
Adaptaciones al Entrenamiento en el Ciclismo de Ruta – Implicancias para el Rendimiento
John A Hawley1 y Nigel K Stepto1
Exercise Metabolism Group, School of Medical Sciences, Faculty of Health Sciences, RMIT University, Bundoora, 3083, Australia.
Artículo publicado en el journal PubliCE, Volumen 0 del año 2001.
Publicado 12 de marzo de 2008
Resumen
Palabras clave: ciclismo de ruta, músculo esquelético, combustibles
INTRODUCCION
Los requerimientos fisiológicos para el éxito en el ciclismo de ruta son únicos y han sido recientemente revisados (1, 2). Los ciclistas compiten en competiciones de etapas múltiples y con salida masiva, las cuales están caracterizadas por cambios estocásticos en la tasa de trabajo y/o la velocidad, los cuales son altamente dependientes del terreno (3, 4) y las tácticas de equipo (1, 5). De manera contraria durante las pruebas contrarreloj individuales (5, 6) y por equipos (5), el ciclista/equipo sostiene la mayor producción de potencia promedio (W) posible para la duración de la competición. Así, los programas de entrenamiento para mejorar el rendimiento en el ciclismo de ruta deberían evocar múltiples adaptaciones que le permiten al atleta incrementar la producción de energía a partir de tanto las vías aeróbicas como las independientes de oxígeno y retrasar el inicio de la fatiga muscular.
Desafortunadamente, nuestros conocimientos científicos actuales acerca de los efectos de las intervenciones de entrenamiento específicas llevadas a cabo por ciclistas profesionales sobre respuestas adaptativas selectas y sus consecuencias para el rendimiento de resistencia son limitados. Aunque muchos datos de entrenamiento han sido recolectados por científicos del deporte afiliados con equipos nacionales y profesionales (Martin DT, comunicación personal), solo pudimos encontrar un estudio publicado que detalló las respuestas de la producción de potencia y la frecuencia cardiaca (HR) de un ciclista profesional a una única sesión de entrenamiento intervalado (7). De este modo, nosotros hemos llevado a cabo una serie de investigaciones para estudiar los efectos del entrenamiento intenso a corto plazo (3 a 6 semanas) sobre adaptaciones metabólicas y de rendimiento en ciclistas bien entrenados (8-13). Aunque los ciclistas de este estudio eran todos de nivel competitivo [máximo consumo de oxígeno (VO2 máx.)>5,0 L/min] que entrenaban entre 300 y 500 km por semana, ellos no alcanzaban los criterios fisiológicos o de rendimiento de un ciclista de elite o de clase mundial (1). Desafortunadamente, es extremadamente difícil conducir estudios que incorporen incluso técnicas invasivas moderadas cuando se estudia a los ciclistas más competitivos del mundo: solo recientemente los científicos de deporte han sido capaces de obtener datos de campo y laboratorio a partir de ciclistas de clase mundial (3, 5, 6, 14).
Las respuestas fisiológicas generales al entrenamiento de resistencia en individuos moderadamente entrenados han sido bien documentadas (15). Es interesante para esta revisión destacar aquellas adaptaciones fisiológicas que ocurren como respuesta a los programas de entrenamiento de volumen e intensidad extremadamente altos adoptados por los ciclistas profesionales. Naturalmente el entrenamiento tiene un impacto sobre el rendimiento en ciclistas competitivos de nivel nacional genéticamente dotados, pero la pregunta es la siguiente: “¿Qué adaptaciones fisiológicas son responsables de los incrementos observados en el rendimiento?”. Debido a la escasez de estudios científicos en esta área, presentamos un modelo teórico acerca de las principales adaptaciones inducidas por el entrenamiento en el músculo esquelético que probablemente determinan y mejoran la capacidad de rendimiento en ciclistas de elite. Una premisa de trabajo es que los cambios inducidos por el entrenamiento en el músculo esquelético como resultado del entrenamiento de volumen e intensidad elevados son al menos parcialmente responsables de las mejoras observadas en el rendimiento. Usando datos experimentales proporcionamos evidencia que apoya el modelo.
EL ESTIMULO DE ENTRENAMIENTO
Los componentes clave de cualquier programa de entrenamiento que tenga como objetivo mejorar el rendimiento en el ciclismo son el volumen, la frecuencia y la intensidad (16). La suma de estas tres variables de entrenamiento puede ser cuantificada como el impulso de entrenamiento (17) que puede mejorar (nivel de entrenamiento incrementado) o disminuir (fatiga) el rendimiento. A cualquier tiempo, el “índice de rendimiento predicho” es la diferencia entre el nivel de entrenamiento incrementado y la fatiga (18). Una pregunta importante de interés para un ciclista es la siguiente: “¿Cómo puedo alcanzar mi mejor rendimiento en un día determinado?”. De manera similar, una pregunta importante de interés para los técnicos y científicos del deporte es la siguiente: “¿Cómo diseñamos un programa de entrenamiento para producir un rendimiento máximo (en un día determinado)”?. Desafortunadamente, las respuestas a estas preguntas no pueden estar basadas en resultados salidos de estudios publicados y bien controlados llevados a cabo en ciclistas de elite, debido a que tales datos no existen. En cambio contamos con reportes anecdóticos y esporádicos acerca de los hábitos de entrenamiento de unos pocos ciclistas de elite (1, 7), y observaciones empíricas basadas en el campo de algunos entrenadores destacados (8, 19).
La Tabla 1 muestra un resumen de un “programa típico” llevado a cabo por un ciclista de ruta profesional de clase mundial durante un período de entrenamiento de pretemporada de 3 semanas. Aunque esta información da una idea acerca del volumen de entrenamiento llevado a cabo por ciclistas profesionales durante una fase específica de su programa, no proporciona indicación alguna acerca de la intensidad de entrenamiento (i.e., producción de potencia, velocidad, FC), ni del tiempo que el ciclista ha estado realizando este régimen. El bloque de entrenamiento identificado en la Tabla 1 refleja solo una parte del estímulo de entrenamiento total de este ciclista, el cual incluye recorrer > 30000 km por año y competir ~ 100 días por año! (1).
