
Cómo influye el ciclo menstrual sobre índices del daño muscular inducido por la contracción (CIMD)
Publicado 4 de mayo de 2015, 14:05

Cuando hacemos un ejercicio no habitual o un ejercicio que involucra un componente excéntrico pesado, dichos ejercicio se los asocia con el daño muscular inducido por la contracción (CIMD, contraction-induced muscle damage). El dolor y el daño asociados con este fenómeno son temporales, pero no obstante son también poco agradables. Otros efectos colaterales transitorios del daño muscular pueden incluir dolor, pérdida del rango de movimiento, y reducción en la fuerza (Brentano, 2011). Mientras el CIMD puede ser el resultado de una variedad de actividades, en un laboratorio, por ejemplo, es común utilizar contracciones excéntricas para inducir el CIMD. Sin embargo, incluso en una situación de laboratorio controlada, pueden aparecer diferencias en la respuesta de los participantes al CIMD por una variedad de razones, incluyendo algún reciente daño del propio músculo, nivel de actividad, tipo de actividad, y condición de salud. También se ha propuesto que, en el caso de las mujeres, el estrógeno puede ejercer un efecto protector sobre el músculo (Feng, 2004). Aunque algunos investigadores no han encontrado una diferencia relacionada con el sexo en el daño muscular (Hubal, 2008; Rinard, 2000), varios estudios recientes (Sayers, 2001;Tiidus, 2003) hacen pensar en una diferencia del sexo en respuesta a la carga excéntrica, de tal forma que las mujeres pueden experimentar menos daño muscular que los hombres. Si la diferencia del sexo es debida a las mujeres que tienen una concentración superior de estrógeno circulante, entonces las diferencias en respuesta a la carga excéntrica muscular pueden existir dentro de las fluctuaciones hormonales normales del ciclo menstrual de una mujer.
El daño muscular tiene muchas medidas subjetivas, pero una elevación de la creatina kinasa en suero (CK) se usa a menudo como indicador objetivo del daño muscular. La creatina kinasa está presente en las células musculares y una presencia elevada en el suero después del ejercicio indica daño muscular (Peñaililli, 2013). El daño muscular, independientemente de la causalidad, producirá una pérdida de la integridad de la membrana como es reflejado por la CK libre de las células. La concentración de la creatina kinasa no puede cuantificar el daño muscular pero sí puede indicar su presencia (Evans, 1991).
El estrógeno puede conferir protección contra el CIMD a través de 2 mecanismos propuestos: sosteniendo la integridad de las membranas celulares y protegiendo la célula contra el ataque de los radicales libres a través de las propiedades antioxidantes (Dykens, 2003; Wang, 2003). Por ejemplo, un estudio 'in vitro' de mioblastos encontraron que la presencia de estrógeno promueve un mejor mantenimiento de los niveles básicos de ATP (Tiidus, 2003). Esto es importante porque una mejor conservación de la homeostasis del ATP compensaría el daño que indujo un aumento de iones intracelulares de calcio, lo cual precipita eventos que llevan a la acción del radical libre sobre la célula.
Ayres y cols. (1998) dirigieron algunos experimentos
'in vitro' para examinar el rol protector del estrógeno contra los radicales
libres y otros oxidantes. La presencia de estrógeno significativamente inhibió
la oxidación de la lipoproteína de baja densidad y protegió contra el hidrógeno
el daño del ácido desoxirribonucleico inducido por el peróxido. Cuando el
estrógeno se combinó con otros residuos conocidos de los radicales libres, la
protección no fue magnificada. En base a los resultados de éstos y otros
estudios (Claassen, 2005), se ha supuesto que el estrógeno ayuda a estabilizar
las membranas celulares. Stupka y cols. (2000) investigaron las diferencias del
daño muscular entre los sexos en respuesta al ejercicio realizado a una misma
intensidad relativa (Figura 1). Tendencias estadísticas existieron para las diferencias
del sexo en las concentraciones de la CK y en las concentraciones de leucocitos
que, cuando están elevadas, es indicativo de una respuesta inflamatoria en el
tejido dañado. Tanto las concentraciones de CK como de leucocitos tendieron a
ser superiores en los hombres. Adicionalmente, sólo los hombres tenían una
elevación significativa en las concentraciones de granulocitos circulantes, otra
medida de la respuesta inflamatoria.
