
Entrenamiento de la Fuerza de Alta Intensidad y Metabolismo Muscular durante el Ejercicio Prolongado
C. Goreham1, H. J Green1, M. Ball-Burnet1 y D. Ranney1
Department of Kinesiology, McMaster University, Hamilton, Ontario, Canada L8S 4K1.
Artículo publicado en el journal PubliCE, Volumen 0 del año 1999.
Publicado 3 de diciembre de 2003
Resumen
Palabras clave: entrenamiento de la fuerza, ejercicio cíclico, adaptación metabólica
INTRODUCCION
La realización regular de ejercicio prolongado resulta en una bien definida serie de adaptaciones en el metabolismo muscular y la utilización de sustratos. Después del entrenamiento, un protocolo de ejercicio submáximo dado puede ser realizado con una menor disminución en el potencial de los fosfatos de alta energía, un menor incremento en la glucogenólisis y la glucólisis, y un incremento en la utilización de grasas (20). Acompañando a las adaptaciones en el metabolismo muscular y la selección de sustratos están las adaptaciones en la capilarización y la organización enzimática en el músculo. Estas incluyen un incremento en el número de capilares por fibra muscular y un incremento en el potencial mitocondrial (1, 20). Se ha postulado que estas adaptaciones representan los mecanismos fundamentales subyacentes a los cambios en el funcionamiento metabólico de los músculos (1, 20). Sin embargo, no están reconocidos los cambios que ocurren en la hipertrofia muscular. En los varones, los incrementos en el área de las fibras pueden acercarse al 20% en el vasto lateral con entrenamiento de resistencia en bicicleta (1), un incremento que no es substancialmente diferente del incremento de 14-24% que puede ser inducido en el mismo músculo con muchas semanas de entrenamiento de fuerza de alta intensidad (HRT; ver Ref. 24). Una cuestión que se encuentra bajo discusión es las consecuencias de los efectos independientes de la hipertrofia muscular en la explicación de las alteraciones metabólicas observadas después del entrenamiento de resistencia.
Existen fundamentos razonables para postular que la hipertrofia que acompaña al entrenamiento pude, por ella misma, influenciar eventos metabólicos en el músculo. El incremento en el área de sección cruzada muscular que es inducido por el HRT es también acompañado por un incremento en las capacidades de generación de fuerza voluntaria máxima (MVC; para revisión ver Ref. 24). Para realizar una tarea repetitiva a una producción de fuerza constante, después del HRT, disminuye el porcentaje de fuerza generada con cada repetición. En la ausencia de cambios en la estrategia de reclutamiento y asumiendo que la hipertrofia se extiende a ambos tipos de fibras (tipo I y tipo II; ver Ref. 24), se reduce el porcentaje MVC por fibra. Es bien conocido que al menos para los grupos musculares locales, el porcentaje MVC es un importante determinante de la contribución de las vías metabólicas para la generación del ATP. A medida que se incrementa la MVC, se incrementa también el potencial de fosforilación, así como la glucólisis (2, 8).
Sin embargo, para estas situaciones para producir los efectos postulados sobre el metabolismo muscular, la hipertrofia de las fibras no debe estar acompañada por una disminución en la densidad capilar o en el potencial mitocondrial. Aunque hay evidencia que sugiere que la hipertrofia puede resultar en un incremento desproporcionado en el área de las fibras relativa a la neoformación capilar y a la biogénesis mitocondrial (24), los trabajos recientes no indican una disminución de estos parámetros con la hipertrofia de las fibras inducida por el HRT (17, 35).
En este estudio, nosotros examinamos los efectos dependientes del tiempo de los incrementos en el área de sección cruzada de las fibras, inducidos por medio del HRT, sobre el metabolismo muscular durante una tarea estandarizada en bicicleta. Nosotros hipotetizamos que la hipertrofia muscular, que ocurre en la ausencia de cambios en la densidad capilar y el potencia mitocondrial, resultaría, al menos cualitativamente, en alteraciones metabólicas similares a aquellas observadas en el entrenamiento prolongado. Estas incluyen un mayor potencial de fosforilación más protegido, una reducción en la acumulación de lactato, y una reducción en la tasa de depleción de glucógeno muscular. Ha sido realizado otro estudio que examinó las alteraciones en el tipo de fibras usando procedimientos histoquímicos.
MATERIALES Y MÉTODOS
Sujetos
7 hombres sanos desentrenados y ocasionalmente activos se ofrecieron para participar en el estudio. La media y desvío estándar de la muestra para la edad y el peso fueron de 20.0±0.7 años y 88.7±7.0 kg. El estudio fue aprobado por la Oficina de Investigación con Humanos luego de la presentación de todos los detalles de los experimentos. Como es requerido, los procedimientos y los riesgos fueron completamente explicados a cada participante antes de obtener el consentimiento por escrito.
Diseño del Estudio
El programa HRT utilizado implico entrenar 3 veces por semana por un período de 12 semanas. Fueron empleados 3 ejercicios durante cada sesión de entrenamiento, todos destinados para cargar los músculos cuadriceps. Durante cada sesión de entrenamiento fueron realizadas 3 series, con 6 a 8 repeticiones máximas (RM) para cada serie, para cada ejercicio (sentadilla, prensa de piernas inclinada, y extensión de rodilla). Todas las sesiones de entrenamiento, las cuales fueron supervisadas, también incluyeron una serie de entrada en calor de 10 repeticiones de cada ejercicio, a ≈ 50% de las 6-8 RM de cada participante. Este programa fue modelado después de un programa empleado por Staron et al. (34), el que se demostró que produce una apreciable hipertrofia de los tipos de fibras. El programa de entrenamiento fue dividido en tres segmentos (0-4 semanas, 4-7 semanas, y 7-12 semanas). Antes del comienzo del entrenamiento, y después de cada segmento del entrenamiento, fueron realizadas una serie de evaluaciones para evaluar los efectos del entrenamiento. Así como también antes del comienzo de un nuevo segmento de entrenamiento, los pesos empleados en el entrenamiento fueron incrementados para mantener el estímulo de entrenamiento de 6-8 RM. Los pesos promedio empleados durante el segmento de entrenamiento inicial fueron de 199±20, 364±22, 90±6 kg para la sentadilla, prensa de piernas, y extensión de rodillas, respectivamente. Durante el segmento de entrenamiento final, los pesos empleados se incrementaron en un 62% para la sentadilla, 72% para la prensa de piernas, y 56% para la extensión de rodillas.
