
Adaptaciones Musculares como Respuesta a tres Regímenes de Entrenamiento de la Fuerza: Especificidad de las Zonas de Entrenamiento de Repeticiones Máximas
William J Kraemer4, Gerson E Campos1, Thomas J Luecke1, Heather K Wendeln1, Kumika Toma1, Fredrick C Hagerman1, Thomas F Murray2, Kerry E Ragg3, Nicholas A Ratamess4 y Robert S Staron1
1Deparment of Biomedical Sciences, College of Osteophatic Medicine, Ohio University, Irvine Hall, rm 430, Athens, OH 45701, Estados Unidos.
2Com-Admin and Diagnostic Services, College of Health and Human Services, Ohio University, Athens, OH 45701, Estados Unidos.
3Student Health Service, Ohio University, Athens, OH 45701, Estados Unidos.
4Human Performance Laboratory, Department of Kinesiology, The University of Connecticut, Storrs, CT 06269, Estados Unidos.
Artículo publicado en el journal PubliCE, Volumen 0 del año 2002.
Publicado 17 de diciembre de 2003
Resumen
Palabras clave: músculo esquelético humano, tipos de fibras, histoquímica, capilarización, miosina de cadena pesada
INTRODUCCIÓN
El músculo esquelético humano es un tejido heterogéneo compuesto de tipos de fibras funcionalmente diversos (Staron, 1997). La mezcla de diferentes tipos de fibras permite al músculo llevar a cabo una variedad de demandas funcionales. Una característica única adicional del músculo esquelético es la habilidad de alterar su perfil fenotípico en respuesta a estímulos específicos (Pette y Staron, 2001). Para el entrenamiento de la fuerza, estas alteraciones usualmente contribuyen a incrementos significativos en la fuerza y el tamaño muscular (McDonagh y Davies, 1984; Tesch, 1988; Abernethy et al., 1994). Las investigaciones en esta área se han focalizado frecuentemente sobre varias combinaciones de series y repeticiones para optimizar estas adaptaciones específicas (Tan, 1999). Por ejemplo, un estudio anterior de Berger (1962) sugirió que 3 series de 4-8 repeticiones producen ganancias óptimas en la fuerza en comparación a varias otras combinaciones de series/repeticiones. Estos datos sugieren que las adaptaciones específicas de las fibras musculares están asociadas con las diferentes ganancias de fuerza.
Esta claro que la manipulación de varias variables de entrenamiento de fuerza (e.g., número de series, duración de los períodos de descanso entre series y ejercicios, intensidad, o carga) pueden estresar los músculos de maneras muy diferentes. Así, parecería haber relaciones específicas entre el estímulo de entrenamiento y la respuesta adaptativa. Tomando el entrenamiento de la fuerza hacia varios extremos. El trabajo clásico de DeLorme (1945) sugirió que un programa de entrenamiento de la fuerza que use bajas repeticiones/alta carga favorecía las adaptaciones de fuerza/potencia, mientras que el entrenamiento con altas repeticiones/baja carga incrementaba la resistencia muscular.
Anderson y Kearney (1982) evaluaron la hipótesis de DeLorme investigando los efectos de tres programas de entrenamiento de la fuerza muy diferentes sobre las adaptaciones de fuerza. 45 hombres de edad universitaria fueron asignados aleatoriamente a uno de tres grupos: alta carga/bajas repeticiones (3 series de 6-8 repeticiones máximas, RM), carga intermedia/intermedias repeticiones (2 series de 30-40 RM), y baja carga/altas repeticiones (1 serie de 100-150 RM). Luego de 9 semanas de entrenamiento durante 3 días/semana, el grupo de alta carga/bajas repeticiones presentó el mayor incremento en la fuerza máxima (una repetición máxima, 1 RM) y el más pobre incremento en la resistencia relativa (máximo número de repeticiones usando el 40% de 1 RM) en comparación con los otros dos grupos. Resultados similares fueron obtenidos en un estudio más reciente usando mujeres (Stone y Coulter, 1994). Este estudio, aunque fue diseñado después de Anderson y Kearney (1982), usó un protocolo de “resistencia” menos extremo para el grupo de baja carga/altas repeticiones. 45 mujeres de edad universitaria fueron asignadas a uno de tres grupos: alta carga/bajas repeticiones (3 series de 6-8 RM), resistencia intermedia/intermedias repeticiones (dos series de 15-20 RM), y baja carga/altas repeticiones (1 serie de 30-40 RM). Después de 9 semanas de entrenamiento durante 3 días/semana, el entrenamiento de alta carga/bajas repeticiones resultó en mayores ganancias de fuerza, mientras que el entrenamiento de baja carga/altas repeticiones produjo mayores ganancias de la resistencia muscular. A partir de estos estudios, ha sido hipotetizado un “continum de repeticiones de entrenamiento” (Anderson y Kearney, 1982) o un “continuum de repeticiones máximas” (Fleck y Kraemer, 1988), tal que el número de repeticiones permitidas por la carga va a resultar en adaptaciones al entrenamiento muy específicas.
Aunque varios estudios acerca del entrenamiento de la fuerza han sido publicados demostrando adaptaciones musculares específicas de fuerza y resistencia (e.g., Anderson y Kearney, 1982; Stone y Coulter, 1994), existe escasa información concerniente a las adaptaciones intramusculares específicas en respuesta a diferentes combinaciones de series y repeticiones. El propósito de la presente investigación fue comparar los efectos de tres programas diferentes de entrenamiento de la fuerza sobre las adaptaciones dentro del músculo vasto lateral. Con este objetivo, fueron elegidas rutinas en tres puntos diferentes dentro del continuum fuerza-resistencia teorizado, para investigar y comparar adaptaciones musculares específicas. El diseño del presente estudio se parece más de cerca a aquel de Stone y Coulter (1994), y utilizó un régimen de entrenamiento de fuerza más práctico sobre el objetivo de resistencia del continuum, en comparación a aquel estudio de Anderson y Kearney (1982).
MÉTODOS
32 sujetos sanos de sexo masculino (media±DS, edad 22.5±5.8 años, talla 178.3±7.2 cm, masa corporal 77.8±11.9 kg) se ofrecieron par participar en el presente estudio. Antes de su participación, todos los sujetos fueron informados acerca de los procedimientos, riesgos y beneficios, y firmaron un documento de informe de consentimiento aprobado por el Comité de Revisión Institucional de la Universidad de Ohio. Aunque los sujetos eran físicamente activos, todos fueron considerados desentrenados y no habían participado en un programa de ejercicio regular por al menos 6 meses antes del inicio del estudio. 27 varones sanos fueron divididos aleatoriamente en tres grupos de entrenamiento: un grupo de bajas repeticiones (LR, n=9; media±DS, edad 21.1±1.5 años, talla 179.8±6.5 cm), un grupo de repeticiones intermedias (IR, n=11; media±DS, edad 20.7±2.9 años, talla 179.6±7.4 cm) y un grupo de altas repeticiones (HR, n=7; media±DS, edad 20.4±3.5 años, talla 174.3±8.6 cm). 6 individuos adicionales sirvieron como sujetos controles que no realizaban ejercicio (Con). Un sujeto del grupo control comenzó a realizar entrenamientos de resistencia durante el transcurso del estudio y fue sacado del mismo, dejando un grupo control final de 5 sujetos (media±DS, edad 31.6±9.8 años, talla 178.1±5.5 cm).
Evaluaciones Antropométricas
Las mediciones antropométricas (masa corporal total, masa libre de grasa estimada, y porcentaje estimado de grasa corporal) fueron determinadas antes y después del período de 8 semanas de entrenamiento (Tabla 1). Las mediciones de pliegues cutáneos fueron obtenidas a partir de tres sitios (muslo anterior, pliegue axilar, y abdomen) antes de la extracción de las muestras de biopsias musculares y fueron usadas en la ecuación propuesta por Jackson y Pollock (1978) para los análisis de composición corporal (e.g., estimación del porcentaje de grasa corporal).