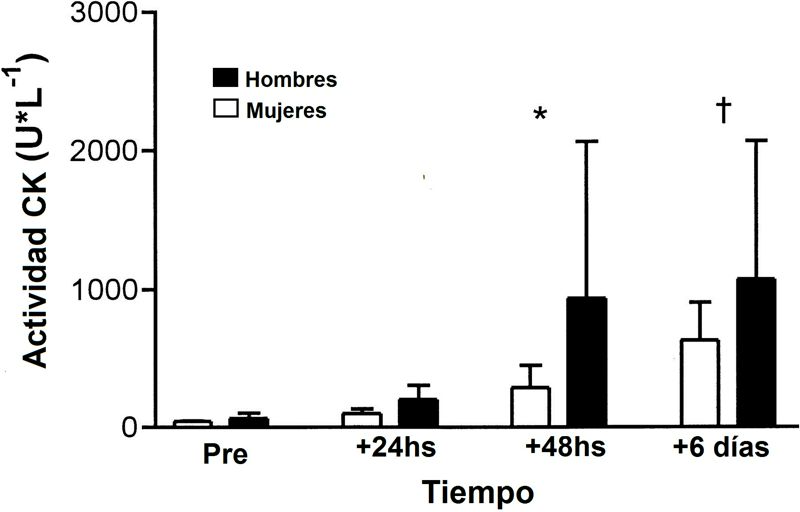
Figura 1. La actividad de la creatina kinasa en plasma (CK) fue significativa a las 24 hs (+24h), 48 hs (+48h), y 6 días (+6 días) post-ejercicio comparado con la línea de base (pre) (*,† P<0.05); no hubo ninguna diferencia sexual significativa (P = 0.14). Los valores son promedios ±SD. Stupka y cols. (2000).
Porque las mujeres tienen concentraciones de estrógenos circulantes significativamente mayores que los hombres, los resultados de este estudio apoyan la hipótesis que el estrógeno puede conferir una ventaja protectora contra el trabajo excéntrico muscular. Estas tendencias relacionadas con el sexo son poco probablemente relacionadas a las diferencias en la masa muscular total, que es a menudo mayor en los hombres. Graves y cols. (1987) reportaron que aunque los hombres generalmente tienen una masa muscular mayor que las mujeres, el volumen de la masa muscular activa no parece influir sobre la magnitud del cambio en la CK en respuesta al daño muscular. En el estudio de Graves y cols. (1987), un diseño equilibrado fue usado para administrar el daño a 1 brazo, ambos brazos, o 1 pierna. Ninguna diferencia significativa en la CK circulante fue observada — concluyendo así que la elevación en la CK es independiente del área muscular implicada.
Aunque estudios que usan a animales y hombres son importantes, ellos no pueden ser representativos de la respuesta en las mujeres humanas. Hay evidencia para indicar que el estrógeno influye en marcadores del CIMD; sin embargo, es desconocido si acciones musculares excéntricas, cuando son realizadas durante los períodos de estrógeno alto vs estrógeno bajo, producirían diferencias perceptibles en los índices del daño muscular.
Pero existe un estudio reciente de Melissa M. Markofski y cols. (2014), que examinaron si los síntomas del daño muscular difieren cuando la carga excéntrica se administra durante la fase folicular vs la fase lútea del ciclo menstrual. La fase lútea está asociada con cantidades superiores de estrógeno que la fase folicular. Si el estrógeno confiere un rol protector, entonces los síntomas del daño muscular pueden diferir entre las 2 fases, con el grupo de la fase lútea experimentando indicios y síntomas menos severos del daño muscular.
