
El concepto de calorías en nutrición viola la segunda ley de la termodinámica
Publicado 18 de agosto de 2015, 17:07

Termodinámica
La nutrición arrastra desde hace décadas uno de los mitos más perversos y de mayor daño en la salud de las personas: “una caloría es una caloría”, esto implica básicamente que lo más importante de cualquier estrategia nutricional son las calorías y que la fuente de dichas calorías no importa, por lo tanto 2000 Kcal de brócoli, pechuga de pollo o caramelos son lo mismo.
Si bien, espero, ningún profesional de la salud en su sano juicio diría que esto es así, a la hora de analizar las estrategias empleadas para mejorar la composición corporal, principalmente tratar de disminuir el tejido adiposo, no son pocos los que optan por esta opción.
La realidad es que si uno tiene en cuenta la primera ley de la termodinámica (la energía no se crea ni se destruye, se conserva), las calorías, sea cual sea su precedencia, tienen la misma cantidad de energía potencial. Por definición 1 kcal. Es la cantidad de calor necesario para elevar la temperatura del agua (pura) 1˚C (desde 14,5 a 15,5 y a presión atmosférica) por lo tanto, esta energía potencial se puede traducir en calor o trabajo y se conservara en su magnitud. Sin embargo cuando entra en juego la biología las cosas no son tan simples.
El cuerpo humano es un sistema altamente complejo, con un elaborada e intrincada regulación del balance energético. Diferentes nutrientes tienen un impacto totalmente diferente tanto hormonal como en las vías metabólicas, las cuales son más o menos eficientes en las reacciones que contienen lo cual conlleva una menor o mayor perdida de energía en forma de calor. Es acá donde entra en escena la 2˚ Ley de la termodinámica.
La 2˚ ley, nos dice dos cosas, la primera es que en cualquier proceso irreversible, la ENTROPÍA (en la química se traduce como desorden, Ej: proteínas = orden, Aminoácidos = desorden) tiene que aumentar, y es esta ley la que dirige las reacciones químicas y la razón de la espontaneidad de dichas reacciones (a modo de ejemplo ilustrativo, si uno tira una gota de sangre en un vaso de agua, la misma se dispersará y será imposible volver a concentrarla en un pequeño volumen, a menos que se gaste energía, esta tendencia al desorden es la entropía), en segundo lugar, en toda reacción química parte de la energía utilizada se disipará en calor en mayor o menor medida según su eficiencia (a mayor eficiencia, menor calor disipado). Por lo tanto, la primera ley nos dice que el total de energía atribuible a trabajo, calor y a la composición química será constante, lo que no nos dice es si esa reacción efectivamente ocurrirá, ni tampoco la distribución relativa de dichas formas de energía e implica que “algo se perdió y no puede ser recuperado”(energía o incluso información).
Sin embargo hay una pequeña “trampa” cualquier sistema biológico es un sistema abierto con lo cual la entropía interna y la externa serán inversamente proporcionales, para ejemplificarlo de mejor manera tomaremos la siguiente situación: cuando ingresa alimento, el cuerpo lo “desarma” por lo tanto la entropía aumenta, sin embargo en muchísimas ocasiones es necesario rearmarlo (síntesis de proteínas, almacenamiento de glucógeno, síntesis de ADN, etc.), obviamente en forma predecible, por lo tanto la entropía disminuirá. Lo que nos dice la segunda ley es que si en un sistema la entropía decrece, en el ambiente (universo) se incrementará aún más, por lo que este incremento se debe a la radiación de calor por parte del cuerpo y por ende respeta una de las leyes mas fundamentales de la física que nos dice que el universo tiende a aumentar su entropía.
La primera ley entonces nos dice cuanta energía se necesita para por ej. Sintetizar ADN sumada a la perdida de calor para satisfacer la segunda ley (sin tomar en cuenta que el desacoplamiento mitocondrial, por ej. genera calor sin generar trabajo (Non-Shivering Thermogenesis) (Ricquier y Bouillaud, 2000).
Para un mayor entendimiento del tema, los invito a leer sobre el “demonio de Maxwell” o “paradoja de Maxwell” lo cual podría ser una explicación a la aplicación biológica del concepto de entropía, el de sistema cerrado y abierto y el rol de las enzimas.
Por lo tanto no hay forma que “una caloría sea una caloría” en un sistema biológico. Este sistema trata de mantener baja entropía (alta entropía interna = muerte) y por ende aumenta la del ambiente (universo) perdiendo energía.
Volviendo a la nutrición, en dietas cuyo objetivo es la pérdida de peso, la ineficiencia es deseada y esta altamente relacionada a las actividades enzimáticas y niveles hormonales, dicha ineficiencia es medida en el llamado “efecto térmico de los alimentos o “termogénesis inducida por la dieta”, esto es el calor generado al digerir la comida, absorberla y metabolizarla, en lo cual hay vasta literatura al respecto, incluyendo una excelente review de Jéquier (2002), donde concluye que este efecto térmico de los alimentos seria de 2-3% para los lípidos, 6-8% para los carbohidratos y 25-30% para las proteínas, lo que deja en claro que la eficiencia no es la misma y la entropía varia en su magnitud.
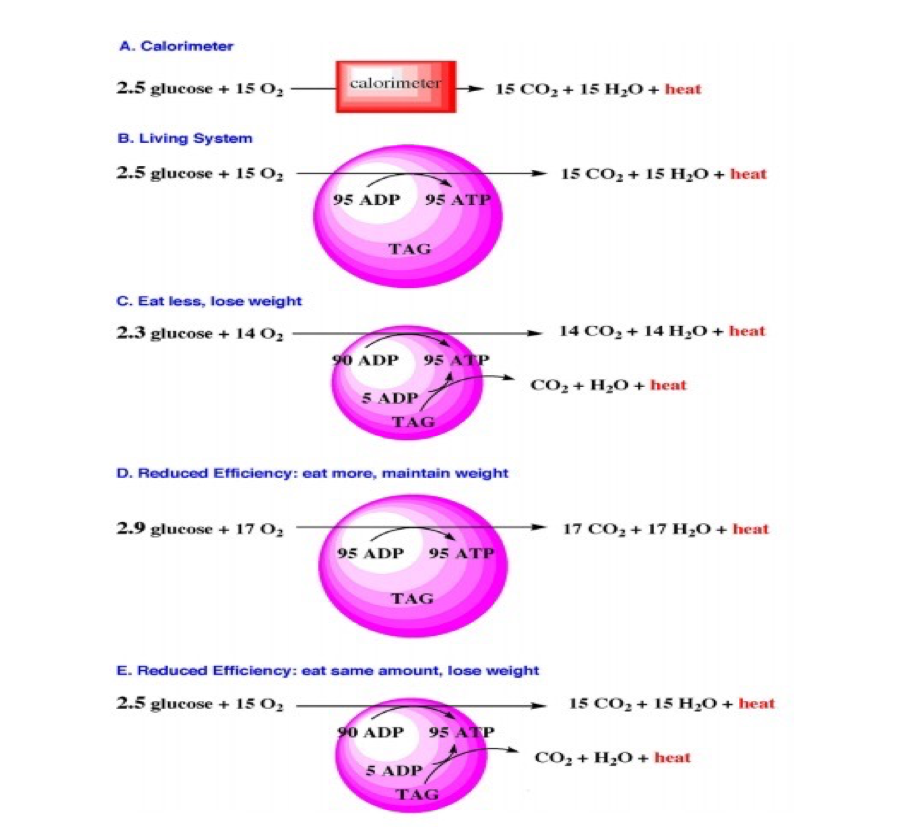
Fig.1: A: la oxidación de glucosa en un calorímetro es 100% ineficiente. Los productos de la oxidación son CO2, H2O y el total de la energía liberada en forma de calor. B: para ilustrar la primera ley de la termodinámica en sistemas biológicos, debemos considerar la que la conservación de la materia y energía implica excreción de productos al ambiente. Ni CO2 ni H2O permanecen en el organismo, solo el ATP, representando la energía útil, es retenido. El calor eliminado constituye el 60% de la energía de oxidación, mientras que la eficiencia es reflejada en el ATP retenido. Las reservas de grasas del cuerpo se muestran en forma de TAG (triacilglicerol). C: la forma mas común de pensar la reducción de peso es reduciendo la ingesta de calorías. Si el sujeto ingiere 2,3 moles de glucosa y produce solo 90 moles de ATP, la utilización de las reservas de TAG será necesaria para suplir el faltante de 5 ATP y mantener la homeostasis. CO2 y H2O (y Calor) serán excretados (e Irradiados) generando perdida de peso. D: si la eficiencia es reducida, entonces nuestro sujeto, deberá comer mas (ej.: 2,9 moles de glucosa) para producir 95 moles de ATP y mantener el peso. CO2 y H2O adicionales serán excretado manteniendo el peso constante. E: ante una reducción de la eficiencia metabólica (de 40% a 38% en este ejemplo), 90 moles de ATP serán producidos por la oxidación de 2,4 moles de glucosa. Los faltantes 5 moles de ATP necesarios para mantener la homeostasis provendrán de la oxidación de las reservas de lípidos (también podría ser de tejido muscular según contexto). Como resultado se vera una disminución del peso. Extraído de Fine y Feinman, 2004.
