
Aumento de la Fuerza mediante el Estiramiento.
Force Enhancement by Stretch
Artículo publicado en el journal Revista de Entrenamiento Deportivo, Volumen 26, Número 4 del año 2012.
Publicado 15 de diciembre de 2012
Resumen
Palabras clave: fuerza, estiramiento
Abstract
Keywords: strength, elongation
Ingen Schenau, Bobbert y Haan concluyen que el almacenaje y la reutilización de la energía elástica no tiene mayor importancia para el desarrollo del rendimiento mecánico aumentado de los músculos del cuerpo después del preestiramiento. Los autores también afirman que la “potenciación” del mecanismo contráctil después de un preestiramiento, probablemente no aumenta el rendimiento muscular bajo condiciones “in vivo”. Para ampliar esta parte de la discusión, puede ser útil considerar algunos asuntos relevantes del aumento de la fuerza durante y después del estiramiento explorados en un músculo aislado. Las investigaciones aludidas en esta investigación se llevaron a cabo en fibras musculares únicas de una rana a temperaturas bajas (1-3 º C). La fibra muscular de la rana se adapta bien a este género de investigación debido a su variabilidad cuando se estudia al exterior del cuerpo en un baño del órgano. Existe toda la razón para creer que los mecanismos que subyacen al aumento de la fuerza mediante el estiramiento en los músculos anfibios, también se pueden aplicar al músculo esquelético de un mamífero.
En esta discusión, es esencial distinguir entre las dos principales componentes del aumento de la fuerza inducida por estiramiento (ver Figura 1). La primera es un aumento de la velocidad-dependiente en la fuerza, la cual rápidamente se desarrolla por entero después del comienzo del estiramiento y permanece a través del periodo de estiramiento. Esta componente desaparece gradualmente en 4 ó 5 segundos, después del final del estiramiento a la baja temperatura ya estudiada. La segunda es un aumento de la velocidad-independiente en la fuerza que persiste por tan largo periodo como permanezca activado el músculo después de la rampa del estiramiento. Esta componente del aumento de la fuerza no aparece cuando el estiramiento se lleva a cabo alrededor de la longitud óptima del sarcómero (Figura 1A), pero destaca a mayores longitudes (Figura 1B). Anteriormente nos hemos referido a esto como “el aumento de la fuerza residual después del estiramiento” (Edman, Elzinga, y Noble, 1978).
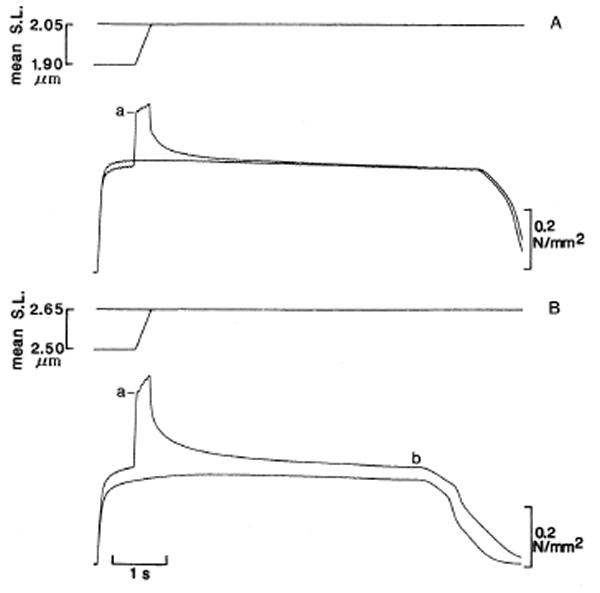
Figura 1. Registros del desplazamiento y de la fuerza de una única fibra muscular de una rana durante el tétanos en dos longitudes de sarcómero diferentes. (A) Estiramiento durante la actividad de 1.90 a 2.05 μm de la longitud del sarcómero comparado con un tétanos isométrico ordinario a 2.05 μm. (B) Comparación del estiramiento de 2.50 a 2.65 11m de longitud del sarcómero con un tétanos isómétrico a 2.65 μm. a denota el aumento de la fuerza de la velocidad-dependiente durante el estiramiento, mientras que b indica el aumento de la fuerza residual después del estiramiento; el último sólo aparece por encima de una longitud óptima del sarcómero. Tomado de “Strain of passive elements during force enhacement by strech in frog muscle fibers”, por K.A.P. Edman & T. Tsuchiya, 1996. Journal of Physiology, 490, p. 193.
