
Análisis ‘clásico’ de las Respuestas Metabólicas al Ejercicio Intermitente
Publicado 29 de julio de 2013, 4:05

Siguiendo con la temática del ejercicio intermitente
y su uso en ambientes de fitness y salud, es imprescindible comprender y
revisar (críticamente) los fundamentos que históricamente fueron la base de
este tipo de metodología, aunque aún se los sigue citando en la actualidad. Nos
referimos a la serie de estudios de Essén y de Edwards (en
menor medidia) acerca del ejercicio intermitente. Los estudios de Essén fueron
publicados en el Acta Physiologica Scandinava en los finales de la década del
setenta (hoy se llama Acta Physiologica y pertenece a la Federation of European
Physiological Societies). Se titulaba “Studies on the Regulation of Metabolism in Human Skeletal
Muscle Using Intermittent Exercise as an Experiment Model” llevados a cabo por el
Department of Clinical Physiology, del Karolinska Institute (Suecia, 1978).
Tiempo atrás, se pensaba que durante el trabajo de moderada intensidad, los lípidos constituían una gran proporción de la utilización de los sustratos, al menos si la duración del ejercicio era de 30’-40’. A medida que aumenta la intensidad, los CHO se tornan más importantes como fuente de sustratos. Pero como advirtieran Ǻstrand y Christensen en la década de los años sesenta, que las situaciones de la vida diaria son de carácter intermitente, los lípidos constituyen relativamente más a la utilización de sustratos que durante un ejercicio continuo de la misma intensidad.
En uno de esos estudios, se
intentó cuantificar la utilización de sustratos tanto intra- como
extra-musculares de los lípidos y los CHO durante un período prolongado de
ejercicio intenso intermitente (15” de trabajo y 15” de descanso pasivo) y
durante el ejercicio continuo, con el mismo VO2
promedio (≈ potencia promedio). Cinco sujetos varones (24 años; 70kg
de peso corporal; 1,81 m de altura y 4,2
l.min-1 de VO2máx,
valores promedios) realizaron un protocolo de ejercicio de pedaleo en forma
intermitente (15”x15”) durante 60’, con una carga promedio de 299 W (270-343 W)
correspondiente al 55% del VO2máx en promedio (pero en la fase de
trabajo, se ejercitaba 15” al 110% del VO2máx). A los 6 meses, los
mismos sujetos realizaron un protocolo de ejercicio continuo, con una carga
promedio de 157 W (138-180 W), es decir casi al 50% de la carga del protocolo
intermitente, y al 55% del VO2máx durante 60’ también. A todos los
sujetos se les realizaban biopsias musculares en los distintos momentos del
ejercicio, lo que podemos imaginar lo cruento que habrá sido tal toma de
muestras de biopsias musculares. También muestras de sangre y de VO2. Algunos
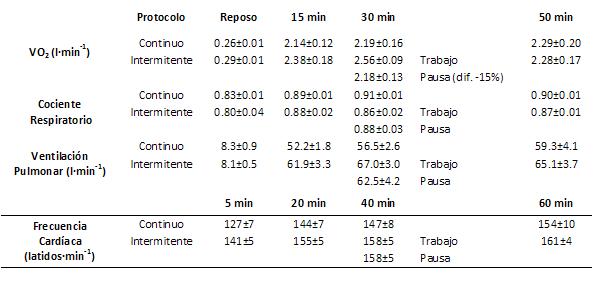
resultados se muestran en la Tabla 1. El VO2 aumentó 9-10 veces
respecto al reposo. El VO2 durante los primeros 30’ en el ejercicio
intermitente fue más alto que en el ejercicio continuo. Al comparar los
períodos de trabajo y pausa en el ejercicio intermitente, el VO2 osciló
ligeramente alrededor del valor calculado para la potencia promedio, y fue más
alto durante el período de trabajo que durante la pausa. El cociente
respiratorio también fluctuó ligeramente, salvo que en el ejercicio continuo
estuvieron los valores más altos. La FC aumentó progresivamente poco desde los
5’ hasta los 60’ en ambas situaciones.
Tabla 1. Consumo de oxígeno, cociente respiratorio, ventilación pulmonar y frecuencia cardíaca en reposo y durante el ejercicio continuo e intermitente (cada figura representa el promedio ± de la media).
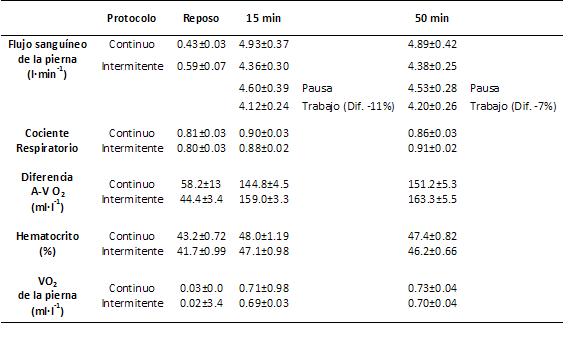
Otros resultados del mismo estudio se pueden ver en la Tabla 2, con el dato llamativo que durante el período de descanso en la modalidad intermitente reveló una tendencia hacia un mayor flujo (un +9%) durante el período de descanso (4,56±0,34 l·min-1) que durante el período de ejercicio (4,18±0,25 l·min-1). La diferencia arteriovenosa femoral aumentó 3 ó 4 veces con el ejercicio, y fue significativamente más alta durante el ejercicio intermitente a los 15’ como a los 50’ de ejercicio con un valor promedio de 161±4 ml·l-1 comparado con 148±5 ml·l-1 en el ejercicio continuo.