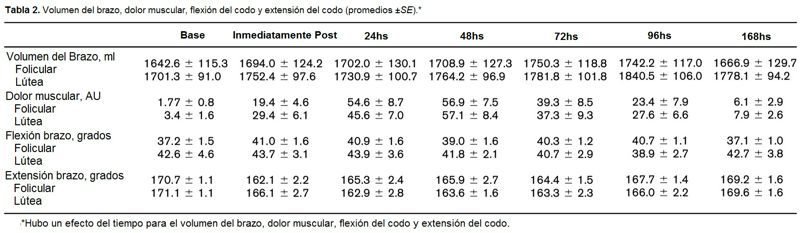
Para ello, realizaron un diseño de investigación de medidas repetidas (tiempo x fase) para determinar si había diferencias fase-dependientes del ciclo menstrual en la respuesta y recuperación al CIMD. El dolor, volumen del brazo, extensión del codo, flexión del codo, y el ángulo articular del codo relajado, fueron medidos antes del ejercicio excéntrico e inmediatamente, 24, 48, 72, 96, y 162 horas después del ejercicio. La creatina kinasa fue medida antes del ejercicio excéntrico y 48, 72, 96, y 162 horas después del ejercicio. El estradiol fue medido para confirmar la fase del ciclo menstrual (Tabla 1). Dieciocho mujeres entrenadas en fuerza que menstruaban regularmente entre edades de 18–38 años fueron reclutadas para este estudio. Todas las mujeres reportaron un ciclo menstrual regular al menos de entre 21 y 35 días para los 12 meses precedentes.
Aunque la duración del ciclo era inconstante entre las mujeres, dentro de cada mujer participante que reportaba una duración del ciclo consistente, esto se usó como criterio para la inclusión. Ocho participantes eran usuarias de píldoras anticonceptiva s (PA) y las restantes 10 no lo eran (sin-PA). Fueron empleadas participantes entrenadas en fuerza en este estudio ya que ellas tendrían una experiencia previa con el dolor muscular y molestia y, por lo tanto, tolerarían mejor la magnitud del daño muscular. Las voluntarias tenían que someterse en un mínimo de 2 sesiones de entrenamiento de la fuerza (que incluían ejercicios de tren superior) por semana para un mínimo de 3 meses continuos, precedentes al estudio y se les pidió abstenerse del ejercicio de fuerza mientras eran sometidas a la evaluación. Todas las participantes se evaluaron dentro de un período de 2 meses. Para minimizar la variabilidad, cada participante se reportó al laboratorio en el mismo momento del día. Las participantes aleatorizadas al grupo folicular fueron testeadas en día 2 o 3 de la fase folicular, y las participantes en la fase lútea en día 1 o 2 de la fase lútea. La aleatoriedad en los grupos de la fase lútea o folicular se realizó en cierto modo que ayudó a asegurar una distribución igual de usuarias de PA y usuarias no-PA. Se describen las características de las participantes en la Tabla 1.
Al menos 4 días antes del turno de ejercicio excéntrico, las participantes se reportaron al laboratorio para un test de 1 máxima repetición (1RM) de curl de bíceps en el brazo no dominante. Para ayudar a mantener los mismos ángulos articulares para todas las pruebas, un banco 'predicador' estándar (Life Fitness, Inc., Schiller Park, IL, USA) se usó para el test de 1RM, turno de ejercicio excéntrico, evaluación del dolor activo, y las siguientes valoraciones de fuerza.
En el día de la sesión del ejercicio excéntrico, las mediciones básicas eran inmediatamente reunidas antes del ejercicio. Estas mediciones incluían la determinación del dolor básico, el volumen del brazo, una valoración de la fuerza (descrita antes), y ángulo articular del codo relajado y la flexión pasiva. En suma, una muestra de 5 ml de sangre venosa fue reunida de una vena antecubital para la determinación más tarde de la concentración de la CK y la concentración del estrógeno. Las participantes se abstuvieron de tomar NSAIDs (drogas antiinflamatorias no esteroideas); realizar cualquier ejercicio de fuerza de tren superior o tren inferior de alta intensidad o carrera en pendiente; y realizar enfriamiento, calentamiento, estiramiento, o masaje al brazo para la duración del estudio.