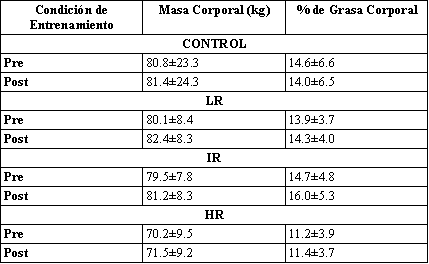
Tabla 1. Masa corporal total y porcentaje de grasa corporal estimado. Los
valores son presentados como media±DS. (LR, grupo de bajas repeticiones; IR,
grupo de repeticiones intermedias, HR, grupo de repeticiones altas; Pre, pre-entrenamiento;
Post, post-entrenamiento).
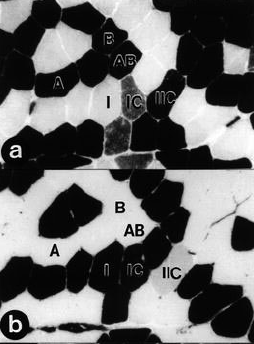
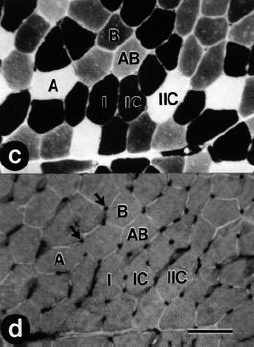
Figura 1. Secciones transversales en serie de muestras musculares tomadas
a partir de un sujeto control demostrando la delineación de los tipos de fibras
usando histoquímica de la adenosín trifosfatasa miofibrilar después de la
preincubación a un pH de 10.4 (a), 4.3 (b), y 4.6 (c), y aporte capilar usando
histoquímica de agglutinin lectina Ulex europaeus (d). Las flechas en el gráfico
d indican capilares. (1, fibras musculares tipo I; 1C, fibras musculares tipo
1C; IIC, fibras musculares tipo IIC; A, fibras musculares tipos IIA; AB, fibras
musculares tipos IIAB; B, fibras musculares tipo IIB). Barra =100 μm.
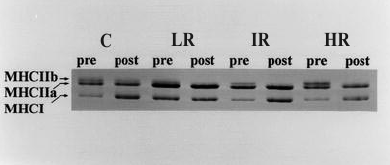
Figura 2. Análisis de miosina de cadena pesada (MHC) de las muestras de
las biopsias obtenidas a partir de un sujeto representativo en cada uno de los
cuatro grupos (C, control; LR, bajas repeticiones; IR, repeticiones intermedias;
HR, altas repeticiones) al comienzo (pre) y el final (post) del estudio. Notar
la disminución en la banda correspondiente a la MHCIIb desde la condición pre- a
post-entrenamiento para los sujetos de entrenamiento. (MHCIIb, miosina de cadena
pesada IIb; MHCIIa, miosina de cadena pesada IIa; MHCI, miosina de cadena pesada
I).
Máximo Consumo de Oxígeno
Un estudio previo de este laboratorio (Hagerman et al., 2000) demostró mejoras significativas en el máximo consumo de oxígeno (VO2 máx.) y el tiempo hasta el agotamiento luego de 16 semanas de entrenamiento de fuerza en hombres ancianos. De este modo, las capacidades aeróbicas fueron determinadas para todos los sujetos al inicio y al final del estudio. La evaluación fue administrada en un cicloergómetro Monark, usando un protocolo incremental que incremento la intensidad a intervalos regulares. Los sujetos comenzaron pedaleando a una frecuencia de 60 rpm a 60 W. La carga de trabajo fue progresivamente incrementada cada minuto a razón de 30 W. La evaluación finalizaba cuando el sujeto no podía continuar manteniendo la potencia requerida o se detenía voluntariamente debido al agotamiento. Una evaluación era considerada válida si uno de los siguientes criterios era observado: 1. era alcanzada la frecuencia cardiaca máxima predicha, 2. el consumo de oxígeno (VO2) se nivelaba o declinaba, o 3. era alcanzado un índice de intercambio respiratorio (R) mayor a 1.1 (ver Howley et al., 1995). El VO2 fue medido cada 20 s. La frecuencia cardiaca y la tasa de esfuerzo percibido fueron medidas en cada intensidad del ejercicio. El VO2 máx., la ventilación pulmonar (VE), dióxido de carbono espirado, y R fueron medidos usando espirometría de circuito abierto computarizada-semiautomática (Vacumed, Ventura, Calif., Estados Unidos) que incluyó un medidor de gases secos Parkinson-Cowan, y analizadores infrarrojos de dióxido de carbono, y analizadores paramagnéticos de oxígeno. La frecuencia cardiaca fue monitoreada cada minuto usando transmisores y receptores Polar CIC. El tiempo hasta el agotamiento fue registrado como un múltiplo de 20 s a partir del inicio de la evaluación. Inmediatamente después de la evaluación de ciclismo, los sujetos permanecieron sentados para tener un período de recuperación de 5 min antes de que fuera obtenida una muestra de sangre para los análisis del lactato sanguíneo total.
Evaluaciones de Fuerza Máxima y Resistencia Muscular
Todos los sujetos (incluyendo a los controles) participaron en un programa de
orientación de 1 semana para la familiarización con los equipos y con los
ejercicios (Dudley et al., 1991). Una técnica de levantamiento apropiada fue
mostrada y los sujetos practicaron cada uno de los tres ejercicios para el tren
inferior (prensa de piernas, sentadilla, extensión de rodilla). La fuerza máxima
dinámica (1 RM) y la resistencia muscular local (máximo número de repeticiones
realizadas con el 60% de 1 RM) fueron evaluadas para cada uno de los ejercicios
al inicio y al final del estudio. Debido a la naturaleza exhaustiva de la
evaluación de resistencia, la evaluación de fuerza máxima fue siempre realizada
primero. Después de la entrada en calor, la carga fue establecida al 90% de la 1
RM predicha, y fue incrementada después de cada levantamiento exitoso hasta el
fallo (Staron et al., 1990). Los períodos de descanso (aproximadamente 4-5 min)
fueron asignados entre cada intento para asegurar la recuperación. Una
evaluación era considerada válida si el sujeto usaba la forma apropiada y
completaba el levantamiento completo de una forma controlada sin ayuda. Una vez
que la 1 RM era determinada, era calculado el 60% de este valor para la
evaluación de resistencia muscular local. Luego de un período de recuperación
suficiente (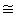 4-5 min), los sujetos realizaban tantas repeticiones como fuera
posible con el 60% de 1 RM hasta el fallo.
4-5 min), los sujetos realizaban tantas repeticiones como fuera
posible con el 60% de 1 RM hasta el fallo.
