
Almacenamiento y Recobro de Energía Elástica en Músculos Esqueléticos Humanos de tipo de Fibras Lentas y Rápidas
Carmelo Bosco1, J. Tihani2, Paavo V Komi1, G. Fekete2 y P. Apor2
1Department of Biology of Physical Activity, University of Jyväskilä, Finland and Research Unit for Sport and Physical Fitness, Jyväskilä, Finland.
2Research Institute of the Hungarian College for Physical Education, Budapest, Hungary.
Artículo publicado en el journal PubliCE, Volumen 0 del año 1982.
Publicado 4 de enero de 2003
Resumen
Palabras clave: elasticidad muscular, fibras musculares, mecanismos musculares, energía elástica, ciclo estiramiento
INTRODUCCIÓN
Los ejercicios con ciclo estiramiento-acortamiento se caracterizan por aumentar el rendimiento cuando se los compara con el trabajo realizado en condiciones de solo acortamiento. Este incremento en el trabajo ha sido atribuido a un almacenamiento de energía elástica en el músculo durante el estiramiento y su re-uso como trabajo mecánico durante la fase concéntrica, si esta fase es realizada inmediatamente.
La utilización de energía elástica ha sido observada en una serie de saltos con rebote y con desplazamiento angular de la rodilla corto y largo (Bosco et al. 1982). En este estudio fue notado que la contribución al trabajo positivo total debido al re-uso de energía elástica, explico cerca del 50 %, en los saltos de amplitud corta y 30 %, en los saltos de amplitud larga. Sin embargo, en valores absolutos, en la condición posterior, el re-uso de energía fue mayor que en la anterior y a pesar del hecho de que los saltos de amplitud larga estuvieron caracterizados por un período de transición largo entre las fases excéntricas y concéntricas. Para examinar este otro fenómeno, fue pensado que la utilización de energía elástica podría derivar diferentemente a partir de las fibras de contracción rápida y lenta, que poseen diferentes tiempos de unión. Ya ha sido sugerido un modelo mecánico de este comportamiento (Bosco 1982). El presente estudio fue diseñado para evaluar esta hipótesis en sujetos con diferentes composiciones musculares de fibras y que realizaron saltos verticales con diferentes desplazamientos angulares de la rodilla.
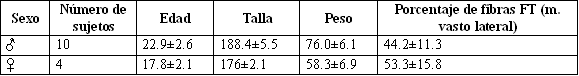
Tabla 1. Características físicas de los sujetos hombres y mujeres.
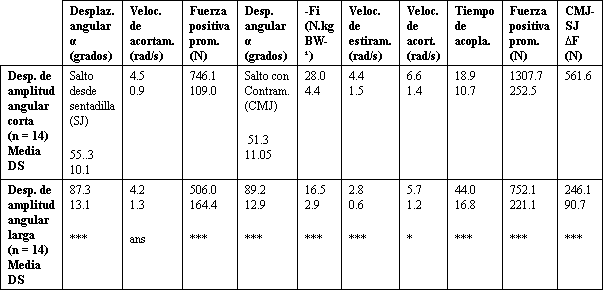
Tabla 2. Características mecánicas de los CMJ y SJ, de amplitud larga y corta.
Las estrellas denotan la significancia estadística (test-t de Student) entre
saltos de amplitud larga y corta (*p<0.05, **p<0.01, ***p<0.001, ans = no
significativo).
