
¿Cuál es el Componente Elástico en serie del Músculo Esquelético?
What is the Series Elastic Component in Skeletal Muscle?
Laboratorio de Rendimiento Humano, Universidad de Calgary (Canadá).
Artículo publicado en el journal Revista de Entrenamiento Deportivo, Volumen 26, Número 4 del año 2012.
Publicado 16 de diciembre de 2012
Resumen
INTRODUCCION
Debido a las limitaciones de espacio, concentraré mis comentarios en asuntos relacionados con la mejora del trabajo e ignoraré todos los aspectos discutidos acerca de la eficiencia. Comenzaré con unos pocos comentarios generales destinados a ilustrar los problemas conceptuales que encontré al leer el manuscrito; después analizaré las afirmaciones hechas por los autores, que espero que proporcionen información adicional o claridad sobre un tema sin resolverCOMENTARIOS GENERALES
1. En la Parte I, Ingen Schenau y otros aportaron cuatro posibles explicaciones para la mejor capacidad de trabajo de un músculo concéntricamente contraído después de un estiramiento (tiempo disponible para el desarrollo de la fuerza, el almacenamiento y reutilización de energía elástica, la potenciación de la maquinaria contráctil y la contribución de los reflejos). Dedujeron el apoyo para sus argumentos de un modelo muscular tipo Hill para las dos primeras explicaciones y de un modelo tipo Huxley para la tercera explicación. Pasar de un modelo conceptual a otro argumento medio contribuyó a facilitar la argumentación que intentaba resolver todos los razonamientos dentro de un marco consistente. (Procuraré ilustrar más tarde con ejemplos específicos cuán difícil es reconciliar todas las ideas propuestas en un marco estable; es decir, un modelo tipo Huxley.) 2. En el manuscrito entero, la noción de los elementos elásticos del músculo es muy importante, concretamente el elemento elástico en serie. Sin embargo, en todo el manuscrito, nunca se define el elemento elástico en serie salvo como el elemento elástico unido al elemento contráctil (aunque el elemento contráctil tampoco está definido). Generalmente se acepta que el puente cruzado tiene elasticidad en serie con su sitio productor de fuerza (Ford, Huxley & Simmons, 1981; Huxley & Simmons, 1971). Además, experimentos recientes han demostrado que existe una considerable elasticidad en los miofilamentos delgados y posiblemente en los gruesos (Higuchi, Yanagida & Goldman, 1995; Huxley, Sosa & Irving, 1994; Kojima, Ishijima & Yanagida, 1994; Wakabayashi y otros, 1994). Estas elasticidades en serie parecen ignoradas en la discusión de Ingen Schenau y otros; antes bien, sus consideraciones generalmente parecen ser correctas sólo si el componente elástico en serie está exclusivamente asociado con las aponeurosis y los tendones. 3. Lamenté que en las discusiones sobre los cambios de longitud in vivo de las fibras musculares y los tendones, Ingen Schenau y otros ignoraban en gran parte la evidencia experimental (ej., Griffiths, 1989; Hoffer, Caputi, Pose & Griffiths, 1989; Roberts, Marsh, Weyand & Taylor, 1997) aunque contaban con información indirecta.COMENTARIOS ESPECÍFICOS