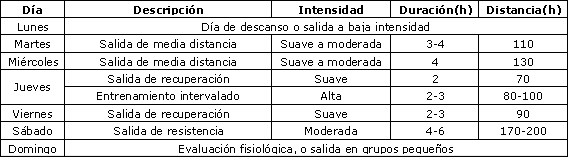
Tabla 1. Programa de entrenamiento de un ciclista de ruta de Clase Mundial durante un período de entrenamiento de pretemporada de 2-3 semanas. Datos de Hawley & Burke (16).
EL MODELO
En la Figura 1 se presenta un modelo teórico de tres áreas principales de adaptación en el músculo esquelético que probablemente contribuyen al rendimiento exitoso de ciclistas de elite. El modelo es una extensión de un modelo desarrollado originalmente por Coyle (20) que describía factores fisiológicos determinantes de la capacidad de rendimiento de resistencia en corredores de larga distancia, ciclistas, y marchistas. Para poder tener acceso a una descripción completa del modelo, se refiere al autor a las revisiones de Coyle (20, 21).
Nuestro modelo consiste de 3 componentes altamente interdependientes: la magnitud de cualquier adaptación inducida por el entrenamiento dentro de un componente tiene probabilidades de estar directamente relacionada al estímulo general del entrenamiento. La premisa de trabajo es que la suma de las adaptaciones dentro de los componentes del modelo ejerce finalmente una influencia importante sobre la capacidad de rendimiento de un ciclista en un tiempo dado.
Morfología del Músculo Esquelético
El componente morfológico del modelo incluye, pero no está limitado, a: (i) tipo de fibras musculares del músculo esquelético; (ii) actividad de las enzimas glucolíticas y oxidativas; (iii) densidad capilar; (iv) volumen magro del muslo; y (v) reclutamiento neuromuscular. Varios de estos factores morfológicos han sido discutidos previamente (20). Una alta proporción de fibras tipo I en el músculo vasto lateral está asociado con un menor costo de oxígeno submáximo (i.e., una mayor eficiencia general o gross efficiency) durante el ciclismo (22), posiblemente debido a un menor recambio o turnover de ATP durante la contracción (23). Teniendo en cuenta que las fibras tipo I también poseen una mayor densidad capilar y un mayor potencial oxidativo que las fibras tipo II (24), estos factores pueden influenciar la eficiencia general en el ciclismo. Sin embargo, los datos transversales revelan que el número de fibras tipo I en el músculo vasto lateral de un ciclista está altamente relacionado al número de años invertidos realizando entrenamiento de resistencia (25). Así, aquellos ciclistas con un gran porcentaje de fibras tipo I son más competitivos y avanzados en el ranking profesional, o la conversión de tipo de fibras desde tipo II a tipo I está ocurriendo. Por consiguiente, si ocurre la transformación de fibras, uno esperaría que la eficiencia general mejore a través del transcurso de la carrera de un ciclista. Desafortunadamente, no hay investigaciones longitudinales que hayan reportado tal fenómeno. Tales estudios están garantizados debido a que pequeñas mejoras en la eficiencia de ciclismo tiene el potencial de resultar en grandes mejoras en el rendimiento. Por ejemplo, Jeukendrup et al. (1) han calculado que para un ciclista de 70 kg que puede sostener una producción de potencia de 400 W durante 1 hora, un incremento del 1% en la eficiencia general resultaría en una mejora de 48 segundos en el tiempo de una prueba contrarreloj de 40 km.
Además de la importancia de una alta proporción de fibras tipo I en los músculos que trabajan, el rol de las fibras tipo IIa para alcanzar un rendimiento exitoso en el ciclismo de ruta no debería ser pasado por alto. Las fibras tipo IIa tienen un potencial oxidativo similar que las tipo I y a intensidades de competición tienen una mayor capacidad de generación de fuerza (24).
Maximal sustainable power output: Máxima Producción de Potencia que puede ser sostenida; Volume: Volumen; Intesity: Intensidad; Frequency: Frecuencia; Years of endurance training: Años de entrenamiento de resistencia; Training stimulus: Estímulo de Entrenamiento; Recovery: Recuperación; Skeletal muscle adaptation: Adaptación del músculo esquelético; Morphological: Morfológico; Type I fibres: Fibras Tipo I; Enzime activity: Actividad enzimática; Capillary density: Densidad capilar; Lean thigh volume: Volumen magro del muslo; Neural recruitment: Reclutamiento neural; Fuel supply: Aporte de combustibles; Fat oxidation: Oxidación de grasas; IMTG content: Contenido de IMTG; FABP and FAT: FABP y FAT; Muscle glycogen stores: Reservas de glucógeno muscular; GLUT4: GLUT 4; Blood glucose ox: Oxidación de glucosa sanguínea; Muscle glycogen utilisation: Utilización de glucógeno muscular; Acid-base status: Nivel ácido-base; Muscle and blood lactate concentration: Concentración de lactato muscular y sanguíneo; Lactate threshold: Umbral del lactato; MCT1 and MCT4: MCT1 y MCT4; ßm: ßm; Lactate turnover and oxidation: Recambio y oxidación de lactato.