La tasa de dolor activa se realizó aproximadamente con un peso correspondiente al 40% de la 1RM de la participante. Después de realizar el ejercicio, ellas indicaban en una escala analógica visual que cuantificaba que tan doloroso eran las fases concéntricas y excéntricas del levantamiento. El volumen del brazo era medido por volumetría de agua que es un método establecido para el volumen e hinchazón del brazo (Taylor, 2006). Brevemente, la participante sumergiría su brazo en un tubo de plástico autoestable lleno de agua y el volumen de agua cambiado de sitio era medido. La medición era dos veces repetida, con una tercera prueba si las mediciones variaban por más de 20 ml. El ángulo articular del codo relajado y la flexión pasiva eran medidas con un goniómetro. El turno del ejercicio excéntrico consistió en 5 series de 15 repeticiones cada una, con una pausa de 2 minutos entre cada serie. Ya que el turno de ejercicio excéntrico, la participante realizaba un precalentamiento ligero de 2 series, la primera serie era de 6–8 repeticiones al 30% de 1RM, seguido por un segundo precalentamiento más pesado (4–6 repeticiones al 60% 1RM). La primer serie de la acción muscular excéntrica correspondió al 140% de la 1RM de la participante. La participante se sentó en el banco predicador de curl de bíceps para realizar el ejercicio. Un investigador ayudaba a la participante a levantar el peso pero no ayudaba en el trabajo excéntrico. Un metrónomo fue usado para poner el ritmo de repetición de 4 segundos de la acción muscular excéntrica y de 2 segundos para la contracción concéntrica con ayuda. Una vez que la participante no era capaz de sostener una acción muscular excéntrica de 4 segundos en forma controlada, el peso se reducía al 130% de 1RM. Una vez que ocurría el fallo, la carga se reducía al 120% de 1RM. Todas las participantes pudieron completar todas las series al menos con un peso correspondiente al 120% de su 1RM. La reducción del peso aseguró que todas las participantes pudieran completar 75 contracciones. Todas las mediciones de fuerza concéntrica se dirigieron después que el ejercicio excéntrico se evaluaba como porcentaje de la 1RM inicial. El primer intento usó una masa que era aproximadamente 40% de la 1RM del participante. Después de un período de descanso de 2 minutos, el próximo intento estaba en el 50% de 1RM y continuó subiendo en incrementos de 10% hasta que la participante no era capaz de levantar el peso. Las muestras de sangre fueron tomadas de una vena antecubital del brazo no dañado. El estradiol básico fue medido no sólo para cuantificar el estradiol mismo sino para confirma la fase. También fue medida la creatina kinasa. En suma a la muestra básica, la sangre fue tomada 48, 72, 96, y 168 horas después del protocolo excéntrico. Todas las otras mediciones fueron repetidas inmediatamente después del turno del ejercicio excéntrico y a las 24, 48, 72, 96, y 168 hs post-ejercicio.
Los Resultados
Durante el análisis estadístico, 2 participantes debieron ser removidas por datos incompletos durante este análisis, (1 sin complacencia; 1 enfermedad no relacionada), siendo el tamaño de la muestra finalmente de 16 participantes (n=9 folicular; lútea n=7) y 6 de estas participantes eran usuarias de PA (folicular = 4; lútea = 2). No hubo ninguna diferencia estadísticamente significativa entre los grupos folicular y lútea para cualquier variable, excepto el estrógeno.