La componente de velocidad-dependiente (a en la Figura 1) se recupera plenamente mediante una rampa de estiramiento que solamente es la 12-14 nm/mitad del sarcómero en amplitud, suficiente para alargar los puentes-cruzados más allá del punto crítico donde son forzados a separarse para permitir que los filamentos delgados se deslicen. La fuerza aumentada puede ser considerada como una fuerza de fricción en tanto que los puentes de miosina continuamente se separan y se juntan cuando son arrastrados a lo largo del filamento delgado. La fuerza alcanzada durante el estiramiento está cercana al doble de la fuerza producida durante la fase de preestiramiento isométrico.
Se podría pensar que el alto nivel de fuerza alcanzado durante el estiramiento (componente a) aumentaría la capacidad del músculo para producir trabajo positivo cuando al músculo se le deja acortarse después del estiramiento. Sin embargo, la energía elástica almacenada en los puentes-cruzados alargados es completamente descargada por un pequeño movimiento. Un acortamiento de menos de 0.50 % de la longitud del músculo (3-4 nm/mitad del sarcómero; Piazzesi, Francini, Linari y Lombardi, 1992) es, así, suficiente para restaurar la fuerza hasta un nivel isométrico de control. La situación no sería fundamentalmente diferente en el músculo actuando “in situ” en el cuerpo, debido a la alta (no lineal) rigidez de los tendones en el intervalo alto de fuerza (ver Figura 2 en Cleworth y Edman, 1972). La rápida desaparición de la fuerza aumentada por un pequeño escape, es suficiente evidencia contra la idea (Cavagna, Heglund, Harry y Mantovani, 1994) de que un estiramiento antecedente lleva al almacenamiento de la energía mecánica en un “elemento debilitado” dentro de los puentes-cruzados (para una mayor discusión, ver Edman y Tsuchiya, 1996). Es posible concluir que la fuerza extra sostenida por los puentes-cruzados durante el estiramiento (componente a) no constituye un recurso que pudiera aumentar notablemente el output de trabajo del músculo durante una fase de acortamiento siguiente.
La naturaleza de la segunda componente de larga duración del aumento de la fuerza por estiramiento (el aumento de la fuerza residual [b], Figura 1B) ha sido recientemente aclarada (ver Edman y Tsuchiya, 1966, para más referencias). Es esencial notar que el aumento de la fuerza residual después del estiramiento, sólo aparece en tensión. La causa principal de este fenómeno parece ser la no uniforme distribución del cambio de longitud dentro de las miofibrillas cuando el músculo se estira durante la actividad. El cambio de longitud no uniforme conduce a diferencias en el solapamiento del filamento a lo largo de las miofibrillas con diferentes grados de escalonamiento de los filamentos gruesos dentro de los sarcómeros de las miofibrillas (Brown y Hill, 1991; Edman y Lou, 1991; Talbot y Morgan, 1996). Las regiones con un solapamiento del filamento mayor y menor, relativo al que se da antes del estiramiento, serán creadas de este modo, esparcidas a lo largo de las miofibrillas. Las regiones “débiles”, es decir, las partes que han adquirido un menor solapamiento por el estiramiento, serán ayudadas por los elementos elásticos en paralelo que son recuperados por la tensión de los componentes elásticos (tales como titina; ver Wang y Ramírez-Mitchell, 1993) dentro de la matriz del sarcómero. Las regiones debilitadas pueden, de ese modo, ser completadas apropiadamente con una fuerza elástica en paralelo que las capacite para igualar a las partes “más fuertes” en serie. Ya que los elementos coadyuvantes actúan en paralelo con las unidades contráctiles, tendrían disminuidas las propiedades elásticas. Para mayor información concerniente a la naturaleza de los elementos elásticos y su relación con el aumento de la fuerza residual después del estiramiento, ver Edman y Tsuchiya (1996).
Las regiones más fuertes, las cuales han conseguido una mayor cantidad de solapamiento del filamento, tenderán a aumentar el output de la fuerza del músculo por encima del nivel de control, siempre que el estiramiento se realice por encima de la longitud óptima del sarcómero. El no aumento de la fuerza residual después del estiramiento se puede esperar a una longitud óptima del sarcómero según el mecanismo propuesto, ya que la fuerza de control isométrica aquí representa el máximo de lo que el sistema contráctil es capaz. Numerosos resultados experimentales avalan este punto de vista (Edman y otros, 1978; Edman, Elzinga y Noble, 1982; Julián y Morgan, 1979).