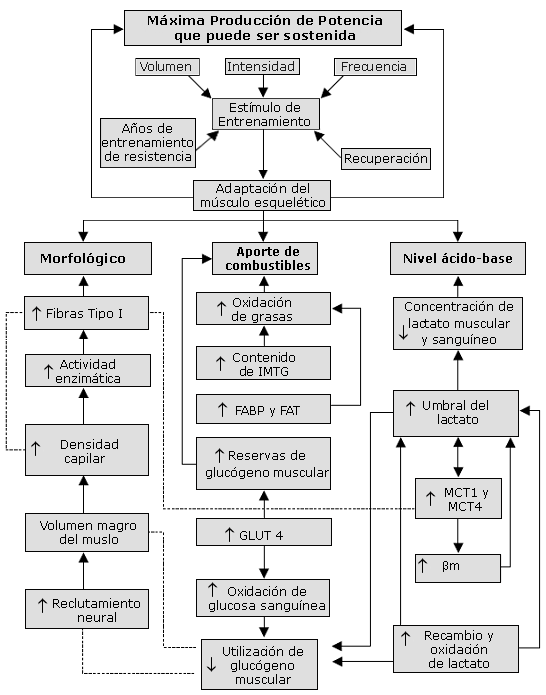
Figura 1. Modelo de algunas de las adaptaciones principales que ocurren en el músculo esquelético como resultado del entrenamiento de alto volumen e intensidad llevado a cabo por los ciclistas profesionales. FABP: proteína ligadora de ácidos grasos; FAT: translocasa de ácidos grasos; GLUT4: proteína transportadora de glucosa; IMTG: triacilglicéridos intramusculares; MCT: transportador monocarboxilato; ßm: capacidad buffer de músculo; ?: indica incremento; ?: indica disminución.
Aunque ha sido reportado que las actividades de enzimas oxidativas selectas del músculo esquelético (i.e., citrato sintetasa) son más altas en un grupo de ciclistas de “elite” versus ciclistas competitivos de “buen” nivel (25), otros autores han encontrado que la actividad enzimática no correlaciona bien con la capacidad de resistencia en el ciclismo (13). Así, en algunos casos otros factores diferentes a la capacidad oxidativa del músculo son importantes para alcanzar un rendimiento exitoso en el ciclismo. Uno de esos factores podría ser la densidad capilar muscular: una alta densidad capilar incrementaría la perfusión por medio de la reducción de la distancia de difusión y facilitaría la remoción de lactato desde los músculos (26).
El volumen magro del muslo de los ciclistas de elite es similar al de los ciclistas amateurs altamente entrenados (9, 25), por lo que este componente morfológico no puede explicar las grandes diferencias en las producciones de potencia máximas y submáximas sostenibles entre estos grupos de ciclistas. Por el contrario, Coyle et al. (25) han reportado que ciclistas con una capacidad de rendimiento superior generan una mayor producción de potencia cuando pedalean, adoptando una estrategia de pedaleo que permite desarrollar mayores fuerzas verticales durante la fase descendente del pedaleo. Esta técnica superior puede estar relacionada al número de años de entrenamiento de resistencia (25). Lo que bien puede ocurrir es que un componente importante de entrenamiento sostenido e intenso a través de un período de muchas temporadas puede alterar directamente la contractilidad muscular (27).
Parte de esta contractilidad muscular alterada puede estar relacionada a la mejora de los patrones de reclutamiento neuromuscular que se adaptarían en respuesta a patrones de sobrecarga específicos para el deporte. Datos recientes (28) indican que ocurren cambios neuromusculares marcados [electromiograma (EMG)/índice de fuerza] tanto en los músculos agonistas como antagonistas de las piernas durante series de esprints de 15 x 5 segundos: a medida que la fatiga se incrementa (i.e., declina la potencia) hubo una “coordinación intermuscular” para transferir fuerza al pedal. Tal adaptación podría estar mediada centralmente y/o ocurrir debido a cambios reflejos causados por los órganos tendinosos de Golgi. Con respecto a esto, es interesante señalar que la vasta mayoría de las mejoras en la fuerza muscular dinámica observadas después del entrenamiento de la fuerza de corta duración son atribuidas al incremento de de la activación voluntaria del músculo (i.e., mejor reclutamiento neural) en vez de a cualquier incremento mensurable en el tamaño de las fibras musculares (29). Además, el rendimiento de resistencia puede ser mejorado independientemente de los cambios en el VO2 máx. y la economía de esfuerzo (30). Tomadas colectivamente estas observaciones sugieren que el entrenamiento de resistencia prolongado e intenso conducido a través de muchas temporadas puede inducir adaptaciones neuromusculares que le permiten a los atletas reclutar un mayor número de fibras musculares y así distribuir la producción de potencia sobre una mayor masa muscular activa mientras se pedalea, en comparación con los ciclistas menos entrenados (25).
Nivel Ácido-Base
Una premisa central del modelo propuesto por Coyle (20) es que el umbral del lactato (LT) individual constituye una “capacidad funcional” importante que está altamente relacionada a la máxima producción de potencia sostenible por el ciclista (20, 31). Efectivamente, uno de los principales atributos de los ciclistas profesionales de ruta es su capacidad para sostener altas producciones de trabajo absolutas por períodos de tiempo prolongados mientras mantienen niveles de lactato sanguíneo en estado estable (6).
Actualmente está ampliamente aceptado que el lactato juega un rol importante como un compuesto rico en energía durante el ejercicio sostenible de intensidad elevada (32, 33). Efectivamente, durante el ciclismo progresivo, MacRae et al. (34) ha reportado que hasta el 80% del lactato es oxidado y explica ~ el 45% de la oxidación total de carbohidratos. Por consiguiente, la capacidad de transportar lactato y H+fuera de las fibras musculares, y la capacidad del músculo esquelético de captar lactato son probablemente determinantes claves de la producción de potencia de un ciclista en el LT.
