
Efecto de la Sesión Repetida Provocado por la Caminata en Descenso Hacia Atrás
Carmel Nottle1 y Kazunori Nosaka1
School of Exercise, Biomedical and Health Sciences, Edith Cowan University, Joondalup, WA, Australia.
Artículo publicado en el journal PubliCE, Volumen 0 del año 2005.
Publicado 16 de abril de 2008
Resumen
Palabras clave: daño muscular inducido por el ejercicio, tríceps sural, fuerza isométrica, creatinquinasa, sensibilidad muscular
INTRODUCCION
La caminata en descenso hacia atrás ha sido utilizada previamente como un modelo de estudio para estudiar el daño muscular inducido por el ejercicio (EIMD) en el tríceps sural, y se ha comprobado que produce síntomas similares a los observados en otros grupos musculares luego de la realización de ejercicio excéntrico (1). Sin embargo, la magnitud del daño muscular inducido por la caminata en descenso hacia atrás no es tan grande como la observada luego de la realización de ejercicio excéntrico máximo (2), pero es similar a la observada luego del ejercicio excéntrico submáximo (3). Debido a que en la caminata en descenso hacia atrás se utilizan los mismos músculos de las piernas que realizan el ejercicio excéntrico submáximo repetido durante la locomoción diaria, estos grupos musculares son ideales para investigar el EIMD que se produce durante el ejercicio diario o en las actividades deportivas recreativas. Adicionalmente, el uso de un movimiento de paso durante la caminata en descenso hacia atrás le permite al modelo de ejercicio aislar eficazmente al tríceps sural de una pierna solamente, lo que hace que sea muy útil en estudios dónde se desea establecer comparaciones entre ambos miembros.
Está bien documentado que una serie de ejercicio excéntrico confiere protección contra el EIMD, y esta adaptación se conoce como el efecto de la sesión repetida (2, 8, 13). El efecto de la sesión repetida se produce cuando luego de realizar una primera serie, se repite el mismo ejercicio excéntrico o un ejercicio similar (2). El resultado que se observa es una reducción sustancial y en algunos casos, la ausencia de síntomas de inicio retardado de dolor muscular y de eflujo de creatinquinasa (CK) plasmática, además de una recuperación más rápida de la pérdida de fuerza (3, 4). Así como el daño muscular inducido por el ejercicio (EIMD) y el efecto de la sesión repetida son dos fenómenos ampliamente estudiados y publicados, los músculos más comúnmente estudiados son los flexores del codo y los extensores de la rodilla. El efecto de la serie repetida también ha sido estudiado mediante modelos de ejercicio excéntrico máximo. Dado que se ha establecido que la magnitud del EIMD también depende de la intensidad y duración del ejercicio (5), podría esperarse que el grado de protección conferido también dependa de la intensidad y duración de la primera serie de ejercicio. Por esta razón, es posible que el efecto de la sesión repetida provocado por el ejercicio excéntrico submáximo sea diferente al provocado por el ejercicio excéntrico máximo.
Hay evidencia para sostener que durante la locomoción lenta, se reclutan preferentemente las unidades motoras tipo I durante las contracciones excéntricas (6). Debido a que se supone que la velocidad de contracción muscular durante la caminata en descenso hacia atrás es lenta, es posible asumir que las fibras reclutadas sean principalmente las fibras de contracción lenta. Por lo tanto, el efecto de la sesión repetida observado a continuación de la primera serie de caminata en descenso hacia atrás podría ser diferente al observado cuando se utilizan movimientos excéntricos máximos donde probablemente se reclutan las unidades motoras tipo II. Como la caminata en descenso hacia atrás reproduce más estrechamente la intensidad de la contracción realizada rutinariamente en la actividad diaria, sería razonable asumir que la protección sobre el tríceps sural ya debería existir como respuesta a la carga de las actividades diarias. Sin embargo, ningún estudio ha investigado el efecto de la sesión repetida de la caminata en descenso hacia atrás. Por lo tanto, el objetivo de este estudio consistió en comparar los cambios que se producen en los indicadores habituales de EIMD luego de una primera sesión y de sesiones posteriores repetidas de caminata en descenso hacia atrás. Se planteó como hipótesis que el efecto de la sesión repetida no podría haber sido proporcionado por la primera sesión de caminata en descenso hacia atrás, porque se asumió que el tríceps sural ya estába adaptado a la carga excéntrica como resultado de las actividades diarias.