Tabla 2. Flujo sanguíneo, cociente respiratorio, diferencia
arteriovenosa, hematocrito y consumo de oxígeno de la pierna estudiada, en reposo
y durante el ejercicio continuo e intermitente (cada figura representa el promedio ± de la media)
Las concentraciones arteriales de sustratos se pueden observar en las Figuras 1, 2 y 3.
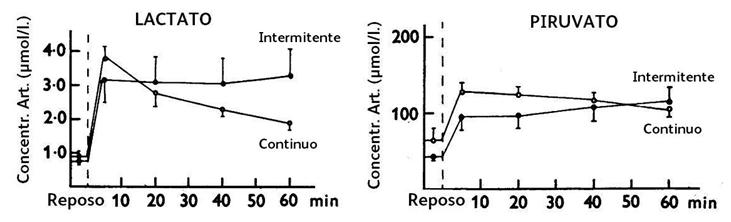
Figura 1. Concentración arterial de lactato y piruvato, en reposo y durante el ejercicio intermitente y continuo.
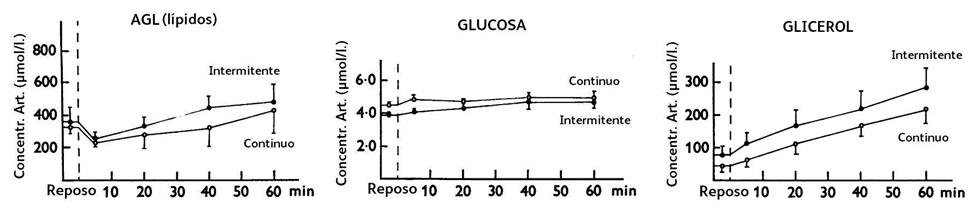
Figura 2. Concentración arterial de AGL, Glucosa y Glicerol, en reposo y durante el ejercicio continuo e intermitente.
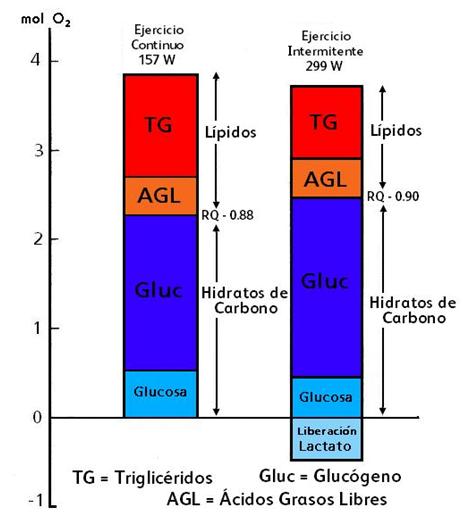
Figura 3. Sustratos sanguíneos utilizados durante 60’ de ejercicio continuo a una intensidad de 157 W y ejercicio intermitente alternando períodos de 15” de trabajo (a 299 W) y 15” de descanso pasivo. Las barras muestran la producción de energía en base a mediciones de consumo de oxigeno de la pierna. La contribución a partir de la oxidación de AGL y glucosa a partir de la sangre y la contribución para los CHO y oxidación de grasas, respectivamente, como también la liberación de lactato. El uso de glucógeno muscular es estimado como la diferencia entre la oxidación total de CHO y consumo de glucosa y la utilización intramuscular de triglicéridos es determinada como la oxidación total de las grasas sustrayendo la oxidación relacionada al consumo de grasas.
Se determinó la contribución al metabolismo total oxidativo a partir de los sustratos circulantes en la sangre: fue del 13% para los AGL (ácidos grasos libres) y del 11% para la glucosa en el ejercicio intermitente, y ocurrió lo mismo en el ejercicio continuo donde el 11% fue derivado de los AGL y el 13% de la glucosa. Es decir, los AGL y la glucosa juntos constituyen casi el 30% del metabolismo oxidativo al final del ejercicio, tanto en forma intermitente como en forma continua. Sin embargo, el RQ de la pierna indica que los AGL contribuyen al 40% de la utilización de las grasas totales en el ejercicio intermitente y solamente el 28% durante el ejercicio continuo. Esto está en línea con la utilización ligeramente mayor de los triglicéridos intramusculares durante el ejercicio continuo que durante el ejercicio intermitente. La glucosa sanguínea cubre aproximadamente el 17% de la utilización de los CHO en el ejercicio intermitente y el 23% en el ejercicio continuo. La restante proporción corresponde al glucógeno intramuscular.
En las Figura 4 y 5, se pueden observar los sustratos
intramusculares. Allí se ve una caída del 20% del glucógeno muscular ya después
de 5’ en ambos tipos de ejercicio y del 60% el final del ejercicio, siendo
siempre un poco más alta esa caída durante la modalidad intermitente. Los
triglicéridos disminuyeron después de los 5’ en ambas situaciones y la
utilización de los mismos después de los 60’ fue también descendiendo, habiendo
una diferencia significativa mayor en el ejercicio continuo que en el ejercicio
intermitente. Respecto al contenido de ATP-CP, se observó una caída del 20% a
los 5’ y a los 60’ en el ejercicio intermitente comparado al ejercicio
continuo. Y durante el ejercicio intermitente específicamente, el CP disminuyó
(-26%) al final del período de trabajo comparado al período de pausa. El ATP se
mantuvo casi sin cambios significativos. El lactato intramuscular fue más alto
(+24%) durante el período de trabajo que durante el período de reposo en el
protocolo intermitente. La concentración de citrato aumentó con el ejercicio
continuo desde reposo hasta los 5’ (+35%) y a los 60’ (+82%). Durante el
ejercicio intermitente, la concentración de citrato fue más alta (+25%) al
final del período de pausa que durante el período de trabajo (ver Figura 6).