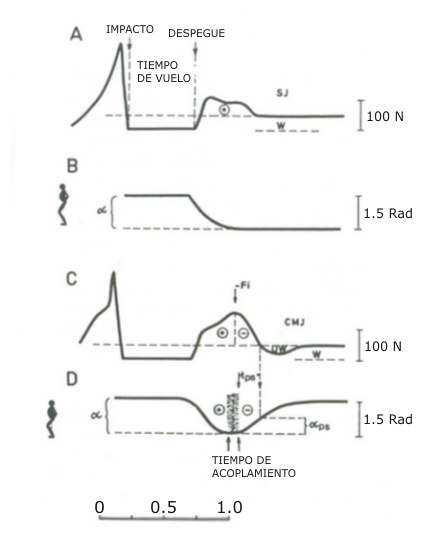
Figura 1. Ejemplos de curva fuerza vertical-tiempo y desplazamiento angular
durante saltos estáticos (SJ) y con contramovimiento (CMJ). En los SJ se leen
valores positivos de la curva fuerza-tiempo durante el despegue como es
indicado por el signo +. En el CMJ, la fuerza de reacción vertical del suelo
es primero negativa durante la fase inferior al peso corporal (UW), pero da
valores positivos, durante la fase de deceleración – y en la fase de trabajo
positivo +. El tiempo de acoplamiento en los registros de desplazamiento
angular, indica el período de transición entre las fases excéntrica y
concéntrica.
MÉTODOS
14 sujetos que realizaban deportes de potencia, bien entrenados (10 hombres y 4 mujeres) fueron usados como muestra. Todos los sujetos se familiarizaron con los test usados antes de ser evaluados.
Esto consistió en realizar saltos verticales máximos en una plataforma de fuerza (frecuencia natural de 100 Hz) con y sin contramovimiento. Estos saltos fueron llamados, respectivamente, salto con contramovimiento (CMJ) y salto desde sentadilla (SJ). Cada salto produjo una curva de fuerza vertical – tiempo, ejemplos de los cuales son dados en la Fig. 1A-C. La fuerza registrada durante el contacto es siempre positiva en los SJ, mientras que en los CMJ es primero negativa (inferior al peso corporal) durante la fase inferior al peso corporal (UW), pero se mantiene positiva durante la fase de desaceleración – y durante la fase de trabajo positivos +. La curva fuerza – tiempo y el tiempo de vuelo (Tair) dieron las bases para calcular los siguientes parámetros: velocidad vertical de despegue (Vv), fuerza positiva promedio (F). En el CMJ la fuerza instantánea desarrollada al final del estiramiento (- Fi) fue también computada. La velocidad vertical de despegue fue obtenida de la fórmula:
Vv = ½ x Tair x g
Donde g = aceleración de la gravedad (9.81 m x s-1).
La fuerza promedio (F) fue luego calculada con la siguiente fórmula: F = M x Vv / t
Donde t = el tiempo respectivo de la curva fuerza – tiempo.
M = masa del sujeto
Vv = Velocidad final de despegue
Con el objetivo de examinar la amplitud de movimiento durante el SJ y CMJ, un electrogoniómetro (Elgon) fue colocado en el lado lateral de la articulación de la rodilla de los sujetos. A partir del desplazamiento angular total (α) que incluía a ambas, fase inferior al peso corporal y fase de estiramiento en el CMJ, la segunda fase también pudo ser computada como es demostrado por el α ps en la Fig. 1. El área sombreada de la Fig. 1 representa la posición del grabador Elgon, donde el ángulo de la rodilla se mantenía igual y supuestamente no ocurrían ningún cambio en los músculos extensores de la rodilla. Esta fase ha sido llamada por Bosco et al. (1981) “Fase de Acoplamiento”, para definir el período de transición entre las fases de estiramiento (excéntrica) y acortamiento (concéntrica). La duración de esta fase fue medida manualmente usando la tangente de las curvas de desplazamiento angular de la rodilla. La exactitud de estas mediciones fue ±2 y ±3, para saltos de amplitud corta y larga, respectivamente.
Determinación de la composición de fibras
La técnica de biopsia introducida por Bergström (1962) fue usada para obtener muestras del músculo vasto lateral. Se realizaron cortes criostáticos con las muestras que luego fueron teñidas para la ATPasa miofibrilar (Adenosin Trifosfatasa) (Padykula & Herman 1965) para la clasificación de las fibras FT y ST de acuerdo con Gollnick et al. (1972).
