
Factores Limitantes del Máximo Consumo de Oxígeno y Determinantes del Rendimiento de Resistencia
David R Bassett1 y Edward T Howley1
Department of Exercise Science and Sport Management, University of Tennesse, Knoxville, TN.
Artículo publicado en el journal PubliCE, Volumen 0 del año 2000.
Publicado 7 de abril de 2004
Resumen
Palabras clave: cardiorrespiratorio, aptitud física, ejercicio, transporte de oxígeno, maratón, economía de carrera
INTRODUCCION
El máximo consumo de oxígeno (VO2 máx.) esta definido como la tasa más alta a la cual el oxígeno puede ser captado y utilizado por el cuerpo durante el ejercicio intenso. Es una de las principales variables en el campo de la fisiología del ejercicio, y es frecuentemente usado para indicar la aptitud cardiorrespiratoria de un individuo. En la literatura científica, un incremento del VO2 máx. es el método más común de demostrar un efecto de entrenamiento. Además, el VO2 máx. es frecuentemente usado en el desarrollo de una prescripción de ejercicio. Dadas estas aplicaciones del VO2 máx., ha habido un gran interés en identificar los factores fisiológicos que limitan el VO2 máx. y determinar el rol de esta variable en el rendimiento de resistencia.
El concepto actual de VO2 máx. empezó con el trabajo de Hill et al. (41, 42) en 1923-24. Su visión acerca del máximo consumo de oxígeno ha sido validada, aceptada y extendida por muchos fisiólogos del ejercicio mundialmente famosos (2, 17, 58, 68, 74, 83). Sin embargo, ha sido recientemente discutido que el paradigma del VO2 máx. de Hill es un concepto anticuado, en base a defectos críticos de lógica (62). Para considerar este punto de vista, nosotros discutimos los argumentos de ambos lados y concluimos en 1997 (5) que la visión “clásica” del VO2 máx. era correcta. El presenta artículo es un intento de clarificar nuestros enfoques sobre el VO2 máx. y para presentar evidencia en apoyo de la teoría de Hill.
La parte I de este artículo revisa la historia del concepto de VO2 máx. La parte II describe la evidencia de cada uno de los potenciales factores limitantes del VO2 máx. La parte III discute el rol del VO2 máx. y otros factores en la determinación del rendimiento de resistencia.
PARTE I: HISTORIA DEL MÁXIMO CONSUMO DE OXIGENO
El término “máximo consumo de oxígeno” fue creado y definido por Hill et al. (41, 42) y Herbst (39) en la década del 20 (74). El paradigma del VO2 máx. de Hill y Lupton (42) postula que:
- hay un límite superior para el consumo de oxígeno.
- hay diferencias interindividuales en el VO2 máx.
- un alto VO2 máx. es un prerrequisito para el éxito en la carrera de media y larga distancia.
- el VO2 máx. está limitado por la capacidad del sistema cardiorrespiratorio para transportar el O2 a los músculos.
En 1923, Hill y Lupton (42) hicieron cuidadosas mediciones del consumo de oxígeno en un sujeto (A.V.H.), que corrió alrededor de una pista de pasto de 85 m. El gráfico presentado en la Figura 1 fue desarrollado principalmente para el propósito de ilustrar el cambio del VO2 en función del tiempo a tres velocidades (181, 203, y 267 m.min-1). En un estudio publicado el año siguiente, Hill et al. (41) reportaron más mediciones del VO2 en el mismo sujeto. Después de 2.5 min de carrera a 282 m.min-1 su VO2 alcanzó un valor de 4080 L.min-1 (o 3730 L.min-1 arriba del VO2 medido en reposo, con el sujeto de pie). Ya que el VO2 a las velocidades de 259, 267, 271, y 283 m.min-1 no se incrementó más allá de aquel medido a 243 m.min-1, esto confirma que a altas velocidades el VO2 alcanza un máximo, más allá del cual ningún esfuerzo puede aumentarlo.
Actualmente, está universalmente aceptado que hay un límite fisiológico superior de la capacidad del cuerpo de consumir oxígeno. Esto es mejor ilustrado en el clásico gráfico de Astrand y Saltin (4) presentado en la Figura 2. En un protocolo de evaluación discontinua, los intentos repetidos de conducir el consumo de oxígeno a valores más altos incrementando la tasa de trabajo son inefectivos. La tasa de crecimiento en el VO2 se incrementa con cada intento sucesivo, pero la “límite superior” alcanzado en cada caso es aproximadamente el mismo. Los sujetos simplemente alcanzaban el VO2 máx. antes con las producciones de potencias más altas. El VO2 máx. no continua incrementándose indefinidamente con el incremento de la tasa de trabajo (o velocidad de carrera). Este hallazgo fue predicho por Hill y Lupton ((42), p. 156), los cuales establecieron eventualmente, “…sin embargo por mucho que la velocidad (o tasa de trabajo) se incrementada más allá de este límite, no puede ocurrir un incremento subsiguiente en la captación de oxígeno”.
No todos los sujetos muestran un plateau en el VO2 al final de una evaluación de ejercicio gradual (GXT), cuando se la grafica vs. la intensidad de trabajo. Ha sido repetidamente demostrado que cerca del 50% de los sujetos no demuestran un plateau cuando son estresados hasta un esfuerzo máximo (46). La falla para alcanzar un plateau no significa que estos sujetos han fallado en alcanzar su “verdadero” VO2 máx. (26). En primer lugar, con un protocolo continuo de GXT un sujeto se puede fatigarse justo cuando es alcanzado el VO2 máx.. Así, el plateau puede no ser evidente aunque el VO2 máx. haya sido alcanzado (69). Segundo, aún con un protocolo discontinuo de GXT la mayoría de los investigadores requieren que los sujetos completen 3-5 min en cada etapa (3, 26, 83). Así, si un sujeto alcanza el VO2 máx. en 2 min a una intensidad supramáxima y luego se fatiga demasiado para continuar, estos puntos de datos no lo graficarían. En este caso, el plateau del VO2 no va a ser aparente en el gráfico final de la tasa de trabajo versus el consumo de oxígeno, aunque el VO2 máx. haya sido alcanzado (Figura 3). Por estas razones, el plateau del VO2 no puede ser usado como el único criterio de logro del VO2 máx. Esta es la razón de porque es recomendado que criterios secundarios sean aplicados para verificar un esfuerzo máximo. Esto incluye un índice de intercambio respiratorio> 1.15 (47) y un nivel de ácido láctico sanguíneo> 8-9 mM (2), un enfoque que ha sido confirmado en nuestro laboratorio (26).
El plateau del VO2 representa una nivelación en el gasto cardíaco y la diferencia a-v de oxígeno que puede ser vista hacia el final de una GXT. Ya que el VO2 falla en mantenerse al ritmo con la demanda incremental de oxígeno, hay un incremento de la dependencia sobre las vías independientes de oxígeno (i.e., glucólisis anaeróbica). La significancia del plateau de VO2 ha sido frecuentemente mal interpretada. En 1988, fue sugerido que la ausencia de un plateau de VO2 en algunas personas significaba que el VO2 máx. no estaba limitado por el sistema cardiovascular (61). Esto condujo a la sugerencia acerca de que “factores musculares” deben ser importantes para limitar el VO2 máx. Sin embargo, como nosotros destacamos, el plateau del VO2 no es la principal evidencia de una limitación cardiovascular (5).
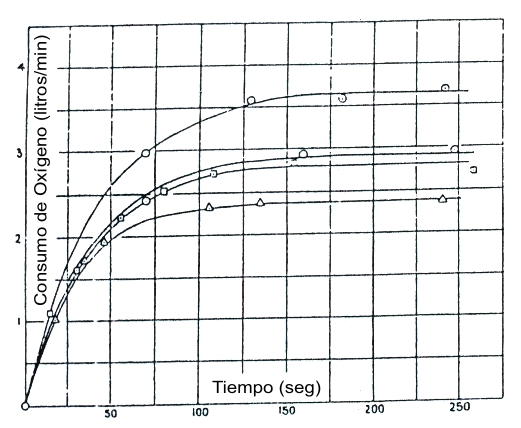
Figura 1. El logro de un “estado estable” para la captación de oxígeno, a
varias velocidades constantes. El eje horizontal describe el tiempo desde el
inicio de cada carrera; el eje vertical muestra la captación de oxígeno (L.min-1).
Las velocidades son 181, 203, 203, y 267 m.min-1 (desde abajo hasta
arriba). Las tres curvas de más abajo representan un estado estable genuino,
mientras que en la curva de arriba los requerimientos de oxígeno exceden el
consumo de oxígeno medido. Tomado de la referencia 42, Hill, A.V. y H. Lupton.
Muscular exercise, lactic acid, y supply and utilization of oxygen. Q. J. Med.
16: 135-171, 1923. Usado con el permiso de Oxford University Press.
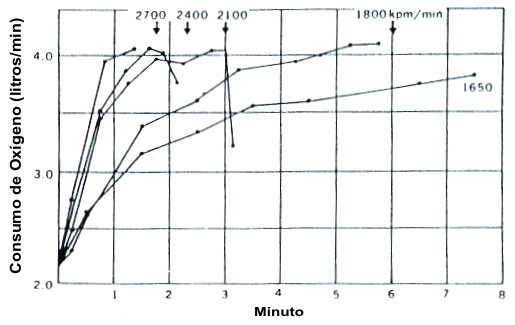
Figura 2. Transcurso de tiempo del incremento en la captación de oxígeno
durante el ejercicio intenso en un cicloergómetro. Las flechas muestran el
tiempo cuando el sujeto paraba el ejercicio debido a la fatiga. La producción de
potencia (W) para cada serie también es presentada. El sujeto pudo continuar
ejercitándose a una producción de potencia de 275 W por más de 8 min. Tomado de
la referencia 3. Astrand, P.-O. y K. Rodahl. Textbook of Work Physiology. New
Cork: McGraw-Hill, 1970. Usado con permiso.
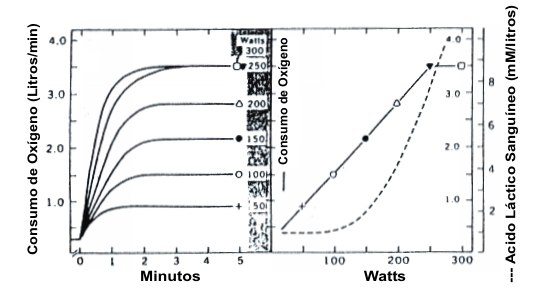
Figura 3. Consumo de oxígeno y respuesta del lactato sanguíneo a una
evaluación de ejercicio discontinuo. El lado derecho muestra el incremento de la
captación de oxígeno (L.min-1) graficado versus la producción de
potencia (W). Notar que la producción de potencia de 250 W provocó este VO2
máx. del sujeto y que los 300 W no incrementaron el consumo de oxígeno. Este
incremento en la producción de potencia fue alcanzado a través de procesos
anaeróbicos. Tomado de la referencia 3, Astrand, P.-O. y K. Rodahl. Textbook of
Work Physiology. New Cork: McGraw-Hill, 1970. Usado con permiso.
Más recientemente, ha sido sugerido que el plateau de VO2 significa una nivelación del gasto cardíaco, causada por la isquemia progresiva e irreversible (63). Sin embargo, no hay evidencia para apoyar este enfoque. De hecho, el plateau del VO2 ocurre en cerca de la mitad de los adultos sanos que realizan ejercicio máximo (46), sin signos o síntomas de acompañamiento de isquemia miocárdica. Una explicación más reciente es que el gasto cardíaco máximo está limitado por la máxima tasa de despolarización del nodo sino-atrial (SA) y los límites estructurales del ventrículo.
Con respecto a la variabilidad en el VO2 máx., Hill y Lupton ((42), p. 158) establecieron, “Un hombre puede no ser un buen corredor por la razón de un bajo consumo de oxígeno, un débito máximo de oxígeno bajo, o un alto requerimiento de oxígeno”. Esto demuestra claramente que ellos reconocieron la presencia de diferencias interindividuales en el VO2 máx. Ellos no creían en el VO2 máx. universal de 4.0 L.min-1, como ha sido sugerido (63). Además, ellos reconocieron la importancia del un alto VO2 máx. para los deportistas de alto rendimiento (42). Ellos también establecieron que otros factores fisiológicos, tales como la economía de carrera, influenciarían el resultado de la carrera (42). Subsecuentemente los investigadores han verificado este punto (ver discusión en la Parte III).