Al menos 1 semana antes del comienzo del estudio y después de cada segmento de entrenamiento, fueron administradas 4 evaluaciones diferentes. Una evaluación utilizó un ejercicio en bicicleta progresivo hasta la fatiga para la medición de la potencia aeróbica pico (VO2 pico), mientras que una segunda evaluación implico ciclismo continuo prolongado, inicialmente por 30 min al 57% del VO2 pico, seguido de un máximo de 30 min o hasta la fatiga al 72% del VO2 pico. La intensidad y duración del ejercicio fue usada para investigar tanto las adaptaciones metabólicas musculares como las adaptaciones de los sustratos (glucógeno) al HRT. Antes del entrenamiento, el tiempo de ejercicio promedio fue de 56.4±2.9 min. Las adaptaciones metabólicas fueron examinadas a las mismas intensidades absolutas a través de todo el estudio, a saber 151±4.7 y 202±6.8 W. Fue usada una tercera evaluación para medir la MVC que podía ser generada durante las extensiones de rodillas mientras que era usada una cuarta evaluación para monitorear los cambios en la potencia mecánica sobre un rango de diferentes ajustes de la resistencia de la bicicleta. Dada la especificidad de las adaptaciones neurales que parecen acompañar al HRT (6), este último protocolo fue considerado esencial para evaluar el grado en el cual los incrementos en la MVC se traducían al rendimiento sobre la bicicleta. Como fue enfatizado, el ejercicio de ciclismo fue usado para examinar las adaptaciones metabólicas que ocurrían en respuesta al HRT. Al menos una semana antes del comienzo del entrenamiento, fue realizada la evaluación progresiva de ciclismo. Esta evaluación fue seguida por la evaluación de ciclismo prolongado, realizada por lo menos dos días después de la evaluación progresiva. La evaluación de MVC y las evaluaciones de potencia mecánica fueron administradas en días separados una de la otra y separadas de las evaluaciones submáximas. Después de cada segmento de entrenamiento, la evaluación submáxima fue administrada primero, ≈24-36 h después de la última sesión de entrenamiento, seguida por las evaluaciones de MVC, potencia mecánica, y la evaluación progresiva de ciclismo. Las evaluaciones de MVC y potencia mecánica fueron administradas sobre un período de 1 a 2 días, en días separados de la evaluación progresiva de ciclismo. Fueron necesarios 3-4 días para realizar todas las evaluaciones. Todas las evaluaciones fueron realizadas a temperaturas entre 24 y 26 ºC y a una humedad relativa entre 49-50 %.
Protocolos de Evaluación
Ciclismo Progresivo
La medición de VO2 pico fue obtenida usando un cicloergómetro frenado eléctricamente (modelo Siemens 380B B) a una velocidad de 60 revoluciones/min. La carga de trabajo inicial fue 16.3 W y fue incrementada en 16.3 W cada minuto hasta la fatiga. El aire espirado fue recolectado y analizado por medio de un sistema de recolección de gases computarizado on-line IBM-PC como fue previamente descrito (21), que consistía de un Digital Pneumotach modelo Hewlett Packard 47363A y un Analizador de Gases Médico modelo Beckman 6B-2. El intercambio respiratorio de gases fue calculado sobre segmentos de 30 s, con el VO2 pico representando el mayor valor registrado.
Ciclismo Prolongado
La evaluación de ciclismo prolongado fue realizada usando el mismo sistema de ciclismo y de recolección de gases que fue usado en la evaluación progresiva. El ajuste del trabajo estuvo basado en los valores del VO2 pico obtenidos para cada individuo y fueron diseñados para producir respuestas de ≈ 60 y 70 % del VO2 pico durante cada segmento. Durante la evaluación, el intercambio respiratorio de gases fue medido durante los 5 min finales de cada carga de trabajo donde fue aplicable. Las mediciones de frecuencia cardiaca (HR) usando un electrocardiograma Cambridge modelo VS-4, fueron también registradas en ese tiempo. Además, las biopsias musculares fueron extraídas a partir del músculo vasto lateral en reposo y al final de cada fase de la evaluación submáxima, i.e., a los 30 y 60 min (o justo antes del comienzo de la fatiga, dependiendo que ocurriera primero). El tejido muscular fue rápidamente extraído en la finalización del ejercicio a partir de incisiones previamente realizadas, y fue inmediatamente metido en nitrógeno líquido y almacenado a -80º C hasta los análisis. Una segunda biopsia fue obtenida a partir del mismo sitio y fue usada para mediciones histoquímicas. Para cada segmento de entrenamiento, fue empleado un procedimiento idéntico, con el muestreo en puntos de tiempo similares. Las biopsias (12 en total) fueron equilibradas entre las piernas y fueron aleatorias de acuerdo al orden para el ejercicio y el entrenamiento. Este procedimiento aseguro que, después de un segmento de entrenamiento dado, las muestras de tejido fueran extraídas a partir de las dos piernas durante el ejercicio. Las evaluaciones tuvieron lugar aproximadamente a la misma hora del día y aproximadamente 4 h después de la ingestión de cualquier comida.