METODOS
Sujetos y Protocolos de Ejercicio
Diecisiete sujetos fueron convocados para el estudio. Los mismos firmaron un consentimiento informado luego de la aprobación por parte del Comité de Investigación Ética con Seres Humanos de la Universidad Edith Cowan siguiendo las Normas Éticas de la Asociación Médica Mundial. Durante el período experimental, se solicitó a los sujetos que no tomaran ninguna medicación, que no realizaran cambios en su dieta y ni ninguna actividad enérgica. Los sujetos que habían realizado entrenamiento de fuerza para los miembros inferiores en los últimos 12 meses fueron excluidos del estudio.
Las mediciones para establecer comparaciones fueron realizadas en 13 ocasiones a lo largo de un período de 26 días. Cada sujeto realizó el protocolo de ejercicio dos veces. Los tests de evaluación fueron realizados 72 y 48 horas antes de la caminata en descenso hacia atrás (Tiempo 0), y 0, 5, 24, 48, 72 y 96 horas luego de la primera sesión de caminata (sesión 1). Para la sesión repetida, las medidas fueron tomadas 48 horas antes, y 0, 5, 24, 48, 72 y 96 horas después de la caminata (sesión 2). Se dejó transcurrir un período de 7 días entre la realización de las pruebas de la sesión 1 y de la línea de base (tiempo 0), antes del inicio de la sesión 2. Este período fue establecido para permitir la recuperación de todas las mediciones a los niveles basales previos, antes de la segunda serie, en base al período de tiempo de recuperación previamente evaluado (1).
El protocolo del ejercicio consistió en 60 min de caminata en descenso hacia atrás con un movimiento de pasos realizados con una sola pierna en una cinta caminadora con motor Trackmaster (TM500) (JAS manufacturing, TX, E.E.U.U.). La cinta caminadora fue modificada para permitir que la correa rotase en reversa, con una pendiente fijada en -15%. Los sujetos caminaron hacia atrás con un movimiento dedo-talón con la pierna izquierda (miembro que realizó el ejercicio) y la pierna derecha (miembro que no realizó ejercicio) fue mantenida junto a la izquierda. La velocidad de la cinta caminadora fue determinada durante la primera sesión de ejercicio con un ritmo individual de 30-35 pasos/min, y esa misma velocidad fue empleada durante la sesión repetida. Todos los tests fueron realizados en ambas piernas, sin embargo no se establecieron comparaciones entre ellas.
Procedimientos de Medición
Usando un dinamómetro isocinético Cybex 6000 (Cybex, NY, EE.UU.), se determinaron la fuerza isométrica voluntaria máxima y la fuerza isocinética de los flexores plantares del tobillo. Para ello, los sujetos permanecieron en una posición reclinada con la rodilla fija con una flexión de 90º. El torque isométrico fue determinado durante 5 s con el pie en tres posiciones: neutral (el 0°), dorsiflexión (-10°), y flexión plantar (10°). En cada uno de los ángulos se realizaron tres esfuerzos máximos con veinte segundos de descanso entre cada esfuerzo, y 60 segundos de descanso entre cada ángulo evaluado. El torque máximo producido en cada ángulo se registró para el análisis posterior. Se determinó la fuerza isocinética voluntaria máxima a velocidades angulares de 30, 60 y 120 °/s a lo largo de un intervalo completo de movimiento del tobillo de los participantes. Los sujetos realizaron tres repeticiones máximas a partir de las que se registró el torque máximo, con un intervalo de descanso de 60 s entre cada una de las velocidades evaluadas. La actividad de la creatinquinasa plasmática (CK) fue determinada mediante extracción de una muestra de 30 µL de sangre por punción que fue luego colocada mediante una pipeta en una tira comercial para la determinación de la CK (Bohringer-Mannheim, Indiana, EE.UU.) y analizada por medio de un Reflotrón (Bohringer-Mannheim, Indiana, EE.UU.). El dolor y la sensibilidad muscular fueron evaluados en 5 sitios encima del tríceps sural. Para el análisis posterior se utilizó el promedio obtenido a partir de los 5 sitios. Dos sitios proximales fueron localizados 5 cm por debajo de la fosa poplítea, encima del vientre de los gastrocnemios medio y lateral. Dos sitios distales se localizaron 5 cm sobre la inserción de las cabezas lateral y medial de los gastrocnemios por encima del vientre del músculo. Un sitio fue establecido en el soleo, 5 cm debajo de la inserción de las cabezas media y lateral de los gastrocnemios a lo largo de la línea media de la pierna. Se cuantificó el dolor muscular utilizando una escala de 1 a 10 donde 1 era dolor normal y 10 muy, muy doloroso. Los sujetos expresaban la magnitud del dolor mientras caminaban por una superficie plana y estable. La sensibilidad muscular fue determinada mediante un miómetro (Dobros) con punta de goma de 1,5 cm. El miómetro se colocó en los cinco sitios establecidos con presión creciente (con un valor máximo de 100 kPa [14,5 psi]), y los sujetos debían informar el momento en que percibían el dolor. De esta manera, la presión necesaria para causar dolor se registró como sensibilidad, de modo que a mayor presión necesaria, menor era la sensibilidad.