Protocolos de Entrenamiento de Fuerza
Los sujetos que entrenaban participaron en un programa de 8 semanas de alta intensidad para el tren inferior. El entrenamiento fue realizado 2 días/semana por las primeras 4 semanas y 3 días/semana por las 4 semanas finales. Los sujetos que entrenaban usaron uno de los tres regímenes diferentes. Los programas de entrenamiento fueron adaptados a partir de varios estudios previos (Anderson y Kearney, 1982; Jackson et al., 1990; Stone y Coulter, 1994) y fueron diseñados para ser aproximadamente iguales en volumen (carga x repeticiones x series) con los períodos de descanso entre series y ejercicios ajustados de acuerdo a el continuum fuerza-resistencia (Fleck y Kraemer, 1997). De este modo, aquellos individuos que trabajaban en el extremo de resistencia del continuum realizaban menos series y tenían períodos de descanso más cortos en comparación con los otros grupos de entrenamiento. Los ejercicios fueron realizados en el orden fijo de press de piernas, sentadilla, y extensión de rodilla. Después de entrar en calor el grupo LR realizaba cuatro series de 3-5 repeticiones máximas (3-5 RM) con 3 min de descanso entre series y ejercicios, el grupo IR realizaba 3 series de 9-11 RM con dos minutos de descanso, y el grupo HR realizaba 2 series de 20-28 RM con 1 min de descanso. Durante el estudio, la resistencia fue progresivamente incrementada para mantener este rango de repeticiones por serie. Para cada serie, los sujetos realizaron las repeticiones hasta el agotamiento. Si los sujetos realizaban repeticiones más allá de la zona de entrenamiento prescripta, el peso era suficientemente incrementado para llevar el número de repeticiones de nuevo dentro de la zona de entrenamiento de RM. Todos los sujetos fueron supervisados y alentados verbalmente durante cada serie. Las frecuencias cardiacas máximas fueron medidas durante el entrenamiento en las semanas 2 y 7 para comparar el estrés cardiorrespiratorio de cada grupo de entrenamiento. Las frecuencias cardiacas de entrenamiento fueron calculadas como un porcentaje de la frecuencia máxima obtenida durante el VO2 máx. final en el cicloergómetro pre- y post-evaluación. Los valores de la semana 2 fueron calculados como un porcentaje de la frecuencia cardiaca máxima obtenida a partir de las evaluaciones máximas pre-entrenamiento y aquellos de la semana 7 como un porcentaje de la evaluación máxima post-entrenamiento. Los entrenamientos comenzaron y finalizaron con 10-15 min de ejercicios calinestésicos, estiramientos, y ciclismo de baja intensidad.
Muestreo de Biopsia Muscular
Las muestras de biopsia muscular (80-160 mg) fueron extraídas a partir de una región superficial (3-4 cm de profundidad) del músculo vasto lateral (aproximadamente la mitad de la aguja) usando la técnica de biopsia con aguja percutanea (Bergström, 1962). Las muestras musculares fueron sacadas de la aguja, dispuestas en goma de tragacanto, e inmediatamente congeladas en isopentano enfriado por medio de nitrógeno líquido a -159 ºC, y almacenadas a –74 ºC hasta que los análisis posteriores pudieran ser realizados. Las muestras de biopsias fueron obtenidas al inicio y al final del estudio. Debido a posibles variaciones en la distribución del tipo de fibras desde la superficie hasta la profundidad y desde un punto de referencia proximal hasta otro distal (Blomstrand y Ekblom, 1982), fueron hechos intentos para extraer las muestras de tejidos pre- y post-entrenamiento, dentro de un área de músculo pequeña, usando la cicatriz pre-biopsia y las marcas de profundidad en la aguja. Así, fueron hechas incisiones sucesivas apartadas en aproximadamente 0.5 cm. Los sitios donde se realizaron las biopsias pre- y post-entrenamiento estuvieron suficientemente distanciados de modo que la inserción de la primera aguja de biopsia y la extracción de tejido no afectara el área de la segunda biopsia. Para asegurar tamaños de muestra adecuados, fueron obtenidos grandes especimenes de biopsias usando un método double-chop (Staron et al., 1990, 1991, 1994) combinado con la succión (Evans et al., 1982).
Determinaciones de Tipos de Fibras y Área de Sección Transversal
Los especimenes de biopsias congelados fueron descongelados, llevados a -24ºC y seccionados en fascículos (de 12 μm de ancho) para los análisis histoquímicos. Para determinar la composición de los tipos de fibras musculares, fue realizada histoquímica de adenosín trifosfatasa miofibrilar (mATPasa) usando valores de pH de preincubación de 4.3, 4.6, y 10.4 (Guth y Samaha, 1969; Brooke y Kaiser, 1970). Fueron distinguidos 6 tipos de fibras (I, IC, IIC, IIA, IIAB y IIB) en base a la intensidad de la tinción (Figura 1 A-C). A los tipos de fibras tipo IIAB y IIB se los ha calificado más recientemente como IIAX y IIX, respectivamente (Smerdu et al., 1994; Ennion et al., 1995). La secciones transversales de los especimenes de biopsias pre- y post-entrenamiento de un individuo fueron colocados en un coverslip de vidrio de modo que las mismas pudieran ser simultáneamente analizadas para la actividad de la mATPasa. Fue hecho un fotomontaje compuesto de cada preparación de mATPasa después de la preincubación a un pH de 4.6 usando micrografías Polaroid (x 56 de aumento). Las mismas fueron usadas en combinación con las otras preparaciones de mATPasa para determinar los porcentajes de tipos de fibras y el número total de fibras en cada muestra de biopsia. El área de sección transversal fue determinada sobre al menos 50 fibras por tipo de fibra muscular principal (I, IIA y IIB) por muestra de biopsia usando un software de imageria NIH (versión 1.55).
Análisis de la Miosina de Cadena Pesada (MHC)
Los análisis MHC fueron realizados sobre las muestras de biopsias usando técnicas electroforéticas de dodecil sulfato de sodio (SDS)-poliacrilamida (Figura 2). El protocolo para el análisis de los especimenes estuvo basado en procedimientos de Perrie y Bumford (1986) con modificaciones usadas para el análisis de fibras aisladas (Staron 1991; Staron y Hikida, 1992). Brevemente, de cuatro a seis secciones transversales (20 μm de ancho) de cada muestra de biopsia fueron colocados dentro de 5 ml de un buffer que contenía 10% (peso/volumen) de glicerol, 5% (vol/vol) de 2-mercaptoetanol, y 2-3 % (peso/volumen) de SDS en 62.5 mM de un buffer HCl tris (hidroximetil) aminometano (pH 6.8) y fueron calentadas durante 10 min a 60 ºC. Pequeñas cantidades de extracto (3-5 μl) fueron cargadas en gels SDS-policacrilamida de un gradiente de 4-8% con gels al 4% (Bär y Pette, 1998), fueron mantenidos durante toda la noche (19-21 h) a 120 V, y teñidas con Coomassie Blue. Las isoformas de MHC fueron identificadas de acuerdo a su masa molecular aparente en comparación con aquellas proteínas marcadoras y patrones de migración a partir de los análisis de fibras aisladas. El contenido relativo de la isoforma de MHC fue subsecuentemente determinado usando un densitómetro laser.
Evaluación de los Capilares
Los capilares fueron identificados sobre secciones transversales consecutivas a aquellas usadas para la determinación del tipo de fibras por medio de histoquímica de agglutinin I (UEA-I) lectina Ulex europaens, de acuerdo a los procedimientos de Holthöfer et al. (1982) (Figura 1 D). UEA-1 es un marcador sensible y confiable para el endotelio. Los datos de los capilares fueron recolectados a partir de al menos 50 fibras por tipo de fibra principal (I, IIA, IIB) por muestra de biopsia. Las fibras que se encontraban sobre el borde del fascículo muscular no fueron incluidas en los análisis. También, los capilares en el borde de un área de muestra fueron tomados juntos y divididos por dos para corregir para la división de las fibras, de acuerdo a Plyley y Groom (1975). En la presente investigación fueron usadas varias mediciones de los capilares musculares. El número de capilares por unidad de área (densidad capilar, CD=capilares/mm2) fue medido para dar una indicación del número de capilares presentes en un área estándar. El número de capilares por área de tipo de fibra (CFTA) fue determinado para evaluar las diferencias relativas en el aporte capilar para las fibras individuales. Finalmente, el número de capilares por tipo de fibra (CFT) fue reportado para demostrar una medición absoluta del aporte capilar para cada tipo de fibra.