El transporte de lactato a través del sarcolema del músculo esquelético está mediado por los transportadores monocarboxilato de protones (MCT), el sistema de transporte de la membrana con la mayor capacidad de transporte de H+durante el ejercicio intenso (35-37). Han sido clonadas dos isoformas de transportadores de lactato/H+: MCT1 y MCT4 (37, 38). En los humanos, el contenido de MCT1 es mayor en las fibras tipo I, mientras que el contenido de MCT4 parece ser bastante similar en diferentes músculos (37, 39). Es interesante señalar el hallazgo que indica que los individuos con una alta proporción de fibras tipo I en sus músculos activos van a tener también un alto contenido de MCT1 en los fibras tipo II en comparación con los sujetos que poseen menos fibras tipo I (37). Teniendo en cuenta que un músculo que tiene principalmente fibras tipo II tiene el 50% de la capacidad de transporte de lactato de un músculo compuesto de fibras tipo I, es obvia la importancia de estas proteínas transportadoras unidas a la membrana para el rendimiento en el ciclismo de ruta.
Pilegaard et al. (39) estudiaron la capacidad de transporte de lactato a través del sarcolema en individuos de diferente nivel de entrenamiento. La relación entre la capacidad de transporte de lactato y la capacidad aeróbica es presentada en la Figura 2. La capacidad de transporte de lactato fue significativamente mayor en los atletas en comparación con los individuos desentrenados. Sin embargo, la capacidad para transportar lactato a través del sarcolema no estuvo directamente relacionada al VO2 máx. del individuo. Contrariamente, el transporte de lactato estuvo positivamente correlacionado con el número de fibras tipo I del vasto lateral (r=0,48, p<0,01), de las cuales los atletas entrenados en resistencia tuvieron una proporción significativamente mayor (39). Aunque estos datos transversales no permiten hacer inferencias acerca de la causalidad entre el transporte de lactato y el nivel de entrenamiento, solo aquellos atletas entrenados en resistencia con un VO2 máx. > 65 ml.kg-1.min-1tenían una capacidad elevada de transporte de lactato (39). En este estudio (39), la densidad capilar muscular fue similar para todos los atletas entrenados en resistencia y no estuvo relacionada a la capacidad de transporte de lactato. Así, parece que solo un gran volumen de entrenamiento de resistencia constituye un estímulo insuficiente para mejorar la capacidad de transporte de lactato.
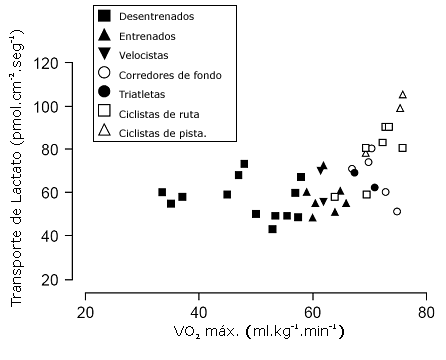
Figura 2. Relación entre la capacidad de transporte de lactato muscular y el máximo consumo de oxígeno (VO2 máx.) en individuos desentrenados (n=13); individuos entrenados (n=7); velocistas (n=2); corredores de fondo (n=5), triatletas (n=3), ciclistas de ruta (n=6) y ciclistas de pista (n=3) [adaptado de Pilegaard et al. (39) con permiso de la Sociedad Fisiológica Americana].
Efectivamente, los mayores valores de transporte de lactato fueron observados en 2 ciclistas de pista cuya especialidad eran los 4000 m de persecución (uno de los ciclistas había ganado la medalla de bronce en los Juegos Olímpicos de 1992). Estos ciclistas estaban entrenando un promedio de ~ 700 km por semana, incluyendo varias sesiones de pista de alta intensidad (39). Este hallazgo destaca la importancia de incluir sesiones de entrenamiento de alta intensidad como preparación para las competiciones tanto de ruta como de pista. También podría explicar porque un programa de entrenamiento supramáximo de corta duración (6 sesiones de 12 x 30 s a ~ 650 W) fue tan efectivo para incrementar el rendimiento en una prueba de ciclismo contrarreloj de 40 km (que duró ~ 55 min) como series de entrenamiento intervalado “aeróbico” prolongadas (6 sesiones de 8 x 4 minutos a ~ 330 W) realizadas a un ritmo cercano al de la prueba contrarreloj de 40 km (10). Efectivamente, el mínimo aparente en la mejora del rendimiento en 40 km luego de las series de entrenamiento supramáximas versus las series de entrenamiento intervalado sugieren que puede haber más de un mecanismo por el cual el entrenamiento intervalado mejora el rendimiento de resistencia.
Ha sido reportado que la capacidad buffer muscular (ßm) aumenta en un grado similar (~16%) luego de el entrenamiento de esprint de alta intensidad (40) y cuando el ciclista entrenado en resistencia reemplaza una parte de su entrenamiento de base por series intervaladas aeróbicas (5 minutos) sostenidas (13). Ya que el entrenamiento de alta intensidad mejora la capacidad de transporte de lactato/H+del sarcolema, así como el contenido de proteínas MCT1 y MCT4 en el músculo esquelético humano (41), y teniendo en cuenta que tales adaptaciones inducidas por el entrenamiento son probablemente importantes para la regulación del lactato y el pH muscular, creemos que es razonable recomendar que sean incorporadas a la preparación para la competición de todos los ciclistas cargas de trabajo de esprint repetidos supramáximos.
Aporte de Combustibles
El ciclismo de ruta induce múltiples adaptaciones profundas en muchos sistemas fisiológicos (i.e., muscular, cardiovascular, endocrino). Un efecto importante de estas adaptaciones implica la modificación de las tasas a las cuales son utilizados las fuentes de combustibles principales (carbohidratos, y grasas) durante el ejercicio prolongado e intenso. Quizás la adaptación al entrenamiento aislada más impresionante que puede aumentar el rendimiento en el ciclismo es que en comparación con los ciclistas menos entrenados, los ciclistas altamente entrenados oxidan menos carbohidratos y más grasa durante el ejercicio realizado a la misma producción de potencia o consumo de oxígeno (VO2), absolutos (42), aunque generalmente tienen reservas de glucógeno iniciales más altas (43). Teniendo en cuenta que el agotamiento de las reservas de carbohidratos endógenas del cuerpo es un factor relacionado a la fatiga durante el ciclismo prolongado y hasta el agotamiento (44, 45), este desplazamiento inducido por el entrenamiento en la selección de sustratos por los músculos que trabajan indudablemente juega un rol importante en la mejora observada en la capacidad de resistencia luego del entrenamiento de resistencia.