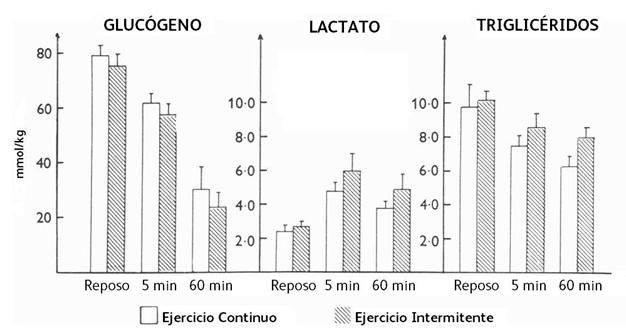
Figura 4. Concentración intramuscular de glucógeno, lactato y triglicéridos en reposo y después de 5’ y 6’’ de ejercicio de pedaleo intermitente (intensidad, 299 W; y períodos de 15”x15”) y continuo (intensidad, 157 W).
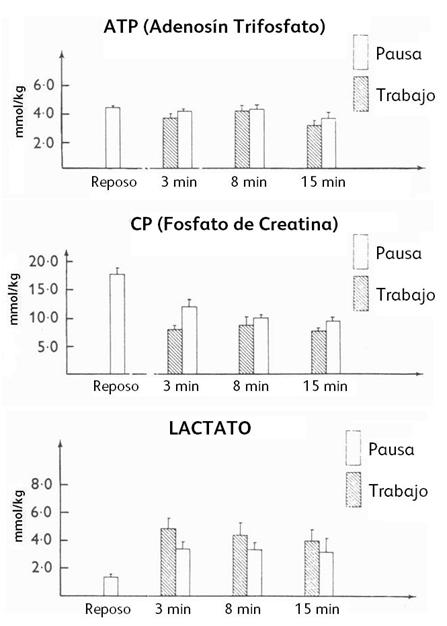
Figura 5. Concentración intramuscular de ATP, CP y lactato, en reposo y al final de un período de trabajo y pausa a los 3’, 8’ y 15’ de ejercicio intermitente.
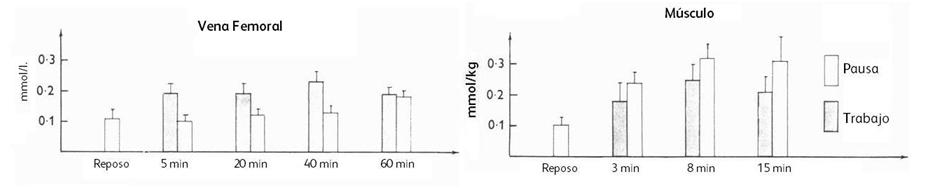
Figura 6. Concentración de citrato en la vena femoral en
reposo y durante los períodos de trabajo y pausa en el ejercicio intermitente a
los 5’, 20’, 40’ y 60’ y concentración intramuscular de citrato en reposo y al
final de los períodos de trabajo y pausa a los 3’, 8’ y 15’.
Todos estos datos, según los autores indicaban que la proporción de energía derivaba de las grasas y los CHO en forma similar tanto en el ejercicio intermitente como en el ejercicio continuo. Y en la Figura 7, los presentes datos permiten el cálculo de la contribución del metabolismo aeróbico y anaeróbico en las piernas durante cada carga de trabajo. A pesar del doble de carga durante el ejercicio intermitente respecto al ejercicio continuo, los hallazgos del estudio de Essen muestran similares contribuciones de las grasas y los CHO al metabolismo energético.
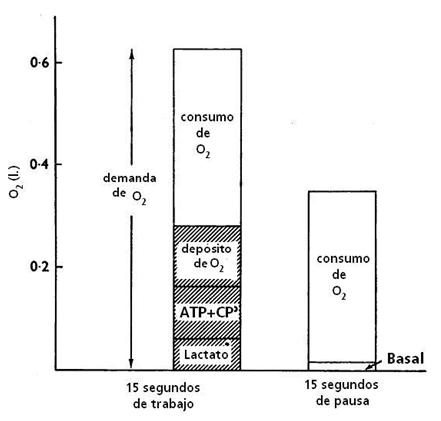
Figura 7. La demanda de oxígeno durante 15 segundos de trabajo y
el consumo de oxígeno promedio medido en los períodos de trabajo y pausa en las
piernas que se ejercitan en el ejercicio intermitente. La contribución al
déficit de oxígeno durante los períodos de trabajo a partir de la acumulación
de lactato, depleción de los fosfágenos y de la mioglobina, son indicados en
las áreas sombreadas.
Sin
embargo, Essén concluye que el ejercicio intermitente realizado en períodos de
15” de trabajo por 15” de descanso (pasivo), una liberación de energía aeróbica
predomina con una sustancial contribución de la utilización de los lípidos.