Potencia Mecánica
Las mediciones de fuerza velocidad fueron conducidas en una serie de diferentes ajustes de resistencia para determinar los efectos del HRT en un rango de determinaciones de velocidad y producciones de potencia. El fundamento detrás de este procedimiento fue determinar si el programa HRT resultaría o no en un incremento en la producción de potencia a velocidades en la región que fue usada durante la evaluación prolongada. Estas evaluaciones fueron conducidas en un cicloergómetro Monach, equipado con un registrador de revoluciones conectado a un registrador de banda (Hewlett Packard modelo 7402 A). Fueron empleados cuatro ajustes de resistencia diferentes y aleatoriamente asignados de acuerdo al orden durante cada sesión de evaluación. En cada caso, un período de entrada en calor de 2 min a una carga cero precedió cada medición. No hubo ninguna interrupción entre la entrada en calor y la evaluación. La velocidad pico para cada ajuste de resistencia fue calculada a partir de registros obtenidos durante el límite de tiempo de 10 s permitido para cada evaluación.
MVC
Las mediciones de MVC generadas por el cuadriceps fueron medidas durante las extensiones de rodilla por métodos previamente descritos (12).
Análisis Bioquímicos
El tejido muscular fue analizado para fosfatos de alta energía y metabolitos relacionados, nucleótidos de adenina, glucógeno, y una serie de intermediarios glucolíticos. Con la excepción de los nucleótidos de adenina (ATP, ADP, AMP) e IMP, fueron usadas, modificadas en nuestro laboratorio (16), técnicas fluorométricas (26), para determinar las respectivas concentraciones. Los nucleótidos de adenina e IMP fueron medidos en la misma homogeneización con el uso de técnicas de cromatografía líquida de alta eficacia (22) como se detallo previamente (15). Todos los valores fueron corregidos para creatina total (TCr) para cada sujeto, usando el promedio de todas las biopsias para cada individuo, para ajustar para la contaminación con sangre y tejido conectivo. La glucosa, piruvato, y lactato fueron también corregidos para TCr, aunque los mismos estaban en el espacio extracelular, ya que la corrección corregiría al menos los contaminantes musculares y proveería más valores estables (13).
Las mediciones de distribución de tipo de fibras (tipo I, IIA, IIAB y IIB), área de las fibras, capilares, y actividad de la succinato dehidrogenasa (SDH) fueron realizadas sobre muestras histoquímicas (11). En este artículo, nosotros reportamos el área promedio de las fibras, la cual esta basada en la distribución porcentual de cada tipo de fibra en las respectivas áreas (17).
Procedimientos Estadísticos
Para los datos metabólicos, fue usado un análisis ANOVA a dos vías para mediciones repetidas para determinar los efectos del entrenamiento y el ejercicio. Para las mediciones de MVC, fuerza velocidad, y VO2 pico fue empleado un análisis ANOVA a una vía. Fue realizado un procedimiento Student Newman-Keuls para comparar medias específicas donde era indicada la significancia. El nivel de confianza fue establecido al 95% para todas las comparaciones.
RESULTADOS
Índices de Rendimiento
Además de monitorear los incrementos en las 6-8 RM para los ejercicios individuales usados en el programa HRT, nosotros también registramos criterios de rendimiento adicionales. Fueron empleadas dos mediciones, a saber MVC y producción de potencia determinada en un cicloergómetro. En las semanas 0, 4, 7 y 12, la MVC (en newtons) fue de 682±41, 646±45, 696±44, y 735±53. Fueron observados incrementos en la MVC, pero no hasta el segmento final de entrenamiento. En la semana 12, ocurrió un incremento de 7.8% en la MVC, en comparación con los valores pre-entrenamiento. También se encontró que la producción de potencia, determinada a diferentes ajustes de resistencia, también se incremento en la semana 12 de entrenamiento, pero solo para los mayores ajustes de resistencia (Tabla 1). A estas cargas de trabajo, las cuales produjeron velocidades de pedaleo equivalentes a aquellas usadas en la evaluación de ciclismo prolongado, fueron observados incrementos de 21 y 57%.
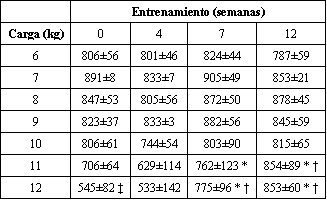
Tabla 1. Alteraciones en la potencia mecánica en bicicleta con el entrenamiento
de fuerza de alta intensidad. Los valores están expresados como medias±DS, los
mismos están expresados en W; n=7 sujetos. * Significativamente diferente
(p<0.05) con respecto a la semana 4; † Significativamente diferente (p<0.05) con
respecto a la semana 0, ‡ Significativamente diferente (p<0.05) con respecto a
6, 7, 8, 9, 10, y 11 kg.
VO2 pico
El HRT no resulto en incrementos en el VO2 pico, pico de HR, o ventilación pico (VE). Antes del entrenamiento (semana 0) y en las semanas 0, 4, 7 y 12 del HRT, los valores fueron 3.53±0.10, 3.40±0.11, 3.53±0.15, y 3.57±0.15 L/min, respectivamente. Los valores comparables para HR pico fueron 196±2.4, 193±1.4, 196±1.8, y 191±2.2 lat/min. Así como con el VO2 pico, las diferencias no fueron significativas.