1.La idea de que la producción de fuerza en el músculo esquelético se produce a través de los puentes cruzados que conectan los miofilamentos gruesos con los delgados ha sido aceptada en la comunidad científica, ya que existe la idea de que el puente cruzado incluye un componente elástico en serie con el extremo del puente cruzado productor de fuerza (Figura 1 en Huxley & Simmons, 1971). La fuerza en un puente cruzado individual y por tanto su trabajo potencia dependen directamente de la distancia x (Figura 1), una medida de la elongación del elemento elástico en el puente cruzado.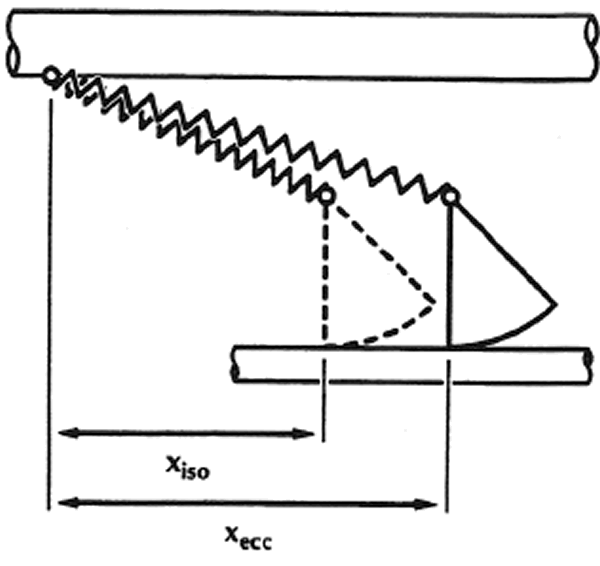
Figura 1. Dibujo esquemático de dos puentes cruzados de acuerdo con el modelo del puente cruzado propuesto por Huxley y Simmons (1971). xiso y xexc representan esquemáticamente la distancia desde la fijación del puente cruzado en el grueso filamento (miosina) (barra superior) hasta el enlace del puente cruzado en el filamento delgado (actina) (barra inferior). Las observaciones experimentales y la estricta aplicación de la teoría del puente cruzado sugieren que la distancia x media aumenta después del estiramiento, en comparación con la situación en la que el músculo es contraído isométricamente. (Nota: la distancia x mostrada aquí no es exactamente la misma que la definida por Huxley & Simmons, 1971).Cuando se estira un músculo activo, la distancia x media se vuelve más larga en comparación con una concentración isométrica o concéntrica, incrementando por tanto la fuerza y el trabajo potencial de los puentes cruzados individuales (ej., Cale, van den Bogert, Herzog & Gerritsen, 1996). Por lo tanto, no coincido del todo con Ingen Schenau y otros en que la cantidad de energía elástica almacenada al comienzo de una contracción concéntrica no está determinada por la cantidad de trabajo negativo sino sólo por la fuerza. Lo que los autores se olvidan de decir es que la fuerza depende (en algunos casos) del trabajo negativo; las dos no pueden separarse en entidades completamente independientes (si se acepta la teoría del puente cruzado y la idea de que existe la elasticidad en serie directamente dentro del mecanismo del puente cruzado).
2.Los autores argumentan (Parte I, Almacenamiento y reutilización de energía elástica) que la creciente elongación del elemento elástico en serie, para un origen dado hasta la distancia de inserción del complejo músculo-tendón, reduce la longitud del elemento contráctil y, consiguientemente, decrece el trabajo potencial durante la siguiente fase de acortamiento. Este argumento no es del todo correcto por varios motivos. Me gustaría mencionar un caso teórico primero (que podría considerarse poco realista) y una consideración totalmente realista después. Primero, imaginar un músculo con un tendón y aponeurosis rígidos que incluye toda su elasticidad en serie en el puente cruzado. Si tal músculo era activado isométricamente en una primera prueba y se activaba con una longitud menor que la longitud isométrica, entonces se estiraba hasta la longitud del objetivo isométrico y después se mantenía isométricamente en una segunda prueba, la fuerza en el último caso sería mayor y se conservaría superior durante un largo período de tiempo en comparación con el primer caso (ej., Abbott & Aubert, 1952). La esperada (esquemática) geometría del sarcómero de las contracciones isométrica y excéntrico-isométrica se representa en la Figura 2. Parece que bajo estas circunstancias, el trabajo potencial del músculo estirado (Figura 2,parte inferior) podría ser mayor que el del músculo isométrico (Figura 2,parte superior).