Concentraciones de estrógeno
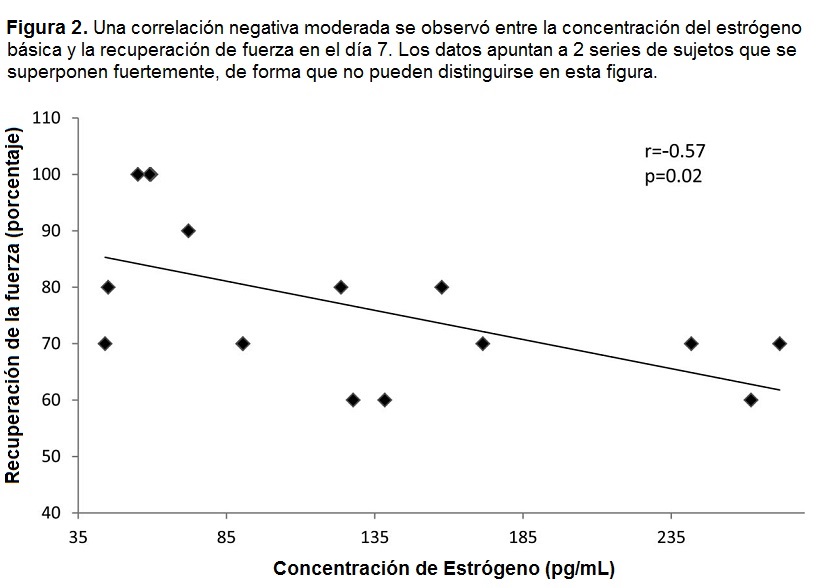
El grupo de la fase lútea tuvo una concentración de estrógeno significativamente (p = 0.0008) más alta que el grupo de la fase folicular (187.8±27.8 vs 77.3±10.7) (Tabla 1). Una correlación negativa moderada estuvo presente entre la concentración de estrógeno básica y la recuperación de fuerza, calculada por el porcentaje de 1RM que la participante pudo realizar (Figura 2).
La Valoración de la fuerza
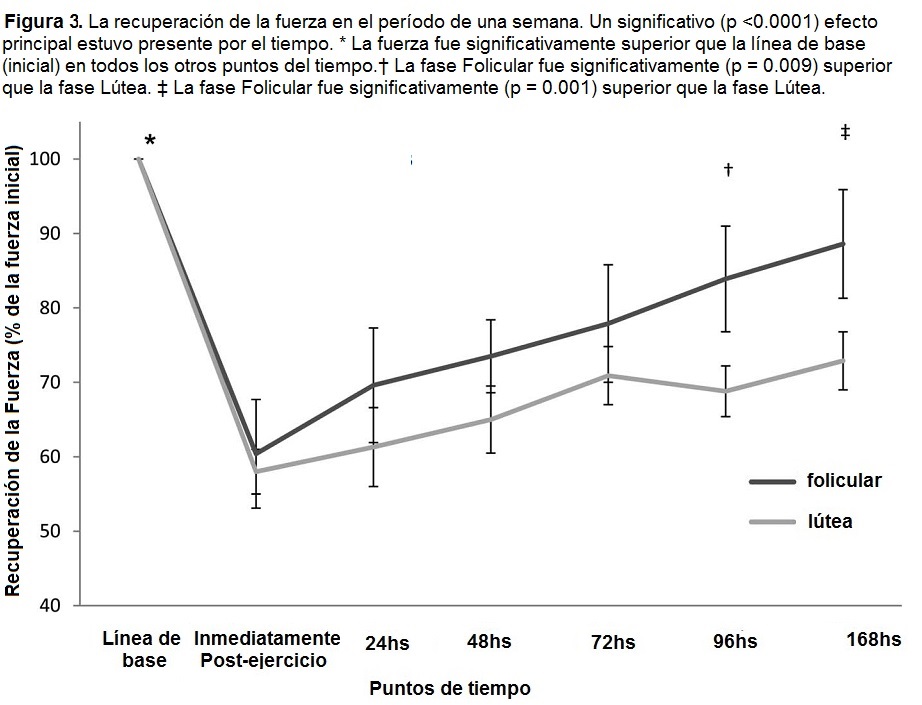
La fuerza se evaluó como porcentaje de cambio a partir de la 1RM inicial. Esto permitió que la fuerza de inicio (de base) sea tenida en cuenta sin tener que usar la línea de base como covariante. Un efecto significativo (p >0.0001) principal estuvo presente durante los puntos de tiempo. La fuerza se recuperó en el tiempo, pero la recuperación de la fuerza completa no ocurrió durante el curso de una semana del estudio (Figura 3). Una interacción significativa (p = 0.04) estuvo presente, en el que el grupo de la fase folicular tuvo la mayor recuperación de la fuerza a las 96 horas después (p = 0.009) y a las 168 horas después (p = 0.0013), que el grupo de la fase lútea.