Termogénesis Adaptativa
Si partimos de la recomendación de “contar calorías”, esto implicaría adoptar una conducta de “comer menos, moverse más” para poder así establecer un déficit calórico o un balance calórico negativo. El problema con dicha premisa radica en su imposibilidad tanto práctica como biológica (Guth, 2014).
Incluso el más motivado e informado individuo es difícilmente capaz de estimar su ingesta de calorías (precisión en la ingesta, precisión en la información de las etiquetas y precisión en la real absorción de las mismas) (Baer y cols., 2010; Urban y cols., 2010; Hall y cols., 2012; Novotny y cols., 2012) y mucho menos su gasto, no solo en actividad física, sino también en el altamente variable gasto de procesos metabólicos y digestivos entre otros, (Feinmann y Fine, 2004; Westertrep y col., 1999; Novotny y col., 2014; Jakubowicz y cols., 2013) Incluso con el uso de tecnología disponible en el mercado (Lee y cols., 2014).
Por otro lado, biológicamente, la ingesta de calorías y el gasto energético están fuertemente regulados, por lo tanto una fuerte reducción de calorías ingresadas, resultara en una compensación proporcional en el gasto y vice - versa (Leibel y cols., 1995; Harris, 1990; Gibson y cols., 2005; Sumithran y Proietto, 2013; Ochner y cols., 2013; Lustig, 2006; Ludwig y Friedman, 2014). Por esta razón, cualquier persona que trate de consumir menos calorías resultará en una respuesta cerebral manifestada como cansancio (para disminuir el gasto) y en la gran mayoría de los casos, un hambre incontrolable y un impulso por alimentos calóricamente densos (Leibel y cols., 1995; Harris, 1990).
Sin embargo algunas personas tienen éxito (mayormente temporal) al adoptar este tipo de estrategias lo cual nos ayuda a plantear algunas preguntas: ¿Porqué y Cómo se llega a un balance neto negativo de gasto energético? O ¿Cómo es posible perder masa magra y seguir aumentando tejido adiposo?, por tanto el concepto de calorías no explica la respuesta no linear a los desbalances calóricos en el tiempo (Shook y cols., 2014; Hall y cols., 2011 y 2013) como tampoco explica porqué y cómo se desarrollan anormalidades metabólicas (Shah y Braverman, 2012; Widman y cols., 2008) y procesos patológicos relacionados con la obesidad aún antes de que haya variaciones en el peso corporal (Coutinho y cols., 2013; Song y cols., 2007; Hamer y Stamatakis, 2012), o incluso porqué mejoras ocurren aún ante un mantenimiento estable del peso (Gannon y Nutall, 2006) o porqué algunas patologías metabólicas no siempre disminuyen con una disminución concurrente del peso corporal (Wing y Bolin, 2013)
Regulación Hormonal y Metabólica
El tejido adiposo es un tejido altamente complejo , el cual tiene un rol fundamental en el control del apetito, regulación del peso y gasto energético. Normalmente cuando los adipocitos se “llenan” (almacén de lípidos) liberan una hormona llamada Leptina, la cual, nuevamente, en condiciones normales, se une a sus receptores en el sistema nervioso central, específicamente en el núcleo supraquiasmático en el hipotálamo (casualmente donde se encuentra el reloj biologico, mas adelante veremos porque es importante) (Harfmann y cols., 2014; Margetic y cols., 2002) donde estimula distintos tejidos periféricos (glándula tiroides, tejido muscular, intestino, etc.) básicamente inhibiendo la ingesta y aumentando el gasto (Sumieran y Proieto, 2013; Lustig; 2006; Speakman y cols., 2011), también tiene varias funciones incluyendo fertilidad, inmunidad, función cerebral, etc. (Allison y Myers, 2014).
Ser resistente a la acción de dicha hormona es uno de los factores que lleva al aumento de peso, principalmente a expensas del tejido adiposo, lo cual tiene lógica, si “llenar” los adipocitos aumenta los niveles de leptina circulante, es el mecanismo compensatorio ideal ante la imposibilidad de la misma a estimular sus receptores y por ende cumplir su función. Lo cual genera un circulo vicioso difícil de romper( Friedman y Halaas, 1998; Allison y Myers, 2014; Dalamaga y cols., 2013; Schwartz y cols., 2000).
Los obesos tienen altos niveles de leptina, lo cual, en cualquier situación normal actuaría en sus receptores de tal manera que señalizaría para regular el apetito y el gasto energético. Sin embargo los altos niveles de leptina no aseguran su correcta unión a sus receptores hipotalámicos, condición que se conoce como “resistencia a la leptina” (Considine y cols., 1996; Myers y cols., 2012; Jung y Kim, 2013). Por ende dicho problema de señalización se traduce en cambios fisiológicos y de comportamiento impulsados por el Cerebro para volver a llenar los depósitos de grasa que cree haber “perdido”, de tal manera que:
- Aumentará el apetito (para no morir de inanición en la que “cree” que está, condición que genera la mala señalización)
- Disminuir el gasto energético (el cerebro conserva la energía que cree que le falta, haciendo que se reduzca el nivel de actividad y se gaste menos incluso en reposo).
Teniendo en cuenta dicho mecanismo podemos concluir que al obeso no es consecuencia de la vagancia y glotonería (se mueve poco y come mucho) sino justamente lo contrario.