Análisis Estadísticos
Fueron usados estadígrafos descriptivos para derivar medias y desvíos estándar (DS) de todas las variables, y los datos fueron presentados en la forma de media±DS. Los análisis estadísticos para cada variable dependiente fueron llevados a cabo usando análisis de varianza (ANOVA) a dos vías separados con mediciones repetidas (diseño 3 x 2: 3 grupos en dos puntos de tiempo). Cuando era alcanzado algún valor F significativo, eran llevadas a cabo comparaciones post-hoc por medio de un test de menor diferencia significativa de Fisher. Usando el software nQuery Advisor (Statistical Solutions, Saugus, Mass., Estados Unidos), el poder estadístico para el n usado varió entre 0.76 a 0.87. Las diferencias fueron consideradas significativas a una p=0.05.
RESULTADOS
Datos Antropométricos
No fueron detectadas diferencias significativas entre o dentro de los grupos para las mediciones antropométricas (Tabla 1).
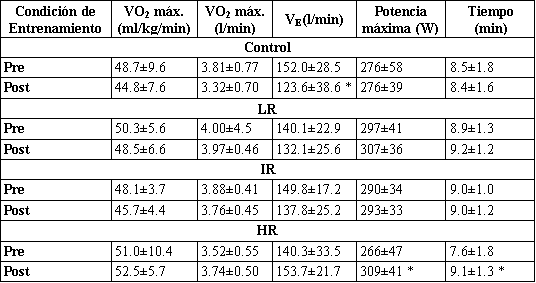
Tabla 2. Datos cardiorrespiratorios obtenidos a partir de las evaluaciones de
resistencia pre- y post-entrenamiento. Los valores dados están presentados como
media±DS. VO2, consumo de oxígeno; VO2 máx., máximo consumo de oxígeno; VE,
ventilación por minuto; Potencia máxima, máxima potencia aeróbica; t, tiempo
hasta el agotamiento. * Diferencias significativas con respecto al valor pre-entrenamiento.
Mediciones Cardiorrespiratorias
El VO2 máx., la VE, el tiempo hasta el agotamiento y la potencia aeróbica fueron monitoreados al inicio y al final del estudio (Tabla 2). Todos los sujetos completaron evaluaciones de VO2 máx. válidas (ver Métodos). Las frecuencias cardiacas promedio fueron 188.2±16.1 y 189±9.6 lat./min para las evaluaciones pre- y post-entrenamiento, respectivamente. Las mediciones de lactato no se diferenciaron entre los grupos, y promediaron 12.4±1.7 y 11.6±2.5 mmol/l para las evaluaciones pre- y post-entrenamiento, respectivamente. El VO2 máx. y la VE no cambiaron al final del estudio para todos los grupos de entrenamiento. El grupo HR fue el único grupo de entrenamiento que mostró un incremento significativo en el tiempo hasta el agotamiento (desde 7.6±1.8 hasta 9.1±1.3 min) y en la potencia aeróbica máxima (desde 265±47 hasta 308±41 W) en la evaluación de resistencia en cicloergómetro. Como se esperaba, el grupo Con no demostró cambios en varios parámetros cardiorrespiratorios con la excepción de una disminución significativa en la VE al final del estudio (Tabla 2).
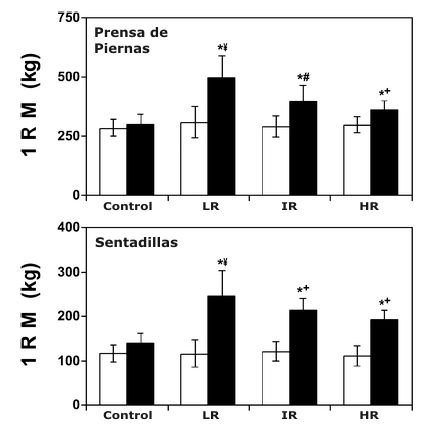
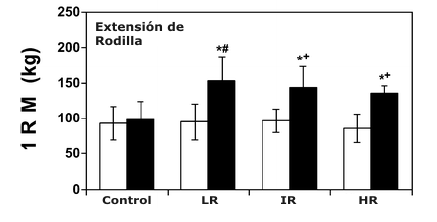
Figura 3. Gráficos de barras comparando los valores de fuerza máxima (1
repetición máxima, 1 RM) (media±DS) para los tres ejercicios del tren inferior
pre- (□) y post-entrenamiento (■). C, grupo control; LR, grupo de bajas
repeticiones; IR, grupo de repeticiones intermedias; HR, grupo de altas
repeticiones. * Significativamente mayor que el valor pre-entrenamiento
correspondiente; ¥, significativamente mayor que todos los valores
post-entrenamiento correspondientes; # significativamente mayor que los valores
post-entrenamiento correspondientes de los grupos altas repeticiones y control;
+ Significativamente mayor que los valores post-entrenamiento correspondientes
del grupo control.
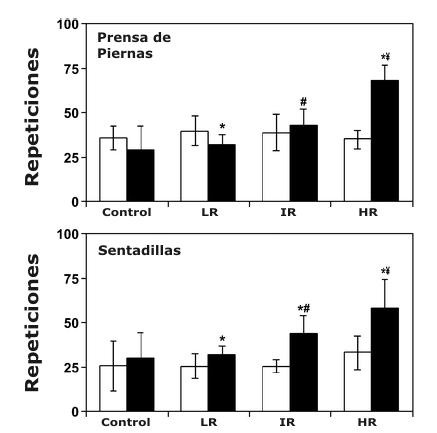
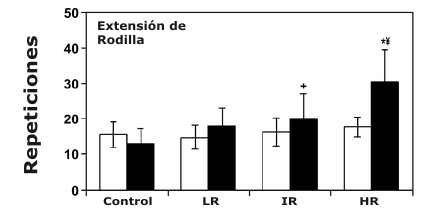
Figura 4. Gráficos de barras comparando el máximo número de repeticiones
usando el 60% de 1 RM (media±DS) para los tres ejercicios del tren inferior pre-
(□) y post-entrenamiento (■). *Significativamente mayor que el valor pre-entrenamiento
correspondiente; ¥, significativamente mayor que todos los valores
post-entrenamiento correspondientes; # significativamente mayor que los valores
post-entrenamiento correspondientes de los grupos bajas repeticiones y control;
+ Significativamente mayor que los valores post-entrenamiento correspondientes
del grupo control.
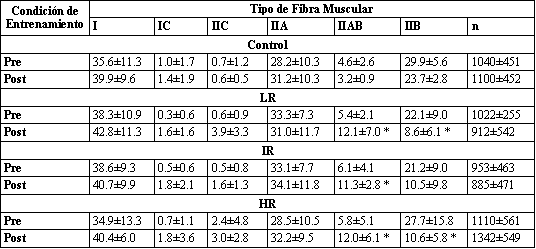
Tabla 3. Porcentajes de los tipos de fibras musculares determinados usando
métodos de histoquímica de adenosín trifosfatasa miofibrilar. Los valores están
presentados como media±DS. n, número promedio de fibras por muestra tomada
mediante biopsia. * Significativamente diferente con respecto al valor pre-entrenamiento
correspondiente.
