
Adaptaciones Enzimaticas Generadas por el Entrenamiento de Fuerza a Largo Plazo
Paavo V Komi2, Keijo Häkkinen2 y Per A Tesch1
1Departamento de Medicina Ambiental, Instituto Karolinska, Estocolmo, Suecia.
2Departamento de Biología de Actividad Física, Univ. de Jyvaskyla, Finlandia.
Artículo publicado en el journal PubliCE, Volumen 0 del año 1997.
Publicado 30 de abril de 2004
Resumen
Palabras clave: fuerza explosiva, entrenamiento de fuerza-resistencia, hipertrofia, tamaño de fibra muscular, fuerza
INTRODUCCION
Es generalmente aceptado que consecuente al entrenamiento de fuerza a largo plazo, las adaptaciones neurales principales, resultante de una activación incrementada de unidades motoras, y un aumento de la síntesis de proteínas contráctiles que conducen a una hipertrofia muscular, provocan un aumento de fuerza muscular (7, 6, 13, 18, 20). En tanto las adaptaciones enzimáticas musculares asociadas con el entrenamiento de resistencia han sido extensivamente estudiadas (por ej., ref. 21), nuestro conocimiento concerniente a la respuesta metabólica al entrenamiento con pesas es escaso. Algunos autores reportaron una actividad incrementada de enzimas que reflejan una función contráctil (26), oxidativa (4, 12) ó glucolítica (4, 14) mejorada, subsecuente al entrenamiento de fuerza. Sin embargo, estos hallazgos no son consistentes (6, 10, 13). Parece ser que las discrepancias son debido a la carga, intensidad y duración del entrenamiento ejecutado, ó por el nivel de actividad física de los sujetos investigados.
La demostración de una menor densidad capilar (23) y la reducción de la densidad mitocondrial (17) en músculos de atletas entrenados en fuerza, comparados con sujetos no entrenados, sugiere que la actividad de marcadores enzimáticas para la «maquinaria de energía aeróbica» está reducida, subsecuente a la hipertrofia muscular inducida por el entrenamiento (3,5). El entrenamiento de fuerza explosiva, menos dependiente de cargas elevadas, y con énfasis en la tasa de producción de fuerza, parece estar asociado a una menor tasa de hipertrofia muscular (9,13). En consecuencia, hay razones para sospechar que las adaptaciones metabólicas en respuesta al entrenamiento de fuerza difieren, dependiendo de la carga del ejercicio y el concomitante grado de hipertrofia muscular.
Este estudio reporta y compara los efectos de 6 meses de entrenamiento de fuerza explosiva y entrenamiento de fuerza-resistencia, y un período subsecuente de 3 meses de desentrenamiento, sobre la actividad de varias enzimas del músculo esquelético. Los cambios relacionados con respecto a la hipertrofia muscular, fuerza muscular, tasa de producción de fuerza, tasa de relajación, y actividad electromiográfica han sido reportados por otros trabajos previos (8,9).
SUJETOS Y METODOS
Los sujetos de este estudio fueron 21 hombres físicamente activos. Todos estaban familiarizados con el entrenamiento con pesas no competitivo. Los sujetos fueron asignados al azar a un programa de entrenamiento de fuerza-resistencia (FR; n = l 1) ó de fuerza explosiva (FE; n =l 0). Las características físicas de los sujetos, y aquellas del grupo de control (n = 8), quienes mantuvieron un nivel de actividad física regular a lo largo del período experimental, son descriptas en la Tabla 1.
Un entrenamiento progresivo fue realizado 3 veces/sem como mínimo y continuado por 6 meses, seguidos de 3 meses de desentrenamiento. El grupo de FR ejecutó sentadillas (1-10 rep/serie, 18-30 rep/ sesión) utilizando una barra con pesas con cargas correspondientes a170 %-100 % de 1 RM (Repetición Máxima). Adicionalmente, ejecutaron 3-5 repeticiones excéntricas «súper pesadas» por sesión durante los meses 3, 5 y 6. El grupo FE ejecutó varios ejercicios de sentadillas (100-200 saltos/sesión) con un esfuerzo máximo, e intentando una mejoría en la tasa de producción de fuerza; por ej.: l) salto vertical en contramovimiento con una barra (10%-60% 1 RM); 2) cinco saltos de parado; 3) cinco saltos con vallas; y 4) saltos con caída seguidos de salto-rebote. Adicionalmente, ambos grupos experimentales ejecutaron ejercicios para músculos del tronco, brazos y piernas utilizando pesas (60 %-80 % 1 RM) 3 veces/ semana. Durante el período de desentrenamiento, el entrenamiento supervisado finalizó, pero los sujetos mantuvieron sus actividades normales diarias (para detalles adicionales, ver refs. 8,9).