Indicando así, que la tasa de glucólisis es menor que durante el ejercicio
continuo a la misma intensidad. El mecanismo
sería el siguiente (según Essén): durante el período de trabajo, el ATP es
hidrolizado (proceso químico en el que una sustancia es dividida en compuestos
más simples por la suma de agua y se libera energía) y la concentración de ADP
y AMP aumentan, acentuando la glucólisis y la tasa del ciclo del citrato (Ciclo
de Krebs). Al final de un período de trabajo o en apenas el comienzo de un
período de pausa, la concentración de ADP-AMP estará alta, pero durante el
período de pausa, el nivel de ATP es resguardado. El citrato, entonces se
acumula y pasa a través de la membrana mitocondrial hacia adentro del
citoplasma. Al aumentar la concentración de citrato en el citoplasma, inhibe a
la glucólisis tanto a nivel de la PFK (fosfofrutokinasa) como de la PDH. Es
decir, cuanta más actividad del ciclo de Krebs es retardada, más citrato se
acumulará debido a una producción de acetil-CoA a partir de la oxidación de
ácidos grasos. Esto hace que el efecto del citrato potencie el efecto
inhibitorio del ATP sobre la PFK. El aumento del ATP al final de los períodos
de descanso en el ejercicio intermitente elevaría más la relación ATP/ADP y
esto es sabido que disminuye la actividad de la isocitratodeshidrogenasa
(enzima alostérica, que es estimulada por la baja carga energética celular, el
ADP, e inhibida por el NADH) en el ciclo de Krebs. Esta acumulación de citrato
encontrado al final de los períodos de descanso potenciaría dicho efecto
inhibidor (como se ve la Figura 8). La acumulación intramitocondrial de
citrato, el acetil-CoA y el NADH generados en la ß-oxidación de los AGL podría
inhibir aún más a la PDH, por tanto, limitaría la utilización de CHO. Por ello,
la continua pero leve liberación de lactato y piruvato que ocurre durante el
período global del ejercicio intermitente podría indicar que el piruvato fue
producido en exceso de lo que podría ser oxidado en actil-CoA. Todo esto
tendería a enlentecer el ritmo de la glucólisis durante el período de trabajo
siguiente, facilitando la utilización de los lípidos para la liberación de
energía aeróbica. Es decir, se creía que esto era causado por la continua
producción de acetil-CoA a partir de la ß-oxidación junto con la menor
actividad del ciclo de Krebs durante el período de pausa.

Figura 8. Contenido muscular de ATP, CP, citrato y lactato, en
reposo y al final de un período de 15” de trabajo por 15” de pausa (a partir de
datos combinados a los 5’, 15’, 30’ y 60’); y en reposo, inmediatamente y
15”-20” después de 5’ de ejercicio intenso continuo; ambas situaciones con
igual alta carga de trabajo.
Y además,
agrega Essen, que las fluctuaciones observadas en las concentraciones entre los
períodos de trabajo y pausa pueden reflejar oscilaciones en la concentración
del citrato muscular con el efecto antes indicado. La relación de tiempo entre
el aumento de citrato y la fase de trabajo no puede ser determinada a partir de
las concentraciones sanguíneas de citrato a causa del retraso (ver Figura 9).
Después de un turno de trabajo en el ejercicio intermitente, el nivel de
citrato aumenta rápidamente mientras que después del ejercicio continuo, el
aumento fue más gradual (ver Figura 8).
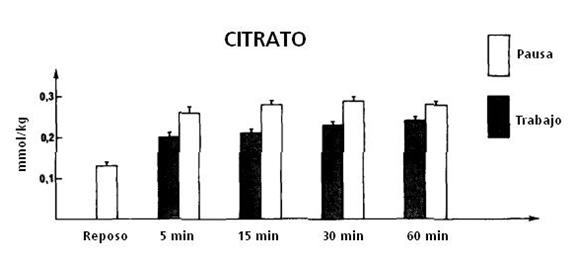
Figura 9. Concentración de citrato durante el ejercicio intermitente (15”x15”) a una intensidad de 300 W, en reposo, y después de 5’,15’,30’ y 60’ de ejercicio.
Sobre este último punto, sobre si el citrato podría ser otro factor que regula el ritmo de la glucólisis en el metabolismo muscular humano durante el ejercicio, Bangsbo indica que es poco probable que el efecto del citrato dé cuenta de todo el ‘empeoramiento’ de la glucólisis (o sea, que sea la primera y única causa) durante el ejercicio repetido de alta intensidad, o al menos, es muy poco claro ese efecto. De todas formas, los depósitos de sustratos intramusculares en la forma de glucógeno y triglicéridos disminuyen rápidamente durante la fase temprana de todos los tipos de ejercicio analizados, indicando que no sólo la glucólisis sino también la lipólisis es activada rápidamente por el ejercicio.
Otros de los estudios de esta serie de Essén
fue investigar el contenido de glucógeno según el tipo de fibras musculares
durante varios tipos de ejercicio. Allí se observó que los contenidos promedio
de glucógeno eran significativamente más altos en las fibras tipo II (402
mmol/kg) que en las tipo I (344 mmol/kg). Luego de 60’ de ejercicio de pedaleo
continuo (a 160 W, rango 138-180 W, un ![]() O2 de 50-60% del
máximo) hubo una pronunciada caída en las fibras tipo I (277 mmol/kg) que en
las fibras tipo IIa y IIb (113 mmol/kg). Con 60’ de
ejercicio de pedaleo intenso (una carga de 299 W, rango 270-343 W) en forma
intermitente (15” de trabajo x 15” de descanso pasivo, un VO2
promedio de 50-60% del máximo) ocurrió una depleción similar en ambos tipos de
fibras (tipo I, 213 mmol/kg y tipo IIa y IIb, 203
mmol/kg). Y con un ejercicio de pedaleo continuo intenso pero hasta el
agotamiento (una duración de 4’-6’, y una carga de 266 W promedio, rango
220-300 W, y un VO2 del 100%), la caída del glucógeno fue más
marcada en las fibras tipo IIa y IIb (118 mmol/kg) que en
las fibras tipo I (74 mmol/kg)(ver Figura 10).