Metabolismo en el Ejercicio Submáximo
La tasa de consumo de oxígeno (VO2) medida durante la evaluación submáxima de dos etapas, conducida al 59 y 72% del VO2 pico, no fue afectada por el entrenamiento (Tabla 2). Similarmente, no fueron encontradas diferencias para la tasa de producción de dióxido de carbono (VCO2), VE, e índice de intercambio respiratorio (RER) sin tener en cuenta la duración del ejercicio. Como se esperaba fueron observados incrementos progresivos (min 0, 30, 56) con cada nivel de ejercicio. La única excepción fue el RER, donde el incremento observado desde el min 0 al 30 no continuo aumentando desde el min 30 al 56. El programa de entrenamiento también fallo en modificar la respuesta de la HR a cualquiera de las dos intensidades de ejercicio. Antes del entrenamiento, la HR fue de 163±3.1 y 195±3.6 lat./min al final de cata etapa. Después del entrenamiento, los respectivos valores fueron 154±4.3 y 187±5.4 lat./min.
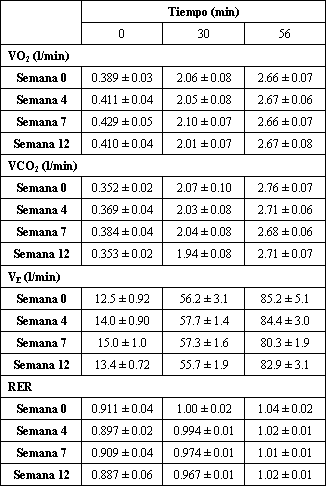
Tabla 2. Entrenamiento de fuerza de alta intensidad e intercambio de gases
durante el ejercicio submáximo. Los valores están expresados como medias±DS; n=7
sujetos. VO2, consumo de oxígeno; VCO2, producción de
dióxido de carbono; VE, ventilación en L/min BTPS; RER, índice de intercambio
respiratorio. Un efecto principal (p<0.05) del tiempo fue encontrado en todas
las variables. Para el VO2, VCO2, VE, 0<30<56 min, para el
RER, 0<30=56 min.
Las concentraciones de nucleótidos de adenina (ATP, ADP, AMP) y las concentraciones totales de nucleótidos de adenina no fueron alteradas ni con el ejercicio ni con el entrenamiento (Tabla 3). Sin embargo, las concentraciones de IMP fuero alteradas. Con la excepción de la semana 0, fueron observados incrementos en el IMP a los 30 min. Durante los 26 min finales de ejercicio, el incremento en el IMP persistió solo en la semana 4. En el caso de la semana 0, el incremento observado en el min 56 fue mayor que en los min 0 y 30. En el min 56, las concentraciones de IMP fueron menores en las semanas 4, 7 y 12 en comparación con la semana 0.
Tanto el ejercicio como el entrenamiento alteraron los niveles de fosfocreatina (PCr) y creatina (Cr) (Tabla 4). En el caso de la PCr, fueron observadas disminuciones progresivas en cada etapa del ejercicio, sin tener en cuenta el estado de entrenamiento. Sin embargo, solo al min 56 fueron observadas diferencias con el HRT. En el min 56, fueron encontrados valores de PCr más altos en las semanas de entrenamiento 4, 7 y 12 en comparación con la semana 0. Como se esperaba, para la Cr fueron encontrados efectos significativos generalmente similares, pero opuestos.
Solo el ejercicio alteró la concentración de los intermediarios glucolíticos glucosa-1-fosfato, glucosa-6-fosfato, fructosa-6-fosfato, y fructosa-1,6-bifosfato (Tabla 5). En todos los casos, los valores observados en los min 30 y 56 fueron mayores que los medidos antes del ejercicio. Solo en el caso de la glucosa-1-fosfato hubo un incremento, observado en el min 56. La glucosa se incremento en los min 30 y 56 de ejercicio, sin embargo, fue también notado un efecto de entrenamiento. En general, los valores de glucosa fueron más altos en las semanas 4 y 7 que en las semanas 0 y 12.
Las concentraciones musculares de lactato se incrementaron con cada segmento de ejercicio antes del entrenamiento y después de cada período de entrenamiento (Figura 1). Sin embargo, la magnitud del incremento fue afectada por el HRT. En el min 56 de ejercicio, el lactato estuvo más bajo en las semanas 7 y 12 en comparación con la semana 0. Tales efectos de entrenamiento no fueron observados en el piruvato. En el caso de este metabolito, el ejercicio incremento los niveles en el min 56, pero no en el min 30.
La velocidad de depleción de glucógeno fue alterada con el entrenamiento, pero no durante el período del ejercicio de los min 30 al 56, un efecto que fue evidente en la semana 4 y persistió en la semana 7 y 12 (Figura 1). Para todos los estados de entrenamiento, la reducción en el glucógeno fue progresiva a través del tiempo. El entrenamiento no tuvo efectos en cuanto a alterar la concentración de glucógeno de reposo.
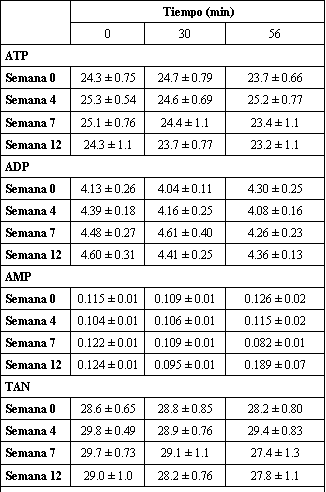

Tabla 3. Entrenamiento de fuerza de alta intensidad, nucleótidos de adenina
musculares e IMP durante el ejercicio submáximo. Los valores están expresados
como medias±DS en mmol/kg de peso seco; n=7 sujetos. TAN, nucleótidos de adenina
totales. * Significativamente diferente (p<0.05) con respecto al minuto 0, †
Significativamente diferente (p<0.05) con respecto al minuto 30, ‡
Significativamente diferente con respecto a las semanas 4, 7, 12.
El área promedio de los tipos de fibras se incrementó con el HRT, pero no hasta la semana 7 (Figura 2). En la semana 7 y 12 del HRT, las áreas promedio de las fibras fueron mayores que en la semana 0 y 4.