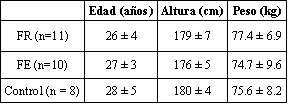
Tabla 1. Características físicas de los grupos de sujetos; FR (entrenamiento
de fuerza-resistencia); FE (entrenamiento de fuerza explosiva); y grupo control.
Los valores son medias ± SD.
Las biopsias musculares percutáneas fueron obtenidas del vasto lateral, en reposo (1). Se utilizaron tinciones histoquímicas para ATPasa miofibrilar (19), para clasificar las fibras como fibras rápidas (FT) ó fibras lentas (ST). Las FT, ST y medias de los tamaños de las fibras se calcularon en un corte seccional, por un método computarizado previamente descripto (27). También se calculó el área relativa ocupada por las fibras FT (% área FT).
Las muestras de tejido para análisis enzimático fueron congeladas en nitrógeno líquido y almacenadas a 80° C para análisis subsecuente. Luego del secado en frío, los paquetes de fragmentos de fibras musculares fueron pesados en una balanza Cahn y subsecuentemente analizados fluorométricamente (15) para determinar la actividad de la hexokinasa (HK; E.C.2.7.1.1), de la ATPasa estimulada por Mg++ (E.C.3.6.1.4), de lacitrato-sintetasa (CS; E.C.4.1.3.7), de la fosfofructokinasa (PFK; E.C.2.7. 1.11), de la lactato-dehidrogenasa (LDH; 1.1.1.27), de la miokinasa (MK: 2.7.4.3), y de la creatin-kinasa (CK; E.C.2.7.3.2).
Se calcularon las medias, desvíos standards y errores standards de las medias, con métodos estadísticos ordinarios. Las diferencias intra-grupos entre valores medios para variables antropométricas e histoquímicas fueron testeadas en búsqueda de significancia, a través del test « t » de Student. Los cambios en la actividad enzimática fueron evaluados por la aplicación de análisis de variancia (ANOVA), utilizando un test de rango «estudiantizado» (11).
RESULTADOS
Los resultados acerca del análisis histoquímico fueron reportados en nuestros trabajos previos, acordando con la mayoría de los parámetros examinados (8, 9). Debido a que algunos de los hallazgos son relevantes en cuando a cambios de actividad enzimática, las adaptaciones hipertróficas son brevemente repetidas. El área de fibras FT se incrementó (P <0.001) en 29 % y 13 % en FR y FE, respectivamente. El área de fibras ST se incrementó algo en el material experimental total (4 %; P <0.05) en respuesta al entrenamiento. Sin embargo, ninguno de los grupos mostraron área de fibras ST incrementada. La media de área por fibras (incluyendo FT y ST) se incrementó en 16 % (P <0.001) en FR, y 9 % (NS) en FE. El área FT relativa se incrementó (P <0.05) de 44 % a 53 % en FR; no ocurrieron cambios en FE. A continuación del período de desentrenamiento, el área de fibras FT siguió siendo mayor que antes del entrenamiento (P <0.05) en FR, pero no en FE (Tabla 2). Luego del período de desentrenamiento, el área ST y la media de área de fibra no difirió de los valores de pre-entrenamiento. Los cambios en las actividades enzimáticas musculares son detalladas en la Tabla 3.
Hexokinasa
La actividad de HK decreció (P <0.001) consecuente al entrenamiento. Este cambio ocurrió en FR (P <0.01) y FE (P <0.01), cuando ambos grupos fueron tratados separadamente, y a su vez, esta modificación persistió luego del desentrenamiento.
ATPasa
La ATPasa decreció (P <0.001) durante el entrenamiento, efecto que fue negativo cuando los grupos fueron tratados separadamente.
Citrato-Sintetasa
La actividad de CS decreció (P <0.05) durante el entrenamiento. No se observaron cambios en cualquiera de los grupos.
Fosfofructokinasa
La actividad de PFK decreció (P <0.01) subsecuente al entrenamiento. Se informa de una tendencia hacia una reducción de la actividad en ambos grupos sujetos al entrenamiento.
Lactato-dehidrogenasa
La actividad de LDH no fue afectada por el entrenamiento.
Miokinasa
La actividad de MK decreció (P <0.05) a continuación del período de entrenamiento, efecto que fue aparente en FR (P <0.05), pero no en FE.