El cuarto punto en el paradigma del VO2 máx. ha sido más controversial. Hill et al. (41) identificaron varios determinantes del VO2 máx. En base a los limitados datos disponibles para ellos, especularon que en el hombre que realiza ejercicio, el VO2 máx. es limitado por la tasa a la cual el O2 puede ser suministrado por el sistema cardiovascular (corazón, pulmones, y sangre). Durante los siguientes 75 años, muchos fisiólogos del ejercicio distinguidos estudiaron este problema usando una amplia variedad de nuevas técnicas experimentales. Ellos han arribado a un consenso que apoya el paradigma original del VO2 máx. de Hill et al. (41). El enfoque actual es que en el humano que hace ejercicio el VO2 máx. es limitado principalmente por la tasa de suministro de oxígeno, y no por la capacidad de los músculos de tomar el oxígeno de la sangre (ver Parte II).
PARTE II: FACTORES LIMITANTES DEL MÁXIMO CONSUMO DE OXIGENO
El camino del O2 desde la atmósfera hasta la mitocondria contiene una serie de pasos, cada uno de los cuales podría representar un impedimento potencial para el flujo de O2. La Figura 4 presenta los factores fisiológicos que podrían limitar el VO2 máx.: 1) la capacidad de difusión pulmonar, 2) máximo gasto cardíaco, 3) capacidad de transporte de oxígeno de la sangre, y 4) características del músculo esquelético. Los primeros tres factores pueden ser clasificados como factores “centrales”; el cuarto es llamado un factor “periférico”. La evidencia para cada uno de estos factores es discutida en las secciones siguientes.
El Sistema Pulmonar
En el individuo promedio que realiza ejercicio al nivel del mar, los pulmones realizan su trabajo saturando la sangre arterial con O2, extremadamente bien. Aún durante el trabajo máximo, la saturación arterial de O2 (%Sa O2) permanece cerca del 95% (65). Hill et al. ((41) p. 161) predijo que una caída significativa en la saturación arterial (Sa O2 <75 %) no ocurre, en base a la apariencia de sus sujetos, “los cuales no han mostrado nunca, aún en el ejercicio más intenso, algún signo de cianosis”. Sin embargo, ellos fueron cuidadosos en asumir que está presente un equilibrio alveolar-arterial completo, debido al rápido pasaje de las células sanguíneas rojas dentro de los capilares pulmonares a altas tasas de trabajo ((42) p. 155).
Los investigadores modernos han verificado que el sistema pulmonar puede en efecto limitar el VO2 máx. bajo ciertas circunstancias.
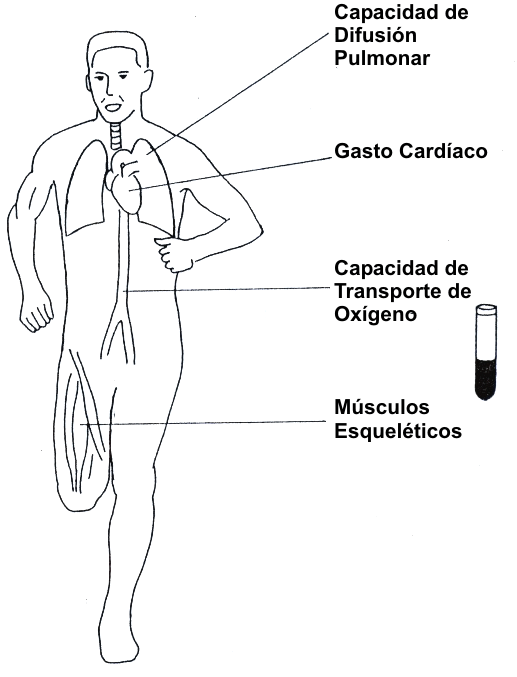
Figura 4. Factores fisiológicos que potencialmente limitan el máximo
consumo de oxígeno (VO2 máx.) en el ser humano que realiza ejercicio.
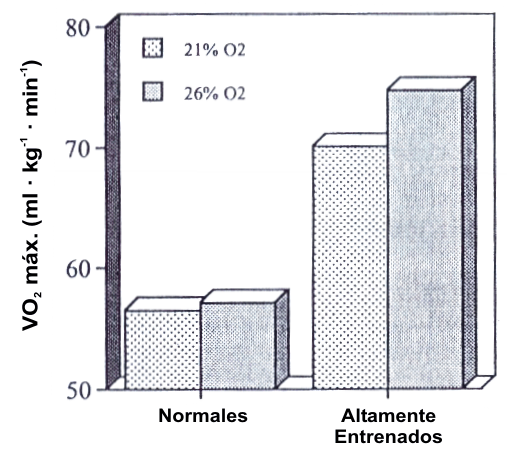
Figura 5. Efectos de la hiperoxia sobre el máximo consumo de oxígeno (VO2
máx.) en individuos normales y altamente entrenados. Los sujetos altamente
entrenados tuvieron un incremento en el VO2 máx. cuando respiraban
aire enriquecido con oxígeno, mostrando la presencia de una limitación pulmonar
en estos sujetos bajo condiciones normóxicas. Tomado de la referencia 65. Powers,
S.K., J. Lawler, J.A. Dempsey, S. Dodd; y G. Landry. Effects of incomplete
pulmonary gas exchange on VO2 max. J. Appl. Physiol. 66:
2491-2495, 1989. Usado con permiso.
Dempsey et al. (25) demostraron que los atletas de elite tienen más probabilidad de sufrir una desaturación de O2 durante el trabajo máximo en comparación con los individuos normales. Los individuos entrenados tienen un gasto cardíaco mucho más alto que los individuos desentrenados (40 vs. 25 L.min-1). Esto conduce a una disminución del tiempo de tránsito de las células rojas sanguíneas en los capilares pulmonares. Consecuentemente, puede no haber suficiente tiempo para saturar la sangre con O2 antes de que salga de los capilares pulmonares.
Esta limitación pulmonar puede ser salvada en atletas altamente entrenados con aire enriquecido con O2. Powers et al. (65) hicieron que sujetos altamente entrenados y sujetos normales realizaran dos evaluaciones de VO2 máx. (Figura 5). En una de las evaluaciones los sujetos respiraban aire de la habitación y en la otra respiraban una mezcla de gases con 26 % de O2. Con el gas hiperóxico, el grupo altamente entrenado tuvo un incremento en el VO2 máx. desde 70.1 hasta 74.7 ml.kg-1.min-1 y un incremento en la saturación arterial de O2 (Sa VO2) desde 90.6% hasta 95.9% durante el trabajo máximo. Ninguno de estos cambios fue observado en los sujetos normales (VO2 máx.= 56.5 ml.kg-1.min-1).
Las limitaciones pulmonares son evidentes en las personas que se ejercitan a alturas moderadamente altas (3000-5000 m) (22, 31, 53). Los individuos con asma y otros tipos de enfermedad pulmonar obstructiva crónica (COPD) sufren de un problema similar (una reducción en la PO2 arterial). Bajo estas condiciones, la capacidad de ejercicio puede ser incrementada con O2 adicional, el cual incrementa la “fuerza de conducción” de la difusión del O2 dentro de la sangre (23, 67). La habilidad de incrementar la capacidad de ejercicio de esta manera muestra la presencia de una limitación pulmonar.
Gasto Cardíaco Máximo
Hill et al. (41, 42) propusieron que el gasto cardíaco máximo fue el principal factor que explicaba las diferencias individuales en el VO2 máx. Esta fue una idea importante dado el estado de conocimiento en 1923. Einthoven había descubierto la electrocardiografía solo una década antes. Hill usó esta nueva técnica para medir las frecuencias cardíacas de aproximadamente 180 lat.min-1 ((41) p. 165). Sin embargo, no fue hasta cerca de 1930 que fue demostrado que los sujetos entrenados tenían menores frecuencias cardiacas a una tasa de trabajo submáxima fija (11), proporcionando evidencia de volúmenes de eyección incrementados. Otros métodos de demostrar el aumento de tamaño de los corazones de los atletas de resistencia (rayos X y ultrasonido) no estuvieron disponibles hasta 1940-1950. Dado el nivel de tecnología en 1923, es increíble que Hill et al. (41, 42) fueran capaz de deducir que los atletas de resistencia tuvieran corazones con capacidades superiores de bombeo. ¿Como llegaron ellos a esta extraordinaria conclusión?. En 1915, Lindhard (55) habían medido los gastos cardiacos de 20 L.min-1, en sujetos promedio durante el ejercicio y demostraron la fuerte relación lineal entre el gasto cardiaco y el VO2. Hill y Lupton ((42), p. 154) especularon que valores de gasto cardiaco máximo de 30-40 L. min-1 eran posibles en atletas entrenados. Estas especulaciones estaban basadas en los conocimientos de la ecuación de Fick y los valores tomados para el VO2 máx., contenido arterial de oxígeno, y contenido de oxígeno venoso mezclado.
Actualmente, nosotros sabemos que el rango normal de valores de VO2 máx. (L.min-1) observado en hombres y mujeres entrenados y desentrenados de la misma edad es debido principalmente a la variación en el volumen de eyección máximo, dado que existe considerablemente menos variación en la HR máxima y la extracción sistémica de oxígeno. Durante el ejercicio máximo, casi todo el oxígeno disponible es extraído de la sangre que perfunde a los músculos activos (76). El contenido de oxígeno de la sangre arterial es aproximadamente 200 mL de O2. L-1; en la sangre venosa que sale de los músculos en el trabajo máximo el mismo cae a 20-30 mL de O2. L-1. Esto muestra que queda poco oxígeno para ser extraído de la sangre durante el ejercicio intenso. Por lo tanto, el mecanismo dominante para el incremento en el VO2 máx. con el entrenamiento debe ser un incremento en el flujo sanguíneo (y transporte de O2). Es estimado que el 70-85% de la limitación en el VO2 máx. está relacionada con el gasto cardiaco máximo (10).
Los estudios longitudinales han mostrado que el incremento inducido por el entrenamiento en el VO2 máx. resulta principalmente de un incremento del gasto cardiaco máximo, en vez de una extensión de la diferencia sistémica a-v de O2 (Figura 6). Saltin et al. (71) examinaron el VO2 máx. en individuos sedentarios después de 20 días de reposo en cama y 50 días de entrenamiento. La diferencia en el VO2 máx. entre los estados desentrenado y entrenado resultó principalmente de la diferencia en el gasto cardiaco. En un estudio similar, Ekblom et al. (27) encontraron que 16 semanas de entrenamiento físico incrementaron el VO2 máx. desde 3.15 a 3.68 L.min-1. Este incremento en el VO2 máx. resultó a partir de un incremento de 8 % en el gasto cardiaco (desde 22.4 hasta 24.2 L.min-1) y un incremento de 3.6 % en la diferencia a-v O2 (desde 138 hasta 143 mL.L-1).
Una forma de disminuir de manera precisa el gasto cardiaco es mediante beta-bloqueantes. Tesch (84) ha escrito una extensa revisión de 24 estudios detallando las respuestas cardiovasculares a los beta-bloqueantes. Los beta-bloqueantes pueden disminuir la frecuencia cardiaca máxima (HR) en un 25-30 %. En estos estudios, el gasto cardiaco máximo disminuyó un 15-20%, mientras que el volumen sistólico se incremento ligeramente. Aunque la disminución del gasto cardíaco es parcialmente compensada por un incremento en la diferencia a-v de O2, el VO2 máx. declinó un 5-15 %. Tesch (84) concluyó que la declinación en el VO2 máx. observada con los beta-bloqueantes cardio-selectivos es causada por la disminución del flujo sanguíneo y el transporte de oxígeno.