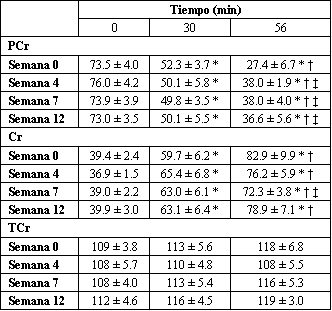
Tabla 4. Entrenamiento de fuerza de alta intensidad y fosfatos de alta energía y
metabolitos durante el ejercicio submáximo. Los valores están expresados como
medias±DS en mmol/kg de peso seco; n=7 sujetos. PCr, fosfocreatina, Cr,
creatina, TCr, creatina total. * Significativamente diferente (p<0.05) con
respecto al minuto 0, † Significativamente diferente (p<0.05) con respecto al
minuto 30, ‡ Significativamente diferente con respecto a la semana 0.
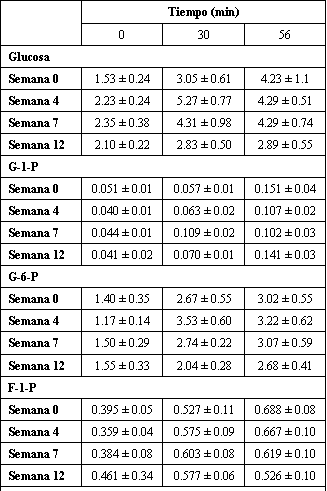
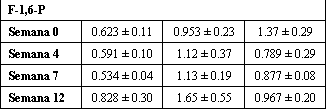
Tabla 5. Entrenamiento de fuerza de alta intensidad y cambios en intermediaries
glucolíticos selectivos con el ejercicio submáximo. Los valores están expresados
como medias±DS en mmol/kg de peso seco; n=7 sujetos. G-1-P, glucosa-1-fosfato;
G-6-P, glucosa-6-fosfato; F-6-P, fructosa-6-fosfato; F-1,6-P, fructosa-1,6-bifosfato.
Fue encontrado un efecto principal (p<0.05) para el tiempo para la glucosa,
G-1-P, G-6-P, y F-6-P. Para la glucosa, G-6-P, F-6-P, 30=56 >0 min; para la
G-1-P, 0 <30<56 min. Fue encontrado un efecto principal (p<0.05) para el
entrenamiento para la glucosa, semanas 4=7 > 0=12.
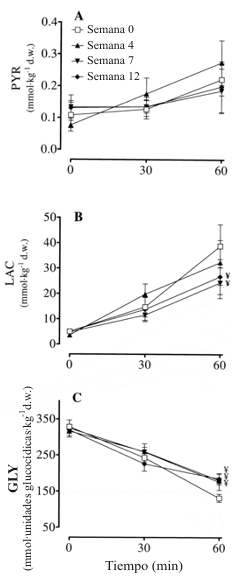
Figura 1. Entrenamiento de fuerza de alta intensidad y piruvato muscular (PYR;
A), lactato (LAC; B), y glucógeno (GLY; C). Los valores están expresados como
medias±DS; n=7 sujetos. Para el piruvato solo fue encontrado un efecto principal
(p<0.05) del tiempo: 56 >30= 0 min. dw, peso seco. ¥ Significativamente
diferente (p<0.05) con respecto a la semana 0. Para ambos, lactato y glucógeno,
el cambio fue progresivo (p<0.05) desde el min 0 al min 30 y desde el min 30 al
56. La única excepción fue el lactato en la semana 7, cuando el cambio desde el
min 0 al min 30 no fue significativo.
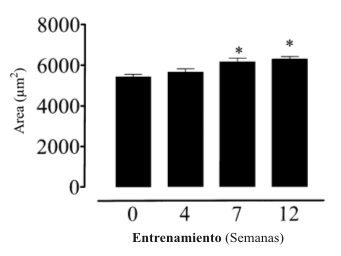
Figura 2. Entrenamiento de fuerza de alta intensidad y área promedio de las
fibras. Los valores están expresados como media±DS, n=6 sujetos. *
Significativamente diferente con respecto a la semana 0 (p<0.05).
DISCUSION
Los resultados de este artículo indican que el HRT resulta en una alteración en la respuesta metabólica muscular al ejercicio de ciclismo submáximo. El contenido de fosfatos de alta energía musculares fue mejor protegido, como fue evidenciado por la menor acumulación de IMP y una disminución menos pronunciada en la PCr. Así como, fueron observadas una atenuación del incremento de lactato, y una reducción de la depleción de glucógeno. Los trabajos previos han demostrado que los cambios en el IMP muscular son un indicador más exacto de los cambios en el ATP, ya que, dada su relativamente baja concentración, pequeños cambios son fácilmente detectables y los cambios son estequimétricos con los cambios en el ATP (19). Interesantemente, las adaptaciones metabólicas (fosfatos de alta energía) que fueron observadas fueron evidentes solo a la mayor intensidad de entrenamiento y dentro de las primeras 4 semanas de entrenamiento. El entrenamiento adicional por las siguientes 8 semanas fallo en exagerar los efectos metabólicos que fueron observados. Los efectos del HRT sobre el metabolismo muscular en ejercicio, al menos cualitativamente, son consistentes con lo que han sido reportados para el entrenamiento prolongado (13, 20). La medición del área promedio de las fibras, una medición basada en las áreas de los respectivos tipos de fibras en conjunción con sus porcentajes de distribución, se incremento con el HRT, pero no hasta la semana 7 de entrenamiento. Así, nuestra hipótesis puede ser rechazada. Las adaptaciones metabólicas que fueron observadas precedieron cualquier cambio en la hipertrofia muscular o en la hipertrofia específica de un tipo de fibras, capilarización, u potencial oxidativo, sin tener en cuenta el tipo de fibras (11). La disociación entre las adaptaciones hipertróficas y metabólicas es enfatizada más tarde en el entrenamiento. En las semanas 7 y 12, donde ocurrió la hipertrofia, no fueron detectados cambios en el metabolismo.