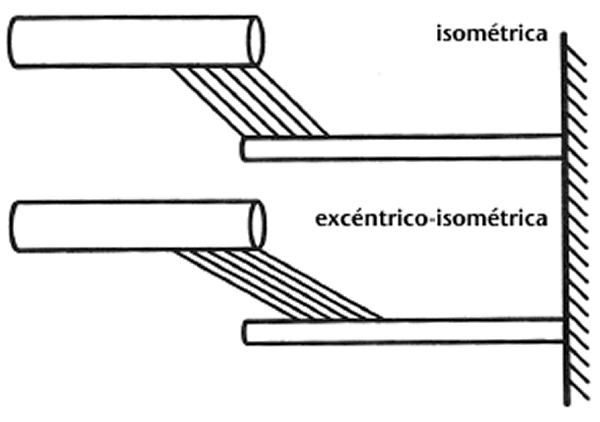
Figura 2. Dibujo esquemático de las distribuciones del puente cruzado (líneas oblicuas paralelas) entre los filamentos gruesos y delgados (las barras horizontales superiores e inferiores, respectivamente) después de una contracción isométrica y otra excéntrico-isométrica de un músculo con aponeurosis y tendones rígidos. Para una longitud dada del músculo-tendón y aponeurosis/tendones rígidos, la longitud del sarcómero sería idéntica en ambos casos aunque la fuerza media del puente cruzado y el trabajo potencial sería mejorado en la contracción excéntrico-isométrica en comparación con la isométrica.Segundo, imaginar un trabajo muscular sobre el miembro decreciente de la relación fuerza-longitud al comienzo de la contracción concéntrica. Una contracción excéntrica, que precede a la contracción concéntrica, produciría más fuerza en el músculo, un elemento elástico en serie más largo, y un elemento contráctil más corto al comienzo de la contracción concéntrica en comparación con un músculo que no se estiraba. Si suponemos ahora que el músculo se acorta una cantidad dada en ambas situaciones, y asumimos además que el trabajo efectuado por los músculos es proporcional al área bajo la curva fuerza-longitud, entonces partir de una menor longitud del elemento contráctil posiblemente fuera ventajoso (Figura 3).
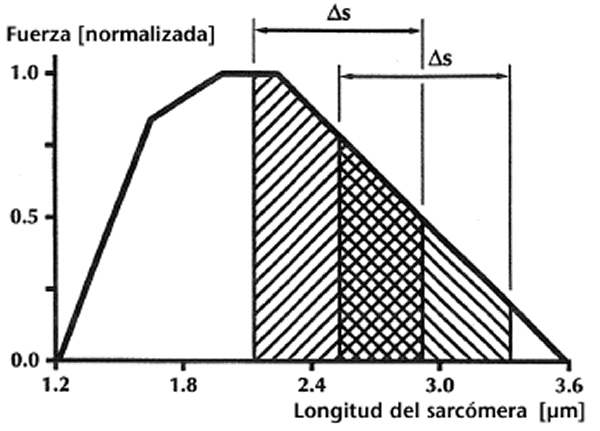
Figura 3. Relación fuerza-longitud esquemática del sarcómero para el músculo esquelético de la rana. Imaginar un complejo músculo-tendón que reduce una distancia dada, s. El trabajo potencial (áreas sombreadas) podría ser mayor para el músculo cuya longitud del elemento contráctil es más corta al comienzo de la fase de acortamiento. El trabajo potencial de un músculo dinámicamente contraído no es igual al área bajo la relación fuerza-longitud isométrica, aunque conceptualmente, este área podría representar adecuadamente la fuerza potencial del músculo.3.Ingen Schenau y otros cuestionan la existencia fundamental del ciclo de estiramiento-acortamiento (Parte I, Potenciación de la maquinaria contráctil, La contribución de los reflejos). Los autores sugieren que “queda por verse si las fibras musculares se alargan siempre” (p. 394), refiriéndose a los ciclos de estiramiento-acortamiento in vivo. Aunque estoy informado de la evidencia indirecta citada en consonancia con esta afirmación, y además conozco la evidencia experimental en dos músculos de la articulación durante la lenta locomoción animal que apoya la idea de que apenas existe el acortamiento de la fibra durante los ciclos de estiramiento-acortamiento in vivo (ej., Hoffer y otros, 1989). Me resulta muy difícil aceptar esta afirmación en general. Por ejemplo, los datos de la longitud de la fibra in vivo presentados por Hoffer y otros (1989) muestran un leve acortamiento de las fibras del gastrocnemio del gato a medida que el músculo se elonga con el contacto de la garra. Sin embargo, este ejemplo se obtenía con una lenta velocidad de marcha; la elongación del músculo en la fase crítica era casi de 1.5-2.0 mm (estimada a partir de la Figura 2 de Hoffer y otros, 1989). Goslow, Reinking y Stewart (1973) también encontraron un incremento de unos 2 mm en la longitud del gastrocnemio medial durante una marcha lenta al comienzo de la fase postural. Sin embargo, cuando el animal corría, la longitud del gastrocnemio medial aumentaba casi 15 mm en esta fase crítica, y la longitud del sóleo crecía unos 16-17 mm. Estos incrementos de la longitud, si el tendón y la aponeurosis los absorbían completamente, corresponderían casi con una tensión del 15% y del 33% en los elementos elásticos en serie del gastrocnemio medias y del sóleo, respectivamente. Dado que la falta de tensión de los tejidos tendinosos es casi del 8-10% (Rigby, Hirai, Spikes & Eyring, 1959), es seguro asumir que los ciclos de estiramiento-acortamiento se producen in vivo, al menos para algunos músculos bajo ciertas condiciones.