Capacidad de Transporte de Oxígeno
Otro método para alterar el transporte de O2 a los músculos es cambiando el contenido de hemoglobina de la sangre (28). El doping sanguíneo es la práctica del incremento artificial del incremento del volumen de una persona de las células rojas sanguíneas totales a través de la remoción, almacenamiento, y subsiguiente reinfusión. Gledhill (35, 36) completó una amplia revisión de 15-20 estudios que han examinado los efectos del doping sanguíneo. La reinfusión de 900-1350 mL de sangre eleva la capacidad de transporte de la sangre. Ha sido demostrado que este procedimiento incrementa el VO2 máx. en un 4-9 % en estudios bien diseñados, doble ciego (35, 36) (Figura 7). No es observada ninguna mejora en los individuos tratados con placebo, que fueron infundidos con un pequeño volumen de solución salina (8). Una vez más, estos estudios proporcionaron evidencia de una relación causa-efecto entre el transporte de O2 y el VO2 máx.
La evidencia acerca de que el VO2 máx. es limitado por el gasto cardiaco, la capacidad de transporte del oxígeno y en algunos casos el sistema pulmonar, es irrefutable. Esta afirmación pertenece a los sujetos sanos que realizan ejercicios dinámicos para todo el cuerpo. Luego vamos a considerar si el músculo esquelético podría también ser un factor limitante del VO2 máx.
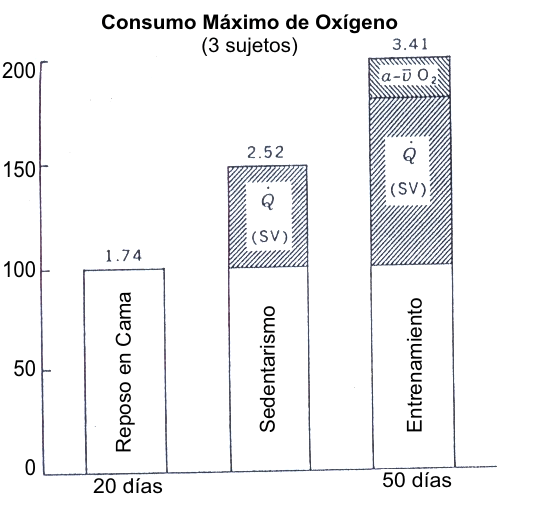
Figura 6. Resumen de los cambios que ocurren en el consumo máximo de
oxígeno (VO2 máx.) luego de el reposo en cama y del entrenamiento
físico. El mayor VO2 máx. bajo condiciones de sedentarismo en
comparación con la condición de reposo en cama resulta de un incremento en el
gasto cardiaco máximo. El incremento posterior después del entrenamiento resulta
de un incremento del gasto cardiaco y, en una menor extensión, un incremento en
la diferencia a-v de O2. Tomado de la referencia 3. Astrand, P.-O. y
K. Rodahl. Textbook of Work Physiology. New Cork: McGraw-Hill, 1970. Usado con
permiso. Los datos son de la referencia 71. Saltin, B., B., Blomquist, J.H.
Mitchell, R.L. Johnson, K. Wildenhal, y C.B. Chapman. Response to submaximal and maximal exercise after bed rest and after training.
Circulation 38 (Suppl. 7): 1-78, 1968. Usado con permiso.
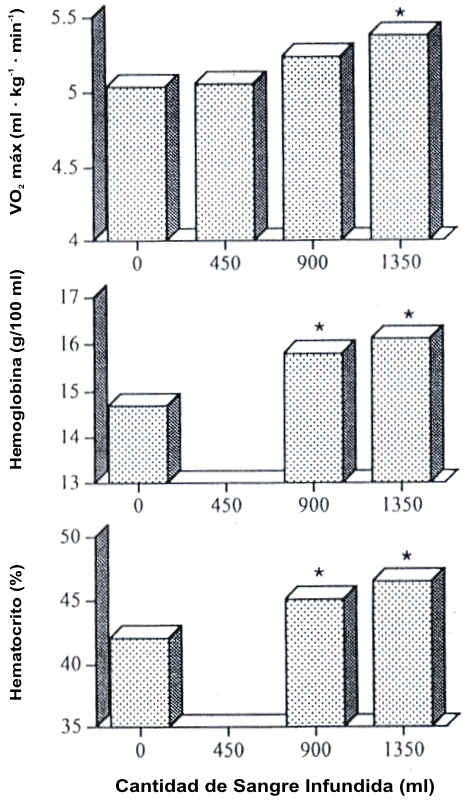
Figura 7. Cambios en la hemoglobina, hematocrito (Hct), y máximo consumo de
oxígeno (VO2 máx.) luego de las transfusiones de sangre de 1, 2, o 3
unidades de sangre total. Un incremento en la capacidad de transporte de oxígeno
de la sangre resulta en un incremento en el VO2 máx. Tomado de:
Spriet, L. L, N. Gledhill, A. B. Froese, and D.L. Wilkes. Effect of graded
erythrocythemia on cardiovascular and metabolic responses to exercise. J. Appl.
Physiol. 61: 1942-1948, 1986. Usado con permiso.
Limitaciones del Músculo Esquelético
Gradientes de Difusión Periféricos
En un simposio sobre los factores limitantes del VO2 máx., Honig et al. (45) presentaron evidencia de una limitación de difusión periférica de O2 en el músculo rojo canino. De acuerdo a sus experimentos y a un modelo matemático, el principal sitio de resistencia a la difusión de O2 ocurre entre la superficie de las células rojas sanguíneas y el sarcolema. Ellos reportaron una gran caída en la PO2 a través de esta corta distancia. Honig et al. (45) concluyeron que el transporte de O2 per se no es un factor limitante. Ellos encontraron que una baja PO2 celular relativa a la PO2 sanguínea es necesaria para mantener la fuerza de conducción para la difusión y así mejorar la conductancia del O2.
El modelo experimental de Honig et al. (45) es completamente diferente de aquel visto en un humano que hace ejercicio. Ellos notaron que simplemente incrementando el flujo sanguíneo a los músculos aislados no es suficiente para causar un incremento en el VO2. El músculo aislado debe experimentar contracciones de modo que las mitocondrias consuman O2 (disminuyendo la PO2 intracelular). Sin un gradiente de difusión periférico, el consumo de oxígeno no se va a incrementar. Su conclusión final es que el VO2 máx. es una propiedad de distribución, dependiente de la interacción del transporte de O2 y la captación de O2 mitocondrial (45). Nosotros coincidimos con esta conclusión. Sin embargo, este modelo no puede determinar cual de estos factores limitan el VO2 máx. en un humano intacto que realiza un esfuerzo máximo.
Niveles de Enzimas Mitocondriales
Los fisiólogos han hecho grandes trabajos para examinar si los niveles de enzimas mitocondriales son un factor limitante del VO2 máx. Dentro de las fibras musculares, la mitocondria es el lugar donde el O2 es consumido en el paso final de la cadena de transporte de electrones, en teoría, doblando el número de mitocondrias se debería doblar el número de sitios para el consumo de O2 en el músculo. Sin embargo, los estudios en humanos muestran que solo hay un modesto incremento del VO2 máx. (20-40 %) a pesar de un incremento de 2.2 veces de las enzimas mitocondriales (72). Esto es consistente con el enfoque acerca de que el VO2 máx., medido durante el ejercicio dinámico de todo el cuerpo, es limitado por el transporte de oxígeno (no las mitocondrias musculares).
Shephard (76) ha preguntado, “¿Si rechazamos el enfoque acerca de que hay una limitación significativa del transporte de oxígeno al nivel de los tejidos, cual explicación alternativa puede ser ofrecida a los teleólogos para explicar la duplicación de actividad enzimática tisular durante el entrenamiento de resistencia?”. En su artículo científico de revisión de referencia de 1984, Holloszy y Coyle (44) propusieron una respuesta a esta pregunta. Ellos establecieron que como consecuencia del incremento de las mitocondrias, el ejercicio a la misma tasa de trabajo, provoca menores disturbios en la homeostasis en los músculos entrenados. Dos efectos metabólicos de un incremento en las enzimas mitocondriales son: 1) los músculos que se adaptan al ejercicio de resistencia van a oxidar grasa a una tasa más alta (así ahorrando glucógeno muscular y glucosa sanguínea) y 2) hay una disminución en la producción de lactato durante el ejercicio. Estas adaptaciones musculares son importantes para explicar la mejora en el rendimiento de resistencia que ocurre con el entrenamiento. (Esto va a ser discutido posteriormente en la Parte III).
El principal efecto del incremento de las enzimas mitocondriales es la mejora del rendimiento de resistencia, en vez de un incremento del VO2 máx. Holloszy y Coyle (43, 44) destacaron que aún en individuos con valores de VO2 máx. prácticamente idénticos puede haber un rango de dos veces en las enzimas mitocondriales (1976). Además, el entrenamiento de baja intensidad puede provocar pequeños cambios en las enzimas mitocondriales sin ningún cambio en el VO2 máx., y viceversa (38, 52, 64). De otro modo, hay alguna evidencia acerca de que el incremento de las mitocondrias juega un rol permisivo para permitir que se incremente el VO2 máx. Holloszy y Coyle (44) destacaron que el valor más bajo para la actividad de la SDH en los corredores de elite estudiados por Costill (16) fue todavía 2.5 veces mayor que aquel hallado en individuos desentrenados en el mismo estudio. El incremento en las mitocondrias musculares puede permitir una extracción de O2 ligeramente mayor por parte de los músculos que trabajan, así contribuyendo en una menor medida a un incremento del VO2 máx. (44).
Densidad Capilar
En 1977 Andersen y Henriksson (1) mostraron que la densidad capilar se incrementa con el entrenamiento. Otros estudios notaron una fuerte correlación entre el número de capilares por fibra en el vasto lateral y el VO2 máx. (ml.kg-1.min-1) medido en cicloergómetro (72). La principal significancia del incremento, inducido por el entrenamiento, en la densidad capilar no es acomodar el flujo sanguíneo, sino mantener o prolongar el tiempo medio de tránsito (70). Esto mejora el transporte de oxígeno manteniendo la extracción de oxígeno (diferencia a-v de O2) aún a una alta tasa de flujo sanguíneo muscular. La capacidad del músculo esquelético para adaptarse al entrenamiento de esta forma es por lejos mayor que aquella observada en los pulmones (24).
¿Limitación Central o Periférica?
La cuestión de factores centrales o periféricos como limitantes del VO2 máx. ha sido un debate por mucho tiempo. Los trabajos conducidos a comienzos de 1970 apoyan la idea de factores centrales como limitantes del VO2 máx. Clausen et al. (12) mostraron que el entrenamiento de bicicleta con las dos piernas resultaba en un incremento en el VO2 máx. de los brazos. Ellos interpretaron correctamente esta evidencia de un efecto de entrenamiento cardiovascular central.
En 1976 Saltin et al. (73) examinaron los efectos de un entrenamiento de ciclismo con una pierna sobre el incremento del VO2 máx. en una pierna entrenada, una pierna de control, y un entrenamiento con las dos piernas. La pierna entrenada tuvo un incremento de 23 % en comparación con el incremento de 7 % en el VO2 máx. de la pierna control (Figura 8). La disparidad entre las piernas fue atribuida a adaptaciones periféricas que ocurrieron dentro del músculo esquelético entrenado. Los autores concluyeron que los factores periféricos fueron dominantes para limitar el VO2 máx. Este estudio fue conducido durante 1970 como un nuevo descubrimiento acerca del tipo de fibra, densidad capilar, y actividades enzimática oxidativas que ocurrían en atletas. En ese tiempo, los investigadores pensaban que estos cambios eran esenciales para incrementar el VO2 max. (73).
Sin embargo, en 1985 Saltin et al. (70) realizaron el experimento definitivo demostrando que el VO2 máx. es limitado por el flujo sanguíneo. Ellos observaron que pasaba cuando un sujeto hacía ejercicio máximo usando solo una pequeña masa muscular (i.e., extensiones de rodilla con solo una pierna). Esto permitió que una mayor proporción del gasto cardiaco sea dirigida a un área aislada. Bajo estas condiciones, el consumo de O2 más alto en un grupo muscular del cuadriceps fue 2-3 veces más alto que aquel medio en el mismo grupo muscular durante un esfuerzo máximo de todo el cuerpo. Ellos concluyeron que el músculo esquelético tiene una tremenda capacidad de incrementar el flujo sanguíneo y el VO2 (70), lo que excede por lejos la capacidad de bombeo del corazón durante el ejercicio máximo de todo el cuerpo. Este experimento probó que el VO2 máx. esta limitado por el transporte de oxígeno y no por la capacidad de la mitocondria de consumir oxígeno.