Otro punto importante es relacionar este mecanismo a la dificultad de sostener dietas bajas en calorías en el largo plazo lo cual no es la regla sino la excepción y una posible explicación es el rol de la leptina (Hinkle y cols., 2013; Wing y cols., 1996). Cuando la leptina cae, el hambre aumenta, disminuye la motivación a moverse y la energía gastada en reposo (Ahima, 2008; Keim y cols., 1998).
Por otro lado la leptina y la insulina están estrechamente relacionadas y la resistencia a la leptina es el paso previo que genera la resistencia a la insulina y la diabetes tipo 2 (German y cols., 2010; Seufert, 2004).
Volviendo al punto en cuestión de las calorías, varios trabajos han demostrado como el consumo de fructosa a diferencia de la glucosa (ambos son isocalóricos = mismas densidad energética), tiene un impacto diferente tanto en la insulina como en la leptina circulante. Siendo que la fructosa es metabolizada principalmente por el hígado ya que es el único tejido que posee el transportador de membrana GLUT 5, el cual no necesita de insulina (sin insulina elevada, se evita el llenado del adipocito y por ende liberación de leptina), y a su vez tiene una alta tasa de extracción de la misma, se podría decir que virtualmente toda la fructosa consumida terminará en el hígado. Una vez en hígado, en caso que las reservas de glucógeno hepático estén depletadas (o el sujeto en estado de inanición), la fructosa será convertida indirectamente en glucógeno (lo que hace una excelente opción en caso de necesitar repletar dichas reservas por ej.: deportista de alto rendimiento) (Douard, 2008; Kim y cols., 1988). En caso contrario la fructosa seguirá la vía glucolítica resultando en un exceso de acetyl-CoA en la mitocondria el cual producirá un exceso de citrato en el ciclo de Krebs que terminará saliendo al citoplasma del hepatocito y aportando a la de Novo lipogénesis (formación de triglicéridos, VLDL y Ácidos grasos libres), lo cual es uno de los elementos clave en la resistencia a la leptina (Banks y cols., 2004) y a la insulina con lo cual dos moléculas con las mismas calorías tienen un impacto metabólico totalmente diferente que pueden hacer una tremenda diferencia en el resultado final.
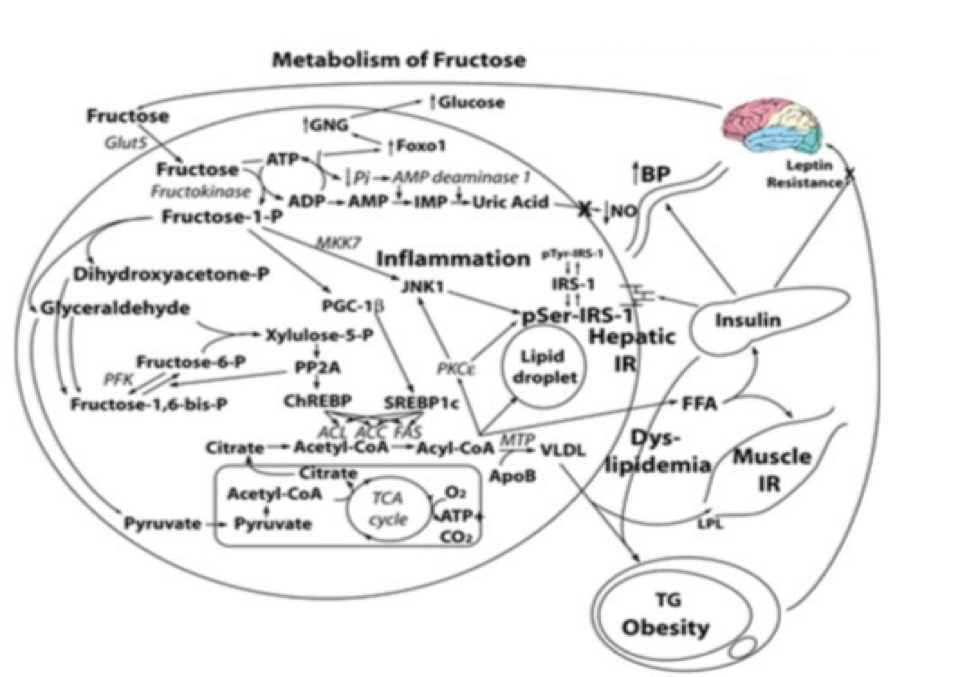
Fig. 2 : Metabolismo hepático de la fructosa. Ante una ingesta de sacarosa (50% glucosa + 50% fructosa), 20% de la glucosa y 100% de la fructosa es metabolizada en el hígado . La Fructosa induce los siguientes procesos : depleción de fosfato sustrato-dependiente, lo cual incrementa los niveles de acido úrico y contribuye a la hipertensión arterial por inhibición de la síntesis endotelial de óxido nítrico, de Novo lipogénesis y dislipidemia, esteatosis e hígado graso, resistencia a la insulina en tejido muscular y hepático, incremento de la activación de FoxO1 lo que promueve gluconeogénesis y por ende hiperglucemia, hiperinsulinemia en el sistema nervioso central lo cual genera resistencia a la leptina y una aumento en la ingesta.
Si bien los mecanismos que generan resistencia a la leptina son varios, el principal factor asociado es la inflamación y la desregulación del ciclo circadiano, ambos procesos íntimamente relacionados a la salud mitocondrial, (verdadero sensor del estado energético celular y del medio ambiente) (Langmesser y Albretch, 2006; Peek y cols., 2013; Rey y Reddy, 2013; de Git y Adan, 2015; Figueiro y cols., 2012) lo cual no sólo tiene consecuencias en la sensibilidad a la leptina sino en varios procesos metabólicos (por no decir todos).
Un claro ejemplo es el trabajo de Arble y cols. (2015) donde uno de los receptores de la leptina fue genéticamente “borrado” en un grupo de ratones lo cual desencadenó todos los signos de la resistencia a la leptina incluyendo obesidad, sin embargo esto ocurría solo cuando los ratones eran expuestos a un ciclo luz/oscuridad normal de 12hs. (se veía una exacerbada hiperglucemia en ayuno, signo de resistencia a la insulina), cuando los mismos ratones genéticamente modificados eran expuestos a 24 hs. de luz, se regulaba la hiperglucemia lo cual muestra cómo la frecuencia de onda de la luz tiene un impacto metabólico marcado, cabe estacar que los ratones son noctámbulos (se alimentan de noche) a diferencia de otros mamíferos (como los humanos), con lo cual habría que ser cauteloso con las implicancias de dicho trabajo y sus posibles aplicación en humanos. Sin embargo no son pocos los trabajos que relacionan los procesos metabólicos con el ciclo circadiano y la exposición a distintos ciclos de luz/oscuridad. (Mendoza, 2007; Yamaguchi y cols., 2013; Froy, 2010 y 2012; Johnston, 2014; Zhang y cols., 2012; Solt y cols., 2012; Shi y cols., 2013; Hartmann y cols., 2014; Oosterman y cols., 2015; Bass y Takahashi, 2010; Oishi y Katsutaka, 2013; Fonken y Nelson, 2014; Green y cols., 2008; Eckel-Mahan y Sassone-Corsi, 2013; Bellet y cols., 2010 y 2012). Los cuales muestran cómo la regulación del peso y la composición corporal trasciende las fronteras simplistas de las calorías y las mismas calorías suministradas con un “timing” o “contexto” determinado tendrán un impacto totalmente diferente.