Debido a que la hipertrofia de las fibras musculares no fue observada al tiempo en el que fueron encontradas las adaptaciones metabólicas con el HRT, otro (s) mecanismo (s) parecen estar implicados. Varios parecen ser posibles. Imaginablemente las adaptaciones metabólicas podrían ocurrir debido a cambios composicionales dentro de la fibra, pero simplemente por medio de un cambio en la estrategia de reclutamiento (6, 28, 32).
Los cambios en el metabolismo muscular que nosotros hemos observado son típicos de aquellos que ocurren con el entrenamiento submáximo prolongado, a saber un potencial de fosforilación más protegido (i.e., una menor disminución en el ATP y la PCr y un menor incremento del ADP y el Pi), una disminución en la acumulación de lactato, y una disminución en la depleción de glucógeno (13, 20). De manera característica, estos cambios han sido atribuidos a los incrementos del potencial oxidativo de la célula (20). De acuerdo con esta hipótesis, a un nivel dado de fosforilación oxidativa (VO2), son necesarios menores incrementos en los moduladores putativos de la respiración mitocondrial como el ADP y el Pi, dado el incremento adaptativo en el potencial mitocondrial (20). También ha sido propuesto que la menor acumulación de estos metabolitos reduciría la activación de la fosforilasa y la fosfofructo quinasa, resultando en una depresión de la glucogenólisis y la glucólisis después del entrenamiento (20).
Debido a que el programa de entrenamiento HRT fallo en alterar la actividad de la SDH sin tener en cuenta el tipo de fibra (11), el incremento en el potencial mitocondrial no puede ser usado para explicar las adaptaciones observadas en este estudio. Interesantemente, nosotros también hemos reportado cambios metabólicos similares dentro de los primeros pocos días de entrenamiento de resistencia prolongado (3, 9) y ocurrieron incrementos anteriores en el potencial oxidativo (3, 9, 10, 30). Este hallazgo ha sido recientemente discutido (33). Los resultados de este estudio parecerían confirmar que al menos las adaptaciones metabólicas iniciales ocurren por algunos otros mecanismos.
Un efecto llamativo del HRT es una transformación temprana de las fibras tipo IIB a fibras tipo IIA y IIAB (11), un efecto que ha sido previamente observado (34). Esta adaptación representa un cambio en el contenido de la isoforma de miosina de cadena pesada (HC) desde HCIIb a HCIIa (34). Debido a que las fibras tipo IIB tienen típicamente un potencial oxidativo bajo, es posible que el cambio hacia las fibras tipo IIA, las cuales tienen un alto potencial oxidativo, podría haber influenciado la respuesta metabólica. Sin embargo, este efecto es cuestionable por varias razones. Primero, el porcentaje de fibras tipo IIB es relativamente bajo, ≈15% en el músculo desentrenado (observación no publicada). Segundo, es desconocido si el potencial oxidativo de las fibras cambia en coordinación con la transformación del contenido de la isoforma de HC. Finalmente, no esta claro el perfil metabólico de los subtipos de fibras tipo II durante el ejercicio prolongado voluntario.
Los cambios en los eventos fuera del los músculos que trabajan, que resultan en un incremento de la disponibilidad de oxígeno, pueden también estar mecánicamente unidos a los cambios metabólicos que fueron observados con el HRT. De acuerdo a este razonamiento, los incrementos en la disponibilidad de oxígeno en la mitocondria podrían resultar en un incremento en la fosforilación oxidativa, reduciendo la dependencia sobre los fosfatos de alta energía y la glucólisis (4). Estos ajustes pueden ser más importantes durante la fase en estado no estable del ejercicio, donde la disponibilidad de oxígeno puede ser el limitante. Tal como con el modelo de entrenamiento prolongado (13, 20), las adaptaciones metabólicas con el HRT ocurren en la ausencia de cambios en el VO2 en estado estable y supuestamente en la fosforilación oxidativa al nivel de los músculos que trabajan. Además, como podría ser esperado con el HRT (25, 27), nosotros no encontramos ningún cambio en el VO2 pico en ninguna etapa del entrenamiento. Como consecuencia, el porcentaje relativo de consumo máximo de oxígeno usado para el ejercicio submáximo permaneció constante.
Con el modelo de entrenamiento de corta duración, nosotros hemos proporcionado evidencia que sugiere que los ajustes metabólicos que ocurren durante el estado no estable (10) parecen ser dependientes de los incrementos en la cinética del VO2 (29) y del aumento del flujo sanguíneo (29). Efectos similares podrían estar ocurriendo con el HRT. Los incrementos en el flujo sanguíneo, sin embargo, no parecerían ser mediados por los incrementos en la capilarización. Los incrementos en el número de capilares en contacto con cada fibra fueron observados con el HRT, pero solo en la semana 12 (11). Quizás, más importante, el incremento en la angiogénesis capilar fue solo suficiente al inicio de la hipertrofia de las fibras, y no fue observado ningún cambio en los capilares por unidad de área de cada fibra.