¿Como podemos nosotros conciliar los resultados de los dos experimentos de Saltin et al. (70, 73)?. En el primer estudio, ellos midieron el VO2 máx. durante un ejercicio de ciclismo con una sola pierna. Sin embargo, debe ser recordado que el gasto cardiaco máximo no es el factor dominante limitante del VO2 máx. en el ejercicio con un grupo muscular aislado (i.e., ciclismo con una sola pierna) (66). El VO2 máx. de todo el cuerpo es limitado principalmente por el gasto cardiaco, mientras que para el ejercicio con grupos musculares chicos el rol del gasto cardiaco es considerablemente menor (10). Ya que la conclusión de 1976 acerca de las limitaciones periféricas no se aplica al VO2 máx. medido en el ejercicio intenso para todo el cuerpo, la conclusión posterior no entra en conflicto con su trabajo anterior. La creencia actual es que el gasto cardiaco máximo es el principal factor limitante del VO2 máx. durante las evaluaciones de bicicleta o carrera (66).
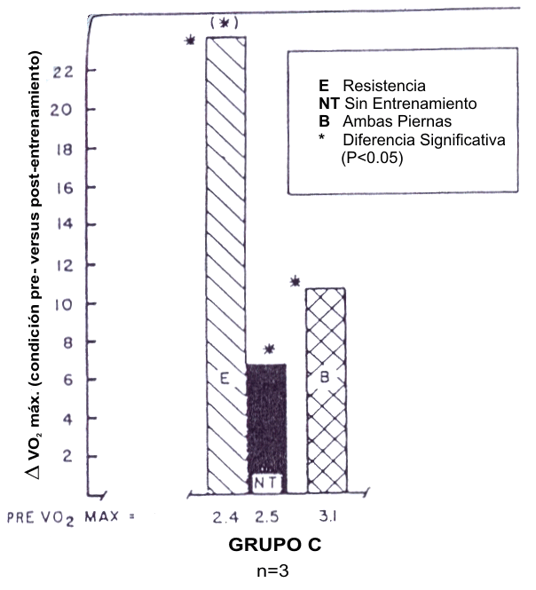
Figura 8. Efectos del entrenamiento de ciclismo con una sola pierna sobre el
VO2 máx. en la pierna entrenada, pierna no entrenada de control y
ambas piernas. El entrenamiento consistió de 4 a 5 sesiones de entrenamiento de
resistencia al 75% del VO2 máx. (1 pierna), 35-45 min por sesión,
durante 4 semanas. Este estudio proporcionó apoyo para el rol de las
adaptaciones periféricas. Tomado de la referencia 73. Saltin, B., K. K. Nazar,
D.L. Costill, et al. The nature of the training response: peripheral and central
adaptations to one-legged exercise. Acta Physiol. Scand. 96: 289-305, 1976.
Usado con permiso.
Fisiología Comparativa y Máximo Consumo de Oxígeno
Taylor et al. (79, 80, 82) y Weibel (87) estudiaron los factores fisiológicos que limitan el VO2 máx. desde una perspectiva diferente. Ellos examinaron diferentes especies animales para ver que factores fisiológicos explicaban el VO2 máx. superior de las especies más atléticas. Estos estudios en fisiología comparativa proporcionaron una vía para evaluar el concepto de “symmorphosis”, el cual hipotetiza que los animales están constituidos de una manera razonable. La suposición subyacente es que todas las partes de la ruta del O2 (desde la atmósfera hasta la mitocondria) están equilibradas con la capacidad funcional del organismo. Si cualquiera de los sistemas implicados en la ruta del O2 son sobreconstruidos, luego habría una redundancia que podría ser derrochadora, desde un punto de vista energético.
La primera serie de experimentos comparó especies de mamíferos de tamaño similar, pero con una diferencia en el VO2 máx. de 2.5 veces (perro vs. cabra, caballo de carrera vs. novillo) (48, 81). Esto es llamado “variación adaptativa” (adaptación fue definida en un sentido evolutivo, como el final-resultado de la selección natural). Los altos valores de VO2 máx. en las especies más atléticas fueron acompañados por un incremento de 2.2 veces en el volumen sistólico, frecuencias cardiacas máximas casi idénticas y un gran incremento de las mitocondrias (49, 81). En general, estas parejas adaptativas muestran diferencias fisiológicas similares a las observadas cuando son comparados humanos entrenados y desentrenados (Tabla 1).
Una segunda serie de experimentos examinó una variedad de especies animales que estaban entre unos pocos gramos hasta 250 kg (79). La diferencia observada en el VO2 máx. en los animales de diferentes masas corporales (Mb) es llamada “variación alométrica”. Los valores de VO2 máx. (L.min-1) se incrementan con la masa corporal a una potencia de 0.81. Sin embargo, cuando son ajustados para la masa corporal, los animales pequeños tienen valores de VO2 máx./Mb que son 8-10 veces mayores que los animales grandes (Figura 9). A través de un amplio rango de especies animales, hay una relación muy cercana entre la densidad mitocondrial y el VO2 máx./Mb (80). Las especies más pequeñas tienen una abundancia de mitocondrias, de modo que la capacidad de los músculos de consumir oxígeno está aumentada. Sería imposible para las especies pequeñas alcanzar tasas metabólicas tan increíblemente altas (200 a 260 ml.kg-1.min-1) sin un incremento en la densidad mitocondrial. Así, puede decirse que los músculos “establecen la demanda de O2” (80).
Los animales más atléticos también tienen un incremento en el tamaño de las estructuras implicadas en el transporte de O2 a los músculos que trabajan. El tamaño y función de los pulmones están adaptados en proporción con el VO2 máx. Además, la capacidad de bombeo del corazón está fuertemente acoplada al VO2 máx. En la variación adaptativa (animales del mismo tamaño), los animales más atléticos alcanzan la misma por un incremento en el tamaño del corazón (80). En la variación alométrica, los animales pequeños alcanzan un incremento en el transporte de VO2 con una mayor frecuencia cardiaca (1300 lat.min-1 en la musaraña) ((87), p. 400). La conclusión general de los estudios es que el principio de symmorphosis es apoyado (80). Las estructuras implicadas en la ruta del O2 están adaptadas en proporción al VO2 máx., lo que significa que los animales están constituidos de una manera razonable (88).
Sin embargo, hay excepciones donde uno observa redundancias a varios niveles en la ruta del O2. Por ejemplo, la capacidad de la mitocondria de consumir O2 excede la capacidad del sistema cardiorrespiratorio de aportar el mismo (80). Para ilustrar este punto, en los animales que se ejercitan al máximo la mitocondria tiene una capacidad respiratoria fija, con un valor invariante de 4-5 mL de O2.mL-1 de mitocondria por minuto a través de las especies (80). Sin embargo, la capacidad respiratoria de una mitocondria aislada ha sido medida en 5.8 mL de O2.mL-1 de mitocondria por minuto (75). Usando estos valores, Taylor y Weibel (80) concluyeron que los animales son capaces de explotar el 60-80% de la capacidad oxidativa in vitro cuando se ejercitan al VO2 máx. La razón de que la mitocondria no puede explotar completamente su capacidad oxidativa es un resultado de la limitación del transporte de O2 impuesto por el sistema cardiovascular central.
Encontrando Consenso sobre los Factores Limitantes del Máximo Consumo de Oxígeno
Los fisiólogos han preguntado frecuentemente, “¿Cual es el factor limitante del VO2 máx.?”. La respuesta depende de la definición de factor limitante y del modelo experimental usado para tratar el problema (R.B. Armstrong, comunicación personal, Febrero, 1999).

Tabla 1. Masa corporal y parámetros de transporte de oxígeno del sistema
cardiovascular en caballos y novillos, durante el ejercicio máximo (media±DS).
Adaptado de Jones et al. (J. Appl. Physiol. 67:862-870, 1999). VO2
máx., máximo consumo de oxígeno; HR máx., frecuencia cardiaca máxima, CO máx.,
máximo gasto cardiaco; C a-v O2, máxima diferencia arteriovenosa de
oxígeno, Hct, hematocrito.
Si uno habla acerca de un ser humano intacto que realiza ejercicio máximo, para todo el cuerpo, entonces el sistema cardiorrespiratorio es el factor limitante (36, 74, 84). Si uno discute los factores que limitan el incremento del VO2 en un miembro trasero de un perro, entonces el gradiente de difusión periférica es el limitante (45). Si uno habla acerca de los factores que explican la diferencia en el VO2 máx. entre las especies, el contenido mitocondrial y la capacidad de transporte de O2 son ambos importantes (80).
Wagner, Hoppeler, y Saltin (86) fueron exitosos en conciliar los diferentes puntos de vista sobre los factores limitantes del VO2 máx. Ellos concluyeron que mientras el VO2 máx. está ampliamente relacionado al volumen mitocondrial entre el rango de especies, en cualquier caso individual el VO2 máx. es determinado por el aporte de O2 a los músculos. Ellos establecieron que en humanos “…la capacidad catabólica de la miosin-ATPasa es tal que la misma supera con mucho la capacidad del sistema respiratorio de transportar energía aeróbicamente. Así, el VO2 máx. debe ser determinado por la capacidad de transportar el O2 a las mitocondrias musculares por medio del sistema de transporte de O2, y no por las propiedades de la maquinaria contráctil muscular (86)”.
Wagner, Hoppeler, y Saltin (86) mantuvieron que no hay ningún único factor limitante del VO2 máx. Ellos concluyeron que “…todos y cada uno de los pasos en la ruta del O2 contribuyen en una forma integrada para determinar el VO2 máx., y una reducción en la capacidad de transporte de cualquiera de los pasos va a previsiblemente reducir el VO2 máx. (85, 86)”. Por ejemplo, una reducción en la PO2 inspirada en la altura va a resultar en una disminución del VO2 máx. (22, 31, 53). Una reducción del nivel de hemoglobina en la anemia va a resultar en una disminución del VO2 máx. (36, 78). Una reducción en el gasto cardíaco con beta-bloqueantes cardioselectivos va a resultar en una disminución del VO2 máx. (84). También hay situaciones donde el aporte de sustrato (no O2) es el factor limitante. Por ejemplo, los defectos metabólicos en el músculo esquelético, como la enfermedad de McArdle (deficiencia de fosforilasa) o deficiencia de fosfofructoquinasa, van a resultar en una disminución del VO2 máx. (54).
En el campo de la fisiología del ejercicio, son discutidos los factores limitantes del VO2 máx., usualmente en referencia a los sujetos humanos, sin enfermedades metabólicas, que realizan ejercicio para todo el cuerpo, al nivel del mar. Bajo estas condiciones, la evidencia demuestra claramente que es principalmente la capacidad del sistema cardiorrespiratorio (i.e., corazón, pulmones, y sangre) de transportar O2 a los músculos, y no la capacidad de las mitocondrias musculares de consumir O2, lo que limita el VO2 máx. Nosotros concluimos que hay un acuerdo general con respecto a los factores que limitan el VO2 máx., y que este acuerdo está basado en evidencia científica válida. En general, los 75 años de investigación subsiguiente han proporcionado fuerte apoyo para las brillantes ideas de Hill et al. (41, 42).
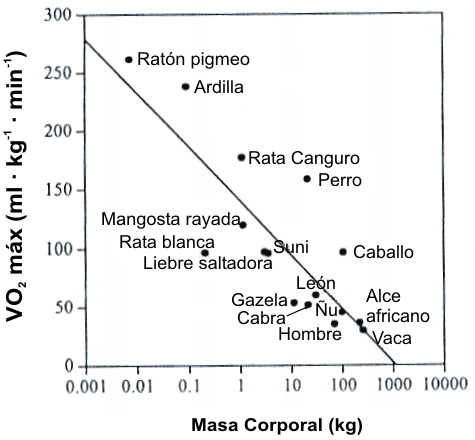
Figura 9. Máximo consumo de oxígeno (VO2 máx./Mb en ml.kg-1.min-1)
de varias especies animales en relación a la masa corporal (Mb). Los animales
más pequeños tiene valores de VO2 máx./Mb increíblemente altos, lo
cual se logra posiblemente por un incremento de la densidad mitocondrial (lo
cual permite a sus músculos consumir más oxígeno), así como por un incremento de
la capacidad para el transporte de oxígeno.
