.R., Sen, C.K., Packer, L (2001). Effects of vitamin E and a-lipoic acid on skeletal muscle contractile properties. J Appl Physiol; 90: 1424-1430
29. Costill, D.L., Cote, R., Fink, W.J (1982). Dietary potassium and heavy exercise: effects on muscle water and electrolytes. Am J Clin Nutr; 36: 266-275
30. Costill, D.L., Cote, R., Fink, W.J (1982). Dietary potassium and heavy exercise: effects on muscle water and electrolytes. Am J Clin Nutr; 36: 266-275
31. Coyle, E.F (2004). Fluid and fuel intake during exercise. J Sport Sci; 22: 39-55
32. Coyle, E.F (2004). Fluid and fuel intake during exercise. J Sport Sci; 22: 39-55
33. Davis J.M., Bailey S.P., Woods J.A., Galiano F.J., Hamilton M., Bartoli W.P (1992). Effects of carbohydrate feedings on plasma free-tryptophan and branched-chain amino acids during prolonged cycling. Eur J Appl Physiol;65: 5139
34. Davis, J.M., Alderson, N.L., Welsh, R.S (2000). Serotonin and central nervous system fatigue: nutritional considerations. Am J Clin Nutr;72(suppl): 573S578S
35. Davis, J.M., Zhao Z., Stock, H.S., Mehl, K.A., Buggy, J., Hand, G.A (1940). Central nervous system effects of caffeine and adenosine on fatigue. Am J Physiol Regul Integr Comp Physiol; 284
36. Davis, J.M., Zhao, Z., Stock, H.S., Mehl, K.A., Buggy, J., Hand, G.A (2003). Central nervous system effects of caffeine and adenosine on fatigue. Am J Physiol Regul Integr Comp Physiol; 284: R399R404
37. Didier L., Schneider, K.E, Prusaczyk, W.K.,Franklin, C., Vogel, A.M., Krssak, M., Falk Petersen, K., Goforth, H.W., Shulman, G.I (2005). Effects of Caffeine on Muscle Glycogen Utilization and the Neuroendocrine Axis during Exercise. J Clin Endocrinol Metab; 85(6): 2170-2175
38. Downey, B.M., Hopkins, W.G (2001). Nutritional Intake Predicts Performance in an Ironman Triathlon. Sportscience; 5(1), sportsci.org/jour/0101/bmd.htm. Acceso 09.06.06 en http://www.sportsci.org/jour/0101/bmd.pdf
39. Earnest, C.P., Lancaster, S., Rasmussen, C., Kerksick, C., Lucia, A., Greenwood, M., Almada, A., Cowan, P., Kreider, R. Low vs (2004). high glycemic index carbohydrate gel ingestion during simulated 64-km cycling time trial performance. J Strength Cond Res; 18(3): 466-472
40. (2004). Effects of EnduroxTM (Ciwujia) supplementation on endurance performance and the metabolic responses to endurance exercise: a brief review. J Exer Physiol online; 7(2): 30-36
41. Fairchild, T.J., Fletcher, S., Steele P., Goodman, P.C., Dawson, B., Fournier, P.A (2002). Rapid carbohydrate loading after a short bout of near maximal-intensity exercise. Med Sci Sports Exerc; 34(6): 980-986
42. Galloway, S.D.R., Wootton, S.A., Murphy, J.L., Maughan, R.J (2001). Exogenous carbohydrate oxidation from drinks ingested during prolonged exercise in a cold environment in humans. J Appl Physiol; 91: 654-660
43. Gandevia, S.C (2001). Spinal and Supraspinal Factors in Human Muscle Fatigue. Physiol Rev; 81(4): 1725-1789
44. Gisolfi, C.V (2000). Is the GI System Built For Exercise?. News Physiol Sc; 15: 114-119