Parte de ente desplazamiento inducido por el entrenamiento en la selección de sustratos (desde carbohidratos a grasas) a la misma tasa de trabajo absoluta ha sido atribuido a la mejora de la sensibilidad del control respiratorio que resulta de un incremento de la densidad mitocondrial muscular (46). Sin embargo, Coyle et al. (31) han reportado previamente que en ciclistas con un VO2 máx. (~67 ml.kg-1.min-1) y actividad enzimática similar, la utilización de glucógeno muscular durante el ciclismo intenso (i.e., 79% del VO2 máx.) fue 2 veces mayor en individuos con una baja producción de potencia en el LT individual en comparación con los individuos con una alta producción de potencia en el LT. Westgarth-Taylor et al. (12) han reportado que el entrenamiento de alta intensidad y de corta duración en 8 ciclistas previamente bien entrenados (VO2 máx. de 5,2 L/min) disminuyó significativamente la tasa de oxidación de carbohidratos (desde 3,2, 4,3 y 5,4 g/min hasta 2,9, 3,6 y 4,8 g/min en el 60, 70, y 80% del VO2 máx. preentrenamiento, respectivamente), a pesar de no lograr cambios en la capacidad oxidativa muscular (13). Teniendo en cuenta todos estos datos parece que el desplazamiento inicial inducido por el entrenamiento en la utilización de sustratos (desde carbohidratos a grasas) está causado por incrementos en la capacidad respiratoria muscular. Sin embargo, en ciclistas con una historia prolongada de entrenamiento de resistencia, quienes presumiblemente han maximizado el potencial oxidativo de sus músculos entrenados, otros factores deben ser importantes (i.e., reclutamiento neuromuscular de una mayor masa muscular, nivel ácido-base, etc.) para los desplazamientos subsiguientes en los patrones de metabolismo de combustibles.
Parte de la reducción en la utilización de carbohidratos observada luego del entrenamiento prolongado e intenso podría ser causada por una disminución en la captación de glucosa en el músculo (47-49), aunque ha sido demostrado que el entrenamiento de resistencia incrementa el contenido de GLUT-4 (49, 50). Un estudio reciente (51) reportó que cuando los niveles de glucógeno muscular eran bajos, la captación de glucosa era mayor en los individuos entrenados versus los desentrenados, trabajando a la misma tasa de trabajo relativa. Esto explicaría porque los ciclistas de ruta altamente entrenados son capaces de utilizar glucosa a tasas notablemente mayores durante el ejercicio prolongado y submáximo en el momento en que las reservas de glucógeno son bajas (45, 52, 53) y porque los carbohidratos orales administrados a los ciclistas bien entrenados (VO2 máx. de 69 ml.kg-1.min-1) durante el ejercicio prolongado (3 horas) resulta en una síntesis de glucógeno muscular neta en las fibras no activas (54).
La reducción inducida por el entrenamiento en la oxidación de carbohidratos observada en los atletas entrenados puede ser completamente compensada por el incremento concomitante en la oxidación de grasas. Ha sido reportado que el nivel de triacilglicéridos intramusculares (IMTG) disminuye después de varias horas de ejercicio submáximo en atletas bien entrenados (55). Además, el contenido de IMTG disminuye durante una serie estandarizada de ciclismo submáximo en individuos que completaron un programa de entrenamiento intenso de 12 semanas (56), aunque otros autores han fallado en observar un fenómeno similar (57). Teniendo en cuenta que el entrenamiento no incrementa la captación neta de ácidos grasos libres (FA) por parte de los músculos que trabajan (58-60), es probable que los FAs adicionales que son oxidados durante el ejercicio sean derivados principalmente de las reservas de IMTG. Es interesante la evidencia reciente acerca de los transportadores de ácidos grasos de cadena larga (LCFA) en el músculo esquelético (61). Bonen y colaboradores han reportado que el transporte de LCFA es mayor en las fibras tipo I que en las tipo II (62), y que cuando la capacidad oxidativa del músculo se incrementa hay un incremento paralelo en la tasa de transporte de FA en la membrana del sarcolema (63). Tales adaptaciones, aunque fueron reportadas en individuos moderadamente a bien entrenados, son probablemente altamente funcionales en atletas mejor entrenados que realizan ejercicio prolongado e intenso, debido a que permiten a los músculos incrementar su metabolismo de FA en un momento en el cual el contenido de glucógeno muscular es bajo.
EVIDENCIA EXPERIMENTAL PARA APOYAR EL MODELO
Nosotros hemos descrito recientemente las demandas musculares y metabólicas del entrenamiento intervalado intenso en ciclistas de resistencia (11). En aquel estudio, 7 ciclistas bien entrenados (edad: 27±5 años, masa: 76±3 kg, VO2 máx.: 5,14±0,23 L/min) que estaban realizando 368±141 km por semana y que habían estado realizando entrenamiento de resistencia regular durante ~ 8 años, realizaron series de trabajo de 8 x 5 min al 86±2 % del VO2 máx (334±18 W) con 60 segundos de recuperación. Las biopsias musculares fueron tomadas del músculo vasto lateral inmediatamente antes y después de la sesión de entrenamiento, mientras eran muestreados el intercambio de gases pulmonares y la sangre venosa a intervalos regulares durante el ejercicio. Comparamos nuestros datos (11) con los de un ciclista profesional que realizó la misma serie de entrenamiento intervalado a la misma intensidad relativa de ejercicio (series de trabajo de 8 x 5 minutos al 86% del VO2 máx., 358 W), bajo las mismas condiciones de laboratorio. Las características del ciclista profesional fueron las siguientes: 36 años, masa 77 kg, VO2 máx. 5,4 L/min. Se encontraba realizando 600 km por semana y había estado realizando entrenamiento de resistencia prolongado durante 19 años.