PARTE III: DETERMINANTES DEL RENDIMIENTO DE RESISTENCIA
Un principio importante en fisiología del ejercicio es que el trabajo requiere energía, y para mantener una tasa de trabajo específica o velocidad de carrera a través de una distancia larga, el ATP debe ser aportado a los puentes cruzados tan rápido como es usado. A medida que se incrementa la duración de un trabajo de máxima intensidad hay una mayor dependencia sobre la producción de ATP por medio de la fosforilación oxidativa para mantener el ciclo de los puentes cruzados. Consecuentemente, la tasa a la que el oxígeno es usado durante el ejercicio submáximo prolongado, es una medida de la tasa a la cual es ATP es generado. En nuestro artículo científico anterior (5) resumimos el entendimiento convencional acerca de cómo la captación de oxígeno esta relacionada al rendimiento en carreras de resistencia. Fueron dirigidas a nuestro trabajo una variedad de críticas, que iban desde sugerencias de que los datos de correlación fueron usados para establecer “causa y efecto”, hasta opiniones acerca de que nuestro modelo no fue adecuadamente explicado (63). En los párrafos siguientes vamos a resumir el modelo fisiológico que relaciona el consumo de oxígeno con el rendimiento en las carreras de distancia.
La Figura 10 muestra que el VO2 mantenido durante una carrera de resistencia (llamado el “VO2 de rendimiento” por Coyle (19)) es igual al producto del VO2 máx. del corredor y el porcentaje del VO2 máx. que puede ser mantenido durante la prueba. El porcentaje del VO2 máx. está relacionado al VO2 medido en el umbral anaeróbico (LT), de modo que para los eventos de resistencia el VO2 esta relacionado de cerca al VO2 en el LT. El VO2 máx. está limitado principalmente por factores cardiovasculares centrales (ver Parte II arriba), mientras que el porcentaje del VO2 máx. que puede ser mantenido esta relacionado principalmente a las adaptaciones en los músculos como resultado del entrenamiento prolongado (44). La velocidad real de carrera lograda a esta tasa de generación oxidativa de ATP (el VO2 de rendimiento) esta determinada por la capacidad del individuo de traducir la energía (e.g., economía de carrera) al rendimiento (19, 20). Nosotros vamos a resumir nuevamente el rol de cada una de estas variables para el rendimiento en las carreras de distancia.
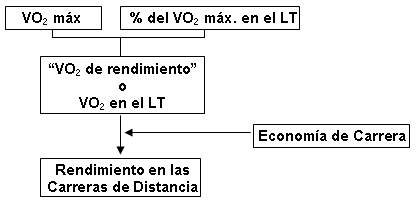
Figura 10. Diagrama simplificado de relación entre la potencia aeróbica
máxima (VO2 máx.), el porcentaje de potencia aeróbica máxima (% del
VO2 máx.) y la economía de carrera y como se relacionan con el
rendimiento en la carrera de distancia.
Rol del Máximo Consumo de Oxígeno en el Rendimiento de Carrera
Previamente nosotros planteamos “el VO2 máx. establece el limite superior para el rendimiento en los eventos de resistencia” (5), y no que es “el mejor predictor de la capacidad atlética” ((63), p. 1381). Los datos de Costill et al. (17) fueron presentados para mostrar la correlación inversa (r= -0.91) entre el VO2 máx. y el tiempo en una carrera de 16 km. Estos investigadores usaron sujetos con un amplio rango de valores de VO2 máx. (54.8 a 81.6 ml.kg-1.min-1) para examinar esta relación. Este fue un diseño de investigación apropiado para observar si existe una correlación entre estas dos variables, ya que tal relación debe ser evaluada a través de un rango de valores apropiado. Si uno tuviera que reducir el rango de valores para los cuales esta relación es examinada, el coeficiente de correlación se aproximaría a cero a medida que el rango de valores se aproxime a cero. Consecuentemente, nosotros reconocemos que el VO2 máx. no fue un buen predictor del rendimiento en corredores con valores similares de VO2 máx. ((5) p. 598). Si Costill et al. (17) hubieran encontrado que la correlación entre el VO2 máx. y el tiempo en una carrera de 16 km en un grupo diverso de corredores era de un r= -0.09 en vez de un r= -0.91, debería haber habido poco debate. Nosotros estamos de acuerdo en que una alta correlación no implica “causa y efecto”, sin embargo, el hecho de simplemente descartar una alta correlación entre dos variables que tienen una alta validez de construcción podría resultar en la pérdida de un punto importante para el investigador.
El VO2 máx. esta directamente vinculado a la tasa de generación de ATP que puede ser mantenida durante una carrera de distancia, aunque las carreras de distancia no son corridas al 100 % del VO2 máx. La tasa de generación de ATP es dependiente del VO2 (ml.kg-1.min-1) que puede ser mantenido durante la carrera, el cual es determinado por el VO2 máx. del sujeto y el porcentaje del VO2 máx. al cual el sujeto puede rendir (Figura 10). Por ejemplo, para completar una maratón en 2:15, debe ser mantenido un VO2 de 60 ml.kg-1.min-1 a través de toda la carrera. Consecuentemente, aún si una maratón pudiera ser corrida al 100% del VO2 máx., el corredor necesitaría un VO2 máx. de 60 ml.kg-1.min-1 para el rendimiento anterior. Sin embargo, ya que el maratón es corrido de manera característica al 80-85% del VO2 máx., los valores de VO2 máx. necesarios para tal rendimiento serían de 70.5-75 ml.kg-1.min-1. De esta forma el VO2 máx. establece el límite superior para producción de energía en eventos de resistencia, pero no determina el rendimiento final. Así como establecimos en el artículo anterior (5) no hay duda que los corredores varían en economía de carrera, así como en el porcentaje del VO2 máx. que puede ser mantenido en una carrera; ambas variables tienen un impacto dramático sobre la velocidad que puede ser mantenida en una carrera de resistencia. Esto va a ser discutido en los párrafos siguientes.
Economía de Carrera
La eficiencia mecánica es el índice de trabajo realizado sobre la energía gastada. El término “economía de carrera” es usado para expresar la captación de oxígeno necesaria para correr a una velocidad dada. Esto puede ser mostrado graficando el consumo de oxígeno (ml.kg-1.min-1) versus la velocidad de carrera (m.min-1) o simplemente expresando la economía como la energía requerida por unidad de masa para cubrir una distancia horizontal (ml de O2.kg-1.km-1). En nuestro artículo científico anterior nosotros mostramos que la economía de carrera explicaba algo de la variabilidad del rendimiento en la carrera de distancia de sujetos con valores de VO2 máx. similares (5). Los datos de Conley y Krahenbuhl (14) fueron usados para mostrar una correlación relativamente fuerte (r=0.82) entre la economía de carrera y el rendimiento en una carrera de 10 km en un grupo de corredores con valores de VO2 máx. similar, pero con un rango de tiempos de 10 km de 30.5-33.5 min. Como fue destacado en la refutación (63), cuando se examinó a los cuatro corredores más rápidos (10 km en 30.5-31 min) hubo una considerable variabilidad en la economía de la carrera (45-49 ml. kg-1.min-1 a 268 m.min-1) sugiriendo una falta de asociación entre las variables. Como fue mencionado arriba, esto puede ser esperado. Un coeficiente de correlación se va a acercar a cero a medida que el rango de valores de una de las variables (en este caso, tiempos de rendimiento que iban desde 30.5 hasta 31 min) se acerque a cero. Es de poca utilidad analizar una correlación a menos que el rango de valores sea suficiente para determinar si existe una relación.
Hay una relación lineal entre la velocidad de carrera submáxima y el VO2 (ml.kg-1.min-1) para cada individuo. Sin embargo, hay considerable variación entre los individuos acerca de cuanto oxígeno consume para correr a una velocidad dada, esto es, economía de carrera (6, 59). La Figura 11 muestra un gráfico de barras de la variación en la economía de carrera (expresada en ml.kg-1.min-1) entre grupos de sujetos que difieren en capacidad de carrera (59). El grupo de corredores de elite tuvo una mejor economía de carrera que los otros grupos de corredores, y todos los grupos de corredores fueron mejores que el grupo de sujetos desentrenados. Sin embargo, uno de los aspectos más reveladores de este estudio fue la variación dentro de cada grupo; hubo una diferencia de 20% entre el corredor menos y el más económico en cualquier grupo (59).
Una de las mejores descripciones acerca de cómo interactúan el VO2 máx. y la economía de carrera para afectar la velocidad de carrera, fue proporcionada por Daniels (20) en su descripción de la “velocidad al VO2 máx.” (v VO2 máx.). La Figura 12 muestra un gráfico de corredores hombres y mujeres iguales en términos de VO2 máx., pero que diferían en economía de carrera (21). Fue establecida una recta a través de la serie de puntos usada para construir una recta de economía de carrera, y fue extrapolada al VO2 máx. de los sujetos. Luego fue graficada una recta perpendicular del valor del VO2 máx. hasta el eje x para estimar la velocidad que el sujeto hubiera alcanzado en el VO2 máx. Esto es una estimación de la velocidad máxima que puede ser mantenida por la fosforilación oxidativa. En este ejemplo, la diferencia en la economía de carrera resultó en una clara diferencia en la velocidad que podría ser alcanzada si la carrera fuera corrida al VO2 máx. En una manera similar, la Figura 13 muestra el impacto que tiene una diferencia en el VO2 máx. en la vVO2 máx. en grupos con valores de economía de carrera similar. La diferencia de 14% en el VO2 máx. resultó en una diferencia de 14% en la v VO2 máx. Consecuentemente, está claro que tanto el VO2 máx. como la economía de carrera interactúan para establecer el límite superior de velocidad de carrera que puede ser mantenida por la fosforilación oxidativa. Sin embargo, ya que las carreras de distancia no son corridas al VO2 máx., la capacidad de un atleta para correr a un alto porcentaje del VO2 máx. tiene un impacto significativo sobre el rendimiento de carrera (17).
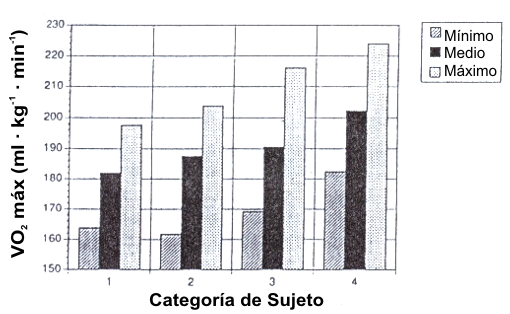
Figura 11. Valores mínimos, medios, y máximos de demanda aeróbica de la
economía de carrera para corredores de elite (Categoría 1), corredores de
subelite (Categoría 2), buenos corredores (Categoría 3), y sujetos desentrenados
(Categoría 4). Tomado de la referencia 59. Morgan, D.W., D.R., Bransford, D.L.
Costill, et al. Variation in the aerobic demand of running among trained and
untrained subjetcs. Med. Sci. Sports. Exerc. 27: 404-409, 1995. Usado con
permiso.
Porcentaje del Máximo Consumo de Oxígeno
La Figura 14, del clásico libro Fisiología del Trabajo Físico de Astrand y Rodahl (3) caracteriza el impacto que el entrenamiento tiene sobre la habilidad del sujeto de mantener un cierto porcentaje del VO2 máx. durante el ejercicio prolongado. Los individuos entrenados trabajaron al 87 % y 83 % del VO2 máx. por 1 h y 2 h, respectivamente, en comparación con solo el 50 % y 35 % del VO2 máx. para los individuos desentrenados. La figura muestra claramente el impacto que el % del VO2 máx. tiene sobre el VO2 real (rendimiento) que una persona puede mantener durante una prueba de resistencia. Además, la Figura 15, tomada del mismo texto, muestra como el VO2 máx. y el % del VO2 máx. cambian a través de los meses de entrenamiento. El VO2 máx. se incrementa durante los primeros 2 meses y se nivela, mientras que el % del VO2 máx. continua cambiando a través del tiempo. Consecuentemente, mientras que los cambios en el VO2 máx. y el % del VO2 máx. impactan los cambios en el rendimiento de un sujeto en la parte inicial de un programa de entrenamiento, los cambios subsiguientes en el VO2 de rendimiento son causados por cambios en el % del VO2 máx. solo. Esta figura clásica esta apoyada por un trabajo posterior que demostró que el VO2 en el LT (% del VO2 máx. en el LT) se incrementa mucho más como resultado del entrenamiento de lo que lo hace el VO2 máx. (ver revisión en (89) p. 59).