Turno de ejercicio del daño muscular inducido por la contracción
La carga excéntrica y el número de repeticiones 'excéntricas' realizadas a cada porcentaje de 1RM (140%, 130%, 120%) no difirieron estadísticamente entre los grupos de participantes.
La flexión y extensión del codo
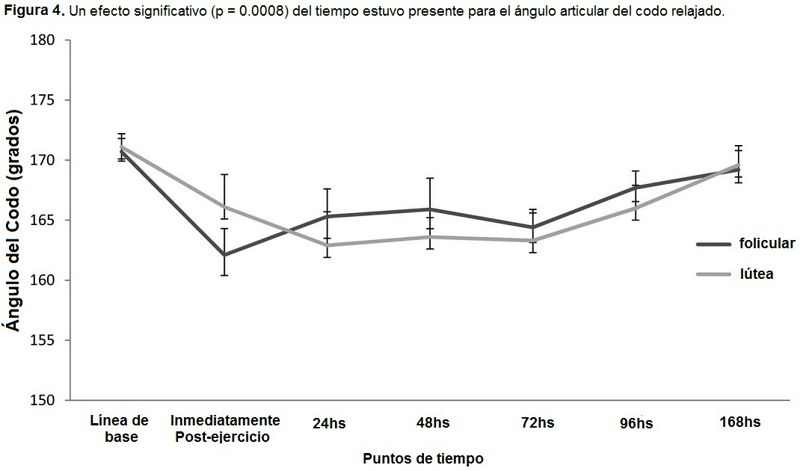
Un efecto del tiempo significativo hubo para el ángulo articular del codo relajado (Figura 4). Un efecto principal para el tiempo (p = 0.04) estuvo presente para la flexión del codo (Tabla 2). No se observaron otros efectos o interacciones. Un efecto principal para el tiempo (p = 0.0002) estuvo presente para la extensión del codo (Tabla 2). No se observaron otros efectos o interacciones.
Análisis de los resultados
Los efectos de tiempo significativos para la flexión y extensión del codo, el volumen del brazo, dolor muscular, y CK estuvieron presentes, indicando que el protocolo del estudio de de Markofski (2014) produjo cambios en estos marcadores comunes del daño muscular. Sin embargo, una concentración mayor de estrógeno no fue asociada con una mejora significativa en los indicadores y síntomas del CIMD. Contrariamente a la hipótesis propuesta por esta autora, las concentraciones fisiológicas de estrógeno de las mujeres que participaron de ese estudio, no se relacionaron a un efecto protector para las variables medidas. En base a los resultados vistos, probablemente parece que la fase del ciclo menstrual tiene un impacto pequeño sobre el CIMD en las mujeres que entrenan en fuerza. Este resultado inesperado puede explicarse parcialmente por algunos recientes estudios y algunas variaciones de estudios en la literatura actual.
A pesar que se ha demostrado previamente que el estrógeno tiene un efecto protector sobre las células (Ayres, 1998; Tiidus, 2003), la recuperación de la fuerza fue mejor en el grupo de la fase folicular (estrógeno bajo). Ya que el estrógeno parece proteger las células, se pensó que concentraciones de estrógeno superiores exhibirían un decremento de la fuerza menos severo; sin embargo, se observó lo contrario en este estudio. Una alta concentración de estrógeno circulante fue asociada con la recuperación de fuerza más débil. La concentración de estrógeno en el momento del daño no pudo tener un efecto sobre la fuerza; en cambio, las concentraciones de estrógeno durante la recuperación pueden jugar un rol más crucial. Los días que las participantes fueron evaluadas fueron seleccionados debido a la diferencia anticipada de concentraciones de estrógeno en esos puntos en el ciclo menstrual. Durante la semana de la recuperación, las concentraciones de estrógeno del grupo de la fase folicular estarían aumentando naturalmente, mientras que en el grupo de la fase lútea habría revelado los niveles bajos del estrógeno. Una investigación más extensa acerca de esta inquietud es necesaria para una mejor conclusión.