O2 de 50-60% del
máximo) hubo una pronunciada caída en las fibras tipo I (277 mmol/kg) que en
las fibras tipo IIa y IIb (113 mmol/kg). Con 60’ de
ejercicio de pedaleo intenso (una carga de 299 W, rango 270-343 W) en forma
intermitente (15” de trabajo x 15” de descanso pasivo, un VO2
promedio de 50-60% del máximo) ocurrió una depleción similar en ambos tipos de
fibras (tipo I, 213 mmol/kg y tipo IIa y IIb, 203
mmol/kg). Y con un ejercicio de pedaleo continuo intenso pero hasta el
agotamiento (una duración de 4’-6’, y una carga de 266 W promedio, rango
220-300 W, y un VO2 del 100%), la caída del glucógeno fue más
marcada en las fibras tipo IIa y IIb (118 mmol/kg) que en
las fibras tipo I (74 mmol/kg)(ver Figura 10).
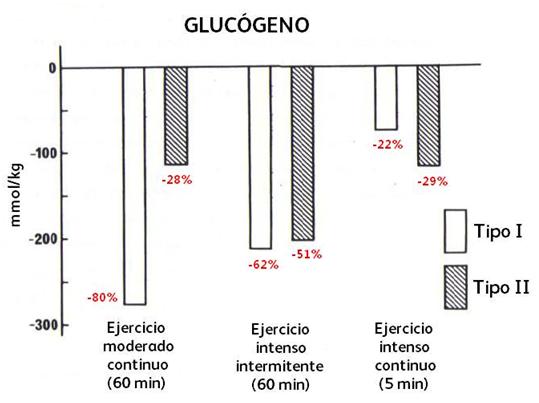
Figura 10. Depleción de glucógeno en las fibras tipo I y II después de 60 minutos de ejercicio continuo moderado y de ejercicio intermitente intenso, y después de 5’ de ejercicio intenso continuo. Los % de las caídas son respecto a los valores de reposo.
Estos datos implican que el patrón de depleción del glucógeno en las fibras musculares está determinado principalmente por la intensidad del trabajo, pero la menor depleción de glucógeno por unidad de tiempo en la modalidad intermitente comparado al ejercicio intenso continuo, indica que el modo y duración del ejercicio es importante también. Essén advirtió que esta baja depleción en el glucógeno en el ejercicio intermitente podría ser explicada por un relativo incremento en la contribución de los lípidos al metabolismo oxidativo, lo cual fue indicado por la propia Essén que es debido a los mayores niveles de ATP, CP y citrato al final de cada período de descanso, suprimiendo así a la glucólisis en la fase temprana del subsiguiente período de trabajo. También, se argumenta la disponibilidad de O2 contenido en la Mb, que se recarga fácilmente en las pausas, como ya se ha visto en los trabajos de con Ǻstrand y otros. Otra explicación de por qué las fibras tipo II utiliza más glucógeno, es probablemente debido al desarrollo rápido de tensión muscular, que ocurre al empezar cada turno de pedaleo y el hecho de que el pico de tensión da cuenta de una significativa fracción de la contracción máxima voluntaria (MCV), que no puede ser alcanzada por las fibras tipo I solamente. También se podría agregar que en el ejercicio de carrera al 70-80% del VO2máx, la depleción de glucógeno es mucha más pequeña que en el ejercicio en bicicleta a intensidades comparables. En la carrera se observa una tasa más grande de glucólisis en los músculos en las primeras fases del ejercicio. Así, como primera conclusión de ese estudio de Essén, se puede decir que la depleción de glucógeno ocurre principalmente en las fibras tipo I con el ejercicio continuo submáximo, y en todos los tres tipos de fibras (I, IIa y IIb), con el ejercicio intenso realizado intermitentemente o continuo. El efecto de ‘ahorro’ del glucógeno en el ejercicio intermitente no es, entonces, dependiente de un patrón específico de activación de las fibras, según Essén
Hay un último estudio, de esta serie, de Essén que investiga los posibles mecanismos regulatorios de la glucólisis en los músculos activos que producen una alterada relación entre la utilización de los CHO y los lípidos durante el ejercicio intermitente. En dicho estudio, 30 sujetos varones (23 años, rango 20-40 años, de edad; 180 cm, rango 174-191 cm, de altura; 74 kg de peso corporal, rango 65-87 kg; y un VO2máx de 3,83 l∙min-1, rango 2,92-4,42). El protocolo intermitente fue realizado en una bicicleta ergométrica a un promedio de carga de 284 W (rango 250-325 W), en forma 15”x15” (trabajo-pausa) durante 60’. El ejercicio continuo fue realizado hasta el agotamiento (4’-6’ de duración) a una carga de trabajo de 280 W (rango 220-320 W). Esas cargas de trabajo en ambos tipos de ejercicio fueron seleccionadas de forma tal, que la demanda aeróbica al realizarlas, el trabajo continuo se igualaba al VO2máx de cada sujeto. En las Figuras 11,12,13,14 y 15 se muestran los resultados.
Se observó que el VO2 durante el ejercicio intermitente correspondió al 55-60% del VO2máx en promedio. El cociente respiratorio estuvo en 0.88 (rango, 0.81-0.93) durante el período global del ejercicio intermitente, indicando una oxidación cubierta por el 60% por CHO y un 40% por lípidos. La FC estuvo, en promedio, a 138 latidos∙min-1 a los 15’ y aumentó a 159 latidos∙min-1 al final del ejercicio. En cambio, durante el trabajo continuo, la FC aumentó a 180 latidos∙min-1. Respecto a los sustratos y metabolitos, el ATP y la CP, durante el ejercicio intermitente al final de los períodos de trabajo en nivel de ATP fue en promedio de -10% y el nivel de CP fue de -45% que el nivel basal (reposo). Al final de un período subsecuente de pausa, tanto el ATP como el CP habían sido parcialmente restituidos, pero el ATP estaba aún casi -5% y la CP -25% por debajo del nivel de reposo. Durante el ejercicio continuo, al final del trabajo el nivel de ATP y de CP fue de -35% y -60%, respectivamente, por debajo del nivel de reposo. Una recuperación gradual ocurrió 15”-30” de recuperación tanto en el ATP (-8%) como en la CP (-45%), por debajo del nivel de reposo (ver figura 11).