A pesar de la mayor tasa de trabajo absoluta de 7% (358 vs. 334 W) y la elevación asociada en la ventilación (VE) [111±5 vs. 107±8 L/min] y VO2 (4,7±0,3 vs. 4,3±0,3 L/min], el ciclista profesional tuvo un índice de intercambio respiratorio menor (0,92±0,002 vs 4,3±0,3) y menores tasas de oxidación de carbohidratos (300±12 vs. 336±52 µmol.kg-1.min-1) que los ciclistas bien entrenados. Por consiguiente, las tasas de oxidación de grasas fueron mayores en el ciclista profesional en comparación con el grupo bien entrenado (28±8 vs 22±10 µmol.kg-1.min-1). La menor tasa de oxidación de carbohidratos podría ser explicada por una menor tasa de oxidación de glucógeno (Figura 3a). Quizás la observación más notable fue la pequeña perturbación del nivel ácido-base en el ciclista profesional a una alta tasa de trabajo: el nivel de lactato sanguíneo fue 20% menor (3,0±0,3 vs 5,2±1,7 mmol/L) y el nivel de lactato muscular fue un 34% menor que en el grupo de ciclistas bien entrenados (Figura 3b). Fue interesante el hallazgo de que el pH fue similar (Figura 3c) indicando que este parámetro puede no ser la medición más sensible del nivel ácido-base. Los índices de esfuerzo percibido subjetivo en la escala de Borg de 20 puntos (64) tomados a través de todas las series de entrenamiento intervalado a la misma intensidad relativa fueron 8±1 para el ciclista profesional (“muy suave”) versus 13±1 para el grupo de ciclistas bien entrenados (“algo duro”). De manera contraria a estudios anteriores (22, 25), la economía de pedaleo (79 W/L) no fue diferente entre el grupo de ciclistas bien entrenados y el ciclista profesional que había estado realizando entrenamiento de resistencia de alto volumen e intensidad durante un período del doble de duración (19 vs 8 años).
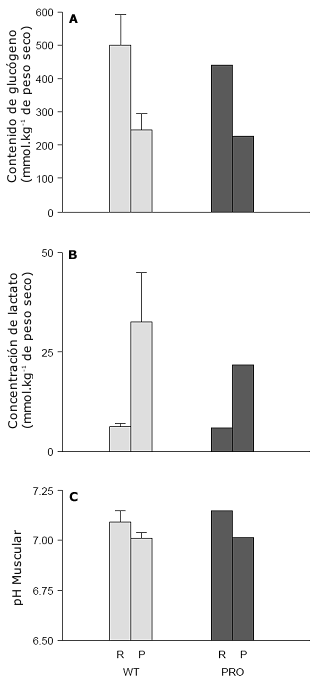
Figura 3. Respuestas del músculo esquelético a una sesión de entrenamiento intervalado aeróbico intenso en un grupo de ciclistas competitivos bien entrenados (WT) (n=7) y un ciclista de ruta profesional (PRO, ver sección Evidencia Experimental para Apoyar el Modelo para ver las características del participante). Luego de una entrada en calor estandarizada, todos los ciclistas realizaron 8 series de trabajo de 5 minutos de duración al 88% del máximo consumo de oxígeno con 60 segundos de recuperación a ~ 100 W. Las biopsias musculares fueron tomadas del músculo vasto lateral en reposo (R) y luego del ejercicio (P). Los datos del grupo pertenecen a Stepto et al. (11).
CONCLUSIONES
Nuestros conocimientos actuales acerca de los efectos específicos de las intervenciones de entrenamiento llevadas a cabo por ciclistas profesionales sobre las respuestas adaptativas en el músculo esquelético y sus consecuencias para el rendimiento, son limitados. Debido a la escasez de estudios científicos, nosotros presentamos un modelo teórico acerca de las principales adaptaciones al entrenamiento que ocurren en el músculo esquelético y que probablemente determinan y mejoran la capacidad de los ciclistas de elite. Una premisa de trabajo fue que tales perturbaciones resultan del entrenamiento de alto volumen e intensidad realizado por estos ciclistas. Usando datos experimentales, nosotros proporcionamos evidencia que apoya este modelo.
Este artículo destaca el hecho de que nuestros conocimientos acerca de las bases fisiológicas de las técnicas de entrenamiento de resistencia empleadas por los ciclistas profesionales son bastante elementales. Desde luego hay muchas preguntas sin contestar que pueden proporcionar áreas fértiles para la investigación para los científicos del deporte. Estas incluyen, pero no están limitadas a:
- (i) los efectos del entrenamiento intenso sobre el recambio o turnover de lactato en ciclistas entrenados;
- (ii) los efectos del entrenamiento intenso sobre los transportadores de lactato en el músculo esquelético;
- (iii) los efectos de diferentes intervenciones de entrenamiento sobre los patrones de reclutamiento muscular y
- (iv) los efectos de diferentes intervenciones de entrenamiento sobre vías de señalización intracelular en el músculo esquelético y sus consecuencias para el control de la utilización de sustratos.
Agradecimientos
Queremos agradecer al Dr David T. Martin por su excelente revisión de este manuscrito y su valioso aporte.