El aumento de la fuerza residual después del estiramiento parece representar un mecanismo potencial mediante el cual el rendimiento del músculo esquelético se puede aumentar durante el ciclo estiramiento-acortamiento. Sin embargo, la magnitud de este efecto de potenciación dependerá de la longitud del músculo al final del estiramiento. La potenciación contráctil puede ser, de este modo, importante en longitudes musculares grandemente alargadas. No obstante, se puede esperar una no potenciación (o una potenciación menor) en la longitud normal de descanso en el cuerpo, donde el espaciamiento del sarcómero es óptimo para la producción de fuerza (ejemplo, James y otros, 1995; Ledvina y Segal, 1995).
En resumen, la evidencia obtenida de las preparaciones del músculo aislado indica que el estiramiento durante la actividad no crea un almacén de energía mecánica (en los puentes-cruzados o en otra parte) que pudiera aumentar notablemente el output de trabajo durante el acortamiento consiguiente. Sin embargo, “el aumento de la fuerza residual después del estiramiento” observado en el músculo aislado no contiene un mecanismo por el cual el rendimiento mecánico de los músculos del cuerpo pueda ser aumentado durante un ciclo de estiramiento-acortamiento.
Referencias
1. Brown, L.M., y Hill, L. (1991). Some observations on variations in fila¬ment overlap in tetanized muscle fibres and fibres stretched during a tetanus, detected in the electron microscope after rapid fixation. . Journal of Muscle Research and Cell Motility, 12, 171-182.
2. Cavagna, G.A., Heglund, N.C., Harry, J.D., y Mantovani, M. (1994). Storage and release of mechanical energy by contracting frog muscle fibres. . Journal of Physiology, 481, 689-708.
3. Cleworth, D.R., y Edman, K.A.P. (1972). Changes in sarcomere length during isometric tension development in frog skeletal muscle. . Journal of Physiology, 227, 1-17.
4. Edman, K.A.P., Elzinga, G., y Noble, M.I.M (1978). Enhancement of mechanical performance by stretch during tetanic contractions of ver¬tebrate skeletal muscle fibres. . Journal of Physiology, 281, b139-155.
5. Edman, K.A.P., Elzinga, G., y Noble, M.I.M. (1982). Residual force enhancement after stretch of contracting frog single muscle fibers. . Journal of General Physiology, 80, 769-784.
6. Edman, K.A.P., y Lou, F. (1991). Unpublished observation of muscle fibres fixed by rapid freezing. .
7. Edman, K.A.P., y Tsuchiya, T. (1996). Strain of passive elements during force enhancement by stretch in frog muscle fibres. . Journal of Physiology, 490, 191-205.
8. Ingen Schenau, G.J. van, Bobbert, M.F., y Haan, A. de. (1997). Does elastic energy enhance work and efficiency in the stretch-shortening cycle? . Journal of Applied Biomechanics, 13, 389-415.
9. James, R.S., Altringham, J.D., y Goldspink, D.F, (1995). The mechanical properties of fast and slow skeletal muscles of the mouse in relation to their locomotory function. . Journal of Experimental Biology, 198, 491-502.
10. Julian, F.J., y Morgan, D.L. (1979). effect on tension of non-uniform distribution of length changes applied to frog muscle fibres. . Journal of Physiology, 293, 379-392.
11. Ledvina, M.A., y Segal, S.S. (1995). Sarcomere length and capillary curvature of rat hindlimb muscles in vivo. . Journal of Applied Physiology, 78, 2047-205 1.
12. Piazzesi, G., Francini, F., Linari M., y Lombardi, V. (1992). Tension transients during steady lengthening of tetanized muscle fibres of the frog. . Journal of Physiology, 445, 659-71 1.
13. Talbot, J.A., y Morgan, D.L. (1996). Quantitative analysis of sarcomere non-uniformities in active muscle following a stretch. . Journal of Muscle Research and Cell Motility, 17, 261-268.
14. Wang, K., y Ramírez-Mitchell, R. (1983). A network of transverse and longitudinal intermediate filaments is associated with sarcomeres of adult vertebrate skeletal muscle. . Journal of Cell Biology, 96, 562-570.