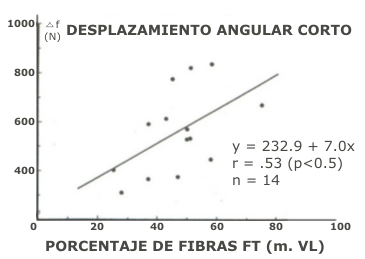
Figura 2. Diferencias de fuerza promedio de los CMJ y SJ (ΔF) realizados con
desplazamiento angular corto, se presentan en función del porcentaje de fibras
de contracción rápida en el músculo vasto lateral.
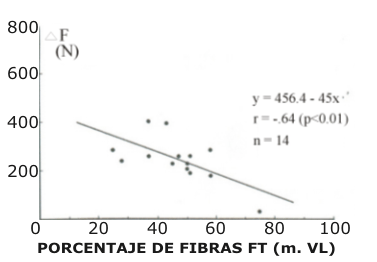
Figura 3. Diferencias de fuerza promedio de los CMJ y SJ (ΔF) realizados con
desplazamiento angular largo, se presentan en función del porcentaje de fibras
de contracción rápida en el músculo vasto lateral.
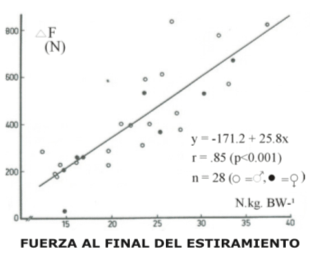
Figura 4. La diferencia de fuerza promedio entre el CMJ y el SJ (ΔF) realizada
con desplazamientos angulares de la rodilla cortos y largos es ploteada versus
la fuerza instantánea (Fi) desarrollada al final de estiramiento y expresada
en Kg. BW-1 para normalizar el peso diferente de los hombres (○) y las mujeres
(•).
RESULTADOS
En la tabla 2 son mostrados los parámetros mecánicos seleccionados, calculados para saltos de amplitud corta y larga. La diferencia de fuerza positiva promedio (ΔF) entre los saltos CMJ y SJ, que utilizaron desplazamientos del ángulo de la rodilla similares en la fase concéntrica, fueron elegidos como parámetro para reflejar la potenciación de estiramiento (Bosco et al. 1981). Cuando el ΔF fue calculado para desplazamientos angulares cortos de la rodilla, mostró una correlación positiva con el porcentaje de fibras de contracción rápida (FT), r = 0.53, p < 0.05 (Fig. 2). De otro modo, cuando el mismo parámetro mecánico ΔF fue calculado para los desplazamientos angulares largos, y ploteado versus la distribución de fibras, fue observado una correlación negativa, r = -0.64, p < 0.05 (Fig. 3).
El efecto de potenciación, ΔF, notado para desplazamientos angulares largos y cortos demostró una fuerte correlación con la fuerza desarrollada al final de la fase de estiramiento (-Fi) en el CMJ, r = 0.84, p < 0.01 (Fig. 4). La –Fi demostró una correlación positiva con la composición de fibras en los desplazamientos angulares cortos (r = 0.57, p < 0.05, Fig. 5) pero no en los desplazamientos angulares largos (Fig. 6)
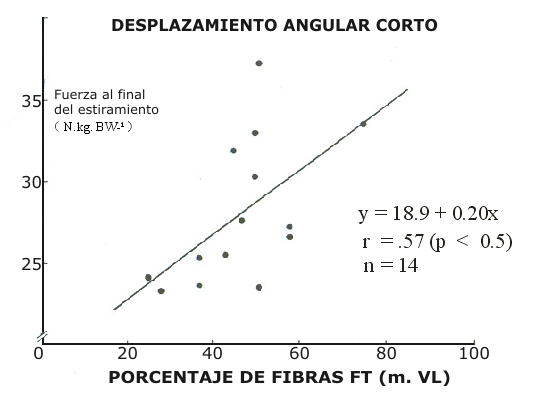
Figura 5. Correlación entre la fuerza instantánea desarrollada al final del
estiramiento en los CMJ de amplitud corta y porcentajes de fibras de
contracción rápida en el músculo vasto lateral.