El lactato se elevó un 83% respecto a los valores de reposo después de transcurridos 5’ de ejercicio intermitente, y después del período de pausa, bajó 15% respecto de aquella elevación. A los 60’ ocurrió algo similar, el lactato se elevó un 68% respecto a los valores de reposo y después del periodo de pausa correspondiente bajó un 19% respecto al aumento de los 60’. El glicerol-1-fostato, un indicador de la glucólisis e implicado en la ‘re-oxidación’ del NADH formado en la glucólisis, se comportó similarmente al lactato durante el ejercicio intermitente (ver figura 12).
Nota: Los triglicéridos séricos son hidrolizados a glicerol y ácidos grasos por efecto de la lipasa. En presencia de ATP y glicerol kinasa (GK) el glicerol es convertido a glicerol-1-fosfato. El glicerol-1-fosfato es oxidado entonces por la glicerol fosfato oxidasa para producir peróxido de hidrógeno (H2O2). Recordemos que el glicerol puede entrar a la glucólisis en el hígado o células adiposas, no en las células musculares.
Durante el ejercicio continuo, el lactato aumentó 10-11 veces inmediatamente después del ejercicio, respecto a los valores de reposo. Con 15”-20” de recuperación, estaba elevado un poco más (casi un +10%) y a los 3’ seguía elevado unas 9-10 veces por encima de los valores de reposo. Con el glicerol-1-fosfato, aumentó 5-6 veces más que en reposo al finalizar el ejercicio continuo. A los 15”-20” se mantuvo elevado un poco más y a los 3’ descendió, pero continuaba elevado 5 veces más que en reposo.
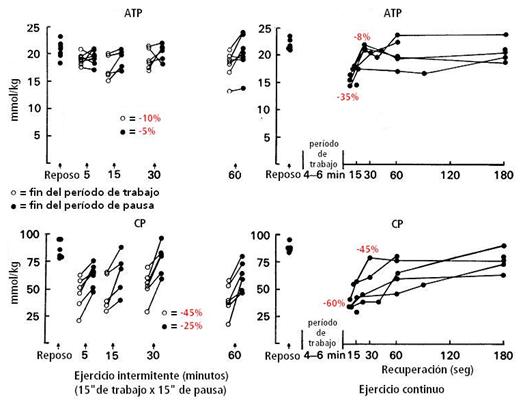
Figura 11.
Concentración muscular de ATP y CP antes (reposo) y al final de un turno de
trabajo y al final del período subsiguiente de pausa, después de 5’, 15’, 30’ y
60’ de ejercicio intermitente; y antes (reposo) y en intervalos de la
recuperación después del ejercicio continuo hasta el agotamiento. Los valores
en % son promedio de todos los períodos tomados en cuenta y el promedio de los
sujetos en su conjunto, respecto a los valores de reposo.
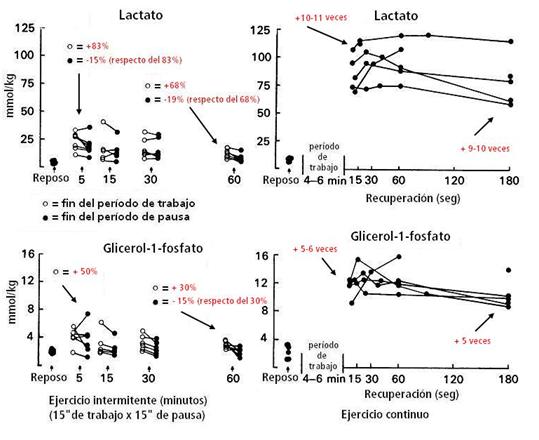
Figura 12.
Concentración muscular de lactato y glucerol-1-fosfato antes (reposo) y al
final de un turno de trabajo y al final del período subsiguiente de pausa,
después de 5’, 15’, 30’ y 60’ de ejercicio intermitente; y antes (reposo) y en
intervalos de la recuperación después del ejercicio continuo hasta el
agotamiento. Los valores en % son promedio de todos los períodos tomados en
cuenta y el promedio de los sujetos en su conjunto, respecto a los valores de
reposo, lo mismo ocurre con los valores expresados como tantas veces
aumentaron, respecto al reposo.
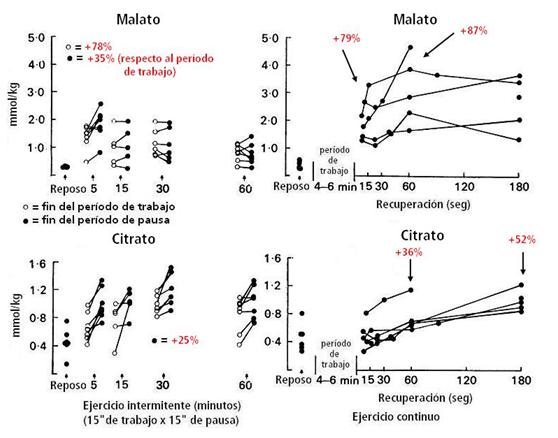
Figura 13.