45. Gisolfi, C.V., Summers, R.W., Lambert, G.P., Xia, T (1998). Effect of beverage osmolality on intestinal fluid absorption during exercise. J Appl Physiol; 85(5): 1941-1948
46. Glace, B.W., Murphy, C.A., McHugh, P.H (2002). Food intake and electrolyte status of ultramarathoners competing in extreme heat. J Am Coll Nutr; 21(6): 553-559
47. Goforth, H.W., Jr., Laurent D., Prusaczyk, W.K., Schneider, K.E., Falk Petersen, K, Shulman, G.I (2003). Effects of depletion exercise and light training on muscle glycogen supercompensation in men. Am J Physiol Endocrinol Metab; 285: E1304-E1311
48. Goforth, Jr., H.W., Arnall, D.A., Bennett, B.L, Law, P.G (1997). Persistence of supercompensated muscle glycogen in trained subjects after carbohydrate loading. J Appl Physiol; 82(1): 342-347
49. Goulet, E.D., Robergs, R.A., Labrecque, S., Royer, D., Dionne, I.J (2006). Effect of glycerol-induced hyperhydration on thermoregulatory and cardiovascular functions and endurance performance during prolonged cycling in a 25 degrees C environment. Appl Physiol Nutr Metab 31(2): 101-109
50. Hargreaves, M., Hawley, J.A., Jeukendrup, A.E (2004). Pre-exercise carbohydrate and fat ingestion: effects on metabolism and performance. J Sports Sci; 22: 3138
51. Hargreaves, M., Hawley, J.A., Jeukendrup, A.E (2004). Pre-exercise carbohydrate and fat ingestion: effects on metabolism and performance. J Sports Sci; 22: 31-38
52. Hew-Butler, T., Almond, C., Ayus, J.C., Dugas, J., Meeuwisse, W., Noakes, T., Reid, S., Siegel, A., Speedy, D., Stuempfle, K., Verbalis, J., Weschler, L (2005). Consensus Statement of the 1st International Exercise-Associated Hyponatremia Consensus Development Conference, Cape Town, South Africa 2005. Clin J Sport Med; 15: 208-213
53. Hollman, W., Hettinger, Th (2000). Sportmedizin: Grundlagen für Arbeit, Training und Präventivmedizin. Stuttgart: Schattauer
54. Horowitz, J.F., Klein, S (2000). Lipid metabolism during endurance exercise. Am J Clin Nutr; 72(suppl):558S-63S
55. Horowitz, J.F., Mora-Rodriguez, R., Byerley, L.O., Coyle, E.F (2000). Preexercise mediumchain triglyceride ingestion does not alter muscle glycogen use during exercise. J Appl Physiol; 88: 219-225
56. Hsu, CC., Ho, MC., Lin LC., Su B., Hsu MC (2005). American ginseng supplementation attenuates creatine kinase level induced by submaximal exercise in human beings. World J Gastroenterol; 11(34): 5327-5331
57. Ivy, J.L (2004). Regulation of muscle glycogen repletion, muscle protein synthesis and repair following exercise. J Sport Sci Med; 3: 131-138
58. Ivy, J.L., Costill, D.L., Fink, W.J., Lower, R.W (1979). Influence of caffeine and carbohydrate feedings on endurance performance. Med Sci Sports Exerc; 11: 611
59. Jackman, M., Wendling, P., Friars, D., Graham, T.E (1996). Metabolic, catecholamine, and endurance responses to caffeine during intense exercise. J Appl Physiol; 81(4): 1658-1663
60. Jackman, M.L., Gibala, M.J., Hultman, E., Graham, T.E (1997). Nutritional status affects branched-chain oxoacid dehydrogenase activity during exercise in humans. Am J Physiol Endocrinol Metab; 272: E233-E238. (Abstract)