Análisis Estadísticos
Cada variable dependiente fue analizada mediante un ANOVA de mediciones repetidas de dos vías (tiempo x series); ambas piernas fueron analizadas por separado (a excepción de la CK plasmática). En aquellos casos en que se encontraba un valor significativo de p (p<0,05), se realizaron comparaciones simples con los valores iniciales (tiempo 0) para determinar cuales intervalos de tiempo eran significativamente diferentes al tiempo 0 para cada sesión por separado. Para establecer las diferencias significativas entre las sesiones, se realizaron test-t de muestras pareadas con un nivel de significancia ajustado a Bonferroni (p<0,0083) entre los intervalos de tiempo para las dos sesiones de ejercicio. Todos los resultados se expresaron como Media±desvío estándar.
RESULTADOS
La edad, talla y peso promedio de los sujetos involucrados en el estudio fueron: edad: 26,6±6,6 años; talla: 169,6±9,0 cm y peso: 72,2±14,3 kg.
Fuerza Voluntaria Máxima
En el caso de la pierna que no realizó ejercicio no se observaron diferencias significativas (p>0,05) con respecto al tiempo 0 para ninguna de las variables de fuerza a lo largo del tiempo. Adicionalmente, no se observaron diferencias significativas (p>0,05) entre las sesiones para ninguna de las mediciones de fuerza. En el caso de la pierna que realizó ejercicio, se observaron disminuciones significativas (p<0,05) con respecto al tiempo 0, en el torque isométrico máximo voluntario con un ángulo de dorsiflexión de 10°, en todos los tiempos que se determinó luego de la primera sesión (Figura 1).
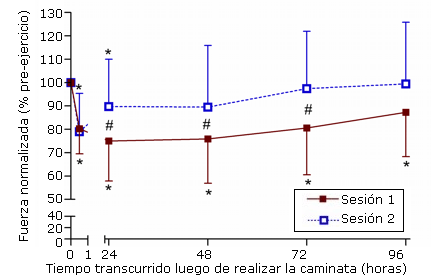
Figura 1. Cambios en el torque isométrico máximo voluntario determinados en dorsiflexión de 10º antes de realizar ejercicio (tiempo 0; 100%) y a las 0, 5, 24, 48, 72, y 96 horas posteriores a la realización de la primera (sesión 1) y segunda sesión (sesión 2) de ejercicio de caminata en descenso hacia atrás (* p<0,05 diferencia con respecto al tiempo 0; # p<0,0083 diferencia entre las series).
Cambios similares fueron observados en los ángulos de 0° (neutral) y 10° de flexión plantar del tobillo, sin embargo, en estos ángulos, sólo se registraron disminuciones significativas con respecto al tiempo 0 en 0, 5 y 24 horas luego de la caminata (Tabla 1). No se observaron diferencias significativas en el torque isométrico máximo voluntario pre-ejercicio entre la primera y segunda sesión. En la segunda sesión, las diferencias significativas (p<0,05) con respecto al tiempo 0, solamente fueron observadas a 0, 5 y 24 horas post-caminata en todos los ángulos evaluados. Adicionalmente, la magnitud de la disminución en la fuerza isométrica fue significativamente (p<0,05) menor en la segunda sesión en comparación con la primera, cuando el pie estaba tanto en la posición de flexión plantar (Tabla 1) como en dorsiflexión (Figura 1).
En Tabla 1 se muestran los cambios en el torque isocinético voluntario máximo normalizados. Las magnitudes de cambio en el torque isocinético fueron similares a las encontradas para el torque isométrico luego de la sesión inicial (Tabla 1 y Figura 1). Se encontraron disminuciones significativas en el torque isocinético luego de la sesión de ejercicio inicial, sin embargo no se observaron disminuciones significativas en ninguna de las velocidades isocinéticas a continuación de la segunda sesión. Aunque se evidenció una tendencia hacia una respuesta más atenuada luego de la sesión 2 en comparación con la respuesta observada en la sesión 1, no se observaron diferencias significativas en la fuerza isocinética entre las dos sesiones.