En resumen, experimenté que las cuestiones presentadas sobre la mejora del trabajo durante los ciclos de estiramiento-acortamiento carecían de convicción debido a que los argumentos se presentaron en diferentes marcos (oportunos) antes que en un marco estable, la elasticidad en serie no estaba definida, y la directa evidencia experimental in vivo sobre la conducta de la longitud de la fibra durante los ciclos de estiramiento-acortamiento se ignoraba en gran parte a costa de estimaciones indirectas.
Palabras clave: músculos, elasticidad
Abstract
Keywords: muscles, elasticity
INTRODUCCION
Debido a las limitaciones de espacio, concentraré mis comentarios en asuntos relacionados con la mejora del trabajo e ignoraré todos los aspectos discutidos acerca de la eficiencia. Comenzaré con unos pocos comentarios generales destinados a ilustrar los problemas conceptuales que encontré al leer el manuscrito; después analizaré las afirmaciones hechas por los autores, que espero que proporcionen información adicional o claridad sobre un tema sin resolver
COMENTARIOS GENERALES
1. En la Parte I, Ingen Schenau y otros aportaron cuatro posibles explicaciones para la mejor capacidad de trabajo de un músculo concéntricamente contraído después de un estiramiento (tiempo disponible para el desarrollo de la fuerza, el almacenamiento y reutilización de energía elástica, la potenciación de la maquinaria contráctil y la contribución de los reflejos). Dedujeron el apoyo para sus argumentos de un modelo muscular tipo Hill para las dos primeras explicaciones y de un modelo tipo Huxley para la tercera explicación. Pasar de un modelo conceptual a otro argumento medio contribuyó a facilitar la argumentación que intentaba resolver todos los razonamientos dentro de un marco consistente. (Procuraré ilustrar más tarde con ejemplos específicos cuán difícil es reconciliar todas las ideas propuestas en un marco estable; es decir, un modelo tipo Huxley.)
2. En el manuscrito entero, la noción de los elementos elásticos del músculo es muy importante, concretamente el elemento elástico en serie. Sin embargo, en todo el manuscrito, nunca se define el elemento elástico en serie salvo como el elemento elástico unido al elemento contráctil (aunque el elemento contráctil tampoco está definido). Generalmente se acepta que el puente cruzado tiene elasticidad en serie con su sitio productor de fuerza (Ford, Huxley & Simmons, 1981; Huxley & Simmons, 1971). Además, experimentos recientes han demostrado que existe una considerable elasticidad en los miofilamentos delgados y posiblemente en los gruesos (Higuchi, Yanagida & Goldman, 1995; Huxley, Sosa & Irving, 1994; Kojima, Ishijima & Yanagida, 1994; Wakabayashi y otros, 1994). Estas elasticidades en serie parecen ignoradas en la discusión de Ingen Schenau y otros; antes bien, sus consideraciones generalmente parecen ser correctas sólo si el componente elástico en serie está exclusivamente asociado con las aponeurosis y los tendones.
3. Lamenté que en las discusiones sobre los cambios de longitud in vivo de las fibras musculares y los tendones, Ingen Schenau y otros ignoraban en gran parte la evidencia experimental (ej., Griffiths, 1989; Hoffer, Caputi, Pose & Griffiths, 1989; Roberts, Marsh, Weyand & Taylor, 1997) aunque contaban con información indirecta.
COMENTARIOS ESPECÍFICOS
1.La idea de que la producción de fuerza en el músculo esquelético se produce a través de los puentes cruzados que conectan los miofilamentos gruesos con los delgados ha sido aceptada en la comunidad científica, ya que existe la idea de que el puente cruzado incluye un componente elástico en serie con el extremo del puente cruzado productor de fuerza (Figura 1 en Huxley & Simmons, 1971). La fuerza en un puente cruzado individual y por tanto su trabajo potencia dependen directamente de la distancia x (Figura 1), una medida de la elongación del elemento elástico en el puente cruzado.