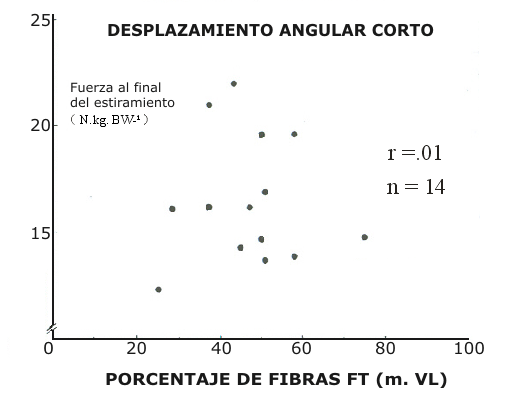
Figura 6. Correlación entre la fuerza instantánea desarrollada al final del
estiramiento en los CMJ de amplitud larga y porcentajes de fibras de
contracción rápida en el músculo vasto lateral.
DISCUSIÓN
El efecto de potenciación, ΔF, expresado como la diferencia de fuerza positiva promedio entre los CMJ y el SJ, demostró una correlación positiva con la composición muscular de fibras en los saltos realizados con desplazamientos angulares cortos (Fig. 2). La explicación de este hallazgo puede ser pensada a través del comportamiento visco-elástico del músculo durante un movimiento balístico rápido.
El recobro de energía elástica durante el trabajo positivo depende de la cantidad de energía elástica almacenada en el músculo durante el trabajo excéntrico. Esta magnitud puede ser representada en este estudio, por la fuerza desarrollada al final del estiramiento. Esta suposición es apoyada por Flitney & Hirst (1978) que señalaron que, durante la fase de estiramiento, parte de la tensión desarrollada es tomada por los elementos elásticos dispuestos en serie con los sarcómeros. Esto puede ocurrir por la rotación de las cabezas de miosina hacia atrás, en contra de su tendencia natural de mantenerse en una posición de mayor energía potencial. De hecho esto significa almacenamiento de trabajo mecánico en los puentes cruzados de los sarcómeros. Esta energía almacenada puede ser re-usada durante el trabajo positivo siguiente, si es permitido que el músculo se contraiga inmediatamente después del estiramiento (Cavagna et al. 1965).
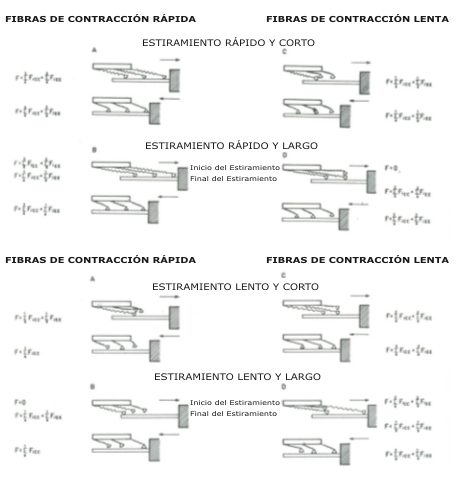
Figura 7. Un modelo esquemático para sugerir un ciclo de unión-separación de
los puentes cruzados en las fibras de contracción rápida (izq.) y lenta
(der.), trabajando a diferentes velocidades y con diferentes longitudes de
estiramiento. Los cálculos consideraron a los puentes cruzados trabajando en
paralelo consigo mismos. En las condiciones B y D, son mostradas dos
ecuaciones para la fase de estiramiento. La ecuación de arriba se refiere al
estado de los puentes cruzados al inicio del estiramiento, y la de abajo al
mismo al final del estiramiento. Este modelo también esta basado en la
hipótesis de Huxley & Simmons (1971) y Cavagna & Criterio (1974) (tomado Bosco
1982).