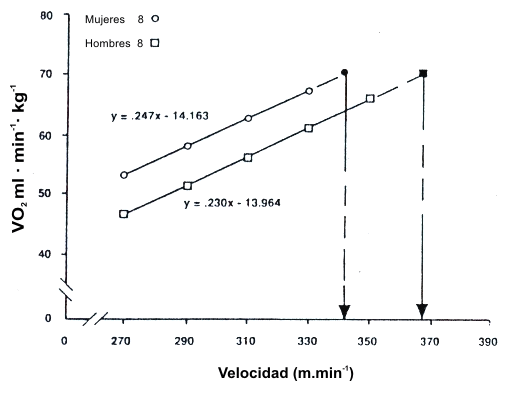
Figura 12. Comparación de corredores hombres y mujeres de igual VO2
máx.. Los hombres son significativamente favorecidos en la economía y en la v VO2
máx. (p<0.05). Tomado de la referencia 21. Daniels, J. and N. Daniels. Running
economy of elite male and elite female runners. Med. Sci. Sports. Exerc. 24:
483-489, 1992. Usado con permiso.
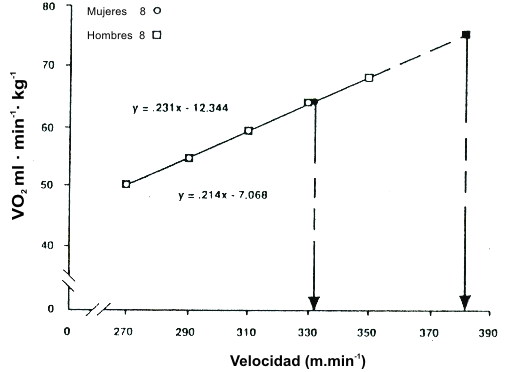
Figura 13. Comparación de corredores hombres y mujeres de igual economía de
carrera, a través de velocidades de evaluación comunes. Los hombres están
favorecidos en el VO2 máx. y en la v VO2 máx. (p<0.05).
Tomado de la referencia 21. Daniels, J. and N. Daniels. Running economy of elite
male and elite female runners. Med. Sci. Sports. Exerc. 24: 483-489, 1992. Usado
con permiso.
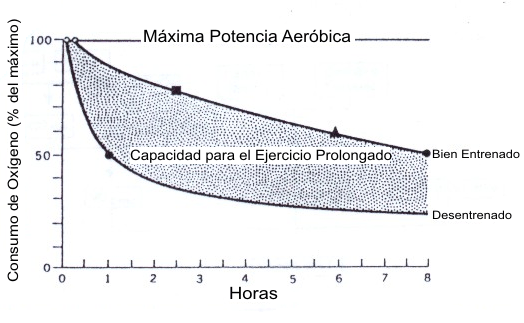
Figura 14. Una ilustración gráfica basada en una pocas observaciones que
mostraron aproximadamente el porcentaje de la potencia aeróbica máxima que un
sujeto, ella/el, puede utilizar durante trabajos de diferentes duraciones y como
es esto afectado por el estado de entrenamiento. Tomado de la referencia 3.
Astrand, P.,-O. and K. Rodahl. Textbook of Work Physiology. New York: McGraw-Hill,
1970, pp. 279-430. Usado con permiso.
Umbral Anaeróbico y Rendimiento de Resistencia
El modelo presentado antes en la Figura 10 muestra como el VO2 máx. y el % del VO2 máx. interactúan para determinar el VO2 de rendimiento y como la economía de carrera determina el rendimiento. En este modelo el VO2 en el LT integra tanto el VO2 máx. como el % del VO2 máx. En nuestro artículo científico anterior (5) nosotros usamos un modelo más detallado para mostrar que la velocidad de carrera en el LT integra las tres variables mencionadas antes (el VO2 máx., el % del VO2 máx., y la economía de carrera) para predecir el rendimiento de la carrera de distancia. Nosotros vamos ahora a usar el mismo modelo (Figura 16) para expandir nuestra discusión con centro en el umbral del lactato.
Para determinar el umbral del lactato, un sujeto completa una serie de evaluaciones a una velocidad de carrera incremental, y luego de cada evaluación es tomada una muestra de sangre para el análisis del lactato. La velocidad a la cual la concentración de lactato cambia de alguna forma (e.g., en una concentración absoluta, un punto de ruptura en la curva, una cantidad delta) es tomada como la velocidad en el LT y es usada como el predictor del rendimiento. Numerosos estudios han demostrado que los indicadores del LT son buenos predoctores del rendimiento en una variedad de actividades de resistencia (e.g., carrera, ciclismo, marcha) y tanto para poblaciones entrenadas como desentrenadas ((89) p. 49). En la mayoría de estos estudios la asociación entre el LT y el rendimiento de resistencia fue evaluada en grupos de atletas que eran heterogéneos en relación al rendimiento. Como fue discutido antes, este es un diseño apropiado para ver si existe una relación (correlación) entre las variables. De otro modo, si uno fuera a estrechar el rango de actividades (o el LT) sobre las cuales va a ser examinada esta relación, uno esperaría que la correlación fuera marcadamente reducida. Esto significa que aunque la velocidad en el LT explica la vasta mayoría de la variación en el rendimiento en las carreras de distancia (30) otros factores pueden todavía influenciar el rendimiento final. Si algún modelo pudiera explicar toda la variación en el rendimiento, las medallas de oro serían conseguidas en el laboratorio!
El modelo clásico que ha sido presentado por nosotros gira entorno a la proposición acerca de que la capacidad para mantener una alta velocidad de carrera esta conectada a la capacidad para mantener una alta tasa de producción oxidativa de ATP. Tanto los datos lógicos como empíricos apoyan aquella proposición: de otro modo para discutir se sugeriría que las fuentes de ATP “independientes de oxígeno” (anaeróbicas) son importantes en tales actividades- una clara imposibilidad dada la pequeña cantidad de energía potencial disponible por medio de tal proceso.
Se ha sabido por algún tiempo (43, 44) que la producción de lactato esta relacionada a un número de variables, incluyendo el contenido mitocondrial del músculo, como fue medido por medio de la actividad enzimática mitocondrial. Las variaciones en el LT a través de diversos grupos de atletas de resistencia y las mejoras que el LT que resultan del entrenamiento están conectadas a las diferencias y a los incrementos en la actividad enzimática mitocondrial, respectivamente ((19), (89), p. 49-59). Una explicación para esta conexión fue proporcionada por Holloszy y Coyle en 1984 (44). Cuando los músculos se contraen para alcanzar una producción de potencia submáxima específica, el ATP es convertido a ADP y Pi para potenciar los puentes cruzados, y las últimas dos moléculas, conducen reacciones en la célula para alcanzar la demanda de ATP asociada con la tasa de trabajo. En la célula muscular con relativamente pocas mitocondrias la concentración de ADP debe subir a un alto nivel para activar el número limitado de mitocondrias para alcanzar la demanda de ATP por medio de la fosforilación oxidativa. Esta alta concentración de ADP también activa otras rutas metabólicas, incluyendo la glucólisis, debido al efecto estimulador del ADP sobre la fosfofructoquinasa (PFK). Esto resulta en una mayor tasa de ruptura de carbohidratos, una acumulación de piruvato y NADH en el citoplasma de las fibras musculares, y un incremento en la producción de lactato (44, 50). Luego del entrenamiento hay un gran incremento (50-100 %) en el número de mitocondrias en los músculos implicados en la actividad.
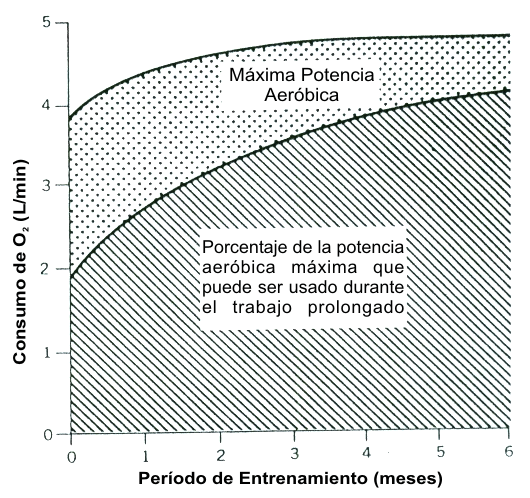
Figura 15. El entrenamiento causa un incremento en el máximo consumo de
oxígeno. Con el entrenamiento un sujeto es también capaz de utilizar un mayor
porcentaje de su máximo consumo de oxígeno durante el trabajo prolongado. Tomado
de la referencia 3. Astrand, P-O., and Rodahl. Textbook of Work Physiology. New
York: McGraw-Hill, 1970, pp. 279-430. Usado con permiso.
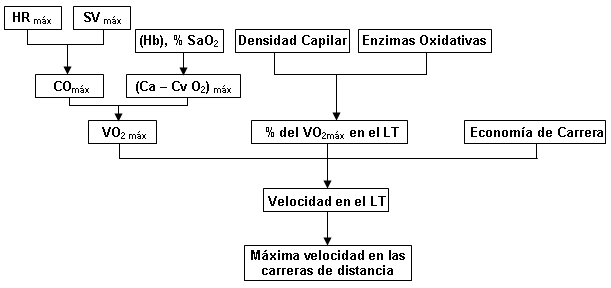
Figura 16. Resumen de las principales variables relacionadas al VO2
máx. y la máxima velocidad que puede ser mantenida en carreras de distancia.
Tomado de Bassett, D.R., Jr. And E.T. Howlwy. Maximal oxygen uptake: “clasical”
versus “contemporary” viewpoints. Med. Sci. Sports Exerc. 29: 591-603, 1997.
Usado con permiso.
Consecuentemente, a la misma tasa de trabajo el consume de oxígeno es compartido por un mayor número de mitocondrias, y la concentración de ADP no tiene que aumentar al mismo nivel que antes del entrenamiento para alcanzar la misma tasa de fosforilación oxidativa (VO2) después del entrenamiento. El menor nivel de ADP después del entrenamiento resulta en una menor estimulación de la PFK y una reducción en el consumo de carbohidratos, y el mayor número de mitocondrias incrementa la capacidad de usar las grasas como combustible. El resultado es una menor formación de lactato (44).
Como nosotros mencionamos a través de esta sección, la relación entre el VO2 máx. y el rendimiento (15), la economía de carrera y el rendimiento (14), y el % del VO2 máx. y el rendimiento (30), usó grupos con grandes variaciones en las variables independientes. A medida que uno reduce el rango de cada una de estas variables, las correlaciones son reducidas en magnitud o eliminadas, sugiriendo que otras variables también influencias el rendimiento. En vez de descartar las relaciones al tener poco valor, los investigadores han usado estas observaciones como motivación para examinar otros factores que podrían estar relacionados al rendimiento de resistencia. Un ejemplo excelente de esto es encontrado en un experimento de Coyle et al. (18).