La flexión y extensión del codo post-ejercicio mostró cada una un efecto de tiempo significativo. Hubo también un efecto de tiempo significativo para el volumen del brazo. Éstos son signos comunes del daño muscular (Glesson, 2003) y no fue inesperado observarlos.
Las concentraciones de la creatina kinasa significativamente aumentaron a través del tiempo, indicando que el ejercicio causó bastante CIMD para deteriorarse la integridad de las membranas celulares. Fue sorprendente que la única interacción fue la alta concentración de la CK del grupo de la fase lútea a las 96 horas. Se suponía que el grupo de la fase lútea tendría menos signos del daño muscular, pero ocurrió lo contrario. La magnitud de la diferencia en ese momento también fue inesperada. Una posible explicación para la falta de diferencias entre los grupos en este estudio es que las mujeres, sin tener en cuenta la fase, pueden tener suficiente estrógeno presente para ofrecer algún nivel de protección contra el daño muscular. Tal vez, un umbral para el efecto protector del estrógeno podría existir. Ayres y cols. (1998) observaron un efecto potente antioxidante del estrógeno, pero estos autores no vieron un beneficio aditivo cuando se usaron otros antioxidantes conocidos junto con el estrógeno.
Junto con la literatura que apoya una diferencia del sexo, los resultados de este estudio y el estudio de Ayres y cols. (1998) indican que el efecto del estrógeno sobre signos y síntomas del daño muscular es un enigma complejo. Es posible que no sea sólo la cantidad de estrógeno lo que tiene un efecto sobre el daño muscular, sino también la interacción del estrógeno con los receptores del estrógeno. Otros investigadores (Pellegrini, 2013, por ejemplo) han reportado diferencias del sexo sobre la actividad del receptor del estrógeno, y la interacción del estrógeno con los receptores del estrógeno puede ser responsable de la respuesta de la diferencia de sexo al CIMD que otros han reportado.
Respecto al uso de pastillas anticonceptiva s, Salvaje y Clarkson (28) compararon el daño muscular en mujeres que tomaban o no dichas pastillas. La única diferencia fue que el grupo que las tomaba tenía una recuperación retrasada de la fuerza. El grupo de la fase folicular tuvo una recuperación de la fuerza mayor que el grupo de la fase lútea. Las diferencias en la respuesta al CIMD varían entre los hombres y mujeres, y varios estudios publicados están en conflicto. El conflicto aparece en la hipótesis planteada por Markofski (2014) acerca que las participantes en este estudio hayan tenido una recuperación de fuerza más pobre aún teniendo altos niveles de estrógeno en el momento del turno del CIMD. Con lo cual se necesitan más estudios de investigación que exploren el rol complejo del estrógeno sobre la protección contra el CIMD, determinado a través del curso de tiempo en eventos que son característicos del daño muscular.
Por lo visto se puede observar que la respuesta de las mujeres a un turno de CIMD es dependiente en su fase del ciclo menstrual. Esto puede tener implicaciones para aquellas mujeres que entrenan la fuerza en forma exhaustiva o máxima como parte de su ejercicio físico, en donde su programa de ejercicio podría sincronizarse de acuerdo a su ciclo menstrual, como vimos en anteriores blogs. Específicamente, puede haber algún beneficio al entrenamiento de la fuerza intenso durante la fase folicular, como opuesto a la fase lútea, del ciclo menstrual.