Referencias
1. Jeukendrup AE, Craig NP, Hawley JA (2000). The bioenergetics of world class cycling. J Sci Med Sport 3: 414-33
2. Mujika I, Padilla S (2001). Physiological and performance characteristics of male professional road cycling. Sports Med; 31 (7): 479-87
3. Padilla S, Mujika I, Cuesta G, et al (1999). Level ground and uphill cycling ability in professional road cycling. Med Sci Sports Exerc; 31: 878-85
4. Palmer GS, Hawley JA, Dennis SC, et al (1994). Heart rates responses uring a 4-d cycle stage race. Med Sci Sports Exerc; 26: 1278-83
5. Padilla S, Mujika I, Orbananos J, et al (2000). Exercise intensity during competition time trials in professional road cycling. Med Sci Sports Exerc; 32: 850-6
6. Padilla S, Mujika I, Angulo F, et al (2000). Scientific approach to the 1-h cycling world record: a case study. J Appl Physiol; 89: 1522-7
7. Jeukendrup AE, van Dieman A (1998). Heart-rate monitoring during training and competition in cyclists. J Sports Sci; 16 Suppl. S91-S9
8. Hawley JA, Myburgh KH, Noakes TD, et al (1997). Training techniques to improve fatigue resistance and enhance endurance performance. J Sports Sci; 15: 325-33
9. Lindsay FH, Hawley JA,Myburgh KH, et al (1996). Improved athletic performance in highly trained cyclists after interval training. Med Sci Sports Exerc; 28: 1427-34
10. Stepto NK, Hawley JA, Dennis SC, et al (2001). Metabolic demands of intense aerobic interval training in competitive cyclists. Med Sci Sports Exerc; 33: 303-10
11. Stepto NK, Martin DT, Fallon KE, et al (2001). Metabolic demands of intense aerobic interval training in competitive cyclists. Med Sci Sports Exerc; 33: 303-10
12. Westgarth-TaylorC, Hawley JA, Rickard S, et al (1997). Metabolic and performance adaptations to interval training in endurance trained cyclists. Eur J Appl Physiol; 75: 298-304
13. Weston AR, Myburgh KH, Lindsay FH, et al (1997). Skeletal muscle buffering capacity and endurance performance after high intensity interval training by well-trained cyclists. Eur J Appl Physiol; 75: 7-13
14. Brosnan MJ, Martin DT, Hahn AG, et al (2000). Impaired interval exercise responses in elite female cyclists at moderate simulated altitude. J Appl Physiol; 89 (5): 1819-24
15. Saltin B. Physiological effects of physical conditioning (1991). Med Sci Sport; 1: 50-6
16. Hawley JA, Burke LM (1999). Peak performance: training and nutritional strategies for sport. Sydney: Allen and Unwinoxidation in the intracellular lactate shuttle. Proc Natl Acad Sci; 96: 1129-34
17. Bannister EW, Good P, Holman G, et al (1986). Modeling the training response. In: Landers DM, editor. The 1984 Olympic Scientific Congress Proceedings. Vol 3. Sport and elite performers. Champaign (IL): Human Kinetics, : 7-23
18. Morton RH (1997). Modelling training and overtraining. J Sports Sci 1997; 15: 335-40
19. Wells CL, Pate RR (1988). Training for performance of prolonged exercise. In: Lamb DR, Murray R, editors. Perspectives in exercise science and sports medicine. Vol 1. Carmel, (IN): Benchmark Press, : 357-91
20. Coyle EF (1995). Integration of the physiological factors determining endurance performance ability. Exerc Sport Sci Rev; 23: 25-63
21. Coyle EF (1999). Physiological determinants of endurance exercise performance. J Sci Med Sport; 2: 181-9
22. Coyle EF, Sidossis LS, Horowitz JF, et al (1992). Cycling efficiency is related to the percentage of type I muscle fibers. Med Sci Sports Exerc; 24: 782-8
23. Wendt IR, Gibbs CL (1974). Energy production of mammalian fastand slow-twitch muscles during development. Am J Physiol; 226: 644-7
24. Saltin B (1977). Fiber types and metabolic potentials of skeletal muscles in sedentary man and endurance runners. Ann NY Acad Sci 1977; 301: 3-29
25. Coyle EF, Feltner ME, Kautz SA, et al (1991). Physiological and biomechanical factors associated with elite endurance cycling performance. Med Sci Sports Exerc; 23: 93-107
26. Tesch PA, Daniels JL, Sharp DS (1982). Lactate accumulation in muscle and blood during submaximal exercise. Acta Physiol Scand; 114: 441-6
27. Noakes TD (1988). Implications of exercise testing for prediction of athletic performance. Med Sci Sports Exerc; 20: 319-30
28. Hautier CA, Arsac LM, Deghdegh K, et al (2000). Influence of fatigue on EMG/force ratio and co-contraction in cycling. Med Sci Sports Exerc; 32: 839-43
29. KraemerWK, Fleck SJ, EvansWJ (1996). Strength and power training: physiological mechanism of adaptation. In: Holloszy JO, editor. Exercise and Sports Science Reviews. Baltimore (MA): Williams and Wilkins, 363-97
30. Daniels JT, Yarborough RA, Foster C (1978). Changes in VO2max and running performance with training. Eur J Appl Physiol; 39: 249-54
31. Coyle EF, Coggan AR, Hopper MK, et al (1988). Determinants of endurance in well-trained cyclists. J Appl Physiol; 64: 2622-30
32. Brooks GA (1998). Mammalian fuel utilization during sustained exercise. Comp Biochem Physiol A Physiol; 120: 89-107
33. Brooks GA, Dubouchard H, Brown M, et al (1999). Role of mitochondrial lactate dehydrogenase and lactate oxidation in the intracellular lactate shuttle. Proc Natl Acad Sci; 96: 1129-34