Coyle et al. (18) estudió a 14 ciclistas (3-12 años de entrenamiento) los cuales eran similares en términos de VO2 máx. (así eliminado a este como variable) para examinar la relación entre el LT y el tiempo hasta la fatiga al 88% del VO2 máx. Los sujetos fueron divididos en un grupo LT alto (media= 81.5 % del VO2 máx.) y un grupo LT bajo (media= 65.8 % del VO2 máx.). El rendimiento en la evaluación al 88% del VO2 máx. resultó en grandes diferencias en el rendimiento (60.8 vs. 29.1 min), y la concentración de lactato post-ejercicio (7.4 vs. 14.7 mM) para los grupos LT alto y LT bajo, respectivamente. La diferencia en el rendimiento entre estos grupos que tenían el mismo VO2 máx., pero diferían en el VO2 máx. en el LT, fue consistente con el modelo descrito más arriba. De otro modo, el hecho de que el vasto lateral de los dos grupos tenía la misma actividad enzimática mitocondrial sugiere una ruptura en la línea de evidencia que une al VO2 máx. con en el LT y la actividad mitocondrial. Esto creó una rara oportunidad para los investigadores de estudiar dos grupos con el mismo VO2 máx. y la misma actividad enzimática mitocondrial, pero con diferencias substanciales en el rendimiento. Los investigadores examinaron la respuesta metabólica de los ciclistas a una evaluación de 30 min al 79% del VO2 máx. Ellos encontraron que mientras el grupo de LT bajo usó 69% más de carbohidratos durante esta serie de ejercicio que el grupo de LT alto, el grupo de LT bajo redujo su concentración de glucógeno muscular del vasto lateral un 134% más que el grupo de LT alto. Esta diferencia en la depleción de glucógeno muscular (relativa a la oxidación total de carbohidratos) sugiere que el grupo de LT alto fue capaz de distribuir la misma tasa de trabajo (y el VO2) en un mayor masa muscular, resultando en una menor carga sobre las fibras musculares reclutadas para hacer el trabajo. El uso de una mayor masa muscular también incrementó la masa de mitocondrias que compartían la producción de ATP por medio de la fosforilación oxidativa ((18, 19) y (87) p. 13). Consecuentemente, el estudio de Coyle et al. (18) indica que la masa de músculo implicada en la actividad (además de la densidad mitocondrial) contribuye al % del VO2 máx. en el LT (así como al rendimiento), de un modo consistente con el modelo antes mencionado.
LT, el Modelo Clásico, y Factores Ambientales
Noakes ha preguntado, “…porque debería el ejercicio de resistencia prolongado en el cual el consumo de oxígeno no es máximo y de este modo no limitante, ser determinado por el transporte de oxígeno a los músculos activos?” ((63) p. 1393). Esta es una buena pregunta acerca de porque los corredores de maratón pueden naturalmente correr a velocidades más altas y a altos valores de VO2 a través de distancias más cortas, pero no sin algunas consecuencias metabólicas. Durante el ejercicio submáximo, el transporte de oxígeno a los músculos está ligado de cerca de la demanda de oxígeno mitocondrial la cual es activada por la carga celular (i.e., (ADP + Pi)) impuesta por el ejercicio. Como fue mencionado antes, la misma carga celular también activa a otras rutas metabólicas, y notablemente, a la glucólisis. Si un maratonista elige correr a una velocidad arriba del LT, la carga celular incrementada necesitada de conducir el VO2 a un nivel más alto, también acelerara la glucólisis. Esto repletaría la limitada reserva de carbohidratos a una tasa más alta; el incremento resultante en la acumulación de lactato sanguíneo sería causado tanto por un incremento en la formación de lactato como por una disminución en la remoción del mismo (7, 50). Dada la necesidad obligada de carbohidratos a altas intensidades de ejercicio (13) y al impacto negativo de la acumulación de iones hidrógeno en la función muscular (29, 56, 57, 60), ninguno de estos cambios es consistente con ser capaz de mantener un ritmo más rápido, a través de una distancia de maratón.
En este y en otros artículos científicos anteriores (5) nosotros tratamos de explicar como las variables de VO2 máx., porcentaje del VO2 máx., y economía de carrera pueden explicar la vasta mayoría de las variaciónes en los rendimientos en las carreras de distancia (30, 89). Además, el modelo también explica el impacto de ciertos factores ambientales sobre el rendimiento de resistencia. La exposición aguda a alturas moderadas resulta en una disminución en la saturación arterial de oxígeno y el VO2 máx. (22, 53). Consecuentemente, el “VO2 de rendimiento” disminuye aunque los corredores puedan todavía rendir a porcentajes similares del VO2 máx., y el rendimiento en eventos de resistencia es afectado de manera adversa (22, 31). Históricamente, este efecto de una menor PO2 causando una disminución en el LT fue interpretado como una “falta de oxígeno” en el músculo. Sin embargo, es ahora reconocido que la menor PO2 resulta en una mayor carga celular para alcanzar el mismo VO2 en estado estable a una tasa de trabajo submáxima fija (50). Estas circunstancias van a resultar en una mayor tasa de glucólisis, una acumulación de NADH+, y un incremento de la producción de lactato.
Los tiempos de rendimiento en la maratón son afectados de manera adversa por altas temperaturas ambientales, siendo los tiempos de rendimiento óptimos a una temperatura de 12-13 °C (34), con una disminución de 40 s esperada por cada 1 °C de aumento en la temperatura (33). El ejercicio en el calor incrementa la tasa de oxidación de carbohidratos, conduciendo a una mayor tasa de depleción de glucógeno muscular y a mayores concentraciones de lactato sanguíneo durante el trabajo prolongado (32). Consecuentemente, los cambios en el metabolismo que resultan de la exposición aguda al calor o la altitura están asociados con una disminución en el rendimiento de resistencia, consistente con el modelo.
Comentario
En conclusión, el modelo “clásico” de VO2 máx. desarrollado por Hill et al. (41, 42) ha sido modificado y expandido por numerosos investigadores. Nosotros tenemos ahora un entendimiento mucho más completo de los determinantes del rendimiento de resistencia de lo que tenían los científicos del ejercicio de 1920. En retrospectiva, Hill et al. (41, 42) se equivocaron en algunos detalles, como la idea de un índice estricto 1/1 de déficit de O2/deuda de O2. Sin embargo, Hill es digno de reconocimiento por su rol principal en el descubrimiento de vías no oxidativas en el músculo aislado de rana y la aplicación de este descubrimiento a los humanos que realizan ejercicio (9, 51). El trabajo de Hill determino a la disciplina emergente llamada Fisiología del Ejercicio (37, 48), y sus ideas continúan influenciando aún en la actualidad.
Hill recibió críticas por sus teorías y recomendó a otros investigadores a que analizaran críticamente sus ideas científicas ((40) p. 363). Esta claro que el divisó que los errores en la interpretación y el debate científico de los méritos de las teorías competentes eran una parte necesaria del progreso ((51) p. 82). En Pruebas y Pruebas en Fisiología (Trials and Trials in Physiology), Hill estableció que, “el conocimiento avanza por la continua acción y reacción entre las hipótesis de un lado y la observación, cálculos, y experimentos del otro ((40), p. 361)”. De manera contraria al enfoque acerca de que las teorías clásicas representan “estructuras antiguas y sin validez” (62), nosotros arribamos a un punto de vista muy distinto. Nuestra conclusión es que las teorías de Hill han servido como una estructura teórica ideal. El trabajo que ha sido desarrollado a partir de esta estructura ha permitido a los científicos del ejercicio aprender mucho acerca de los factores fisiológicos que gobiernan el rendimiento atlético.
Agradecimientos
El apoyo para la realización del estudio fue proporcionado por la University of Tennessee, Knoxville, Exhibit, Perfomance, and Publication Expense (EPPE) Fund.
Dirección para correspondencia
David R. Bassett, Jr., Department of Exercise Science and Sport Management, University of Tennessee, 1914 Andy Holt Ave., Knoxville, TN 37996. Correo electrónico: DBassett@utk.edu
Referencias
1. ANDERSEN, P. and J. HENRIKSSON (1977). Capillary supply of the quadriceps femoris muscle of man: adaptive response to exercise. Capillary supply of the quadriceps femoris muscle of man: adaptive response to exercise
2. ASTRAND, P.- O (1952). Experimental Studies of Physical Working Capacity in Relation to Sex and Age. Cpenhagen: Ejnar Munksgaard, pp. 23-27
3. ASTRAND, P.-O. and K. RODAHL (1970). Textbook of Work Physiology. New York: McGraw-Hill, pp. 279-430
4. ASTRAND, P.-O. and B. SALTIN (1961). Oxygen uptake during the first minutes of heavy muscular exercise. J. Appl. Physiol. 16:971-976
5. BASSETT D. R. and E. T. HOWLEY (1977). Maximal oxygen uptake: classical versus contemporary viewpoints. Med. Sci. Sports Exerc. 9:41-44
6. BRANSFORD, D. R. and E. T. HOWLEY (1977). Oxygen cost of running in trained and untrained men and women. Med. Sci. Sports Exerc. 9:41-44
7. BROOKS, G (1985). Anaerobic Threshold: review of the concert and directions for future research. Med. Sci. Sports Exerc. 17:22-31
8. BUICK, F. J., N. GLEDHILL, A. B. FROESE, L. SPRIET, and E. C. MEYERS (1980). Effect of induced erythrocythemia on aerobic work capacity. J. Appl. Physiol. 48:636-642
9. CASTIGLIONI, A (1947). A History of Medicine. New York: Alfred A. Knopf, pp 942-943
10. CERRETELLI, P. and P. E. DI PRAMPERO (1987). Gas exchange in exercise. In: Hanbook of Physiology. A. P. Fishman, L. E. Farhi, S. M. Tenney and S. R. Geiger (Eds.). Bethesda, MD: American Physiological Society, pp. 297-339
11. CHRISTENSEN, E. H (1931). Beitrage zur Physiologie wchwerer korperlicher. IV: Mitteilung: die Pulsfrequenz wahrend und unmittelbar nach schwerer korperlicher Arbeit. Arbeits Physiol. 4:453-469
12. CLAUSEN, J. P., K. KLAUSEN, B. RASMUSSEN, and J. TRAP-JENSEN (1973). Central and peripheral circulatory changes after training of the arms and legs. Am. J. Physiol. 225:675-682
13. COGGAN, A. R. and E. F. COYLE (1991). Carbohydrate ingestion during prolonged exercise: effect on metabolism and performance. Exerc. Sport Sci. Rev. 19:1-40
14. CONLEY, D.L. and G. KRAHENBUHL (1980). Running economy and distance running performance of highly trained athletes. Med. Sci. Sports Eserc. 12:357-360
15. COSTILL, D. L (1970). Metabolic responses during distance running. J. Appl. Physiol. 28:251-255
16. COSTILL, D. L., W. J. FINK, and M. L. POLLOCK (1976). Muscle fiber composition and enzyme activities of elite distance runners. Med. Sci. Sports 8:96-100
17. COSTILL, D. L., H. THOMASON, and E. ROBERTS (1973). Fractional utilization of the aerobic capacity during distance running. Med. Sci. Sports Eserc. 5:248-252
18. COYLE, E. F., A. R. COGGAN, M. R. HOPPER, and T. J. WALTERS (1988). Determinants of endurance in well-trained cyclists. J. Appl. Physiol. 64:2622-2630