Actividad de la CK Plásmática
Se observaron aumentos significativos en la actividad plasmática de CK con respecto a los valores iniciales (tiempo 0) luego de la primera sesión de ejercicio a las 24, 48, 72 y 96 horas luego de la caminata (Figura 2). La actividad plasmática de la CK no aumentó significativamente a continuación de la sesión de ejercicio. Los valores inferiores observados en la segunda sesión de ejercicio mostraron ser significativamente diferentes (p<0,0083) entre las sesiones, a las 24 y 96 horas luego de la caminata.
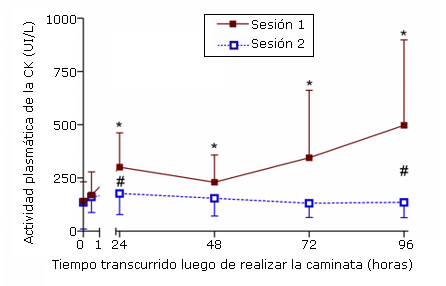
Figura 2. Actividad plasmática de CK antes de realizar el ejercicio (0) y luego de 0, 5, 24, 48, 72,
y 96 horas luego de la primera (sesión 1) y segunda (sesión 2) sesión de ejercicio de caminata en descenso hacia atrás. * p<0,05 diferencia significativa con respecto a la línea de base; # p<0,0083 diferencia significativa entre las dos sesiones.
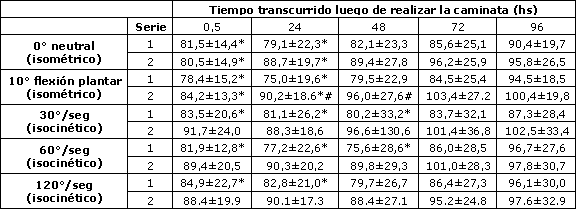
Tabla 1. Cambios (% con respecto a los valores registrados antes de realizar ejercicio) en la fuerza isométrica voluntaria máxima determinada a las 0, 5, 24, 48, 72 y 96 horas luego de la primera (sesión 1) y segunda sesión (sesión 2) de caminata en descenso hacia atrás. * p <0,05: Diferencias significativas con respecto al valor pre-ejercicio); # p <0,0083: Diferencias significativas entre las dos sesiones. Los datos se presentan como (Media±DS).
Dolor y Sensibilidad
Se registraron aumentos significativos luego de la realización de las dos series de ejercicio (p<0,001) en las determinaciones de dolor muscular realizadas a las 24, 48, 72 y 96 horas en la pierna que realizó el ejercicio, con valores máximos de dolor de 6,2±2,5 (sesión 1) y 3,1±2,1 (sesión 2) registrados a las 48 horas luego de realizar el ejercicio (Figura 3). Cuando se compara la magnitud del dolor muscular entre las sesiones, se observa que este fue significativamente (p<0,01, p <0,0083) menor luego de la segunda sesión. Este valor fue aproximadamente 2 puntos más bajo que el valor determinado luego de la primera sesión. No se observaron aumentos significativos (p>0,05) en el dolor en la pierna que no realizó ejercicios en los tiempos estudiados.
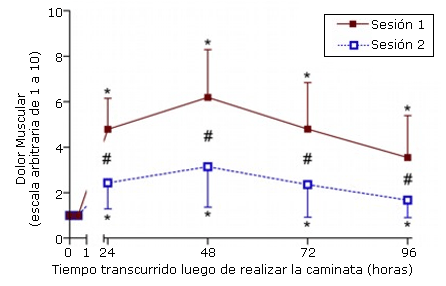
Figura 3. Dolor muscular antes (0) y 24, 48, 72, y 96 horas luego de la primera (sesión 1) y segunda (sesión 2) sesión de ejercicio de caminata en descenso hacia atrás. * p<0,05: diferencia significativa con respecto a la línea de base; # p<0,0083: diferencia significativa entre las sesiones.
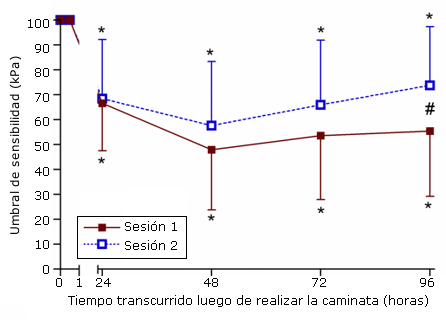
Figura 4. Sensibilidad muscular antes (0) y 24, 48, 72, y 96 horas luego de la primera (sesión 1) y segunda (sesión 2) sesión de ejercicio de caminata en descenso hacia atrás. * p<0,05: diferencia significativa con respecto a la línea de base; # p<0,0083: diferencia significativa entre las sesiones.