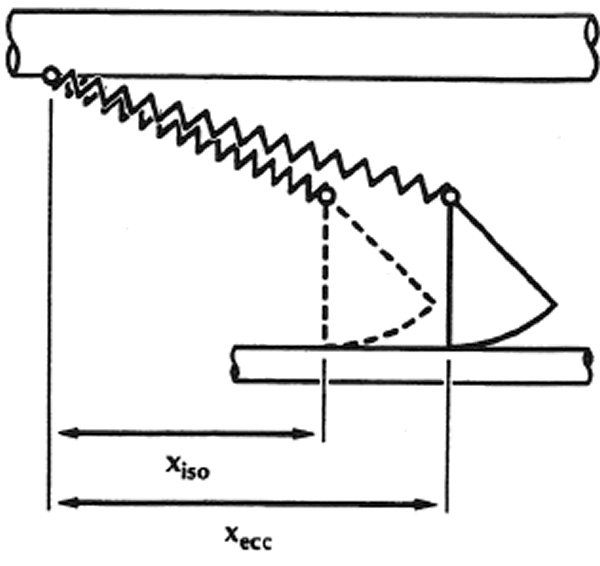
Figura 1: Dibujo esquemático de dos puentes cruzados de acuerdo con el modelo del puente cruzado propuesto por Huxley y Simmons (1971). xiso y xexc representan esquemáticamente la distancia desde la fijación del puente cruzado en el grueso filamento (miosina) (barra superior) hasta el enlace del puente cruzado en el filamento delgado (actina) (barra inferior). Las observaciones experimentales y la estricta aplicación de la teoría del puente cruzado sugieren que la distancia x media aumenta después del estiramiento, en comparación con la situación en la que el músculo es contraído isométricamente. (Nota: la distancia x mostrada aquí no es exactamente la misma que la definida por Huxley & Simmons, 1971).
Cuando se estira un músculo activo, la distancia x media se vuelve más larga en comparación con una concentración isométrica o concéntrica, incrementando por tanto la fuerza y el trabajo potencial de los puentes cruzados individuales (ej., Cale, van den Bogert, Herzog & Gerritsen, 1996). Por lo tanto, no coincido del todo con Ingen Schenau y otros en que la cantidad de energía elástica almacenada al comienzo de una contracción concéntrica no está determinada por la cantidad de trabajo negativo sino sólo por la fuerza. Lo que los autores se olvidan de decir es que la fuerza depende (en algunos casos) del trabajo negativo; las dos no pueden separarse en entidades completamente independientes (si se acepta la teoría del puente cruzado y la idea de que existe la elasticidad en serie directamente dentro del mecanismo del puente cruzado).
2.Los autores argumentan (Parte I, Almacenamiento y reutilización de energía elástica) que la creciente elongación del elemento elástico en serie, para un origen dado hasta la distancia de inserción del complejo músculo-tendón, reduce la longitud del elemento contráctil y, consiguientemente, decrece el trabajo potencial durante la siguiente fase de acortamiento. Este argumento no es del todo correcto por varios motivos. Me gustaría mencionar un caso teórico primero (que podría considerarse poco realista) y una consideración totalmente realista después. Primero, imaginar un músculo con un tendón y aponeurosis rígidos que incluye toda su elasticidad en serie en el puente cruzado. Si tal músculo era activado isométricamente en una primera prueba y se activaba con una longitud menor que la longitud isométrica, entonces se estiraba hasta la longitud del objetivo isométrico y después se mantenía isométricamente en una segunda prueba, la fuerza en el último caso sería mayor y se conservaría superior durante un largo período de tiempo en comparación con el primer caso (ej., Abbott & Aubert, 1952). La esperada (esquemática) geometría del sarcómero de las contracciones isométrica y excéntrico-isométrica se representa en la Figura 2. Parece que bajo estas circunstancias, el trabajo potencial del músculo estirado (Figura 2,parte inferior) podría ser mayor que el del músculo isométrico (Figura 2,parte superior).