19. COYLE, E. F. and J. O. HOLLOSKY (1995). Integration of the physiological factors determining endurance performance ability. In: Exercise and Sport Sciences Reviews. J. O. Holloszy (Ed.). Baltimore, MD: Williams & Wilkins, pp. 25-63
20. DANIELS, J (1985). A physiologists view of running economic. Med. Sci. Sports Exerc. 17:332-338
21. DANIELS, J. and N. DANIELS (1991). Running economy of elite male and female runners. Med. Sci. Sports Exerc. 24:483-489
22. DANIELS, J. and N. OLDRIDGE (1970). The effects of alternate exposure to altitude and sea level on world-class middle-distance runners. Med. Sci. Sports 2:107-112
23. DAVIDSON, A. C., R. LEACH, R. J. D. GEORGE, and D. M. GEDDES (1988). Supplemental oxygen and exercise ability in chronic obstructive airway disease. Thorax 43:965-971
24. DEMPSEY, J. A (1986). Is the lung built for exercise?. Med. Sci. Sports Exerc. 18:143-155
25. DEMPSEY, J. A., P. HANSON and K, HENDERSON (1984). Exercise-induced arterial hypoxemia in healthy humans at sea-level. J. Physiol. (Lond.) 355:161-175
26. DUNCAN, G. E., E. T. HOWLEY, and B. N. JOHNSON (1997). Applicability of VO2max criteria: discontinuous versus continuous protocols. Med. Sci. Sports. Exerc. 29:273-278
27. EKBLOM, B., P. O. ASTRAND, B. SALTIN, J. STRENBERG, and B. WALLSTROM (1968). Effect of training on circulatory response to exercise. J. Appl. Physiol. 24:518-528
28. ERBLOM, B., G. WILSON, and P. O. ASTRAND (1976). Central circulation during exercise after venesection and reinfusion of bed blood cells. J. Appl. Physiol. 40:379-383
29. FABIATO, A. and F. FABIATO (1978). Effects of pH on the myofilaments and the sarcoplasmic reticulum of skinned cells from cardiac and skeletal muscle. J. Physiol. 276:233-255
30. FARRELL, P. A., J. H. WILMORE, E. F. COYLE, J. E. BILLING, and D. L. COSTILL (1979). Plasma lactate accumulation and distance running performance. Med. Sci. Sports Exerc. 11:338-344
31. FAULKNER, J. A., J. KOLLIAS, C. B. FAVOUR, E. R. BUSKIRK, and B. BALKE (1968). Maximum aerobic capacity and running performance at altitude. J. Appl. Physiol. 24:685-691
32. FINK, W., D. L. COSTILL, and P. J. V. HANDEL (1975). Leg muscle metabolism during exercise in the heat and cold. Eur. J. Appl. Physiol. 34:183-190
33. FOSTER, C. And J. DANIELS (1975). Running by the numbers. Runners World 10:14-17
34. FEDERICK, E. E (1983). Hot times. Running 9:52
35. GLEDHILL, H (1982). Blood doping and related issues: a brief review. Med. Sci. Sports Exerc. 14:183-189
36. GLEDHILL, N (1985). The influence of altered blood volume and oxygentransport capacity on aerobic performance. Exerc. Sport Sci. Rev. 13:75-93
37. HAMLEY, E. J. A. V. HILL, C. H., F. R. S. (1976). An appreciation. Br. J. Sports Med 10:195
38. HERIKSSON, J. And J. S. REITMAN (1977). Time course of changes in human skeletal muscle succinate dehydrogenase and cytochrome oxidase activities and maximal oxygen uptake with physical activity and inactivity. Acta Physiol. Scand. 99:91-97
39. HERBST, R (1928). Der Gasstoffwechsel als Mass der korperlichen Leistungsfahigkeit. I. Mitteilung: die Bestimmung des Sauerstoffaufnahmevermogens bein Gesuden. Deut. Arch. Klin. Med. 162: 33-50
40. HILL, A. V (1966). Trails and Trials in Physiology. Bibliography, 1909-1964, with Reviews of certain Topics and Methods and a Reconnaissance for Further Research. Baltimore, MD: W. and W., pp. 359-368
41. HILL, A. V., C. N. H. LONG, and H, LUPTON (1924). Muscular exercise, lactic acid and the supply and utilization of oxygen: Parts VII-VIII. Proc. Roy. Soc. B 97:155-176
42. HILL, A. V. And H. LUPTON (1923). Muscular exercise, lactic acid, and the supply and utilization of oxygen. Q. J. Med. 16:135-171
43. HOLLOSZY, J. O (1973). Biochemical adaptations to exercise: aerobic metabolism. Exerc. Sport Sci. Rev. 1:45-71
44. HOLLOSZY, J. O. and E. F. COYLE (1984). Adaptations of skeletal muscle to endurance exercise and their metabolic consequences. J. Appl. Physiol. 56:831-838
45. HONIG, C. R., R. J. CONNETT, and T. E. J. GAYESKI (1992). O2 transport and its interaction with metabolism: a systems view of aerobic capacity. Med Sci. Sports Exerc. 24:47-53
46. HOWLEY, E. T., D. R. BASSETT, and H. G. WELCH (1995). Criteria for maximal oxygen uptake: review and commentary. Med. Sci. Sports Exerc. 27: 2292-1301
47. ISSEKUTZ, B., N. C. BIRKHEAD, and K. RODAHL (1962). Use of respiratory quotients in assessment of aerobic capacity. J. Appl. Physiol. 17: 47-50
48. JOKL, E (1976). 90th Birthday of Professor A. V. Hill. J. Sports Med. 16: 349
49. JONES, J. H., K. E. LONGWORTH, A. LINDHOLDM. Et al (1989). Oxygen transport during exercise in large mammals. I. Adaptive variation in oxygen demand. J. Appl. Physiol. 67:862-870
50. KATZ, A. and K. SAHLIN (1988). Regulation of lactic acid production during exercise. J. Appl. Physiol. 65:509-518
51. KATZ, B. ARCHIBALD VIVIAN HILL (1978). Biographical Memoirs of Fellows of the Royal Society. 24:71-149
52. KLAUSEN, K., L. B. ANDERSEN, and I. PELLE (1981). Adaptive changes in work capacity, skeletal muscle capillarization and enzyme levels during training and detraining. Acta Physiol. Scand. 113:9-16
53. KOLLIAS, J. And E. BUSKIRK (1974). Exercise and altitude. In: Science and Medicine of Exercise and Sport. W. R. Johnson and E. R. Buskirk (Eds.). New York: Harper and Row, pp. 211-227
54. LEWIS, S. F. And R. G. HALLER (1989). Skeletal muscle disorders and associated factors that limit exercise performance. Exerc. Sport. Sci. Rev. 17:67-113
55. LINDHARD, J (1915). Ueber das minutenvolum des herzens bei ruhe und die muskelarbeit. Arch. F. D. Ges. Physiol. 1915, clxi:233-383
56. METZGER, J. M. And R. L. MOSS (1988). Depression of Ca2+ insensitive tension due to reduced pH in partially troponin-extracted Skinned skeletal muscle fibeers. Biophys. J. 54:1169-73
57. METZGER, J. M. AndR. L. MOSS (1987). Greater hydrogen-ion induced depression of tension and velocity in skinned single fibres of rat fast than slow muscles. J. Physiol. (Lond) 393:727-742
58. MITCHELL, J. H., B. J. SPROULE, and C. B. CHAPMAN (1958). The physiological meaning of the maximal oxygen intake test. J. Clin. Invest. 37:538-547
59. MORGAN, D. W., D. R. BRANSFORD, D. L. COSTILL, J. T. DANIELS, E. T. HOWLEY, and G. S. KRAHENBUHL (1995). Variation in the aerobic demand of running among trained and untrained subjects. Med. Sci. Sports Exerc. 27:404-409
60. NEKAMARU, Y. and A. SCHIWARTZ (1972). The influence of hydrogen ion concentration on calcium binding and release by skeletal sarcoplasmic reticulum. J. Gen Physiol. 59:22-32
61. NOAKES, T (1988). Implications of exercise testing for prediction of athletic performance: a contemporary perspective. Med. Sci. Sports Exerc. 20:319-330
62. NOAKES, T. D (1997). Challenging beliefs: ex Africa semper aliquid novi. Med. Sci. Sports Exerc. 29:571-590
63. NOAKES, T. D (1998). Maximal oxygen uptake: classical versus contemporary viewpoints: a rebuttal. Med. Sci. Sports Exerc. 30: 1381-1398
64. ORLANDER, J., K.H. KIESSLING, J. KARLSSON, and B. EKBLOM (1977). Low intensity training, inactivity and resumed training in sedentary men. Acta Physiol. Scand 101:351-362
65. POWERS, S. K., J. LAWLER, J. A. DEMPSEY, S. DODD and G. LANDRY (1989). Effects of incomplete pulmonary gas exchange of VO2max. J. Appl. Physiol. 66:2491-2495
66. RICHARDSON, R. S. and B. SALTIN (1998). Human muscle blood flow and metabolism studied in the isolated quadriceps muscles. Med. Sci. Sports Exerc. 30:28-33
67. ROOYACKERS, J. M., P. N. DEKHUIJZEN, C. L. VAN-HERWAARDEN, and H. T. FOLGERING (1997). Training with supplemental oxygen in patients with COPD and hypoxaemia at peak exercise. Eur. Respir. J. 10:1278-84
68. ROWELL, L. B (1986). Human Circulation: Regulation During Physical Stress. New York: Oxford University Press. pp. 213-286
69. ROWLAND, T. W (1993). Does peak VO2 reflect VO2max in children?: evidence from supramaximal testing. Med. Sci. Sports Exerc. 25:689-693
70. SALTIN, B (1985). Hemodynamic adaptations to exercise. Am. J. Cardiol. 55:42D-47D
71. SALTIN, B., B. BLOMQUIST, J. H. MITCHELL, R. L. JOHNSON, K. WILDENTHAL, and C. B. CHAPMAN (1968). Response to submaximal and maximal exercise after bed rest and after training. Circulation 38 (Suppl. 7): 1-78
72. SALTIN, B., J. HENRIKSSON, E. NYGAARD, and P. ANDERSEN (1977). Fiber types and metabolic potentials of skeletal muscles in sedentary man and endurance runners. Ann. N. Y. Acad. Sci. 301:3-29
73. SALTIN, B., K. NAZAR, D. L. COSTILL, et al (1976). The nature of the training response; peripheral and central adaptations to one-legged exercise. Acta Physiol. Scand. 96: 289-305
74. SALTIN, B. and S. STRANGE (1992). Maximal oxygen uptake: old and new arguments for a cardiovascular limitation. Med. Sci. Sports Exerc. 24:30-37
75. SCHEWERZMANN, K., H. HOPPELER, S. R. KAYAR, and E. R. WEIDEL (1988). Oxidative capacity of muscle and mitochondria: correlation of physiological, biochemical and morphometric characteristics. Proc. Natl. Acad. Sci, USA 86:1583-1587
76. SHEPHARD, R (1977). Endurance Fitness, 2nd Ed. Toronto and Buffalo: Univ. of Toronto Press, pp. 64-103
77. SPRIET, L. L, N. GLEDHILL, A. B. FROESE, and D. L. WILKES (1986). Effect of graded erythrocythemia on cardiovascular and metabolic responses to exercise. J. Appl. Physiol. 61:1942-1948
78. SPROULE, B. J., J. H. MITCHELL, and W. F. MILLER (1960). Cardiopulmonary physiological responses to heavy exercise in patients with anemia. J. Clin. Invest. 39:378-388
79. TAYLOR, C. R. and E. R. WEIBEL (1981). Design of the mammalian respiratory system I-IX. Respir. Physiol. 44:1-164
80. TAYLOR, D. R. and E. R. WEIBEL (1991). Learning from comparative physiology. In: The Lung: Scientific Foundations. J. B. W. R. G. Crystal. Et al. (ed.) New York: Raven Press, pp. 1595-1607
81. TAYLOR, C. R., E. R. WEIBEL, R. H. KARAS, and H. HOPPELER (1987). Adaptive variation in the mammalian respiratory system in relation to energetic demand I-VIII. Respir. Physiol. 69
82. TAYLOR, C. R., E. R. WEBEL, J. M. WEBER, et al (1996). Design of the oxygen and substrate pathways IVII. Exper. Biol. 199-1643-1709
83. TAYLOR, H. L., E. BUSKIRK, and A. HENSCHEL (1955). Maximum oxygen intake as an objective measure of cardio-respiratory performance. J. Appl. Physiol. 8:73-80
84. TESCH, P. A (1985). Exercise performance and beta-blockade. Sports Med. 2:389-412
85. WAGNER, P. D (1992). Gas exchange and peripheral diffusion limitation. Med. Sci. Sports Exerc. 24:54-58
86. WAGNER, P. D., H. HOPPELER, and B. SALTIN (1991). Determinants of maximal oxygen uptake. In: The Lung: Scientific Foundations. R. G. Crystal, J. B. West, et al. (Eds.) New York: Raven Press, pp. 1585-1593
87. WEIBEL, E. R (1984). The Pathway for Oxygen: Structure and function in the Mammalian Respiratory System. Cambridge, MA: Harvard University Press, pp. 1-425
88. WEIBEL, E. R (1996). A tribute to Charles Richard Taylor. J. Exper. Biol. 199: I-IV
89. WELTMAN, A (1995). The Blood Lactate Response to Exercise. Champaign, IL: Human Kinetics, pp. 1-117