34. MacRae HHS, Noakes TD, Dennis SC (1995). Effects of endurance training on lactate removal by oxidation and gluconeogenesis during exercise. Pflugers Arch 430: 964-70
35. Juel C, Pilegaard H. (1999). Lactate exchange and pH regulation in skeletal muscle. In: Hargreaves M, Thompson M, editors. Biochemistry of exercise X. Champaign (IL): Human Kinetics, 185-200
36. Juel C (1995). Regulation of cellular pH in skeletal muscle fiber types, studies with sarcolemmal giant vesicles obtained from rat muscle. Biochem Biophys Acta 1265: 127-32
37. Pilegaard H, Terzis G, Halestrap A, et al (1999). Distribution of the lactate/H+ transporter isoforms MCT1 and MCT4 in human skeletal muscle. Am J Physiol 276: E843-E848
38. Brooks GA, Brown MA, Butz CE, et al (1999). Cardiac and skeletal muscle mitochondria have a monocarboxylate transporter MCT1. J Appl Physiol 87: 1713-8
39. Pilegaard H, Bangsbo J, Richter EA, et al (1994). Lactate transport studied in sarcolemmal giant vesicles fromhuman muscle biopsies: relation to training status. J Appl Physiol 1858-62
40. Bell GJ, Wenger HA (1988). The effect of one-legged sprint training on intramuscular pH and nonbicarbonate buffering capacity. Eur J Appl Physiol 58: 158-64
41. Pilegaard H, Domino K, Noland T, et al (1999). Effect of high-intensity exercise training on lactate/H+ transport capacity in human skeletal muscle. Am J Physiol 276: E255-E261
42. Coggan AR,Williams BD (1995). Metabolic adaptations to endurance training: substrate metabolism during exercise. In: Hargreaves M, editor. Exercise metabolism. Champaign (IL): Human Kinetics, 177-210
43. Hawley JA, Schabort EJ, Noakes TD, et al (1997). Carbohydrate-loading and exercise performance. Sports Med 24: 73-81
44. Bergstrom J, Hermansen L, Hultman E, et al (1967). Diet, muscle glycogen and physical performance. Acta Physiol Scand 71: 140-50
45. Coyle EF, Coggan ER, Hemmert MK, et al (1986). Muscle glycogen utilization during prolonged strenuous exercise when fed carbohydrate. J Appl Physiol 61: 165-72
46. Holloszy JO, Coyle EF (1984). Adaptations of skeletal muscle to endurance exercise and their metabolic consequences. J Appl Physiol 56: 831-8
47. Bergman BC, Butterfield GE, Wolfel EE, et al (1999). Muscle net glucose uptake and glucose kinetics after endurance training in men. Am J Physiol E81-E92
48. CogganAR, KohrtW, Spina R, et al (1990). Endurance training decreases plasma glucose turnover and oxidation during moderateintensity exercise in men. J Appl Physiol 68: 990-6
49. Richter EA, Jensen P,Kiens B, et al (1998). Sarcolemmal glucose transport and GLUT-4 translocation during exercise is diminished by endurance training. Am J Physiol E89-E95
50. Ebeling P, Bourey RE, Koranyi LI, et al (1992). Mechanism of enhanced insulin sensitivity in athletes: increased blood flow, muscle glucose transport protein (GLUT4) concentration, and glycogen synthase activity. J Clin Invest 92: 1623-31
51. Kristiansen S, Gade J, Wojtaszewski JFP, et al (2000). Glucose uptake is increased in trained vs. untrained muscle during heavy exercise. J Appl Physiol 89: 1151-8
52. Coggan AR, Coyle ER (1987). Reversal of fatigue during prolonged exercise by carbohydrate infusion or ingestion. J Appl Physiol 63: 2388-95
53. Hawley JA, Bosch AN,Weltan SM, et al (1994). Glucose kinetics during prolonged exercise in hyperglycaemic and euglycaemic subjects. Pflugers Arch 426: 378-86
54. Kuipers H, Keizer HA, Brouns F, et al (1987). Carbohydrate feeding and glycogen synthesis during exercise in man. Pflugers Arch 410: 652-6
55. Froberg SO, Mossfeldt F (1971). Effect of prolonged, strenuous exercise on the concentration of triglycerides, phospholipids and glycogen in muscle of man. Acta Physiol Scand 82: 161-71
56. Hurley BF, Nemeth PM, Martin WH, et al (1986). Muscle triglyceride utilization during exercise: effect of endurance training. J Appl Physiol 60: 562-7
57. Kiens B, Richter EA (1998). Utilization of skeletal muscle triacylglycerol during post-exercise recovery in man. Am J Physiol 275: E332-E337
58. Henriksson J (1977). Training induced adaptation of skeletal muscle and metabolism during submaximal exercise. J Physiol 270: 661-7
59. Jansson E, Kaijser L (1987). Substrate utilization and enzymes in skeletal muscle of extremely endurance-trainedmen. J Appl Physiol 662, 999-1005
60. Martin WH, Dalsky GP, Hurley BF, et al (1993). Effect of endurance training on plasma free fatty acid turnover and oxidation during exercise. Am J Physiol 28: E708-E714
61. Bonen A, Dyck DJ, Luiken JJFP (1998). Skeletal muscle fatty acid transport and transporters. In: Richter EA, Kiens B, Galbo H, et al., editors. Skeletal muscle metabolism in exercise and diabetes. New York: Plenum Press, 193-205
62. Bonen A, Luiken JJFP, Liu S, et al (1998). Palmitate transport and fatty acid transporters in red and white muscles. Am J Physiol 275: E471-E478
63. Bonen A,DyckDJ, Azeddine I, et al (1999). Muscle contractile activity increased fatty acid metabolism and transport and FAT/CD36. Am J Physiol 276: E642-E649
64. Borg G (1973). Perceived exertion: a note on history and methods. Med Sci Sports 5: 90-3