61. Jacobson, T.L., Febbraio, M.A., Arkinstall, M.J., Hawley, J.A (2001). Effect of caffeine coingested with carbohydrate or fat on metabolism and performance in endurance trained men. Exp Phys 86(1): 137-144
62. Jentjens, R.L. P. G., Wagenmakers, A.J.M., Jeukendrup, A.E (2002). Heat stress increases muscle glycogen use but reduces the oxidation of ingested carbohydrates during exercise. J Appl Physiol; 92: 15621572
63. Jentjens, R.L.P.G., van Loon, L.J.C., Mann, C.H., Wagenmakers, A.J.M., Jeukendrup, A.E (2001). Addition of protein and amino acids to carbohydrates does not enhance postexercise muscle glycogen synthesis. J Appl Physiol; 91: 839-846
64. Jeukendrup, A.E (2003). High-carbohydrate versus high-fat diets in endurance sports. Schw Z «Sportmedizin und Sporttraumatologie» ; 51 (1): 1723
65. Jeukendrup, A., Brouns, F., Wagenmakers, A.J.M., Saris, W.H.M (1997). Carbohydrate-electrolyte feedings improve 1 h time trial cycling performance. Int J Sports Med; 18: 125129
66. Jeukendrup, A.E., Jentjens, R.L.P.G., Moseley L (2005). Nutritional considerations in triathlon. Sports Med; 35(2): 163-181
67. Johnson, N. A., Stannard, S.R., Mehalski, K., Trenell, M.I., Sachinwalla, T., Thompson, C.H., Thompson, M.W (2003). Intramyocellular triacylglycerol in prolonged cycling with high- and low-carbohydrate availability. J Appl Physiol; 94: 13651372
68. Jordan, A.N., Jurca R., Abraham E.H., Salikhova A., Mann J.K., Morss G.M., Church T.S., Lucia A., Earnest C.P (2004). Effects of oral ATP supplementation on anaerobic power and muscular strength. Med Sci Sports Exerc; 36(6): 983-90, (Abstract)
69. Kalmar, J. M., Cafarelli,E (1999). Effects of caffeine on neuromuscular function. J Appl Physiol; 87(2): 801-808
70. Karlsson, J., Saltin, B (1971). Diet, muscle glycogen, and endurance. J Appl Physiol: 31: 203-206
71. Kavouras, S.A., Armstrong. L.E., Maresh, C.M., Casa, D.J., Herrera-Soto, J.A., Scheett, T.P., Stoppani, J, Mack, G.W., Kraemer, W.J (2006). Rehydration with glycerol: endocrine, cardiovascular, and thermoregulatory responses during exercise in the heat. J Appl Physiol; 100: 442-450. (Abstract)
72. Kichenin, K., Decollogne, Jean Angignard, J., Seman, M (2000). Cardiovascular and pulmonary response to oral administration of ATP in rabbits. J Appl Physiol; 88: 9621968
73. Kichennin K., Seman, M (2000). Chronic oral administration of ATP modulates nucleoside transport and purine metabolism in rats. J Pharmacol Exp Ther; 294(1): 126133
74. Knechtle, B (2005). Kann Fatloading die Ausdauerleistungsfähigkeit verbessern?. Schweizerische Zeitschrift für «Sportmedizin und Sporttraumatologie»; 53 (4): 179-184
75. Koopman, R., Pannemans, D.L.E., Jeukendrup, A.E., Gijsen, A.P., Senden, J.M.G., Halliday, D., Saris, W,H.M., van Loon L.J.C., Wagenmakers, A.J.M (2004). Combined ingestion of protein and carbohydrate improves protein balance during ultra-endurance exercise. Am J Physiol Endocrinol Metab; 287: E712-E720