Creatin-Kinasa
CK se redujo (PL 0.05) después del entrenamiento, un efecto que fue aparente en FR (P<0.05), pero no en FX.
DISCUSION
Este estudio demostró que el entrenamiento de fuerza a largo plazo está asociado a una hipertrofia muscular, como se refleja en varias mediciones antropométricas, y es sustentado por el hallazgo de un incremento en el tamaño de las fibras (ver ref. 8, 9). Estos resultados concuerdan con reportes previos acerca de los efectos de programas de entrenamiento de fuerza de duración más breve (6, 10, 13, 16). Nuestros resultados también sugieren que la carga utilizada, más que el volumen de entrenamiento o tasa de producción de fuerza, influye sobre la magnitud de la hipertrofia. Por ello, el incremento en tamaño de fibra muscular fue mayor en individuos sujetos a programas de entrenamiento de fuerza-resistencia, comparados con los de fuerza explosiva. En ambos tipos de entrenamiento, la hipertrofia estuvo primariamente limitada a fibras FT, lo que concuerda con estudios previos (6, 10, 13, 16, 26). Esto es compatible con la suposición de que la hipertrofia total de levantadores de pesas de alto nivel Olímpico o levantadores de potencia está principalmente atribuída a hipertrofia de fibras FT (22, 24). Como consecuencia de esta hipertrofia de fibras FT selectiva, el área muscular seccional relativa ocupada por fibras FT se incrementó en individuos que ejecutaron un entrenamiento de fuerza de alta resistencia. También parece que la tasa de atrofia muscular después del cese del entrenamiento, fue mayor o se equiparó con aquella hipertrofia inducida por el ejercicio.
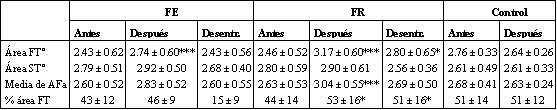
Tabla 2. Cambios en varios índices de área de fibras musculares durante el
entrenamiento y el desentrenamiento. Los valores son medias ± SD. Las
diferencias estadísticamente significativas con los valores de pre-entrenamiento
están expresadas por: (*) = P <0.05, (**) = P <0.01, y (***) = P <0.001. a =
áreas de fibra expresadas en unidades arbitrarias. AF = Área de fibras. Estos
datos fueron presentados previamente en forma gráfica (8,9).
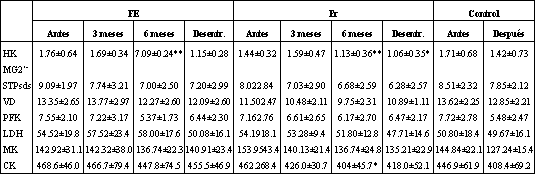
Tabla 3. Actividades enzimaticas antes, durante y después del entrenamiento
de fuerza en grupos sujetos con entrenamiento de fuerza explosiva (FX) y con
fuerza-resistencia (FR), y en un grupo control, antes y despues de 6 meses. Los
valores son medias± SD, y están expresados en pmol/g p.h/min. Los Cambios
estadisticamente significativos son expresados por: () = P <0.05, y (`) = P
<0.01.
El principal hallazgo del presente análisis fue, sin embargo, que 6 meses de trabajo de fuerza no incrementaron la actividad de enzimas que reflejan el metabolismo de energía glucolítico, oxidativo, o de contractilidad del músculo esquelético. Por lo tanto, la tasa de síntesis de esas enzimas parece ser más baja que aquella de las proteínas contráctiles durante el entrenamiento de fuerza. De hecho, la actividad de las enzimas hexokinasa, ATPasa, citrato-sintetasa, fosfofructokinasa y creatin-kinasa se redujo luego del entrenamiento, y ese efecto estuvo acompañado de un incremento de la media del tamaño de fibras, resultante principalmente por la hipertrofia de fibras FT. Los estudios han producido resultados divergentes respecto a los cambios enzimáticos, luego del entrenamiento de fuerza. Sin embargo, en aquellos estudios longitudinales que demuestran actividades enzimáticas incrementadas, la hipertrofia de fibras musculares fue pequeña o no ocurrió (4, 14, 26). Concordando con nuestros resultados, Houston y cols. (10) recientemente no pudo demostrar cambios enzimáticos luego de 8 semanas de entrenamiento de fuerza que produjeron un incremento del 20 % en el tamaño de fibras FT. En contraste, Costill con sus coautores (4) encontraron un incremento de la actividad de PFK, luego de 7 semanas de régimen de entrenamiento, que comprendía 10 series de 5 seg. de contracciones concéntricas, sin generar hipertrofia de fibra muscular. Por lo tanto, estudios de individuos que previamente no habían sido entrenados con fuerza, pueden haber provisto datos que sugieren adaptaciones metabólicas favorables, subsecuente a programas de entrenamiento de fuerza de corto plazo. Sin embargo, esos hallazgos deberían ser interpretados con cautela, especialmente si ellos se extrapolaron a poblaciones entrenadas, que realizaron entrenamiento de fuerza-resistencia de nivel elevado, a largo plazo.