En el caso de la pierna que realizó el ejercicio, se registraron aumentos significativos en la sensibilidad luego de la sesión 1 (p<0,001) y la sesión 2 (p<0,01), con registros de sensibilidad máxima de 48,0±7,0; 48,0±28,9 kPa y 57,6±7,4; 57.6±30,5 kPa respectivamente, registrados 48 horas luego de la caminata (Figura 4). Aunque los valores de sensibilidad tendieron a ser más bajos en la segunda sesión, las diferencias solo fueron significativas (p <0,0083) en los valores registrados a las 96 horas luego del ejercicio. No se registraron cambios significativos (p>0,05), luego de ninguna de las dos sesiones en la sensibilidad de la pierna que no realizó ejercicio.
DISCUSION
En el presente estudio se observaron, luego de la segunda sesión de caminata en descenso hacia atrás, disminuciones significativamente más bajas y recuperaciones más rápidas de la fuerza voluntaria máxima (Figura 1, Tabla 1) y además menores aumentos en la actividad plasmática de CK (Figura 2), un desarrollo más atenuado de DOMS (Figura 3) y de sensibilidad (Figura 4) que los determinados luego de la primera sesión. Los cambios atenuados en las determinaciones realizadas luego de la segunda sesión parecen ser otorgados por la primera sesión de caminata hacia atrás en pendiente, y coinciden con el efecto de la sesión repetida observado en estudios anteriores realizados en otros grupos musculares como los flexores del codo (2) y extensores de la rodilla (3). Éste es el primer estudio que demuestra el efecto de la sesión repetida en el tríceps sural. Planteamos como hipótesis que el efecto de la sesión repetida no podría ser provocado por una primera sesión de ejercicio de caminata en pendiente hacia atrás porque se asumía que el tríceps sural ya se estaba adaptado a la carga excéntrica a través de las actividades diarias. Sin embargo el daño muscular observado a continuación de la primera serie inicial sugiere que no se había conferido ninguna protección. Como resultado de esto, nos preguntamos si el tríceps sural presentaría el efecto de la sesión repetida, similar al observado en otros grupos musculares. A pesar de los trabajos anteriores, no fue posible asumir que el tríceps sural respondería de la misma manera a la carga excéntrica repetida, especialmente debido a que no se conoce el estímulo exacto para producir el efecto de la sesión. Sin embargo, los resultados del presente estudio sugieren que el tríceps sural no constituye una excepción.
Se han propuesto adaptaciones neurales, mecánicas, y celulares para el efecto de la sesión repetida (4), sin embargo todavía existe debate con respecto a cual es el mecanismo principal que lo causa. Es importante señalar que las disminuciones en la fuerza isométrica máxima voluntaria con respecto a los valores iniciales ocurrieron, luego de las dos sesiones de ejercicio, y que la magnitud de pérdida de fuerza inmediatamente después del ejercicio fue similar entre las sesiones (Figura 1). Algunos estudios informaron una pérdida de fuerza atenuada inmediatamente después de la segunda sesión de ejercicio en comparación con la sesión inicial (3, 7). Warren et al. (7) concluyeron que la menor pérdida de fuerza inmediatamente después del ejercicio excéntrico puede deberse a una adaptación neural en la cual se produce una mayor activación de las unidades motoras lentas y una disminución concomitante en la activación de las unidades rápidas. Se ha especulado que un reclutamiento de unidades motoras más eficaz reduce la tensión en algunas fibras musculares que son susceptibles al daño muscular (3). Sin embargo, la pérdida de fuerza similar observada entre las dos sesiones en este estudio, no sugeriría que éste sea el caso para el tríceps sural al realizar la caminata en descenso hacia atrás.