Como fue presentado en la Tabla 3, la –Fi fue de 30.2 ±4.8 y 25± 4.8 N kg BW¹, en los saltos de amplitud corta, para los grupos con predominio de fibras FT y ST, respectivamente. De este modo, los sujetos del grupo FT fueron capaces de almacenar una mayor cantidad de energía mecánica durante la fase excéntrica. Consecuentemente esta –Fi demostró una correlación significativa con la composición de fibras en los desplazamientos angulares cortos (r = 0.57, p< 0.05, Fig. 5), pero no en los desplazamientos angulares largos.
A pesar de esta diferencia en la –Fi, el efecto de potenciación relativo no fue significativamente diferente en los grupos de sujetos (Tabla 3). Esto implica que en los saltos de amplitud corta la fase de almacenamiento de energía (estiramiento) es causada probablemente debido a un reclutamiento más rápido de unidades motoras, en el grupo con predominio de FT. Ha sido presentada evidencia de que tanto la tasa de desarrollo de la fuerza como la fuerza de estiramiento, son mayores en sujetos ricos en fibras de contracción rápida en el vasto lateral (Komi & Bosco 1978, Bosco & Komi 1979).
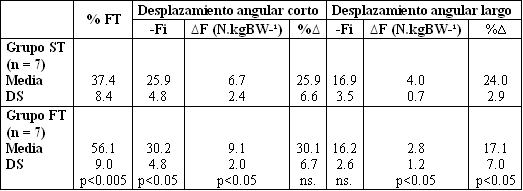
Tabla 3. Media (±DS) de la distribución de fibras, fuerza al final del
estiramiento (-Fi), efecto de potenciación (∆F) y porcentaje de re-uso de la
energía elástica almacenada ∆F-Fi (%∆), calculados en saltos de amplitud larga
y corta, los mismos están expresados separadamente para los grupos de fibras
rápidas y lentas. También es mostrada la significancia estadística (test-t de
Student).
Si el patrón de reclutamiento de unidades motoras es responsable de la diferencia en la obtención de –Fi en los grupos examinados, la razón por la que el porcentaje de recobro de la energía elástica almacenada es similar, puede ser examinado desde el punto de vista del ciclo unión-separación de los puentes cruzados del sarcómero. Los saltos de amplitud corta estuvieron caracterizados por un tiempo de acoplamiento corto (Tabla 2) y este tiempo de transición corto no parece ser un factor limitante para el re-uso de energía elástica almacenada. Aunque la literatura no es uniforme en términos de tiempo de vida de los puentes cruzados (ver Curtin et al. 1974, Stienen et al. 1978) parece que el período de transición disponible entre el estiramiento y el acortamiento fue corto, y no pudo ser observado un re-uso de energía elástica significativamente diferente entre los sujetos con tipos de fibras rápidas y lentas. Así en los saltos de amplitud corta el recobro de energía elástica, expresado en valores absolutos, es proporcional al almacenamiento de energía elástica durante la fase de estiramiento sin tener en cuenta la composición muscular de las fibras.
Los saltos de amplitud larga, de otro modo, representaron movimientos donde pudieron ser observadas diferencias significativas en la potenciación entre los grupos FT y ST (Tabla 3). La fuerza excéntrica final (–Fi) fue, sin embargo, similar 16.2 ±2.6 vs. 16.9 ± 3.5 N.kg.BW¹, respectivamente, y en contraste con los saltos de amplitud corta. En estos saltos de amplitud larga, la fase excéntrica es relativamente larga (147 ms, de promedio) y de este modo también los sujetos ST tienen más tiempo para el reclutamiento de unidades motoras y consecuentemente la –Fi debería ser la misma. La observación de que los valores –Fi fueron diferentes en los dos tipos de saltos en los dos grupos (Tabla 2), es debido a diferencias en las ventajas mecánicas en la posición donde la –Fi fue registrada (ver métodos y Smith 1973). De este modo el hallazgo de que el grupo ST tubo una potenciación mucho más grande que en el grupo FT en los saltos de amplitud larga (24.0 ±2.9 % vs. 17.1 ± 7.0 %, p< 0.05, Tabla 3) es explicable a través de la hipótesis de Bosco (1982), que destaca que el tiempo de acoplamiento largo favorece a los músculos de fibras lentas para retener su energía elástica más tiempo sin la separación de los puentes cruzados. Esta hipótesis esta en línea con la observación de ciclos de puentes cruzados más rápidos en las fibras FT (Länergren 1976). En los saltos de amplitud larga con tiempos de acoplamiento largos, algunos de los puentes cruzados de las fibras FT van a estar desunidos y su potencia elástica, perdida. Esta es probablemente la razón de porque el efecto de potenciación es más alto en sujetos ricos en fibras de contracción lenta en saltos realizados con un desplazamiento angular de la rodilla largo.