Concentración muscular de malato y citrato antes (reposo) y al final de un
turno de trabajo y al final del período subsiguiente de pausa, después de 5’,
15’, 30’ y 60’ de ejercicio intermitente; y antes (reposo) y en intervalos de
la recuperación después del ejercicio continuo hasta el agotamiento. Los
valores en % son promedio de todos los períodos tomados en cuenta y el promedio
de los sujetos en su conjunto, respecto a los valores de reposo.
El citrato (indicador de oxidación de AGL, primer intermediario del ciclo de Krebs y es inhibidor de la PFK, y por tanto, de la glucólisis), durante el ejercicio intermitente, y luego del período de pausa a los 5’,15’, 30’ y 60’ se elevó un 25% (en promedio) respecto a los períodos de carga. El malato (otro intermediario del ciclo de Krebs) aumentó al principio del período de pausa un 35% comparado al período de trabajo a los 5’ de ejercicio intermitente. Después del ejercicio continuo, un aumento gradual del citrato ocurrió a 1’ (+156%) y a los 3’ (+200%) de la recuperación. El malato aumentó después del trabajo 4,7 veces más respecto al valor de reposo y aumentó aún más al 1’ (7,7 veces más respecto al reposo) y se mantuvo así en ese nivel a los 3’ de recuperación.
La glucosa-6-fosfato (un intermediario primario de la glucólisis) durante el ejercicio intermitente aumentó, luego de 5’, un 166% respecto al reposo. Luego, a los 60’, el valor fue menor y no difirió de los valores de reposo. Sin embargo, la G-6-P aumentó en los 2/3 de todos los períodos de pausa, respecto al período de trabajo. La fructosa-1,6-difosfato aumentó durante los períodos de ejercicio intermitente un promedio de 1,6 veces por encima de los valores de reposo. Pero disminuyó en promedio después de los períodos de pausa, a casi los valores de reposo. Durante el ejercicio continuo, la G-6-P aumentó 4 veces más respecto al reposo inmediatamente después de finalizado el ejercicio, y luego de 15”-20” de recuperación aumentó un poco más aún (5,2 veces más). Al 1’ de recuperación disminuyó muy poco y a los 3’ seguía elevado unas 2,8 veces respecto al reposo (pero disminuyó respecto a la finalización del ejercicio). La F-1,6-P2 aumentó 1,5 veces respecto al reposo luego de finalizar el ejercicio, pero siguió disminuyendo (-31%) aún por debajo de los valores de reposo.
Nota: La glucosa-6-fosfato (G-6-P) es una molécula de glucosa fosforilada en el carbono 6 a través de la enzima hexokinasa; esta rápida fosforilación de la glucosa tras su entrada en la célula, es para prevenir su difusión al exterior, ya que la fosforilación añade un grupo fosfato cargado (a través del consumo de un ATP) que impide que la glucosa-6-fosfato pueda atravesar la membrana plasmática. La glucosa-6-fosfato también puede ser producida durante la glucogenólisis, a partir de glucosa-1-fosfato, el primer producto generado en la hidrólisis de los polímeros de glucógeno. La fructosa-1,6-difosfato (F-1,6-P2) es una molécula de fructosa fosforilada en los carbonos 1 y 6, con el fin de impedir que pueda atravesar la membrana plasmática y difundir al medio extracelular. La conversión es de fructosa-6-fosfato a fructosa-1,6-difosfato por la enzima PFK, cuya actividad está regulada por la relación ATP/ADP.
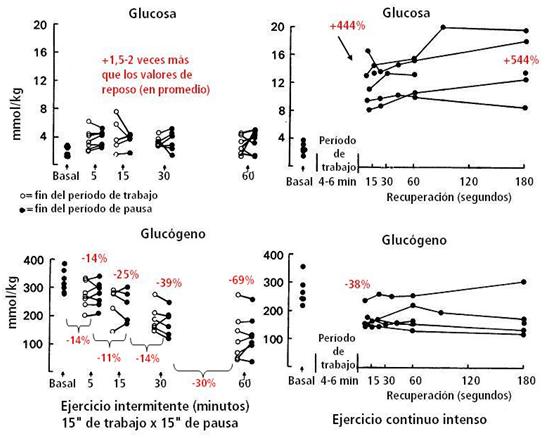
Figura 14. Concentración muscular de glucosa y glucógeno antes (reposo) y al final de un turno de trabajo y al final del período subsiguiente de pausa, después de 5’, 15’, 30’ y 60’ de ejercicio intermitente; y antes (reposo) y en intervalos de la recuperación después del ejercicio continuo hasta el agotamiento. Los valores en % son promedio de todos los períodos tomados en cuenta y el promedio de los sujetos en su conjunto, respecto a los valores de reposo. Los valores entre las llaves significan % de diferencia entre cada caída del contenido de glucógeno entre los momentos tomados como referencia.
El glucógeno durante el ejercicio intermitente disminuyó a lo largo de todo el ejercicio, siendo algo más rápido al inicio (14%) y luego siguió disminuyendo (69%) hasta el final del ejercicio. La glucosa aumentó 1,5-2 veces por arriba los valores de reposo y en ambos casos, con la glucosa y el glucógeno, no hubo diferencias significativas entre los períodos de trabajo y pausa (ver Figura 14). Durante el ejercicio continuo disminuyó (-38%) la cantidad del glucógeno al finalizar el ejercicio y no cambió durante la recuperación. La glucosa aumentó (4,5 veces respecto al reposo) al finalizar el ejercicio y durante la recuperación del ejercicio se mantuvo aún alta (5 veces el valor de reposo).