Los presentes resultados pueden tener implicancias para atletas que participan en actividades que requieren alta capacidad aeróbica, y donde el entrenamiento de fuerza es utilizado como una ayuda para mejorar la velocidad y potencia. De ello se desprende que la hipertrofia muscular inducida por el entrenamiento puede atenuar la capacidad de generación de ATP a través de metabolismo aeróbico y anaeróbico. Dando sustento a este punto de vista están los hallazgos de una densidad de volumen mitocondrial reducida luego de un período de entrenamiento de fuerza (16), o de densidad mitocondrial y flujo capilar más bajos en atletas sujetos a muchos años de entrenamiento de fuerza (17, 23), y observaciones de capacidad oxidativa reducida en músculos hipertrofiados de atletas de fuerza y de potencia (3,5).
Por lo tanto, si el propósito del entrenamiento es principalmente el mejorar la capacidad aeróbica, el efecto hipertrófico del entrenamiento de fuerza puede impedir óptimas ganancias, reduciendo la tasa de síntesis de proteína mitocondrial. La demostración de una menor o comparable capacidad aeróbica para ejercicios del tren superior, o en cicloergómetro, en atletas entrenados con fuerza, comparados con deportistas no entrenados (a pesar de una masa muscular sustancialmente más grande en los primeros), concuerda con dicha presunción (2, 25). Estas nociones también implican especificidad de las adaptaciones a regímenes de entrenamiento de resistencia vs. fuerza/ potencia.
El detrimento en la actividad de la hexokinasa fue más pronunciado que en cualquier otra enzima. La hexokinasa cataliza la ruptura de la glucosa, reacción que es inhibida por la acumulación de glucosa-6-fosfato. Esta vía de energía parece ser de menor importancia durante el ejercicio elevado de fuerza. Asumiendo que los estímulos inducidos por el ejercicio no acrecientan la utilización de glucosa, el hallazgo de una actividad reducida de la hexokinasa podría, por lo tanto, ser explicado sobre la única base de un incremento en la síntesis de material contráctil, resultante en un «efecto de dilución». Por lo tanto, el detrimento en la actividad de la hexokinasa aparece paralelo al incrementato de la masa muscular, y se sugiere como efecto secundario de la hipertrofia muscular inducida por el ejercicio. El cambio en las actividades de otras enzimas puede estar relacionado al mismo mecanismo.
También parece que el incremento sustancial encontrado en el presente estudio en los índices de fuerza máxima y en la tasa de producción de fuerza, consecuente al entrenamiento de fuerza elevado, tai fue reportado en otros trabajos (8), no está relacionado al incremento de la actividad de la ATPasa miofibrilar. Por ello, es que la performance neuromuscular mejorada fue principalmente justificada por una función de adaptaciones neurales e hipertrofia selectiva de fibras FT.
En síntesis, 6 meses de entrenamiento elevado de fuerza-resistencia, o de fuerza explosiva, que resultaron en grandes mejorías de la performance neuromuscular, no indujeron ningún incremento en las actividades enzimáticas que reflejan el metabolismo aeróbico, o el metabolismo anaeróbico, o la contractilidad muscular. En cambio, la hipertrofia muscular inducida por el ejercicio de fuerza estuvo acompañada por la atenuación de las actividades de ciertas enzimas claves. Si bien el presente programa de entrenamiento no estuvo asociado con un incremento de las actividades, no puede excluirse que el entrenamiento de fuerza durante muchos años puede estar ligado a adaptaciones de enzimas contráctiles que podrían ser paralelas a la hipertrofia.