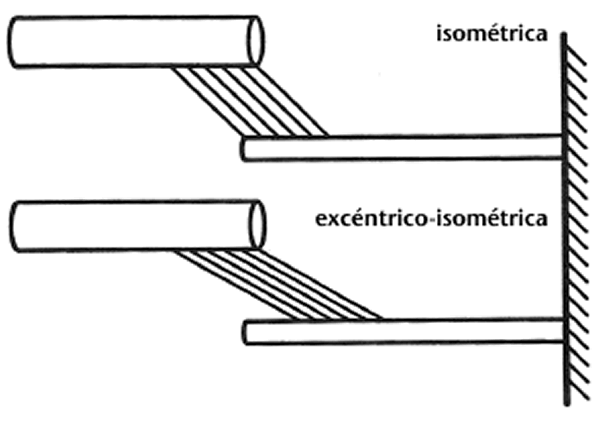
Figura 2: Dibujo esquemático de las distribuciones del puente cruzado (líneas oblicuas paralelas) entre los filamentos gruesos y delgados (las barras horizontales superiores e inferiores, respectivamente) después de una contracción isométrica y otra excéntrico-isométrica de un músculo con aponeurosis y tendones rígidos. Para una longitud dada del músculo-tendón y aponeurosis/tendones rígidos, la longitud del sarcómero sería idéntica en ambos casos aunque la fuerza media del puente cruzado y el trabajo potencial sería mejorado en la contracción excéntrico-isométrica en comparación con la isométrica.
Segundo, imaginar un trabajo muscular sobre el miembro decreciente de la relación fuerza-longitud al comienzo de la contracción concéntrica. Una contracción excéntrica, que precede a la contracción concéntrica, produciría más fuerza en el músculo, un elemento elástico en serie más largo, y un elemento contráctil más corto al comienzo de la contracción concéntrica en comparación con un músculo que no se estiraba. Si suponemos ahora que el músculo se acorta una cantidad dada en ambas situaciones, y asumimos además que el trabajo efectuado por los músculos es proporcional al área bajo la curva fuerza-longitud, entonces partir de una menor longitud del elemento contráctil posiblemente fuera ventajoso (Figura 3).
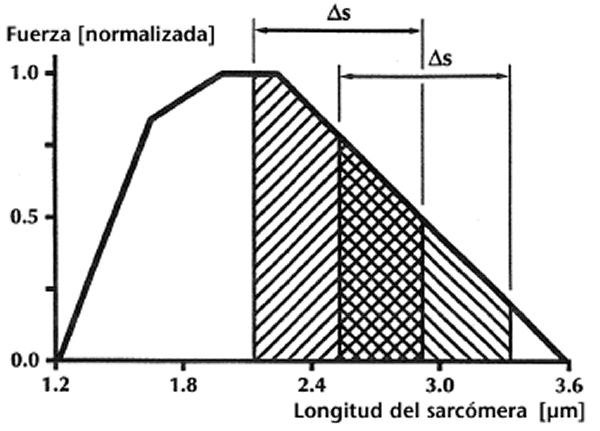
Figura 3: Relación fuerza-longitud esquemática del sarcómero para el músculo esquelético de la rana. Imaginar un complejo músculo-tendón que reduce una distancia dada, s. El trabajo potencial (áreas sombreadas) podría ser mayor para el músculo cuya longitud del elemento contráctil es más corta al comienzo de la fase de acortamiento. El trabajo potencial de un músculo dinámicamente contraído no es igual al área bajo la relación fuerza-longitud isométrica, aunque conceptualmente, este área podría representar adecuadamente la fuerza potencial del músculo.
3.Ingen Schenau y otros cuestionan la existencia fundamental del ciclo de estiramiento-acortamiento (Parte I, Potenciación de la maquinaria contráctil, La contribución de los reflejos). Los autores sugieren que “queda por verse si las fibras musculares se alargan siempre” (p. 394), refiriéndose a los ciclos de estiramiento-acortamiento in vivo. Aunque estoy informado de la evidencia indirecta citada en consonancia con esta afirmación, y además conozco la evidencia experimental en dos músculos de la articulación durante la lenta locomoción animal que apoya la idea de que apenas existe el acortamiento de la fibra durante los ciclos de estiramiento-acortamiento in vivo (ej., Hoffer y otros, 1989). Me resulta muy difícil aceptar esta afirmación en general. Por ejemplo, los datos de la longitud de la fibra in vivo presentados por Hoffer y otros (1989) muestran un leve acortamiento de las fibras del gastrocnemio del gato a medida que el músculo se elonga con el contacto de la garra. Sin embargo, este ejemplo se obtenía con una lenta velocidad de marcha; la elongación del músculo en la fase crítica era casi de 1.5-2.0 mm (estimada a partir de la Figura 2 de Hoffer y otros, 1989). Goslow, Reinking y Stewart (1973) también encontraron un incremento de unos 2 mm en la longitud del gastrocnemio medial durante una marcha lenta al comienzo de la fase postural. Sin embargo, cuando el animal corría, la longitud del gastrocnemio medial aumentaba casi 15 mm en esta fase crítica, y la longitud del sóleo crecía unos 16-17 mm. Estos incrementos de la longitud, si el tendón y la aponeurosis los absorbían completamente, corresponderían casi con una tensión del 15% y del 33% en los elementos elásticos en serie del gastrocnemio medias y del sóleo, respectivamente. Dado que la falta de tensión de los tejidos tendinosos es casi del 8-10% (Rigby, Hirai, Spikes & Eyring, 1959), es seguro asumir que los ciclos de estiramiento-acortamiento se producen in vivo, al menos para algunos músculos bajo ciertas condiciones.