76. Kovacs, E.M.R., Stegen, J.H.C.H.,Brouns, F (1998). Effect of caffeinated drinks on substrate metabolism, caffeine excretion, and performance. J Appl Physiol; 85(2): 709-715
77. L. Havemann, L., West, S.J., Goedecke, J.H., Macdonald, I.A., St Clair Gibson, A., Noakes, T.D., Lambert, E.V (2006). Fat adaptation followed by carbohydrate loading compromises high-intensity sprint performance. J Appl Physiol; 100: 194-202
78. Larson-Meyer, D.E., Bradley R.N., Hunter, G.R (2002). Influence of endurance running and recovery diet on intramyocellular lipid content in women: a 1HNMR study. Am J Physiol Endocrinol Metab; 282: E95E106
79. Latzka, W. A., Sawka, M.N, Montain, S.J, Skrinar, G.S, Fielding, R.A, Matott, R.P., Pandolf, K.B (1997). Hyperhydration: thermoregulatory effects during compensable exercise-heat stress. J Appl Physiol; 83(3): 860-866
80. Liu, Z., Jahn, L.A., Wei, L., Long, W., Barrett, E.J (2002). Amino acids stimulate translation initiation and protein synthesis through an Akt-independent pathway in human skeletal muscle. J Clin Endocrinol Metab; 87: 5553-5558
81. MacLaren, D. P. M., Reilly, T., Campbell, I.T., Hopkin, C (1999). Hormonal and metabolic responses to maintained hyperglycemia during prolonged exercise. J Appl Physiol; 87(1): 124-31
82. Madsen, K., MacLean, D.A., Kiens, B., Christensen, D (1996). Effects of glucose, glucose plus branchedchain amino acids, or placebo on bike performance over 100 km. J Appl Physiol; 81(6): 2644-2650
83. Mannhart, C (2003). Aktuelle Leistungsförderer im Sport. Schw Zeit Sportmed Traum; 51(1): 58-79
84. Maughan, R.J (2003). Nutritional status, metabolic response to exercise and implications for performance. Biochemical Society Transactions; 31(6): 1267-1269
85. Maughan, R.J., Bethell, L.R., Leiper, J.B (1996). Effects of ingested fluids on exercise capacity and on cardiovascular and metabolic response to prolonged exercise in man. Exp Physiol; 81: 847-859
86. Maughan, R.J., Leiper, J.B., McGaw, B (1990). Effects of exercise intensity on absorption of ingested fluids in man. Exp Physiol; 75: 419-421
87. Maughan, R.J., Leiper, J.B., Vist, G.E (2004). Gastric emptying and fluid availability after ingestion of glucose and soy protein hydrolisate solutions in man. Exp Physiol; 89(1): 101-108
88. McConell, G.K., Canny, B.J, Daddo, M.C., Nance, M.J., Snow, R.J (2000). Effect of carbohydrate ingestion on glucose kinetics and muscle metabolism during intense endurance exercise. J Appl Physiol; 89: 1690-1698
89. McLellan, T.M., Bell, D.G., Lieberman, H.R., Kamimori, G.H (2003). The impact of caffeine on cognigtive and physical performance and marksmanship during sustained operations. Canadian Military Journal; 4(4)Winter 2003-2004: 47-54
90. Melinada, M.M (2000). Effect of physical activity on thiamine, riboflavin, and vitamin B-6 requirements. Am J Clin Nutr; 72(suppl): 598S606S
91. Miller, J.M., Coyle, E.F., Sherman, W.W., Hagberg, J.M., Costill, D.L., Fink W.J., Terblanche, S.E., Holloszy, J.O (1983). Effect of glycerol feeding on endurance and metabolism during prolonged exercise in man. Med Sci Sports Exerc; 15(3): 237-242