Referencias
1. Bergstrom I. (1962). Muscle electrolytes In man. Scand J Clin Lab Invest 14: (suppl) 68: 1-110
2. Colliander E.B., Tesch P.A (1960). Onset of blood lactate accumulation in strength trained athletes. (In manuscript)
3. Costill D.L., Daniels J., Evans W., Fink W., Kraehenbuhl G., Saltin B (1976). Skeletal muscle enzymes and fiber composition in male and female track athletes. J Appl Physiol 40: 149-151
4. Costill D.L., Coyle E.F., Fink W.F., Lesmes G.R. Witzmann F.A (1979). Adaptations In skeletal muscle following strength training. J Appl Physiol: REEP 46: 96-99
5. Gollnick P.D., Armstrong R.B., Saubert IV C.W., Pichl K., Saltin B (1972). Enzyme activity and fiber composition in skeletal muscle of untrained and trained men. J Appl Physiol 33: 312-319
6. Hakkinen K., Komi P.V., Tesch P.A (1981). Effect of combined concentric and eccentric strength training and detraining on forcetime, muscle fiber and metabolic characteristics of leg extensor muscles. Scand J Sports Sci 3: 50-58
7. Hakkinen K., Komi P.V (1983). Electromyographic changes during strength training and detraining. Med Sci Sports Exer 15: 455-460
8. Houston M.E., Froese E.A., Valeriote St. P., Green H.J., Ranney D.A (1983). Muscle performance, morphology and metabolic capacity during strength training and detraining: A one leg model. Eur J Appl Physiol 51: 25-35
9. Kirk R.E (1968). Experimental Design: Procedures for the Behavioral Sciences. Belmont, CA, Brooks-Cole
10. Komi P.V., Vlltasalo J., Rauramaa R., Vihko V (1978). Effect of isometric strength training on mechanical, electrical and metabolic aspects of muscle function. Eur JAppl Physiol 40: 45-55
11. Komi P.V., Suominen H., Helkkinen E., Karlsson J., Tesch P (1982). Effects of heavy resistance and explosive-type strength trainIng methods on mechanical, functional, and metabolic aspects of performance. In Komi P.V. (ed): Exercise and Sport Biology. int Series on Sport Sciences. Champaign, ILL, Human Kinetics Publishers, Vol 12, pp 90-102
12. Krotkiewski M., Aniansson A., Grimby G., Bjbrntorp P., Sjostrom L (1979). The effect of unilateral Isokinctic strength training on local adipose and muscle tissue morphology, thickness, and enzymes. Eur JAppl Physiol 42: 271-281
13. Lowry O.H., Passonneau J.V (1973). A Flexible System of Enzymatic Analysis. Academic Press
14. MacDougall J.D., Sale D.G., Moroz J.R., Elder C.C.B., Sutton J.R., Howald H (1979). Mitochondrial volume density in human skeletal muscle following heavy resistance training. Afed Sci Sports 11: 164-166
15. MacDougall J.D., Sale D.G., Elder C.C.B., Sutton J.R (1982). Muscle ultrastructural characteristics of elite powerlifters and bodybuilders. Eur J Appl Physiol 48: 117-126
16. Moritani T., DeVries H (1979). Neural factors versus hypertrophy in the time course of muscle strength gain. Am J Phys Med 58: 115-130
17. Padykula H.A., Herman E (1955). Factors affecting the activity of adenosine triphosphatase and other phosphatases as measured by histochemical techniques. JHistochem Cytochem 3: 161-195
18. Sale D.G., McComas A.J., MacDougall J.D., Upton A.R.M (1982). Neuromuscular adaptation in human thenar muscles following strength training and immobilization. J Appl Physiol: REEP 53: 419-424
19. Saltin B., Gollnick P.D (1983). Skeletal muscle adaptability: significance for metabolism and performance, in Peachy et al. (Eds): Handbook of Physiology - Skeletal Muscle. Baltimore, MD, Williams & Wilkins, pp 555-631
20. Staron R., Hikida R.S., Hagerman F.C., Dudley G.A., Murray T.F (1984). Human skeletal muscle fiber type adaptability to various workloads. J Histochem Cytochem 32: 146-152
21. Tesch P.A., Thorstensson A., Kaiser P (1984). Muscle capillary supply and fiber type characteristics in weight and power lifters. J Appl Physiol: REEP 56: 35-38
22. Tesch P.A., Karlsson J (1985). Muscle fiber types and size in trained and untrained muscles of elite athletes. J Appl Physiol 59: 1716-1720
23. Tesch P.A., Lindeberg S. (1984). Blood lactate accumulation during arm exercise in world class kayak paddlers and strength trained athletes. Eur J Appl Physiol 52: 441-455
24. Viitasalo J., Makinen P (1980). Comparison of five different methods for determination of muscle fibre areas. Presented at the Fifth Puijo Symposium, Kuopio, Finland, August 6-8