Volumen de Entrenamiento y Estrés Cardiorrespiratorio
El volumen promedio de entrenamiento realizado (carga x repeticiones x series) fue calculado para cada grupo de entrenamiento cada semana. No fueron encontradas diferencias significativas en el volumen entre el entrenamiento de los grupos. Los volúmenes tuvieron una tendencia a aumentar ligeramente para los tres grupos para cada ejercicio a través de toda la duración del período de entrenamiento. Sin embargo, no ocurrieron diferencias significativas en el volumen través del tiempo. Del mismo modo, no fueron encontradas diferencias entre los grupos cuando se compararon las frecuencias cardiacas obtenidas durante el entrenamiento en las semanas 2 y 7. Las frecuencias cardiacas de entrenamiento promedio para la semana 2 fueron 87%, 86%, y 93%, y para la semana 7 fueron de 88%, 87% y 91% para los grupos LR, IR, y HR, respectivamente.
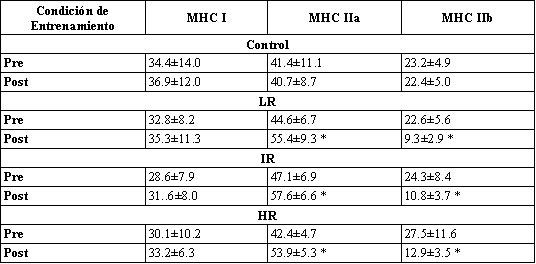
Tabla 4. Porcentajes relativos de las isoformas de miosina de cadena pesada a
partir muestras musculares homogeneizadas determinadas usando electroforesis en
gel de dodecil sulfato de sodio- poliacrilamida. Los valores son presentados
como media±DS. MHCI, miosina de cadena pesada I; MHCIIa, miosina de cadena
pesada IIa; MHCIIb, miosina de cadena pesada IIb. * Significativamente diferente
de los valores pre-entrenamiento.
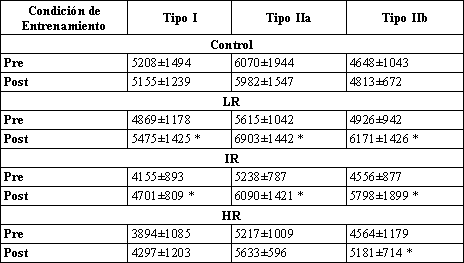
Tabla 5. Área de sección transversal (m2) de los tres tipos principales de
fibras musculares. Los datos son presentados como media±DS. * Significativamente
diferente con respecto a los valores pre-entrenamiento correspondientes.
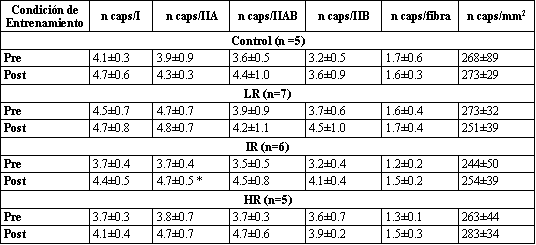
Tabla 6. Aporte capilar a los músculos determinado usando histoquímica de agglutinin lectina Ulex europaeus. Los valores son presentados como media±DS. n
caps/I, número de capilares por fibra tipo I; n caps/IIA, número de capilares
por fibra tipo IIA; n caps/IIAB, número de capilares por fibra tipo IIAB; n caps/IIB,
número de capilares por fibra IIB; n caps/fibra, número de capilares por fibra;
n caps/mm2, número de capilares por milímetro cuadrado. * Significativamente
diferente con respecto a los valores pre-entrenamiento correspondientes.
Fuerza Dinámica Máxima y Resistencia Muscular
Todos los grupos de entrenamiento mostraron incrementos significativos en la fuerza dinámica máxima (1 RM) para los tres ejercicios en comparación con sus respectivos valores pre-entrenamiento (Figura 3). Aunque no fue presentado, esto también fue cierto para la fuerza dinámica máxima relativa a la masa corporal total y magra. No fueron demostrados cambios significativos en la 1 RM desde las condiciones pre- a post-entrenamiento para el grupo Con (Figura 3). En una comparación entre los grupos, los incrementos relativos y absolutos en la fuerza dinámica máxima para los ejercicios de prensa de piernas y sentadilla fueron significativamente mayores para el LR en comparación con los otros grupos (Figura 3). Para la extensión de piernas, el valor de 1 RM post-entrenamiento para el grupo LR fue significativamente mayor que para los grupos HR y Con (Figura 3).
En comparación con los resultados obtenidos para la fuerza dinámica máxima, lo inverso también fue cierto para la evaluación de la resistencia muscular local. El grupo de HR realizó significativamente más repeticiones usando el 60% de 1 RM después del entrenamiento para todos los ejercicios, y estos valores post-entrenamiento fueron mayores que todos los valores de los grupos correspondientes (Figura 4). Aunque todos los grupos de entrenamiento incrementaron significativamente el número de repeticiones usando el 60% de 1 RM en el ejercicio de sentadilla después del entrenamiento, ni el grupo IR ni el grupo LR demostraron mejoras significativas al 60% de 1 RM después del entrenamiento para los ejercicios de prensa de piernas y extensión de piernas (Figura 4). Efectivamente, el grupo LR realizó significativamente menos repeticiones para la prensa de piernas después del entrenamiento (Figura 4). No fueron encontrados cambios significativos en la resistencia muscular total para el grupo Con (Figura 4).
Distribución del Tipo de Fibras y Contenido de MHC
Para todos los grupos de entrenamiento, los porcentajes de fibras tipo IIB disminuyeron, con un incremento concomitante en el porcentaje de fibras clasificadas como tipo IIAB (Tabla 3). No fueron encontrados cambios en la distribución del tipo de fibras para el grupo Con. Estos datos fueron apoyados por los cambios en los porcentajes relativos de isoformas de MHC. Los especimenes de biopsias de los tres grupos de entrenamiento mostraron una disminución significativa en la MHCIIb y un incremento concomitante en la MHCIIa (Figura 2, Tabla 4). No fue encontrado ningún cambio en el contenido MHC para el grupo Con.
Área de Sección Transversal
Fue observado un efecto de hipertrofia después del entrenamiento de resistencia solo para los grupos IR y LR (Tabla 5). Para estos dos grupos, las áreas de sección transversal para todos los principales tipos de fibras (I, IIA, y IIB) fueron significativamente mayores después del entrenamiento. El programa de entrenamiento de fuerza progresivo de 8 semanas de duración causó un incremento en las áreas de sección transversal de los principales tipos de fibras de aproximadamente 12.5% para las fibras tipos I, 19.5% para las fibras tipos IIA, y 26% para las fibras tipo IIB, para ambos grupos IR y LR. No fueron encontrados cambios en el área de los grupos HR y Con (Tabla 5). Sin embargo, hubo una tendencia de incremento en el área de sección transversal de las fibras tipo IIB después del entrenamiento en el grupo HR (p=0.13).
Capilarización
Comparando los valores pre- a post-entrenamiento para todos los grupos, no fueron encontradas diferencias significativas en CD o CF (Tabla 6). Además, no ocurrieron cambios significativos en CFTA para ningún grupo (datos no presentados). Con la excepción de un incremento significativo después del entrenamiento para CFT para las fibras tipo IIA para el grupo IR, no ocurrieron cambios significativos en CFT para ninguno de los tipos de fibras (I, IIA, o IIB; Tabla 6).
DISCUSIÓN
Adaptaciones de la Fuerza
Aunque algunos estudios de entrenamiento de la fuerza han puesto en tela de juicio la teoría de DeLorme (1945) acerca de un continuum fuerza-resistencia (DeLateur et al., 1968; Clark y Strull, 1970; Stull y Clark, 1970), la mayoría de las investigaciones en esta área apoyan la idea de una especificidad en la tarea relacionada a combinaciones de series/repeticiones específicas (e.g., Stull y Clark, 1970; Anderson y Kearney, 1982). Los resultados de la presente investigación apoyan los hallazgos de estos primeros estudios sobre las adaptaciones de la fuerza. Aunque la fuerza dinámica máxima mejoró significativamente en los tres grupos, el grupo LR es el que más la incrementó. Por ejemplo, los incrementos de la fuerza dinámica máxima en el ejercicio de prensa de piernas alcanzaron el 61% para el grupo LR en comparación con el 36% del IR, 32% del HR, y 6% del Con (Figura 3). De otro modo, la resistencia muscular local en el ejercicio de prensa de piernas se incremento más en el HR en comparación con los otros grupos: 94% de incremento en el grupo HR, 10% de incrementos en el IR, -20% en el grupo LR, y –19 % para el grupo Con (Figura 4).