92. Morifuji, M., Sakai, K. Sugiura, K (2005). Dietary whey protein modulates liver glycogen level and glycoregulatory enzyme activities in exercise-trained rats. Exp Biol Med; 230: 23-30
93. Newsholme, E.A., Blomstrand E (2006). Branched-chain amino acids and central fatigue. J Nutr; 136: 274S276S
94. Noakes, T.D (2000). Physiological models to understand exercise fatigue and the adaptations that predict or enhance athletic performance. Scand J Med Sci Sports; 10: 123145
95. Noakes, T.D., Sharwood, K., Speedy, D., Hew, T., Reid, S., Dugas, J., Almond, C., Wharam, P., Weschler, L (2005). Three independent biological mechanisms cause exercise-associated hyponatremia: Evidence from 2,135 weighed competitive athletic performances. PNAS; 102(51): 18550-18555
96. Nybo, L., Nielsen, B (2001). Hyperthermia and central fatigue during prolonged exercise in humans. J Appl Physiol; 91: 10551060
97. Parkin, J. M., M. F. Carey, S. Zhao, and M. A. Febbraio (1999). Effect of ambient temperature on human skeletal muscle metabolism during fatiguing submaximal exercise. J Appl Physiol; 86(3): 902908
98. Pendergast, D.R, Leddy, J.J., Venkatraman, J.T (2000). A perspective on fat intake in athletes. J Am Coll Nutr; 19(3): 345350
99. Plaskett, C. J., Cafarelli, E (2001). Caffeine increases endurance and attenuates force sensation during submaximal isometric contractions. J Appl Physiol 91: 1535-1544
100. Rauch, H.G.L., St Clair Gibson, A., Lambert, E.V., Noakes, T.D (2005). A signalling role for muscle glycogen in the regulation of pace during prolonged exercise. Br J Sports Med; 39: 34-38
101. Roepstorff, C. , Vistisen, B., Roepstorff, K., Kiens, B (2004). Regulation of plasma long-chain fatty acid oxidation in relation to uptake in human skeletal muscle during exercise. Am J Physiol Endocrinol Metab; 287: E696-E705
102. Roepstorff, C., Halberg, N., Hillig, T., Saha, A.K., Ruderman, N.B., Wojtaszewski, J.F.P., Richter, E.A., Kiens, B (2005). Malonyl-CoA and carnitine in regulation of fat oxidation in human skeletal muscle during exercise. Am J Physiol Endocrinol Metab; 288: E133-E142
103. Rose, A.J., Broholm, C., Kiillerich, K., Finn, S.G., Proud, C.G., Rider, M.H., Richter, E.A, Bente, K (2005). Exercise rapidly increases eukaryotic elongation factor2 phosphorylation in skeletal muscle of men. J Physiol; 569(1): 223-228
104. Saur, P (2004). Magnesium und Sport. Dtsch Z Sportmed; 55(1): 23-24
105. SCF (2003). Opinion of the Scientific Committee on Food on Additional information on energy drinks (expressed on 5 March 2003). European Comimission, Health & Consumer protection Directorate-General, Document SCF/CS/PLEN/ENDRINKS/16 Final
106. Schirrefs, S.M (2003). The optimal sport drink. Schw Z Sportmed Traum; 51 (1): 2529
107. Scherman, W., Costill, D., Fink, W., Miller, J (1981). Effect of exrecise-diet manipulation on muscle glycogen and its subsequent utilisation during performance. Int J Sports Med; 114: 114-118