Nuestros resultados inesperados, ambos con respecto a los cambios en el funcionamiento metabólico en el HRT, antes de la hipertrofia muscular y en la ausencia de un efecto de la hipertrofia muscular más tarde en el HRT, pueden ser explicados por los cambios en el reclutamiento neural. Con respecto a la falla de la hipertrofia a los cambios de efecto, un factor central parecería girar en torno a la transferencia de las mejoras en la fuerza muscular, observadas durante el entrenamiento por si mismo, a las mejoras en la potencia mecánica en la bicicleta, la cual fue usada para investigar las adaptaciones metabólicas. Aunque las mejoras no fueron medidas en 1 RM con los pesos libres, fue medido el peso que pudo ser levantado durante las 6-8 RM. Como se esperaba, ocurrieron mejoras dramáticas para los tres ejercicios durante el curso del HRT. Nosotros también encontramos que, en la semana 12, estos incrementos se tradujeron a un incremento en la MVC del cuadriceps, medida isométricamente bajo condiciones estándar. El incremento en la MVC también se tradujo en un incremento en la producción de potencia mecánica, pero solo a los mayores ajustes de resistencia. La producción de potencia no mejoro por medio del HRT a resistencias bajas y altas velocidades, una observación que enfatiza las especificidad de las adaptaciones al HRT previamente reportadas (5, 7, 23). A los mayores ajustes de resistencia, el pico de potencia se produjo a una velocidad que correspondía a la velocidad seleccionada para la evaluación submáxima (360º/s). Sin embargo, la potencia generada durante la evaluación submáxima de dos etapas solo representó el 21 y 37% del nivel pre-entrenamiento. Después del entrenamiento, el porcentaje disminuyó a 17 y 24% respectivamente. Sin embargo, a pesar de esta disminución, el metabolismo en ejercicio no fue afectado.
La falla para hallar un efecto del HRT y un incremento en la producción de potencia mecánica sobre el metabolismo muscular puede resultar a partir de un cambio en la estrategia de reclutamiento para generar fuerza después de la hipertrofia inducida por el HRT. Nosotros hemos propuesto que en la ausencia de cambios en el reclutamiento, sería generada menos fuerza por unidad de área de sección cruzada, y consecuentemente habría menos agotamiento metabólico, aunque las tasas de producción de ATP permanezcan sin cambios. Sin embargo, este no sería el caso. Hay evidencia tanto de electromiografías (18, 28) como de imageria por resonancia magnética (31) que sugiere que ocurre una reducción en la conducción nerviosa y que menos músculo es usado para generar la misma potencia luego del HRT. Esto podría concentrar el esfuerzo metabólico en menos fibras, obteniendo el efecto de la hipertrofia.
Además, no esta claro si la producción de potencia mecánica elevada observada después del HRT fue provocada por el cuadriceps. En el ciclismo, son activados 7 músculos diferentes, con su contribución relativa dependiendo de la carga. A producciones de potencia bajas, los músculos del cuadriceps (vasto lateral, vasto medial, vasto intermedio) son particularmente importantes, pero a medida que la producción de potencia disminuye el glúteo mayor se convierte en el mayor contribuyente (14). Es posible que, aún con la hipertrofia, la contribución el vasto lateral permanezca sin cambios.
De significancia potencial es el hecho que los efectos metabólicos del HRT fueron observados solo a las producciones de potencia más altas, acercándose a las características de alta carga del programa de entrenamiento. Como ha sido repetidamente demostrado con el HRT (28, 32) los cambios en la conducción neural pueden haber ocurrido durante la fase temprana del entrenamiento. Si este es el caso, el efecto de la adaptación neural pudo ocurrir para distribuir la producción de fuerza sobre más unidades motoras y/o para facilitar el flujo sanguíneo y la extracción de oxígeno por los músculos que trabajan. Estas hipótesis proveen dirección para futuros estudios.
Finalmente, debe ser reconocido que podría haber ocurrido un error estadístico, resultado de nuestra inhabilidad para detectar cambios reales en la hipertrofia celular antes de 7 semanas de HRT. Aunque esta posibilidad existe en todos los estudios, parecería que tiene pocos efectos sobre las conclusiones alcanzadas en este estudio. En la semana 4 del HRT, cuando la mayoría de las adaptaciones metabólicas fueron expresadas, la hipertrofia muscular fue mínimamente alterada.
En conclusión, hemos sido capaces de demostrar que el HRT promueve en efecto una alteración en el funcionamiento metabólico muscular observado durante el ejercicio submáximo. Sin embargo, el cambio en el metabolismo parece ser independiente de la hipertrofia muscular.
Agradecimientos: Este estudio fue apoyado por el Consejo Nacional de Investigación en Ciencias e Ingeniería de Canadá.
Dirección para pedido de reimpresiones: H. J. Green, Dept. of Kinesiology, Univ. of Waterloo, Waterloo, Ontario, Canada N2L 3G1.