Adaptaciones Cardiorrespiratorias
Aunque el entrenamiento de fuerza tradicional implica resistencias altas combinadas con bajos números de repeticiones, los regímenes de baja resistencia/altas repeticiones deben ser todavía considerados como formas de entrenamiento e la fuerza. Aún trabajar en el “extremo” de resistencia del continuum fuerza-resistencia (realizando tantas como 150 repeticiones) significa usualmente realizar contracciones submáximas repetidas por al menos 5 minutos/serie (e.g., Stull y Clark, 1970; Anderson y Kearney, 1982). Sin embargo, han sido reportados incrementos en la resistencia de corta duración luego de un programa de entrenamiento de fuerza. Después de 10 semanas de un programa de entrenamiento de resistencia de alta intensidad, Hickson et al. (1980) encontraron incrementos significativos en el tiempo hasta el agotamiento en ciclismo y carrera con ningún incremento significativo en el VO2 máx. Además, Macinik et al. (1991) reportaron que 12 semanas de entrenamiento de la fuerza en circuito mejoraron el rendimiento de resistencia en ciclismo independientemente de los cambios en el VO2 máx. Han sido reportados hallazgos similares cuando al entrenamiento de fuerza se le suma un programa de entrenamiento de resistencia aeróbico en individuos jóvenes (Hickson et al., 1998; Paavolainen et al., 1999), así como en hombres (Grimby et al., 1992) y mujeres (Feketich et al., 1998) ancianos. Los resultados del presente estudio apoyan estos hallazgos. El grupo HR mejoro significativamente su rendimiento en ciclismo (potencia aeróbica máxima y tiempo hasta el agotamiento) sin cambios en el VO2 máx. (Tabla 2). Aunque esto parece contradecir el principio básico de la especificidad del entrenamiento, un incremento en la capacidad de trabajo de larga duración también requiere fuerza muscular y potencia anaeróbica (Tanaka y Swensen, 1998). Además, ha sido demostrado que el entrenamiento de resistencia mejora la economía de carrera (Johnston et al., 1995; Paavolainen et al., 1999) y puede así, también mejorar la economía en ciclismo. Tales incrementos en el rendimiento de resistencia luego de un programa de entrenamiento de la fuerza pueden estar relacionados a incrementos en el umbral del lactato y a la fuerza de miembros inferiores (Marcinik et al., 1991), y de este modo, pueden no necesariamente estar relacionados con incrementos en la capacidad aeróbica.
Tipos de Fibra Musculares y Área Transversal
Aunque las adaptaciones de fuerza y rendimiento ante programas de entrenamiento de la fuerza variados están bastante bien documentadas, existe escasa información con respecto a las adaptaciones neuromusculares. Nuestra esperanza era poder generar una idea acerca de que puede estar pasando dentro del músculo (i.e., cambios en el tamaño de las fibras, tipo de fibra, y capilaridad) que pudiera contribuir potencialmente a estas diferencias documentadas en la fuerza máxima, resistencia muscular local y rendimiento.
Muy pocos estudio han intentado documentar cambios musculares luego de diferentes modos de entrenamiento de fuerza. Algunos estudios han usado métodos indirectos para medir los cambios en el área transversal (imageria de resonancia magnética y exploración con ultrasonido) y han reportado áreas y adaptaciones de la fuerza similares en hombres jóvenes (Chestnut y Docherty, 1999), mujeres jóvenes (Hisaeda et al., 1996), y mujeres posmenopáusicas (Bemben et al., 2000) sujetos a diferentes protocolos de entrenamiento de series/repeticiones máximas. Para nuestro conocimiento, solo dos estudios han investigado los efectos de diferentes tipos de programas de entrenamiento de la fuerza sobre el músculo esquelético, utilizando muestreo de biopsia muscular (Jackson et al., 1990; Taaffe et al., 1996), y ninguno de los mismos estableció específicamente el “continuum fuerza-resistencia”.
Las ventajas del presente estudio en comparación con trabajos previos en esta área incluyen un grupo control que no realizo ejercicio, un rango completo de tipos de fibras histoquímicas, contenido relativo de MHC para validar los datos histoquímicos, área de sección transversal determinada a partir de 50 fibras por tipo de fibra muscular principal, pesos libres para el entrenamiento y las evaluaciones, y la inclusión de varios parámetros cardiorrespiratorios. Además, en el presente estudio nosotros elegimos tres protocolos de entrenamiento de la fuerza distintos, pero prácticos para focalizarnos específicamente sobre las adaptaciones musculares a diferentes puntos a lo largo del continuum fuerza-resistencia.
De manera similar a estudios previos (ver Staron y Johnson, 1993) el presente estudio encontró conversiones de tipos de fibras inducidas por el ejercicio dentro de la población de la fibras rápidas en la dirección de las fibras tipo IIB a las IIA. Esta transformación del tipo de fibras ocurrió en la misma magnitud en los tres grupos de entrenamiento, alcanzando aproximadamente un incremento de dos veces en el porcentaje de fibras clasificadas como tipo IIAB, con una disminución concomitante en las fibras clasificadas como “puras” tipo IIB. El hallazgo de conversiones de tipos de fibras similares entre los tres grupos quizás no es sorprendente considerando datos previamente publicados sobre el acondicionamiento aeróbico y anaeróbico en humanos (Staron y Johnson, 1993). Parece que cualquier estímulo de ejercicio (e.g., fuerza o resistencia) que sea suficiente en duración y/o intensidad tiene el potencial para finalmente causar conversiones dentro de la población de fibras rápidas desde el tipo IIB hasta IIA (Staron y Johnson et. al, 1995). Aunque esta transformación ocurrió en el presente estudio luego de los tres protocolos de entrenamiento, fueron notadas diferencias entre los grupos con respecto a la magnitud de la hipertrofia inducida por el ejercicio.
El resultado lógico y frecuentemente buscado después de un programa de entrenamiento de fuerza es un incremento en el tamaño y producción de fuerza de los músculos ejercitados. La hipertrofia parece ser el resultado de un incremento de la tasa de síntesis de proteínas (Chesley et al., 1992; Phillips et al., 1997), lo cual contribuye a un incremento absoluto en al cantidad de elementos contráctiles (MacDougall et al., 1979; 1982; Lüthi et al., 1986). Numerosos estudios han demostrado una respuesta de hipertrofia para todos los principales tipos de fibras (I, IIA, y IIB) luego del entrenamiento de la fuerza de corta duración en individuos jóvenes y ancianos previamente desentrenados (e.g., Staron et al., 1990, 1991; Hikida et al. 2000). Sin embargo, esta hipertrofia inducida por el ejercicio parece afectar a las fibras rápidas en una mayor magnitud que a las fibras lentas, fibras tipo I (ver Tesch, 1987). En el presente estudio, la respuesta hipertrófica fue minimizada (esencialmente anulada) en aquellos individuos en el grupo HR. Por el contrario, el área de sección transversal de los tres tipos de fibras principales se incremento para aquellos sujetos que entrenaron en el extremo de la fuerza del continuum (Tabla 5). Interesantemente, la respuesta hipertrófica fue similar entre los grupos LR e IR. Ha sido frecuentemente aceptado que el incremento de la fuerza/potencia resulta a partir del entrenamiento de alta intensidad/bajo volumen, mientras que el entrenamiento de baja intensidad/alto volumen maximiza la hipertrofia muscular (Hisaeda et al., 1996). En base a los datos de la presente investigación, esto pude no ser completamente cierto. En efecto, los datos de la presente investigación sugieren que el entrenamiento de RM bajas e intermedias induce adaptaciones musculares similares, al menos después del entrenamiento de corta duración en sujetos previamente desentrenados.