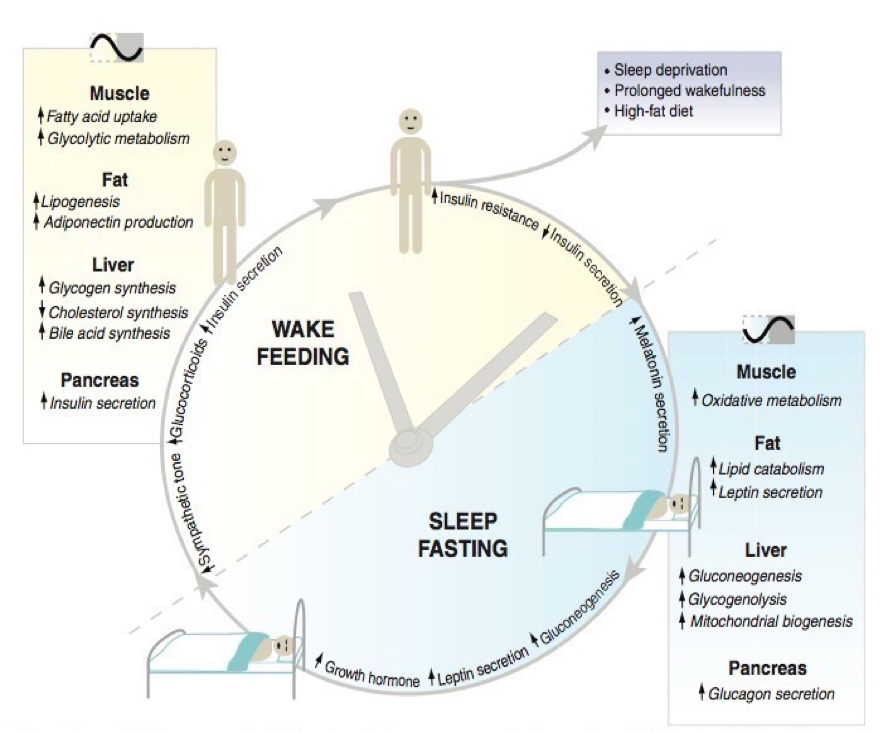
Fig. 3: el comportamiento del Reloj biológico y los procesos metabólicos según momento del día. El “reloj” coordina apropiadamente los procesos metabólicos en los tejidos periféricos según el ciclo de luz/oscuridad. Por ejemplo: el reloj periférico hepático promueve la gluconeogénesis y glucogenólisis durante el periodo de sueño/ayuno, mientras que en periodo despierto/saciedad promueve la síntesis de glucógeno y colesterol. Un adecuado funcionamiento de los “relojes” periféricos mantiene los procesos metabólicos en sincronicidad con el medio ambiente, lo que es critico para la salud de cualquier organismo. Los diferentes tejidos muestran diferentes “relojes” que controlan diferentes funciones los cuales dependen del "reloj" central en el SNC . Extraído de Baas y Takahashi, 2012.
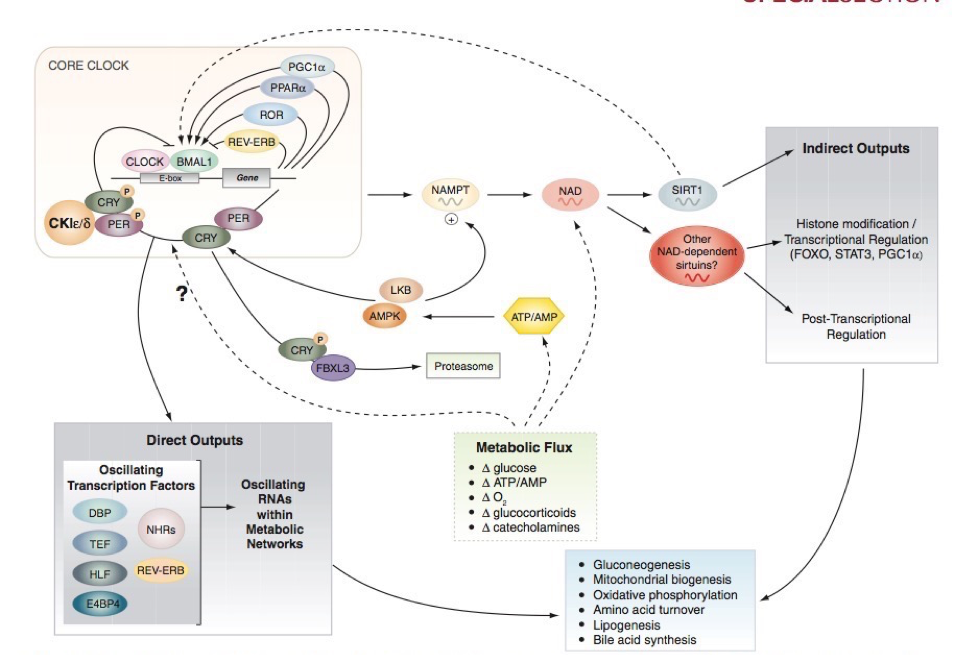
Fig. 4: “Salidas directas e indirectas del “reloj” central. El reloj Central consiste en una serie de factores de transcripción/translación en “bucle” o loop que funcionan a modo de feedback los cuales sincronizan diversos procesos metabólicos directa e indirectamente, incluyendo gluconeogénesis y metabolismo oxidativo. A su vez este “reloj” recibe información reciproca de las cascadas metabólicas (SIRT1 y AMPK), lo cual funciona como regulador y genera que el ciclo circadiano y el flujo metabolico estén atados, especialmente en tejidos periféricos.. Extraído de Bass y Takahashi, 2012.
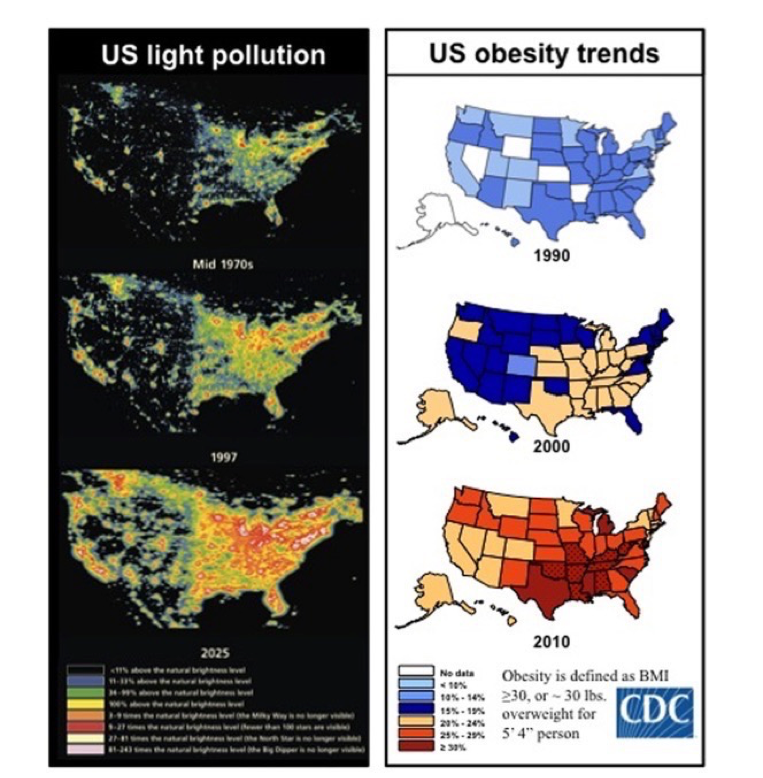
Fig. 5: Exposición a la luz durante la noche y obesidad. En el panel izquierdo la tendencia de polución lumínica en EEUU desde 1970 hasta niveles proyectados para 2025. En el panel derecho, tendencias en obesidad en EEUU en 1990, 2000 y 2010. Extraído de Fonken y Nelson, 2013.