Es importante señalar que la primera serie de caminata en descenso hacia atrás provocó un considerable daño muscular como puede advertirse por la pérdida prolongada de fuerza (Figura 1 y Tabla 1), por los aumentos en la actividad de la CK plasmática (Figura 2), y por el desarrollo de DOMS (Figura 3). Esto sugiere que los movimientos musculares excéntricos realizados durante la caminata en descenso hacia atrás excedieron el nivel de actividad diaria a la que las fibras musculares estaban acostumbradas normalmente. Dado que se ha demostrado que el efecto de la sesión repetida ocurre a continuación de solo un número pequeño de movimientos excéntricos (8), es sorprendente que los movimientos excéntricos realizados durante las actividades diarias no hayan conferido ninguna protección. Aunque la velocidad de alargamiento real de los movimientos excéntricos durante la caminata en descenso hacia atrás no fue determinada en este estudio, esta no parecía ser tan rápida comparada con la velocidad en otras actividades del tríceps sural como correr o saltar. Además, teniendo en cuenta que los sujetos caminaban con una acción de paso controlado, es posible asumir que la magnitud de carga sobre el músculo no excedió a la que se produce cuando se corre o salta (9). Así parecería que la tensión muscular durante el volumen de contracciones (1800 - 2300 contracciones) fue el estímulo principal involucrado en la inducción de daño muscular luego de la sesión inicial de ejercicio, porque la acción del paso con punta-talón, realizado durante la caminata en descenso hacia atrás probablemente excediera el intervalo de movimiento de la articulación en la locomoción normal.
Teniendo en cuenta que ha sido demostrado que durante los movimientos de estiramiento lento se reclutan preferentemente las unidades motoras lentas (6), sería razonable asumir que la mayoría de las fibras dañadas por la caminata en descenso hacia atrás fueron las fibras de contracción lenta. Es posible que algunas fibras musculares de contracción lenta se hayan adaptado a la carga excéntrica durante la caminata en descenso hacia atrás, pero que otras no lo hayan hecho. Además, se ha observado que los movimientos musculares excéntricos inducen una mayor magnitud de daño muscular en los músculos largos que en los músculos cortos (10). Es posible que el movimiento dedo-talón durante la marcha hacia atrás estirara el músculo más de lo habitual y causara daño muscular en la población de fibras de contracción lenta que generalmente serían resistentes al daño muscular inducido por el ejercicio excéntrico.
En respuesta a la pérdida de fuerza inmediatamente después del ejercicio se produce fatiga, falla del acoplamiento excitación-contracción [E-C] y rotura física de fibras musculares, mientras que se ha reportado que la pérdida de proteínas contráctiles prolonga la pérdida de fuerza (11). Dado que no se evidenció ninguna recuperación de fuerza desde 0,5 a las 24 horas después de la primera sesión (Figura 1, Tabla 1), es posible que los factores responsables de la pérdida de fuerza observada, utilizando este modelo de ejercicio, sean la falla de acoplamiento E-C y la ruptura física de las fibras en lugar de la fatiga. La recuperación más rápida de fuerza desde 0,5 a las 24 horas después de la segunda sesión de ejercicio sugiere que se produjo una recuperación más rápida de la falla de acoplamiento E-C y de la reparación de los elementos relacionados con la fuerza dentro del músculo. Es posible que esta reparación más rápida del daño físico reduzca el grado de pérdida de proteínas contráctiles y permita una velocidad de recuperación de fuerza significativamente más rápida luego de la segunda sesión (Figura 1), como se ha demostrado previamente en otros grupos musculares (2, 8).
Se ha sugerido que una respuesta adaptativa durante el proceso de reparación del daño de la sesión inicial pueda producir una reorganización estructural que permita a las fibras musculares volverse más resistentes al daño en las sesiones repetidas. La reorganización propuesta incluye la remoción de sarcómeros debilitados, fortalecimiento de la membrana celular, y adición longitudinal de sarcómeros (12,13). Los resultados con modelos humanos han demostrado que mientras mayor sea el estiramiento del músculo con respecto a la longitud de reposo, mayores serán los síntomas de daño muscular (3). El estiramiento causado por tales contracciones causa el "estallido" de los sarcómeros dentro del músculo, lo que causa la ruptura del filamento de titina, el punto de anclaje del filamento de miosina al disco Z (14). Esta ruptura del filamento inhibe la interacción de las proteínas entre los sarcómeros necesaria para la generación de fuerza y por consiguiente limita la generación de fuerza del músculo afectado (15). La ruptura de los sarcómeros también ha sido utilizada para explicar las observaciones de un cambio en el ángulo óptimo a continuación del ejercicio excéntrico (16). Luego del “estallido”, la reestructuración de los sarcómeros y de otros filamentos intermedios dentro del músculo podría proteger al músculo frente la sesión repetida de ejercicio excéntrico.