Capilaridad
Los resultados de los estudios de entrenamiento de la fuerza que investigaron los cambios capilares son erróneos, con reportes de incrementos, disminuciones y sin cambios desde la condición pre- a post-entrenamiento (e.g., Schantz, 1982; Tesch et al., 1984, 1990; Hather et al., 1991; Wang et al., 1993; McCall et al., 1996; Hagerman et al., 2000). En el presente estudio, hubo una tendencia de incremento en el número de capilares por fibra con el entrenamiento, indicando la formación de nuevos capilares dentro del músculo (Tabla 6). Tales hallazgos han sido reportados previamente luego del entrenamiento de la fuerza en humanos (Hather et al., 1991; McCall et al., 1996; Hepple et al., 1997; Green et al., 1998), y sugieren que los cambios en los capilares son proporcionales a los cambios en el tamaño de las fibras. En otras palabras, el crecimiento capilar puede haber estado enmascarado por el incremento en el área ocupada por las fibras musculares. Así, la densidad capilar no cambio en el presente estudio, aunque ocurrió una cantidad significativa de hipertrofia luego del entrenamiento para los grupos LR e IR.
Aunque no hubo diferencias significativas, ya sea en CD o CF después del entrenamiento en el presente estudio, hubo una tendencia sugiriendo diferencias potenciales entre los grupos. CD cayó un 8% para el grupo LR, aumento un 4% para el grupo IR, y aumento un 7% para el grupo HR. Del mismo modo, hubo una tendencia al incremento en CF en una mayor cantidad luego del entrenamiento para los grupos IR y HR (23% y 18%, respectivamente) en comparación con los grupos LR y Con (4%). Tomados juntos, estos datos sugieren que el entrenamiento continuo puede haber resultado en diferencias significativas entre los grupos, dando apoyo a la idea acerca de que las adaptaciones capilares ocurren en un continuum que esta basado en la duración y la intensidad del entrenamiento. Así, el entrenamiento de fuerza de altas repeticiones/baja carga parece causar adaptaciones capilares más similares al entrenamiento de resistencia aeróbica en comparación con el entrenamiento de fuerza de bajas repeticiones/alta carga.
En conclusión, los tres regimenes de entrenamiento causaron alteraciones similares dentro de la población de tipos de fibras rápidas (conversión de fibras desde IIB a IIA) y en el contenido de MHC. Las diferencias fueron, sin embargo, aparentes entre los tres grupos de entrenamiento, en la respuesta hipertrófica, y para varios parámetros cardiorrespiratorios. Estas adaptaciones post-entrenamiento específicas contribuyeron obviamente a las diferencias encontradas entre los grupos para la fuerza dinámica máxima y la resistencia muscular local. Aquellos individuos que entrenaron con cargas más altas mejoraron más la fuerza máxima, mientras que aquellos que entrenaron con cargas más livianas mejoraron más usando el 60% de 1 RM. Interesantemente, ambos grupos que trabajaron sobre el extremo de la fuerza del continuum fuerza-resistencia (LR e ir) tuvieron una respuesta hipertrófica similar. Ha sido frecuentemente aceptado que las ganancias en la fuerza/potencia resultan a partir del entrenamiento de alta intensidad/bajo volumen, mientras que el entrenamiento de baja intensidad/alto volumen maximiza la hipertrofia muscular (Hisaeda et al., 1996). En base a los datos de la presente investigación, los entrenamientos de bajas e intermedias RM parecen inducir adaptaciones musculares similares, al menos después del entrenamiento de corta duración en sujetos previamente desentrenados. En general, sin embargo, los datos de la presente investigación demostraron que tanto el rendimiento físico como las adaptaciones fisiológicas asociadas, están vinculados con la intensidad y el número de repeticiones realizados.
Agradecimientos
Los autores quieren agradecer a los Departamentos Fotográficos y Gráficos de Medicina Osteopática de la Ohio University Collage por la ayuda con las figuras y tablas. También estamos agradecidos con aquellos individuos que nos ayudaron en la supervisión del entrenamiento y especialmente a los sujetos que se ofrecieron y participaron. Parte de este trabajo fue subvencionado por la O.U Collage of Osteopathic Medice y la Oficina de Investigación. G.E.R Campos fue apoyado por la FAPESP, 95/4744-4, San Pablo, Brasil.
Referencias
1. Anderson T, Kearney JT (1982). Effects of three resistance training programs on muscular strength and absolute and relative endurance. Res Q Exerc Sport 53:1-7
2. Bemben DA, Fetters NL, Bemben MG, Nabavi N, Koh ET (2000). Musculoskeletal responses to highand low-intensity resistance training in early postmenopausal women. Med Sci Sports Exerc 32:1949-1957
3. Berger R (1962). Optimum repetitions for the development of strength. Res Q 33:333-338
4. Blomstrand E, Ekblom B (1982). The needle biopsy technique for fibre type determination in human skeletal muscle - a methodological study. Acta Physiol Scand 116:437-442
5. Brooke MH, Kaiser KK (1970). Three "myosin ATPase" systems: the nature of their pH lability and sulfhydryl dependence. J Histochem Cytochem 18:670-672
6. Chesley A, MacDougall JD, Tarnopolsky MA, Atkinson SA, Smith K (1992). Changes in human muscle protein synthesis after resistance exercise. J Appl Physiol 73:1383-1388
7. Chestnut JL, Docherty D (1999). The effects of 4 and 10 repetition maximum weight-training protocols on neuromuscular adaptations in untrained men. J Strength Cond Res 13:353-359
8. Clarke DH, Stull GA (1970). Endurance training as a determinant of strength and fatigue ability. Res Q 41:19-26
9. DeLateur BJ, Lehmann JF, Fordyce WE (1968). A test of the DeLorme axiom. Arch Phys Med Rehabil 49:245-248
10. DeLorme TL (1945). Restoration of muscle power by heavy resistance exercise. J Bone Joint Surg 27:645-667
11. Dudley GA, Tesch PA, Miller BJ, Buchannan P (1991). Importance of eccentric actions in performance adaptations to resistance training. Aviat Space Environ Med 62:543-550
12. Ennion S, Pereira JS, Sargeant AJ, Young A, Goldspink G (1995). Characterization of human skeletal muscle fibres according to the myosin heavy chains they express. J Muscle Res Cell Motil 16:35-43
13. Evans WJ, Pinney SD, Young VR (1982). Suction applied to a muscle biopsy maximizes sample size. Med Sci Sports Exerc 14:101-102
14. Ferketich AK, Kirby TE, Always SE (1998). Cardiovascular and muscular adaptations to combined endurance and strength training in elderly women. Acta Physiol Scand 164:259-267
15. Fleck SJ, Kraemer WJ (1988). Resistance training: basic principles (Part 1 of 4). Physician Sportmed 16:161-171
16. Fleck SJ, Kraemer WJ (1997). Designing resistance training programs. Human Kinetics, Champaign, Illinois
17. Green H, Goreham C, Ouyang J, Ball-Burnett M, Ranney D (1998). Regulation of fiber size, oxidative potential, and capillarization in human muscle by resistance exercise. Am J Physiol 276:R591-R596