Por ultimo y no menos importante, a medida que uno revisa la literatura, se ve obligado a tomar diferentes posturas, sin embargo hay una frase que se presenta una y otra vez: CONTEXT MATTERS o traducido al español, lo que importa es el contexto, para ejemplificarlo me gustaría incluir algunos trabajos que llamaron mucho mi atención, los dos primero muestran como la elección de diferentes macronutrientes en un contexto determinado tiene impactos diferentes y otro que ejemplifica cómo el contexto manipula el impacto de los macronutrientes.
En un trabajo de José Antonio (2014) unos 30 individuos físicamente activos y sanos (esto quiere decir sin síndrome metabólico, CONTEXTO!!) y fueron asignados ya sea a un grupo control o a un grupo de consumo elevado de proteínas, y cuando digo elevado es elevado (una de las dificultades del estudio fue la adherencia a tal ingesta) (4,4 gr/kg/día) siendo 5,5 veces mayor a la ingesta diaria recomendada durante 8 semanas (la cual me parece insuficiente, incluso absurda), también fueron instruidos a mantener la misma cantidad de carbohidratos y grasas y los hábitos de entrenamiento (Nuevamente, CONTEXTO!!). Entonces para resumir, sólo aumentaron y mucho, su consumo de proteínas, por ende las calorías aumentaron significativamente, por lo que si todas las calorías fueran lo mismo uno esperaría ver un aumento de peso.
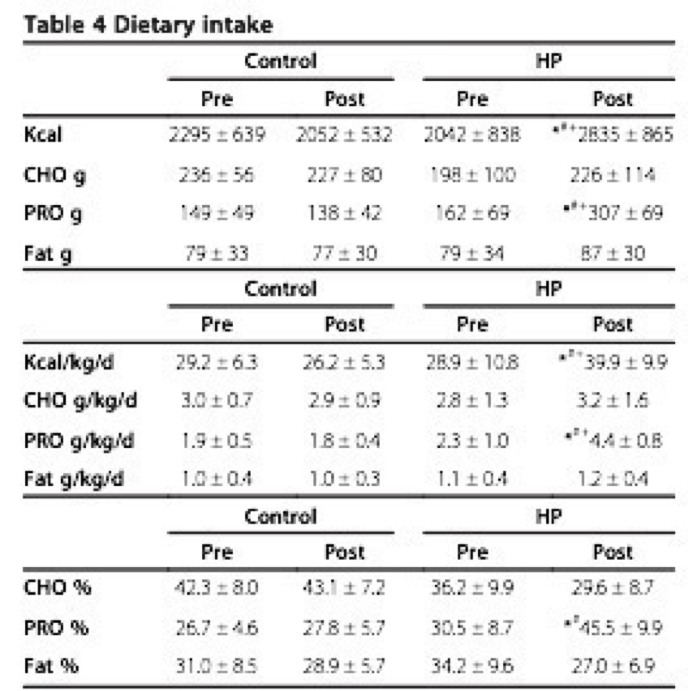
Fig.6: características tanto calóricas como de macronutrientes de los individuos del estudio. Extraído de Antonio, 2014.
Sin embargo a continuación se podrá ver la tabla de los resultados del dicho trabajo
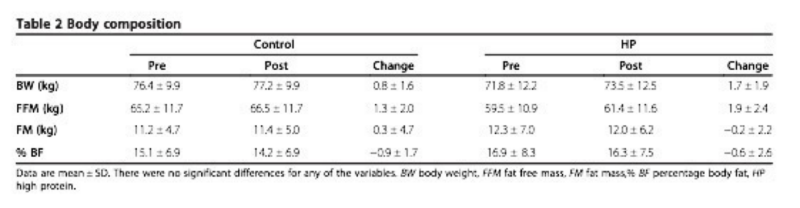
Fig.7: modificaciones en la composición corporal y el peso. Extraído de Antonio, 2014
Claramente se puede apreciar que si bien hubo un aumento en el peso corporal (1,7kg), este aumento no tiene relación con el exceso de calorías consumidas (+800/semana) y no solo eso sino que, analizando la composición corporal, el aumento fue solo a expensas del tejido muscular (+1,9kg) ya que incluso se vio una reducción del tejido adiposo (-0,2kg).
El segundo estudio en cuestión fue llevado a cabo por un grupo de investigadores (Maslow y cols., 2014) en el cual compararon los efectos de dos intervenciones dietarias en pacientes obesos y diabéticos tipo 2 o con resistencia a la insulina. Se dividió a los integrantes del estudio en 2 grupos, uno con una dieta moderada en carbohidratos baja en calorías(MCCR) (45-50% CHO, 165 gramos por día, baja en grasas y con un déficit de calorías de 500/día) y otra muy baja en carbohidratos LCK (‹50 gramos de Cho por día sin restricción de calorías), ambos grupos debían mantener su ingesta de proteínas.
Resultados: ambos grupos cumplieron con los objetivos del estudio en cuanto a macronutrientes aunque la disminución en calorías fue superior en ambos a pesar que el grupo de LCK no tenia una restricción en las mismas, esta se produjo de manera espontánea.
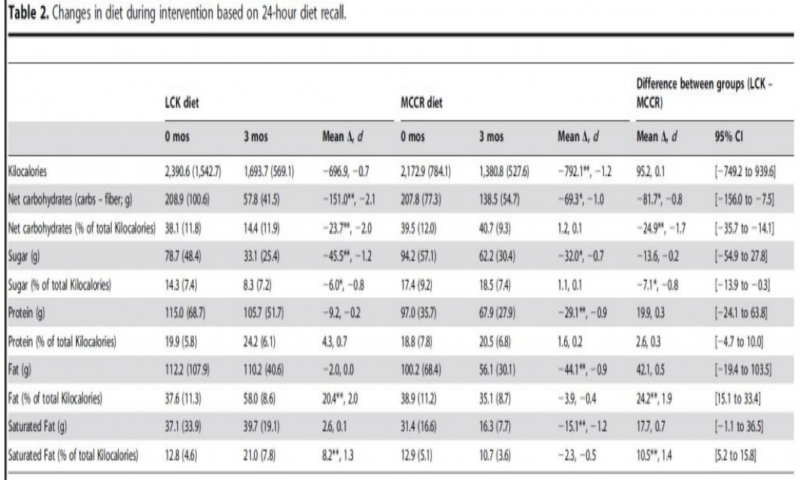
Fig.8: Cambios en las características de la dieta en ambas intervenciones. Extraído de Maslow y cols., 2014.