En conclusión, puede ser sugerido que las fibras ST y FT están caracterizadas por propiedades visco-elásticas diferentes que les permiten beneficiarse de manera diferente a partir de los ciclos estiramiento-acortamiento, dependiendo de si el movimiento es rápido o lento. La Fig. 7 hace un intento de explicar esquemáticamente esta sugerencia.
Este trabajo se realizo en el Research Institute of Hungarian Collage for Physical Education, durante una visita de investigación del autor (C.B.). El trabajo fue apoyado por una subvención de la Asociación Italiana de Agricultura, y por el Ministerio de Educación (Finlandia).
Referencias
1. Bosco, C (1982). Stretch-shortering cycle in skeletal muscle function with special reference to elastic energy and potentiation of myoelectrical activity. Studies in Sport, Physical Education and Health, No 15, University of Jyväskyla, Finland
2. Bosco, C. & Komi, P.V (1979). Mechanical characteristics and the fiber composition of human leg extensor muscles. Eur J Appl Physiol 41: 275-284
3. Bosco, C. & Komi, P.V. & ITO, A (1981). Prestretch potentiation of human skeletal muscle during ballistic movement. Acta Physiol Scand; 111: 135-140
4. Bosco, C., Ito, A., Komi, P. V, Luhtanen, P., Rahkila, P.,Rusko, H. & Viitasalo, J. T (1982). Neuromuscular function and mechanical efficiency of human leg extensor muscles during jumping exercises. Acta Physiol Scand 114: 543-550
5. Cavagna, G. A, & Criterio, G (1974). Effect of streching on the elastic characteristics and contractile component of frog striated muscle. J Physiol (Lond) 239: 1-4
6. Cavagna, G. A., Saibene, F. P. & Margaria, R (1965). Effect of negative work on the amount of positive work performed by an isolated muscle. J Appl Physiol 20: 157-158
7. Curtin, N., Gilbert, C., Kretzschmar, K.M. & Wilkie, D.R (1974). The effect of the performance of work on total energy output and metabolism during muscular contraction. J Physiol (Lond) 238: 455-472
8. Flitney, F. W. & Hirst, D. G (1978). Cross bridge detachment and sarcomere give during strech of active frogs muscle. J Physiol (Lond) 276: 449-465
9. Huxley, A.F. & Simmons, R.M (1971). Mechanical properties of the cross-bridges of frog striated muscle. J Physiol (Lond) 218: 59-60P
10. Komi, P.V., Bosco,C (1978). Utilization of elastic energy in jumping and its relation to skeletal muscle fiber composition. In: Biomechanics VI A (ed. E. Asmussen and K. Jorgersen), pp. 79-85. University Park Press, Baltimore
11. Padykula, R. & Herman, E (1955). The specificity of histochemical method of adenosine triphosphatase. J Histochem Cytochem 3: 170-195
12. Smith, GI (1973). Biomechanical analysis of knee flexion and extension. J Biomechanics 6: 79-82
13. Stienen, G.J.M., Blange, T. & Schneer, M (1978). Tension response of frog sartorius muscle to quick ramp-shaped and some effects of metabolic inhibition. Pflügers Arch Eur J Physiol 376: 97-104