108. Shimomura, Y.,Yamamoto, Y., Bajotto, G.,Sato J., Murakami, T., Shimomura, N., Kobayashi, H., Mawatariyy, K (2006). Nutraceutical effects of branched-chain amino acids on skeletal muscle. J Nutr; 36: 529S-32S
109. Shirreffs, S. M., Maughan, R.J (1997). Whole body sweat collection in humans: an improved method with preliminary data on electrolyte content. J Appl Physiol; 82(1): 336-341
110. Striegel, H., Niess, A.M (2006). Sportgetränke. Deutsche Zeit Sportmed; 57(1): 27-28
111. Suryawan, A., Hawes, J.W., Harris, R.A., Shimomura, Y., Jenkins, A.E., Hutson, S.M (1998). A molecular model of human branched-chain amino acid metabolism. Am J Clin Nutr; 68: 72-81
112. Tarnopolsky, M.; Cupido, C (2000). Caffeine potentiates low frequency skeletal muscle force in habitual and nonhabitual caffeine consumers. J Appl Physiol; 89: 1719-1724
113. Tarnopolsky, M.A (2003). Females and males: should nutritional recommendations be gender specific?. Schweizerische Zeitschrift für «Sportmedizin und Sporttraumatologie»; 51 (1): 3946
114. Tarnopolsky, M.A., Gibala, M., Jeukendrup, A.E., Phillips, S.M (2005). Nutritional needs of elite endurance athletes. Part I: Carbohidrate and fluid requirements. Eur J Sport Sci; 5(1): 3-14
115. Tarnopolsky, M.A., Gibala, M., Jeukendrup, A.E., Phillips, S.M (2005). Nutritional needs of elite endurance athletes. Part II: Dietary protein and the potential role of caffeine and creatine. Eur J Sport Sci; 5(2): 59-72
116. Tarnopolsky, M.A., Zawada, C., Richmond, L.B., Carter, S., Shearer, J., Graham, T., Phillips, S.M (2001). Gender differences in carbohydrate loading are related to energy intake. J Appl Physiol; 91: 225-230
117. (2004). The effect of acute branched-chain amino acid supplementation on prolonged exercise capacity in a warm environment. Eur J App Physiol; 93(3): 306-314
118. (2006). USDA Food Search for Windows. Version 1.0, database version SR17
119. Van Loon, L. J. C., Jeukendrup, A.E., Saris, W.H.M., Wagenmakers, A.J.M (1999). Effect of training status on fuel selection during submaximal exercise with glucose ingestion. J Appl Physiol; 87(4): 14131420
120. Van Loon, L.J.C., Schrauwen-Hinderling, V.B., Koopman, R., Wagenmakers, A.J.M, Hesselink, M.K.C, Schaart, G., Kooi, M.E., Saris, W.H.M (2003). Influence of prolonged endurance cycling and recovery diet on intramuscular triglyceride content in trained males. Am J Physiol Endocrinol Metab; 285: E804E811
121. Van Nieuwenhoven, M. A., Brummer, R.-J. M., Brouns, F (2000). Gastrointestinal function during exercise: comparison of water, sports drink, and sports drink with caffeine. J Appl Physiol; 89: 10791085
122. Voces, J., Cabral de Oliveira, A.C., Prieto, J.G., Vila, L., Perez, A.C., Duarte, I.D.G., Alvarez, A.I (2004). Ginseng administration protects skeletal muscle from oxidative stress induced by acute exercise in rats. Braz J Med Biol Res; 37(12): 1863-1871
123. WADA (2004). he world anti-doping code.
124. Warburton, D.E.R., Welsh, R.C., Haykowsky, M.J., Taylor, D.A., Humen, D.P (2002). Biochemical changes as a result of prolonged strenuous exercise. Br J Sports Med;36: 301-303