A la luz se los resultados de este estudio, la interpretación de Essén es que durante el ejercicio intermitente hubo una menor tasa de degradación de la glucólisis (esto es, menor utilización de glucógeno y, por tanto, menor formación de lactato), además de menor cantidad de intermediarios de la glucólisis, aún cuando el ejercicio fue realizado a alta intensidad (es decir, ante igual carga, el ejercicio intermitente comparado al ejercicio continuo, utiliza menos glucógeno y forma menos lactato). Esto pudo deberse o a una menor utilización de CHO o a una mayor utilización de lípidos. El metabolismo aeróbico fue superior durante los períodos de pausa, por el aumento de sustratos que indican una mayor utilización de lípidos (ATP, CP y citrato, por ejemplo) y que inhiben a la glucólisis. Cabe aclarar, que el intercambio respiratorio ante altas cargas de trabajo, no refleja lo que ocurre en el músculo fehacientemente. También se concluye, finalmente, que durante el ejercicio intermitente la existencia de factores regulatorios a través de la activación e inhibición de diferentes enzimas y de diferentes concentraciones de sustratos, los cuales en forma combinada influencian y retardan la glucólisis en las fases finales de los períodos de pausa (teniendo un corte netamente aeróbico dichas pausas) y al comienzo del período subsiguiente de trabajo, aún con una carga alta de ejercicio.
Por lo tanto, el ejercicio intermitente en estos estudios de Essén resultó en una menor depleción de glucógeno y en una mayor depleción de los triglicéridos intramusculares comparado al esfuerzo continuo de baja intensidad. Esto es consecuencia de una mayor estimulación de la respiración mitocondrial en presencia de ácidos grasos. También se debe tener en cuenta que la eficiencia oxidativa del músculo, puede explicarse por un cambio en la permeabilidad de la membrana interior mitocondrial debido a cambios en la composición de los fosfolípidos, que puede lograrse por el entrenamiento aeróbico de larga duración; por lo cual, el estatus de entrenamiento está más relacionado a esta capacidad oxidativa muscular que la composición del tipo de fibra o de la densidad mitocondrial como antes se pensaba. Otro factor a tener en cuenta en estos estudios de Essén es el % de VO2 utilizado (alrededor el 55-60% en promedio), que bien es sabido que es una carga mayormente aeróbica con base en la oxidación de lípidos. En estudios, en décadas posteriores, veremos porcentajes mayores utilizados. Además, en esos estudios de Essén tampoco se analizaron concentraciones de hormonas, que quizás podrían haber dado más información acerca del comportamiento de las catecolaminas, por ejemplo.
Debemos aclarar también, que a pesar de la antigüedad de estos trabajos de Essén, se resalta el alto valor científico de esas investigaciones que fueron, y aún hoy se mantienen, como las investigaciones predominantes acerca del uso del combustible muscular bajo las condiciones del ejercicio intermitente. Pero no olvidemos que también existen algunas interpretaciones actuales que contradicen estos estudios y que analizaremos más adelante, entre ellos, los estudios de de Christmass M.A. de 1999.
Bibliografía
1.Essen B., Hagenfeldt L., Kaijser L. Utilization of blood-borne and intramuscular substrates during continuous and intermittent exercise in man. J Physiol 265: 489–506, 1977.
2.Edwards, R.H.T., Ekelund L.G., Harris R.C, Hesser C.M., Hultman E, Melcher A., Wigertz O. Cardiorespiratory and metabolic costs of continuous and intermittent exercise in man. J Physiol (Lond) 234: 481-497, 1973.
3.Essen, B. Intramuscular substrate utilization during prolonged exercise. Ann NY Acad Sci 301: 30-44, 1977.
4.Essen B, Kaijser L. Regulation of glycolysis in intermittent exercise in man. J Physiol(Lond) Aug; 281: 499-511. 1978.
5.Essen, B. Studies on the regulation of metabolism in human skeletal muscle using intermittent exercise as an experimental model. Acta Physiol Scand Suppl. 454: 1-32. 1978.
6.Essen, B. Glycogen depletion of different types in human skeletal muscle during intermittent and continuous exercise. Acta Physiol Scand 1978; 103:446.
7.Plowman, S.A.; Smith, D.L Exercise physiology for health, fitness, and performance; 2nd ed.; San Francisco: Benjamin Cummings, Pearson Education, Inc . 2003.
8.Christmass M.A., Dawson B., Passeretto P., Arthur P.G. A comparison of skeletal muscle oxygenation and fuel use in sustained continuous and intermittent exercise. Eur J Appl Physiol Occup Physiol, Oct 1999; 80(5): 423-35.
9.Christmass M.A., Dawson B., Arthur P.G. Effect of work and recovery duration on skeletal muscle oxygenation and fuel use during sustained intermittent exercise. Eur J Appl Physiol Occup Physiol, Oct 1999; 80(5): 436-47.
10.Bangsbo J. Physiology of intermittent exercise. Chapter 5, pp 53-65. Exercise and Sport Science. Garrett W., Kirkendall D. Edit. Lippincott Williams & Wilkins (2000).
11.Bangsbo J., Krustrup P., González-Alonso J., Saltin B. ATP production and efficiency of human skeletal muscle during intense exercise: effect of previous exercise. Am J Physiol Endocrinol Metab, Jun 2001; 280: 956 - 964.
12.Christmass M.A., Dawson B., Arthur P.G. Goodman C. Brief intense exercise followed by passive recovery modifies the pattern of fuel use in humans during subsequent sustained intermittent exercise. Acta Physiol Scand 2001;172: 39-52.