En resumen, experimenté que las cuestiones presentadas sobre la mejora del trabajo durante los ciclos de estiramiento-acortamiento carecían de convicción debido a que los argumentos se presentaron en diferentes marcos (oportunos) antes que en un marco estable, la elasticidad en serie no estaba definida, y la directa evidencia experimental in vivo sobre la conducta de la longitud de la fibra durante los ciclos de estiramiento-acortamiento se ignoraba en gran parte a costa de estimaciones indirectas.
Referencias
1. Abbott, B.C., y Aubert, X.M. (1952). The force exerted by active striated muscle during and after change of length. Journal of Physiology (London), 117, 77-86.
2. Cole, G., van den Bogert, A.J., Herzog, W., y Gemtsen, K. (1996). Modelling of force production in skeletal muscle undergoing stretch. Journal of Biomechanics, 29, 1091-1104.
3. Ford, L.E., Huxley, A.F., y Simmons, R.M. (1981). The relation between stiffness and filament overlap in stimulated frog muscle fibres. Journal of Physiology (London), 311, 219-249.
4. Goslow, G.E., Jr., Reinking, R.M., y Stuart, D.G. (1973). Physiological extent, range and rate of muscle stretch for soleus, medial gastrocnemius and tibialis anterior in the cat. European Journal of Physiology, 341,77-86.
5. Griffiths, R.I. (1989). The mechanics of the medial gastrocnemius muscle in the freely hopping wallaby (Thylogale billardierii). Journal of Experimental Biology, 147, 439-456.
6. Higuchi, H., Yanagida, T., y Goldman, Y.E. (1995). Compliance of thin filaments in skinned fibers of rabbit skeletal muscle. Biophysical Journal, 69, 1000-1010.
7. Hoffer, J.A., Caputi, A.A., Pose, I.E., y Griffiths, R.I. (1989). Roles of muscle activity and load on the relationship between muscle spindle length and whole muscle length in the freely walking cat. In J.H.H. Allum y M. Hulliger (Eds.), Progress in brain research (pp. 75-85). Amsterdam: Elsevier Science Publishers B.V.
8. Huxley, A.F., y Simmons, R.M. (1971). Proposed mechanism of force generation in striated muscle. Nature, 233, 533-538.
9. Huxley, H.E., Stewart, A., Sosa, H., y Irving, T. (1994). X-ray diffraction measurements of the extensibility of actin and myosin filaments in contracting muscles. Biophysical Journal, 67, 2411-242 1.
10. Ingen Schenau, G.J. van, Bobbert, M.F., y Haan, A. de (1997). Does elastic energy enhance work and efficiency in the stretchshortening cycle?. Journal of Applied Biomechanics, 13, 389- 415.
11. Kojima, H., Ishijima, A., y Yanagida, T. (1994). Direct measurement of stiffness of single actin filaments with and without tropomyosin by in vitro nanomanipulation. Proceedings of the National Academy of Sciences of the United States of America, 91, 12962-12966.
12. Rigby, B.J., Hirai, N., Spikes, J.D., y Eyring, H. (1959). The mechanical properties of rat tail tendon. Journal of General Physiology, 43, 265-283.
13. Roberts, T.J., Marsh, R.L., Weyand, P.G., y Taylor, C.R. (1997). Muscular force in running turkeys: The economy of minimizing work. Science, 275, 11 13-1 115.
14. Wakabayashi, K., Sugimoto, Y., Tanaka, H., Ueno, Y., Takezawa, Y., y Amemiya, Y. (1994). X-ray diffraction evidence for the extensibility of actin and myosin filaments during muscle contraction. Biophysical Journal, 67, 2422-2435.