Sin embargo, no se conoce el grado hasta donde la reestructuración de los sarcómeros es responsable del efecto de la sesión repetida, porque investigaciones previas estudiaron los cambios en el ángulo óptimo luego de una sola serie de ejercicio, y demostraron que los cambios sólo se mantuvieron por dos días luego del ejercicio (12, 17). Como se cree que el ángulo óptimo se mueve hacia la derecha después del ejercicio excéntrico, lo que significa que la generación de fuerza máxima se produce en un ángulo de articulación mayor (12), es posible asumir que cualquier adaptación o reestructuración dentro del músculo ejercitado, como resultado de este cambio, ocurriría en una longitud del músculo más larga. Sin embargo, en el presente estudio se observó una diferencia entre las sesiones cuando el pie estaba en ambas posiciones; dorsiflexión (largo) y flexión plantar (corto), sin diferencias entre las series en el ángulo neutro. Parece improbable que esto se hubiera observado si los factores responsables de la adaptación, hubieran sido la reestructuración de los sarcómeros o el cambio en el ángulo óptimo, ya que la protección debería haber ocurrido en el estiramiento o acortamiento, pero no en las dos condiciones. Este planteo se basa en adaptaciones previas demostradas luego de realizar ejercicio excéntrico donde antes de este ejercicio, se había realizado un período de entrenamiento concéntrico (18, 19). Para entender completamente el grado en el que la reestructuración del sarcómero y el cambio en el ángulo óptimo contribuyen al efecto de la serie repetida en términos de pérdida de fuerza, sería necesario estudiar específicamente esta variable luego de realizar ejercicio excéntrico repetido.
En concordancia con resultados de investigaciones previas (20), se registró una reducción significativa en el flujo de CK plasmática a continuación de la segunda sesión de ejercicio en comparación con la sesión inicial (Figura 2). Así como se supone generalmente que el aumento de la actividad enzimática observado luego de realizar ejercicio refleja la existencia de daño en la membrana muscular (5), la elevada actividad de CK plasmática después de la primera sesión de ejercicio sugiere que algunas fibras musculares han sufrido necrosis. Los resultados de estudios anteriores, en los que se cuantificó directamente el daño histológico producido luego de sesiones repetidas de ejercicio excéntrico, apoyan esta idea (21). Es probable que la menor respuesta de CK refleje un daño muscular y una ruptura celular menos severos luego del ejercicio repetido en comparación con la primera sesión de ejercicio (22).
En el presente estudio también se observaron respuestas atenuadas en DOMS y en la sensibilidad del tríceps sural después de la segunda sesión de ejercicio. Esto coincide con varios estudios que también evaluaron el dolor muscular durante movimientos activos como caminar (3, 23). En la bibliografía existen pocas explicaciones acerca de por qué se produce una atenuación del dolor y/o de la sensibilidad luego de una sesión de ejercicio repetido. La adaptación neural es un mecanismo propuesto frecuentemente con relación a la “protección” frente a la pérdida de fuerza, donde una mejor capacidad para reparar la lesión y una menor respuesta inflamatoria se destacan como los mecanismos posibles para reducir el dolor (4). Esto se confirma por la recuperación más rápida de la fuerza y por los menores aumentos en la actividad de la CK plasmática luego de la segunda sesión en comparación con la primera. Sería razonable pensar que el dolor y la sensibilidad atenuados reflejan el menor grado de degeneración de inflamación muscular en la segunda sesión.
El protocolo de ejercicio utilizado en esta investigación fue seleccionado como un modelo para estudiar la magnitud de daño muscular que podría producirse durante la actividad diaria. El grado de daño muscular sugeriría, sin embargo, que el ejercicio es todavía “nuevo”. Habiendo dicho esto, el modelo de ejercicio de caminata en descenso hacia atrás es fácilmente adaptable y posteriores investigaciones en las cuales se varíen la pendiente de la cinta caminadora, el tiempo o la velocidad de caminata, pueden arrojar resultados diferentes. Dado que el efecto de la serie repetida parece ser un rasgo consistente de varios músculos esqueléticos, el estudio del grado de protección conferido por la actividad diaria puede ayudar a comprender cuales son los estímulos responsables de la adaptación.
Conclusiones
En conclusión, la sesión repetida de caminata en descenso hacia atrás provocó un menor indicio de daño muscular en comparación con la sesión inicial. Dado que la sesión inicial de caminata en descenso hacia atrás provocó cambios en todas las mediciones de referencia, sería razonable asumir que los movimientos excéntricos realizados durante la caminata exceden a los de las actividades diarias. Parecería que las fibras dañadas en la primer sesión se adaptaron a los movimientos excéntricos, así el daño muscular se atenuó después de la segunda sesión. El efecto de la sesión repetida demostrado en la presente investigación reafirma que la caminata hacia atrás en pendiente puede ser utilizada como un modelo eficaz para el estudio del daño muscular inducido por el ejercicio en el tríceps sural y apoya el potencial de entrenamiento excéntrico de la caminata hacia atrás.