125. Weight, L.M., Myburgh, K.H., Noakes, T.D (1988). Vitamin and mineral supplementation: effect on them running performance of trained athletes. Am J Clin Nutr; 47: 192-195
126. Williams, J.E (2000). Vitamin E, vitamin C, and exercise. Am J Clin Nutr; 72(suppl): 647S52S
127. Williams, M.H (2004). Dietary supplements and sports performance: introduction and vitamins. J Int Soc Sports Nutr; 1(2):1-6
128. Williams, M.H (2005). Dietary supplements and sport performance: minerals. J Int Soc Sports Nut; 2(1): 43-49
129. Willmert, N., Porcari, J.P., Foster, C., Doberstein, S., Brice, G (2002). The effects of oxygenated water on exercise physiology during incremental exercis and recovery. JEPonline; 5(4): 16-21
130. Wittbrodt, E.T (2003). Maintaining fluid and electrolyte balance during exercise. J Pharm Prac; 16(1): 4550
131. World Health Organization (1999). WHO monograps on selected medicinal plants. Malta
132. Yoshiharu S., Murakami, T., Nakai, N., Nagasaki, M., Harris, R.A (2004). Exercise promotes BCAA catabolism: effects of BCAA supplementation on skeletal muscle during exercise. J Nutr;134: 583S-587S
133. Zderic, T.W., Davidson, C.J., Schenk, S., Byerley, L.O., Coyle, E.F (2004). High-fat diet elevates resting intramuscular triglyceride concentration and whole body lipolysis during exercise. Am J Physiol Endocrinol Metab; 286: E217E225
134. Zhu, I.H, Haas, J.D (1997). Iron depletion without anemia and physical performance in young women. Am J Clin Nutr; 66: 334-341
135. Zimmermann, M.B (2003). Vitamin and mineral supplementation and exercise performance. Schweizerische Zeitschrift für «Sportmedizin und Sporttraumatologie»; 51 (1): 5357
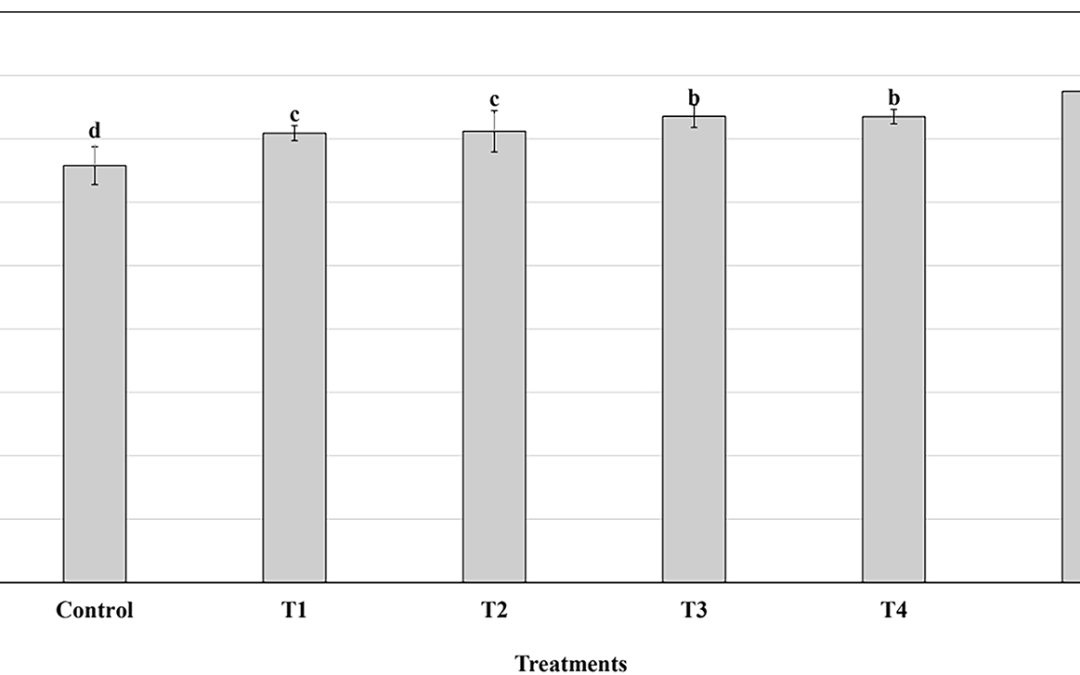
Mejora de las propiedades nutricionales y de textura del jerky reestructurado con polvo de sangre de pato para aplicaciones en kits de comida
Resumen del Estudio sobre el Uso de Polvo de Sangre de Pato en Jerky Reestructurado En un contexto...
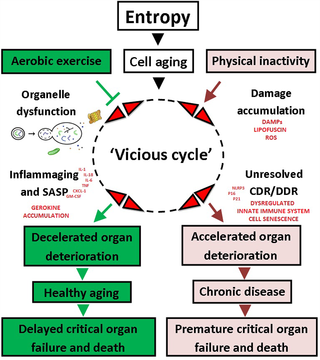
El ejercicio aeróbico a lo largo de la vida protege contra la inflamaging y el cáncer
Ejercicio Aeróbico y su Impacto en el Envejecimiento y el Riesgo de Cáncer La investigación...
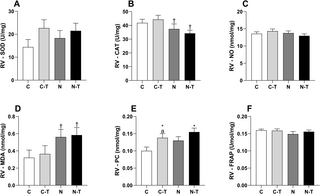
La combinación de nandrolona y entrenamiento de resistencia indujo remodelación cardíaca y estrés oxidativo a pesar de la contractilidad mejorada de los cardiomiocitos
Este artículo investiga cómo la combinación de entrenamiento de resistencia (RT) y el uso de...
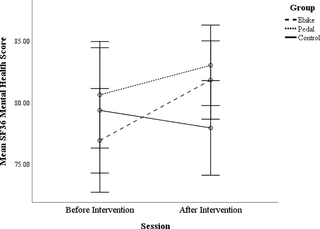
El efecto del ciclismo en la función cognitiva y el bienestar en adultos mayores
El impacto del ciclismo al aire libre en la función cognitiva y bienestar de los adultos mayores...
Predicción de las salidas de potencia mecánica anaeróbica del Wingate a partir de una prueba de esfuerzo cardiopulmonar incremental máxima utilizando un enfoque de aprendizaje automático
El Wingate Anaerobic Test (WAnT) se ha establecido como un método efectivo para medir la potencia...