En cuanto al peso corporal a pesar de consumir menos calorías, los pacientes asignados a MCCR perdieron menos peso que los asignados a LCK , la composición corporal no fue medida lo cual podría sugerir que la disminución de peso podría estar asociada a perdidas de masa muscular, sin embargo el grupo LCK fue el que menos disminución en la ingesta de proteínas tuvo al tiempo que redujo menos las calorías con lo cual difícilmente sea el caso.
En cuanto a los parámetros metabólicos, proteína C reactiva (parámetro que marca inflamación) mejoró más marcadamente en la dieta LCK como así también niveles de insulinemia y glucemia en ayuno.
Por ultimo el punto mas relevante a mi juicio de este trabajo:
En el grupo LCK 7/11 pacientes (64%) redujeron su consumo de medicamentos anti diabéticos mientras que en el grupo MCCR solo 2/13 lo hicieron. Lo mas interesante de todo es que la dieta MCCR es el tipo de dieta que recomienda la ADA (American Diabetes Association) para el control de la diabetes tipo 2.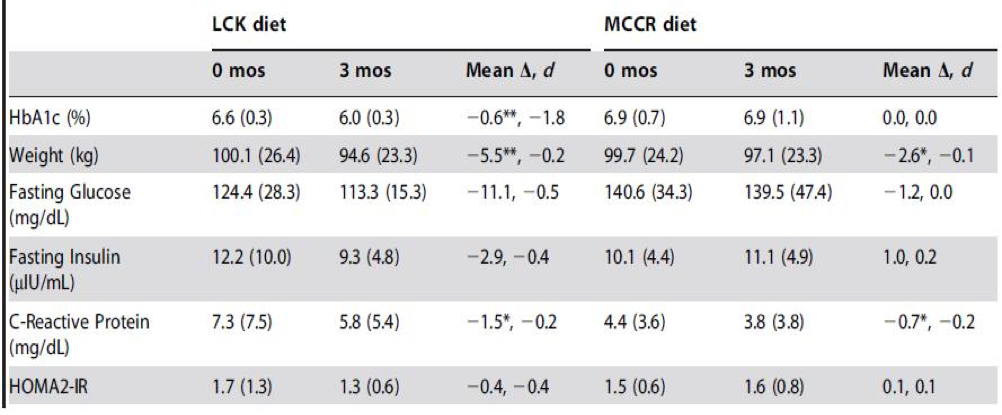
Fig.9: cambios en las variables metabólicas analizadas en ambos grupos. Extraído de Maslow y cols., 2014.
Como anteriormente comenté el contexto es una de las claves, un reciente trabajo de Nedeltcheva y cols.(2010) investigó el impacto de diferentes regímenes de sueño y por ende diferentes ciclos de luz/oscuridad sobre la composición corporal y el metabolismo .
10 sujetos con sobrepeso fueron expuestos a 14 días de restricción calórica moderada y asignados tanto a 8,5 como a 5,5 horas de sueño.
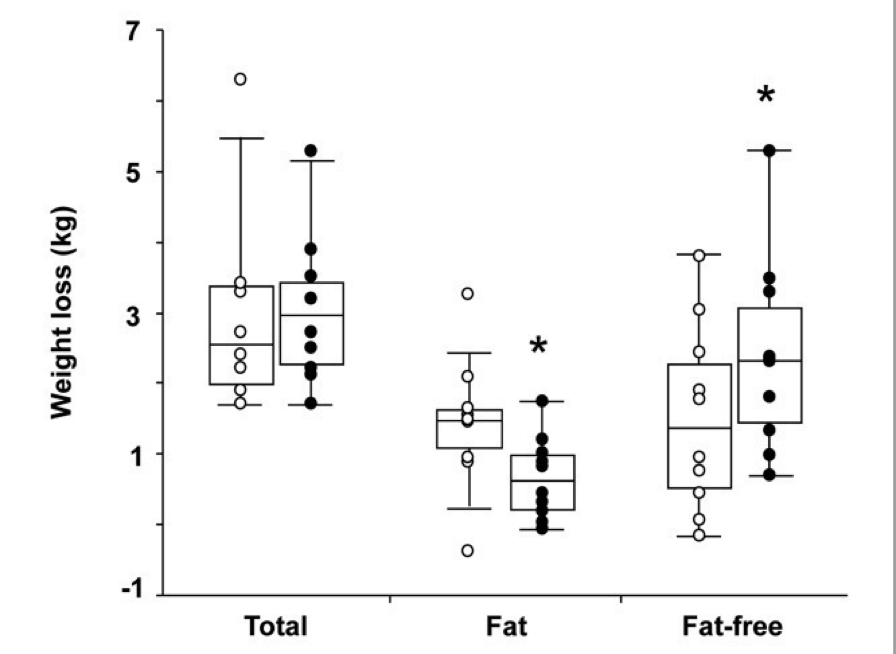
Fig.10: cambios en la composición corporal y peso en los grupos de 8,5hs (círculos claros) y de 5,5 hs. (Círculos oscuros). Extraído de Nedeltcheva y cols., 2010.
A modo de resumen, aquellos que durmieron 8,5hs. por noche perdieron la mitad de su peso en masa grasa y la mitad en masa muscular, en cambio los que fueron privados de horas de sueño, solo 1/5 del peso perdido fue tejido adiposo a pesar de mantener tanto macronutrientes como cantidad de calorías. En un contexto de ciclo luz/oscuridad y un ritmo circadiano normal la misma estrategia nutricional tiene un impacto muy diferente que en un contexto de ciclo circadiano alterado. Cabe destacar que ambas intervenciones tuvieron una disminución de tejido muscular inaceptable posiblemente debido a la baja ingesta de proteínas y de calorías que conlleva a un balance nitrogenado negativo.
Para pasarlo en limpio y mostrarlo en números hagamos el siguiente análisis. Ambos grupos consumían 1448 Kcal/día, controlado en el laboratorio, la comida pesada, etc., etc. El gasto energético se midió mediante el método de agua doblemente marcada (el Gold estándar en dicha medición con una tremenda precisión) lo cual daba en promedio para ambos grupos 2136 Kcal/día, lo que nos deja con un déficit de 688 Kcal/día. Sin embargo el grupo que dormía 8,5 hs/día perdió 50% más calorías de las que debería (347) y el grupo con sueño restringido, 20% menos(134), lo que nos lleva a una diferencia de 481 Kcal. (no incluí la cuenta completa para no marear al lector) que “mágicamente” desaparecieron.
Como conclusión final a la hora de garantizar el éxito de una intervención nutricional, la biología no entiende de calorías, sino de información la cual integrará para su correcta regulación y funcionamiento expresada en una adecuada señalización que generará o no los resultados deseados. Partiendo de esta base las calorías son solo una parte de la información, solo una página de un libro mucho mas grande con varios capítulos cuyo titulo es CONTEXTO.
Estos temas y muchos más trataremos en nuestro próximo Webinar
Webinar de Estrategias Nutricionales Para el Manejo de la Composición Corporal.
Referencias
.Ahima, R.S. (2008) Revisiting leptin’s role in obesity and weight loss. The Journal of clinical investigation,
.Allison, M.B. & Myers, M.G. (2014) 20 YEARS OF LEPTIN: Connecting leptin signaling to biological function. Journal of Endocrinology, 223, T25-T35.