18. Guth L, Samaha FJ (1969). Qualitative differences between actomyosin ATPase of slow and fast mammalian muscle. Exp Neurol 25:138-152
19. Hagerman FC, Walsh SJ, Staron RS, Hikida RS, Toma K, Ragg KE (2000). Effects of high-intensity resistance training on untrained older men. I. Strength, cardiovascular, and metabolic responses. J Gerontol 55A:B336-B346
20. Hather BM, Tesch PA, Buchanan P, Dudley GA (1991). Influence of eccentric actions on skeletal muscle adaptations to resistance training. Acta Physiol Scand 143:177-185
21. Hepple RT, Mackinnon SLM, Thomas SG, Goodman JM, Plyley MJ (1997). Quantitating the capillary supply and the response to resistance training in older men. Pflügers Arch 433:238-244
22. Hickson RC, Rosenkoetter MA, Brown MM (1980). Strength training effects on aerobic power and short-term endurance. Med Sci Sports Exerc 12:336-339
23. Hickson RC, Dvorak BA, Gorostiaga EM, Kurowski TT, Foster C (1988). Potential for strength and endurance training to amplify endurance performance. J Appl Physiol 65:2285-2290
24. Hikida RS, Staron RS, Hagerman FC, Walsh SJ, Kaiser E, Shell S, Hervey S (2000). Effects of high intensity resistance training on untrained older men. II. Muscle fiber characteristics and nucleo-cytoplasmic relationships. J Gerontol 55A:B347-B354
25. Hisaeda H, Miyagawa K, Kuno S-Y, Fukunaga T, Muraoka I (1996). Influence of two different modes of resistance training in female subjects. Ergonomics 39:842-852
26. Howley ET, Bassett DR Jr, Welch HG (1995). Criteria for maximal oxygen uptake: review and commentary. Med Sci Sports Exerc 27:1292-1301
27. Jackson AS, Pollock ML (1978). Generalized equations for predicting body density of men. Br J Nutr 40:497-504
28. Jackson CGR, Dickson AL, Ringel SP (1990). Skeletal muscle fiber area alterations in two opposing modes of resistance-exercise training in the same individual. Eur J Appl Physiol 61:37-41
29. Johnston RE, Quinn TJ, Kertzer R, Vroman NB (1995). Improving running economy through strength training. Strength Cond 17:7-13
30. Kraemer WJ, Patton JF, Gordon SE, Harman EA, Deschenes MR, Reynolds K, Newton RU, Triplett NT, Dziados JE (1995). Compatibility of high-intensity strength and endurance training on hormonal and skeletal muscle adaptations. J Appl Physiol 78:976-989
31. MacDougall JD, Sale DG, Moroz JR, Elder GCB, Sutton JR, Howald H (1979). Mitochondrial volume density in human skeletal muscle following heavy resistance training. Med Sci Sports 11:164-166
32. MacDougall JD, Sale DG, Elder GCB, Sutton JR (1982). Muscle ultrastructural characteristics of elite powerlifters and bodybuilders. Eur J Appl Physiol 48:117-126
33. Marcinik E, Potts J, Schlabach G, Will S, Dawson P, Hurley B (1991). Effects of strength training on lactate threshold and endurance performance. Med Sci Sports Exerc 23:739-743
34. McCall GE, Byrnes WC, Dickinson A, Pattany PM, Fleck SJ (1996). Muscle fiber hypertrophy, hyperplasia, and capillary density in college men after resistance training. J Appl Physiol 81:2004-2012
35. McDonagh MJN, Davies CTM (1984). Adaptive responses of mammalian skeletal muscle to exercise with high loads. Eur J Appl Physiol 52:139-155
36. Perrie WT, Bumford SJ (1986). Electrophoretic separation of myosin isoenzymes. Implications for the histochemical demonstration of fibre types in biopsy specimens of human skeletal muscle. J Neurol. Sci 73:89-96
37. Pette D, Staron RS (2001). Transitions of muscle fiber phenotypic profiles. Histochem Cell Biol. 115:359-372
38. Phillips SM, Tipton KD, Aarsland A, Wolf SE, Wolfe RR (1997). Mixed muscle protein synthesis and breakdown after resistance exercise in humans. Am J Physiol 273:E99-E107
39. Plyley MJ, Groom AC (1975). Geometrical distribution of capillaries in mammalian striated muscle. Am J Physiol 228:1376-1383
40. Schantz P (1982). Capillary supply in hypertrophied human skeletal muscle. Acta Physiol Scand. 114:635-637
41. Smerdu V, Karsch-Mizrachi I, Campione M, Leinwand L, Schiaffino S (1994). Type IIx myosin heavy chain transcripts are expressed in type IIb fibers of human skeletal muscle. Am J Physiol. 267:C1723-C1728
42. Staron RS (1991). Correlation between myofibrillar ATPase activity and myosin heavy chain composition in single human muscle fibers. Histochemistry 96:21-24
43. Staron RS (1997). Human skeletal muscle fiber types: delineation, development, and distribution. Can J Appl Physiol 22:307-327
44. Staron RS, Hikida RS (1992). Histochemical, biochemical, and ultrastructural analyses of single human muscle fibers with special reference to the C fiber population. J Histochem Cytochem 40:563-568
45. Staron RS, Johnson P (1993). Myosin polymorphism and differential expression in adult human skeletal muscle. Comp Biochem Physiol 106B:463-475
46. Staron RS, Malicky ES, Leonardi MJ, Falkel JE, Hagerman FC, Dudley GA (1990). Muscle hypertrophy and fast fiber type conversions in heavy resistance-trained women. Eur J Appl Physiol. 60:71-79
47. Staron RS, Leonardi MJ, Karapondo DL, Malicky ES, Falkel JE, Hagerman FC, Hikida RS (1991). Strength and skeletal muscle adaptations in heavy-resistance-trained women after detraining and retraining. J Appl Physiol. 70:631-640
48. Staron RS, Karapondo DL, Kraemer WJ, Fry AC, Gordon SE, Falkel JE, Hagerman FC, Hikida RS (1994). Skeletal muscle adaptations during early phase of heavy-resistance training in men and women. J Appl Physiol 76:1247-1255
49. Stone WJ, Coulter SP (1994). Strength/endurance effects from three resistance training protocols with women. J Strength Cond Res 8:231-234
50. Stull GA, Clarke DH (1970). High-resistance, low-repetition training as a determiner of strength and fatigability. Res Q 41:189-193
51. Taaffe DR, Pruitt L, Pyka G, Guido D, Marcus R (1996). Comparative effects of high- and low-intensity resistance training on thigh muscle strength, fiber area, and tissue composition in elderly women. Clin Physiol 16:381-392
52. Tan B (1999). Manipulating resistance training program variables to optimize maximum strength in men: a review. J Strength Cond Res 13:289-304
53. Tanaka H, Swensen T (1998). Impact of resistance training on endurance performance. A new form of cross-training?. Sports Med 25:191-200
54. Tesch PA (1987). Acute and long-term metabolic changes consequent to heavy-resistance exercise. Med Sport Sci 26:67-89
55. Tesch PA (1988). Skeletal muscle adaptations consequent to long-term heavy resistance exercise. Med Sci Sports Exerc 20:S132-S134
56. Tesch PA, Thorsson A, Kaiser P (1984). Muscle capillary supply and fiber type characteristics in weight and power lifters. J Appl Physiol 56:35-38
57. Tesch PA, Thorsson A, Colliander EB (1990). Effects of eccentric and concentric resistance training on skeletal muscle substrates, enzyme activities and capillary supply. Acta Physiol Scand 140:373-380
58. Wang N, Hikida RS, Staron RS, Simoneau J-A (1993). Muscle fiber types of women after resistance training - quantitative ultrastructure and enzyme activity. Pflügers Arch 424:494-502