Sin embargo, ante una falta de evidencia fuerte en apoyo al rol protector del estrógeno contra el daño muscular en el estudio de Markofski (2014), probablemente parece que la fase menstrual tendría un pequeño impacto sobre los síntomas del CIMD.
Otras lecturas de interés:
Más información relacionada a este tópico puede ser
encontrada en las siguientes webinar:
Bibliografía
Brentano, MA; Martins Kruel, LF. A review on strength exercise induced muscle damage: Applications, adaptation mechanisms and limitations. J Sports Med Phys Fitness 51: 1–10, 2011.
Markofski, MM, Braun, WA. Influence of menstrual cycle on indices of contraction-induced muscle damage. J Strength Cond Res 28(9): 2649–2656, 2014.
Ayres, S, Abplanalp, W, Liu, J, Subbiah, M. Mechanismsinvolved in the protective effect of estradiol-17beta on lipid peroxidation and DNA damage. Am J Physiol 274: E1002–1008, 1998.
Feng, X, Li, G; Wang, S. Effects of estrogen on gastrocnemius muscle strain injury and regeneration in female rats. Acta Pharmacol Sin 25: 1489–1494, 2004.
Hubal, M, Rubinstein, S, Clarkson, P. Muscle function in men and women during maximal eccentric exercise. J Strength Cond Res 22: 1332–1338, 2008.
Rinard, J, Clarkson, P, Smith, L, Grossman, M. Response of males and females to high-force eccentric exercise. J Sports Sci 18: 229–236, 2000.
Sayers, S, Clarkson, P. Force recovery after eccentric exercise in males and females. Eur J Appl Physiol 84: 122–126, 2001.
Stupka, N, Lowther, S, Chorneyko, K, Bourgeois, J, Hogben, C, Tarnopolsky, M. Gender differences in muscle inflammation after eccentric exercise. J Appl Physiol (1985)89: 2325–2332, 2000.
Graves, J, Clarkson, P, Litchfield, P, Kirwan, J, Norton, J. Serum creatine kinase activity following repeated bouts of isometric exercise with different muscle groups. Eur J Appl Physiol Occup Physiol 56: 657–661, 1987.
Gleeson, N, Eston, R, Marginson, V, and McHugh, M. Effects of prior concentric training on eccentric exercise induced muscle damage. Br J Sports Med 37: 119–125, 2003.
Taylor, R, Jayasinghe, UW, Koelmeyer, L, Ung, O, Boyages, J. Reliability and validity of arm volume measurements for assessment of lymphedema. Phys Ther 86: 205–214, 2006.
Tiidus, P. Influence of estrogen on skeletal muscle damage, inflammation, and repair. Exerc Sport Sci Rev 31: 40–44, 2003.
Pellegrini, M, Bulzomi, P, Lecis, M, Leone, S, Campesi, I, Franconi, F, Marino, M. Endocrine disruptors differently influence estrogen receptor band androgen receptor in male and female rat VSMC. J Cell Physiol 2013.
Peñailillo, L, Blazevich, A, Numazawa, H, Nosaka, K. Metabolic and muscle damage profiles of concentric versus repeated eccentric cycling. Med Sci Sports Exerc 45: 1773–1781, 2013.
Evans, WJ, Cannon, JG. The metabolic effects of exercise-induced muscle damage. Exerc Sport Sci Rev 19: 99–125, 1991.
Claassen, H, Schu¨nke, M, Kurz, B. Estradiol protects cultured articular chondrocytes from oxygen-radical-induced damage. Cell Tissue Res 319: 439–445, 2005.
Dykens, J, Simpkins, J, Wang, J, Gordon, K. Polycyclic phenols, estrogens and neuroprotection: A proposed mitochondrial mechanism. Exp Gerontol 38: 101–107, 2003.
Wang,
X, Sharma, R, Sikka, S, Thomas, AJ, Falcone, T, Agarwal, A. Oxidative stress is associated with increased
apoptosis leading to spermatozoa DNA damage in patients with male factor infertility.
Fertil Steril 80: 531–535, 2003.