Referencias
1. Nottle C (2004). Proprioceptive and muscle activation changes associated with exercise-induced muscle damage. Unpublished doctoral dissertation Edith Cowan University, Perth, Western Australia
2. Newham D, Jones D, Clarkson P (1987). Repeated high-force eccentric exercise: effects on muscle pain and damage. J Appl Physiol 63:1381-1386
3. Rowlands AV, Eston RG, Tilzey C (2001). Effect of stride length manipulation on symptoms of exercise-induced muscle damage and the repeated bout effect. J Sports Sci 19:333-340
4. McHugh MP, Connolly DA, Eston RG, Gleim GW (1999). Exercise-induced muscle damage and potential mechanisms for the repeated bout effect. Sports Med 27:157-170
5. Tiidus PM, Ianuzzo CD (1983). Effects of intensity and duration of muscular exercise on delayed soreness and serum enzyme activities. Med Sci Sports Exerc 15:461-465
6. Walmsley B, Hodgson JA, Burke RE (1978). Forces produced by medial gastrocnemius and soleus during locomotion in freely moving cats. J Neurophysiol 41:1203-1216
7. Warren GL, Hermann KM, Ingalls CP, Masselli MR, Armstrong RB (2000). Decreased EMG median frequency during a second bout of eccentric contractions. Med Sci Sports Exerc 32:820-829
8. Nosaka K, Sakamoto K, Newton M, Sacco P (2001). How long does the protective effects on eccentric exercise-induced muscle damage last?. Med Sci Sports Exerc 33:1490-1495.
9. Hof AL, VanZandwijk JP, Bobbert MF (2002). Mechanics of human triceps surae in walking, running and jumping. Acta Physiol Scand 174:17-30
10. Nosaka K, Sakamoto K (2001). Effect of elbow joint angle on the magnitude of muscle damage to the elbow flexors. Med Sci Sports Exerc 33:22-29
11. Warren GL, Ingalls CP, Lowe DA, Armstrong RB (2001). Excitation-contraction uncoupling: major role in contraction-induced muscle injury. Exerc Sport Sci Rev 29:82-87
12. Jones C, Allen T, Talbot J, Morgan DL, Proske U (1997). Changes in the mechanical properties of human and amphibian muscle after eccentric exercise. Eur J Appl Physiol Occupat Physiol 76:21-31
13. Friden J, Seger J, Sjostrom M, Ekblom B (1983). Adaptive response in human skeletal muscle subjected to prolonged eccentric training. Inter J Sports Med 4:177-183
14. Flitney F, Hirst D (1978). Cross-bridge detachment and sarcomere give during stretch of active frogs muscle. J Physiol 276:449-465
15. Lindstedt SL, LaStayo PC, Reich TE (2001). When active muscles lengthen: properties and consequences of eccentric contractions. News Physiol Sci 16:256-261
16. Whitehead NP, Allen TJ, Morgan DL, Proske U (1998). Damage to human muscle from eccentric exercise after training with concentric exercise. J Physiol 512:615-620
17. Whitehead NP, Morgan DL, Gregory JE, Proske U (2003). Rises in whole muscle passive tension of mammalian muscle after eccentric contractions at different lengths. J Appl Physiol 95:1224-1234
18. Lynn R, Talbot JA, Morgan DL (1998). Differences in rate skeletal muscles after incline and decline running. J Appl Physiol 85:98-104
19. Whitehead NP, Allen TJ, Morgan DL, Proske U (1998). Damage to human muscle from eccentric exercise after training with concentric exercise. J Physiol 512:615-620
20. Hortobagyi T, Houmard JA, Fraser DD, Dudek R, Lambert J, Tracy J (1998). Normal forces and myofibrillar disruption after repeated eccentric exercise. J Appl Physiol 84: 492-498
21. Ebbeling C, Clarkson PM (1989). Exercise-induced muscle damage and adaptation. Sports Med 7:207-234
22. Paddon-Jones D, Muthalib M, Jenkins D (2000). The effect of a repeated bout of eccentric exercise on indices of muscle damage and delayed onset muscle soreness. J Sci Med Sport 3:35-43.
23. McHugh MP, Connolly DA, Eston RG, Gartman EJ, Gleim GW (2001). Electromyographic analysis of repeated bouts of eccentric exercise. J Sports Sci 19:163-170