.Antonio, J. et al. (2014) The effects of consuming a high protein diet (4.4 g/kg/d) on body composition in resistance-trained individuals. Journal of the International Society of Sports Nutrition, 11, 19.
.Arble, D.M. et al. (2015) The melanocortin-4 receptor integrates circadian light cues and metabolism. Endocrinology, 156, 1685-1691.
.Baer, D.J., Gebauer, S.K. & Novotny, J.A. (2012) Measured energy value of pistachios in the human diet. Br J Nutr, 107, 120-125.
.Banks, W.A. et al. (2004) Triglycerides induce leptin resistance at the blood-brain barrier. Diabetes, 53, 1253-1260.
.Bass, J. & Takahashi, J.S. (2010) Circadian integration of metabolism and energetics. Science, 330, 1349-1354.
.Bellet, M.M. et al. (2011) The time of metabolism: NAD+, SIRT1, and the circadian clock. Cold Spring Harb Symp Quant Biol, 76, 31-38.
.Bellet MM, Sassone-Corsi P. (2010) Mammalian circadian clock and metabolism - the epigenetic link. Journal of Cell Science. ;123(22):3837-3848.
.Charles H. Bennett, "Demons, Engines and the Second Law", Scientific American, pp.108-116 (November, 1987).
.Considine, R.V. et al. (1996) Serum immunoreactive-leptin concentrations in normal-weight and obese humans. N Engl J Med, 334, 292-295.
.Coutinho, T. et al. (2013) Combining body mass index with measures of central obesity in the assessment of mortality in subjects with coronary disease: role of ?normal weight central obesity?. Journal of the American College of Cardiology, 61, 553-560.
.Douard, V. & Ferraris, R.P. (2008) Regulation of the fructose transporter GLUT5 in health and disease. Am J Physiol Endocrinol Metab, 295, E227-37.
.Dyck, D.J. (2005) Leptin sensitivity in skeletal muscle is modulated by diet and exercise. Exercise and sport sciences reviews, 33, 189-194.
.Eckel-Mahan, K. & Sassone-Corsi, P. (2013) Metabolism and the circadian clock converge. Physiol Rev, 93, 107-135.
.Feinman, R.D. & Fine, E.J. (2003) Thermodynamics and metabolic advantage of weight loss diets. Metab Syndr Relat Disord, 1, 209-219.
.Feinman, R.D. & Fine, E.J. (2004) A calorie is a calorie” violates the second law of thermodynamics. Nutr J, 3, 9.
.Feinman, R.D. & Fine, E.J. (2007) Nonequilibrium thermodynamics and energy efficiency in weight loss diets. Theor Biol Med Model, 4, 27.
.Figueiro, M.G., Plitnick, B. & Rea, M.S. (2012) Light modulates leptin and ghrelin in sleep-restricted adults. International journal of endocrinology, 2012,
.Fine, E.J. & Feinman, R.D. (2004) Thermodynamics of weight loss diets. Nutr Metab (Lond), 1, 15.
.Fonken, L.K. & Nelson, R.J. (2014) The effects of light at night on circadian clocks and metabolism. Endocrine reviews, 35, 648-670.
.Friedman, J.M. (2000) Obesity in the new millennium. Nature, 404, 632-634.
.Friedman, J.M. & Halaas, J.L. (1998) Leptin and the regulation of body weight in mammals. Nature, 395, 763-770.
.Froy, O. (2010) Metabolism and circadian rhythms?implications for obesity. Endocrine reviews, 31, 1-24.
.Froy, O. (2012) Circadian rhythms and obesity in mammals. ISRN obesity, 2012,
.Gannon, M.C. & Nuttall, F.Q. (2006) Control of blood glucose in type 2 diabetes without weight loss by modification of diet composition. Nutr Metab (Lond), 3, 16.
.German, J.P. et al. (2010) Leptin deficiency causes insulin resistance induced by uncontrolled diabetes. Diabetes, 59, 1626-1634.
.Git, K.C.G. & Adan, R.A.H. (2015) Leptin resistance in diet induced obesity: the role of hypothalamic inflammation. Obesity Reviews, 16, 207-224.
.Green, C.B., Takahashi, J.S. & Bass, J. (2008) The meter of metabolism. Cell,
.Guth, E. (2014) JAMA patient page. Healthy weight loss. JAMA, 312, 974.
.Hall, K.D. et al. (2013) Dynamics of childhood growth and obesity: development and validation of a quantitative mathematical model. The Lancet Diabetes & Endocrinology, 1, 97-105.
.Hall, K.D. et al. (2012) Energy balance and its components: implications for body weight regulation. The American journal of clinical nutrition, 95, 989-994.
.Hall, K.D. et al. (2011) Quantification of the effect of energy imbalance on bodyweight. The Lancet, 378, 826-837.
.Hamer, M. & Stamatakis, E. (2012) Metabolically healthy obesity and risk of all-cause and cardiovascular disease mortality. The Journal of Clinical Endocrinology & Metabolism, 97, 2482-2488.
.Harfmann, B.D., Schroder, E.A. & Esser, K.A. (2014) Circadian rhythms, the molecular clock, and skeletal muscle. Journal of biological rhythms, 0748730414561638.
.Harris, R.B. (1990) Role of set-point theory in regulation of body weight. FASEB J, 4, 3310-3318.
.Heymsfield, S.B. et al. (2007) Why do obese patients not lose more weight when treated with low-calorie diets? A mechanistic perspective. The American journal of clinical nutrition, 85, 346-354.
.Hinkle, W. et al. (2013) Effects of reduced weight maintenance and leptin repletion on functional connectivity of the hypothalamus in obese humans. PloS one, 8, e59114.
.Jakubowicz, D. et al. (2013) High caloric intake at breakfast vs. dinner differentially influences weight loss of overweight and obese women. Obesity, 21, 2504-2512.
.Jequier, E. (1993) Body weight regulation in humans: the importance of nutrient balance. Physiology,
.Jéquier, E. (2002) Leptin signaling, adiposity, and energy balance. Annals of the New York Academy of Sciences,
.Jequier, E. (2002) Pathways to obesity. International Journal of Obesity,
.Jéquier, E. & Schutz, Y. (1988) Energy expenditure in obesity and diabetes. Diabetes/metabolism reviews, 4, 583-593.
.Jéquier, E. & Tappy, L. (1999) Regulation of body weight in humans. Physiological reviews, 79, 451-480.
.Johnston, J.D. (2014) Physiological links between circadian rhythms, metabolism and nutrition. Experimental physiology, 99, 1133-1137.
.Jung, C.H. & Kim, M.-S. (2013) Molecular mechanisms of central leptin resistance in obesity. Archives of pharmacal research, 36, 201-207.
.Keim, N.L., Stern, J.S. & Havel, P.J. (1998) Relation between circulating leptin concentrations and appetite during a prolonged, moderate energy deficit in women. The American journal of clinical …,
.Kim, H.S. et al. (1988) Effects of several simple sugars on serum glucose and serum fructose levels in normal and diabetic subjects. Diabetes Res Clin Pract, 4, 281-287.
.Kohsaka, A. & Bass, J. (2007) A sense of time: how molecular clocks organize metabolism. Trends Endocrinol Metab, 18, 4-11.