Referencias
1. Anderson, P., and J. Henriksson (1977). Capillary supply of the quadriceps femoris muscle of mann. Adaptive response to exercise. J. Physiol. (Lond.) 270: 677-690
2. Boska, M (1984). ATP production rates as a function of force level in the human gastrocnemius/soleus using 31P MRS. Magn. Reson. Med. 32: 1-10
3. Connett, R. J., C. R. Honig, T. E. J. Gayeski, and G. A. Brooks (1990). Defining hypoxia: a systems view of VO2, glycolysis, energetics and intracellular PO2. J. Appl. Physiol. 68: 833-842
4. Coyle, E. F., D. C. Feiring, T. C. Rotkis, R. W. Cote III, F. B. Ruby, W. Lee, and J. H. Wilmore (1981). Specificity of power improvements through slow and fast isokinetic training. J. Appl. Physiol. 51: 1437-1442
5. Enoka, R. M (1988). Muscle strength and its development. New perspectives. Sports Med. 6: 146-168
6. Ewing, J. L., Jr., D. R. Wolfe, M. A. Rogers, M. L. Amundson, and G. A. Stull (1990). Effects of velocity of isokinetic training on strength, power and quadriceps muscle fibre characteristics. Eur. J. Appl. Physiol. 61: 159-162
7. Fowler, M. D., T. W. Ryschon, R. E. Wysong, C. A. Combs, and R. S. Balaban (1997). Normalized metabolic stress for 31P-NMR spectroscopy studies of human skeletal muscle: MVC vs. muscle volume. J. Appl. Physiol. 83: 875-883
8. Green, H., R. Helyar, M. Ball-Burnett, N. Kowalchuk, S. Symon, and B. Farrance (1992). Metabolic adaptations to training precede changes in muscle mitochondrial capacity. J. Appl. Physiol. 72: 484-491
9. Green, H., C. Goreham, J. Ouyang, M. Ball-Burnett, and D. Ranny (1999). Regulation of fiber size, oxidative potential, and capillarization in human muscle by resistance exercise. Am. J. Physiol. 276 (Regulatory Integrative Comp. Physiol. 45): R591-R596
10. Green, H. J., and S. R. Jones (1989). Does post-tetanic potentiation compensate for low frequency fatigue?. Clin. Physiol. 9: 499-514
11. Green, H. J., S. Jones, M. Ball-Burnett, B. Farrance, and D. Ranney (1995). Adaptations in muscle metabolism to prolonged exercise and training. J. Appl. Physiol. 78: 138-145
12. Green, H. J., and A. E. Patla (1992). Maximal aerobic power: neuromuscular and metabolic considerations. Med. Sci. Sports Exerc. 24: 38-46
13. Green, H. J., J. Sutton, P. Young, A. Cymerman, and C. S. Houston (1989). Operation Everest II: muscle energetics during maximal exhaustive exercise. J. Appl. Physiol. 66: 142-150
14. Green, H. J., J. A. Thomson, and M. E. Houston (1987). Supramaximal exercise after training-induced hypervolemia. II. Blood/muscle substrates and metabolites. J. Appl. Physiol. 62: 1954-1961
15. Hather, B. M., P. A. Tesch, P. Buchanan, and G. A. Dudley (1991). Influence of eccentric actions on skeletal muscle adaptations to resistance training. Acta Physiol. Scand. 143: 177-185
16. Hochachka, P. W., and G. O. Matheson (1992). Regulating ATP turnover over broad dynamic work ranges in skeletal muscles. J. Appl. Physiol. 73: 1697-1703
17. Holloszy, J. O., and E. F. Coyle (1984). Adaptations of skeletal muscle to endurance exercise and their metabolic consequences. J. Appl. Physiol. 56: 831-838
18. Hughson, R. L., J. M. Kowalchuk, W. M. Prime, and H. J. Green (1980). Open-circuit gas exchange analysis in the non-steady state. Can. J. Appl. Physiol. 5: 15-18
19. Ingebretson, D. C., A. M. Bakken, L. Segadal, and M. Farstad (1982). Determination of adenine nucleotides and inosine in human myocardium by ion pair reversed phase high performance liquid chromatography. J. Chromatogr. Sci. 242: 119-126
20. Jones, D. A., and O. M. Rutherford (1987). Human muscle strength training. The effects of three different regimes and the nature of the resultant changes. J. Physiol. (Lond.) 391: 1-11
21. Kraemer, W. J., S. J. Fleck, and W. J. Evans (1996). Strength and power training: physiological mechanisms of adaptation. In: Exercise and Sport Science Reviews, edited by J. O. Holloszy. Baltimore, MD: Williams and Wilkins, p. 363-397
22. Kraemer, W. J., J. F. Patton, S. E. Gordon, E. A. Harman, M. R. Deschenes, K. Reynolds, R. U. Newton, N. T. Triplett, and J. E. Dziados (1995). Compatability of high-intensity strength and endurance training on hormonal and skeletal muscle adaptations. J. Appl. Physiol. 78: 976-989
23. Lowry, O. H., and J. V. Passonneau (1972). A Flexible System of Enzymatic Analysis. New York: Academic, p. 146-218
24. Marcinik, E. J., J. Potts, G. Schlabach, S. Will, P. Dawson, and B. F. Hurley (1991). Effects of strength training on lactate threshold and endurance performance. Med. Sci. Sports Exerc. 23: 739-743
25. Moritani, T., and H. A. deVries (1979). Neural factors versus hypertrophy in the time course of muscle strength gain. Am. J. Phys. Med. Rehabil. 58: 115-130
26. Phillips, S. M., H. J. Green, M. J. MacDonald, and R. L. Hughson (1995). Progressive effect of endurance training on VO2 kinetics at the onset of submaximal exercise. J. Appl. Physiol. 79: 1914-1920
27. Phillips, S. M., H. J. Green, M. A. Tarnopolsky, G. J. F. Heigenhauser, and S. M. Grant (1996). Progressive effect of endurance training on metabolic adaptations in working skeletal muscle. Am. J. Physiol. 270 (Endocrinol. Metab. 33): E265-E272
28. Ploutz, L. L., P. A. Tesch, R. L. Biro, and G. A. Dudley (1994). Effects of resistance training on muscle use during exercise. J. Appl. Physiol. 76: 1675-1681
29. Sale, D. G (1992). Neural adaptation to strength training. In: Strength and Power in Sports. The Encyclopaedia of Sports Medicine Oxford, edited by P. Komi. Oxford, UK: Blackwell, p. 249-265
30. Spina, R. J., M. M.-Y. Chi, M. G. Hopkins, P. M. Nemeth, O. H. Lowry, and J. O. Holloszy (1996). Mitochondrial enzyme increase in muscle in response to 7-10 days of cycle exercise. . J. Appl. Physiol. 80: 2250-2254
31. Staron, R. S., E. S. Malicky, M. J. Leonardi, J. E. Falkel, F. C. Hagerman, and G. A. Dudley (1990). Muscle hypertrophy and fast fiber type conversions in heavy resistance-trained women. Eur. J. Appl. Physiol. 60: 71-79
32. Wang, N., R. S. Hikida, R. S. Staron, and J. A. Simoneau (1993). Muscle fiber types of women after resistance training-quantitative ultrastructure and enzyme activity. Pflügers Arch. 424: 494-502