.Kramer, C.K., Zinman, B. & Retnakaran, R. (2013) Are metabolically healthy overweight and obesity benign conditions?: A systematic review and meta-analysis. Annals of internal medicine, 159, 758-769.
.Langmesser, S. & Albrecht, U. (2006) Life time-circadian clocks, mitochondria and metabolism. Chronobiol Int, 23, 151-157.
.Lee, J.M., Kim, Y. & Welk, G.J. (2014) Validity of consumer-based physical activity monitors. Med Sci Sports Exerc, 46, 1840-1848.
.Leibel, R.L., Rosenbaum, M. & Hirsch, J. (1995) Changes in energy expenditure resulting from altered body weight. N Engl J Med, 332, 621-628.
.Group, L.A.H.E.A.D.R. (2013) Cardiovascular effects of intensive lifestyle intervention in type 2 diabetes. The New England journal of medicine, 369, 145.
.Ludwig, D.S. & Friedman, M.I. (2014) Increasing adiposity: consequence or cause of overeating. JAMA, 311, 2167-2168.
.Lustig, R.H. (2006) The ‘skinny’ on childhood obesity: how our western environment starves kids’ brains. Pediatr Ann, 35, 898-902, 905.
.Lustig, R.H. (2013) Fructose: it’s “alcohol without the buzz”. Adv Nutr, 4, 226-235.
.Mandrup-Poulsen, T. (2013) Type 2 diabetes mellitus: a metabolic autoinflammatory disease. Dermatologic clinics, 31, 495-506.
.Margetic, S. et al. (2002) Leptin: a review of its peripheral actions and interactions. International journal of obesity and related metabolic disorders: journal of the International Association for the Study of Obesity, 26, 1407-1433.
.Mendoza, J. (2007) Circadian clocks: setting time by food. J Neuroendocrinol, 19, 127-137.
.Myers, M.G. et al. (2012) Challenges and opportunities of defining clinical leptin resistance. Cell Metab, 15, 150-156.
.Nedeltcheva, A.V. et al. (2010) Insufficient sleep undermines dietary efforts to reduce adiposity. Annals of internal medicine, 153, 435-441.
.Novotny, J.A., Gebauer, S.K. & Baer, D.J. (2012) Discrepancy between the Atwater factor predicted and empirically measured energy values of almonds in human diets. The American journal of clinical nutrition, 96, 296-301.
.Ochner, C.N. et al. (2013) Biological mechanisms that promote weight regain following weight loss in obese humans. Physiol Behav, 120, 106-113.
.Oishi, K. & Itoh, N. (2013) Disrupted daily light-dark cycle induces the expression of hepatic gluconeogenic regulatory genes and hyperglycemia with glucose intolerance in mice. Biochem Biophys Res Commun, 432, 111-115.
.Oosterman, J.E. et al. (2015) Impact of nutrients on circadian rhythmicity. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology, 308, R337-R350.
.Peek, C.B. et al. (2013) Circadian clock NAD+ cycle drives mitochondrial oxidative metabolism in mice. Science, 342, 1243417.
.Rey, G. & Reddy, A.B. (2013) Protein acetylation links the circadian clock to mitochondrial function. Proc Natl Acad Sci U S A, 110, 3210-3211.
.Ricquier, D. & Bouillaud, F. (2000) The uncoupling protein homologues: UCP1, UCP2, UCP3, StUCP and AtUCP. Biochem J, 345 Pt 2, 161-179.
.Saslow, L.R. et al. (2014) A randomized pilot trial of a moderate carbohydrate diet compared to a very low carbohydrate diet in overweight or obese individuals with type 2 diabetes mellitus or prediabetes. PloS one, 9, e91027.
.Schwartz, M.W. et al. (2000) Central nervous system control of food intake. Nature, 404, 661-671.
.Seufert, J. (2004) Leptin effects on pancreatic -cell gene expression and function. Diabetes, 53, S152-S158.
.Shah, N.R. & Braverman, E.R. (2012) Measuring adiposity in patients: the utility of body mass index (BMI), percent body fat, and leptin. PloS one, 7, e33308.
.Shapiro, A. et al. (2008) Fructose-induced leptin resistance exacerbates weight gain in response to subsequent high-fat feeding. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology, 295, R1370-R1375.
.Shi, S.- et al. (2013) Circadian disruption leads to insulin resistance and obesity. Current Biology, 23, 372-381.
.Shook, R.P., Hand, G.A. & Blair, S.N. (2014) Top 10 research questions related to energy balance. Research quarterly for exercise and sport, 85, 49-58.
.Solt, L.A. et al. (2012) Regulation of circadian behaviour and metabolism by synthetic REV-ERB agonists. Nature, 485, 62-68.
.Song, Y. et al. (2007) Comparison of usefulness of body mass index versus metabolic risk factors in predicting 10-year risk of cardiovascular events in women. The American journal of cardiology, 100, 1654-1658.
.Song, Y. et al. (2007) Comparison of usefulness of body mass index versus metabolic risk factors in predicting 10-year risk of cardiovascular events in women. The American journal of cardiology, 100, 1654-1658.
.Speakman, J.R. et al. (2011) Set points, settling points and some alternative models: theoretical options to understand how genes and environments combine to regulate body adiposity. Disease models & mechanisms, 4, 733-745.
.Speakman, J.R., Levitsky, D.A. & Allison…, D.B. (2011) Set points, settling points and some alternative models: theoretical options to understand how genes and environments combine to regulate body adiposity. Disease models & …,
.St Clair Gibson, A. et al. (2005) Metabolic setpoint control mechanisms in different physiological systems at rest and during exercise. J Theor Biol, 236, 60-72.
.Sumithran, P. & Proietto, J. (2013) The defence of body weight: a physiological basis for weight regain after weight loss. Clin Sci (Lond), 124, 231-241.
.Teff, K.L. et al. (2004) Dietary fructose reduces circulating insulin and leptin, attenuates postprandial suppression of ghrelin, and increases triglycerides in women. The Journal of Clinical Endocrinology & Metabolism, 89, 2963-2972.
.Urban, L.E. et al. (2010) The accuracy of stated energy contents of reduced-energy, commercially prepared foods. Journal of the American Dietetic Association, 110, 116-123.
.Westerterp, K.R., Wilson, S.A. & Rolland, V. (1999) Diet induced thermogenesis measured over 24h in a respiration chamber: effect of diet composition. Int J Obes Relat Metab Disord, 23, 287-292.
.Wildman, R.P. et al. (2008) The obese without cardiometabolic risk factor clustering and the normal weight with cardiometabolic risk factor clustering: prevalence and correlates of 2 phenotypes among the US population (NHANES 1999-2004). Archives of internal medicine, 168, 1617-1624.
.Wing, R.R. et al. (1996) Relationship between weight loss maintenance and changes in serum leptin levels. Hormone and metabolic research= Hormon-und Stoffwechselforschung= Hormones et metabolisme, 28, 698-703.
.Yamaguchi, Y. et al. (2013) Mice genetically deficient in vasopressin V1a and V1b receptors are resistant to jet lag. Science, 342, 85-90.
.Zhang, L. et al. (2012) PKC participates in food entrainment by regulating BMAL1. Proceedings of the National Academy of Sciences, 109, 20679-20684.