
Factores Moduladores de la Potenciación Post- Activación y su Efecto sobre el Rendimiento en Actividades Explosivas Subsiguientes
Facoltà di Scienze Motorie, Università degli Studi di Verona, Verona, Italia.
Artículo publicado en el journal PubliCE, Volumen 0 del año 2009.
Publicado 25 de mayo de 2009
Resumen
Palabras clave: tasa de desarrollo de la fuerza, unidades motoras, producción de potencia, esprint, potenciación
POTENCIACION POST-ACTIVACION (PAP)
La potenciación post-activación (PAP) o potenciación post-tetánica (PTP) se refiere a los fenómenos por los cuales las características del rendimiento muscular se realzan extremadamente como resultado de su historia contráctil. [1,2]. La diferencia entre PAP y PTP está definida por la naturaleza de la contracción de acondicionamiento.
La PTP es inducida por una contracción tetánica involuntaria, mientras que la PAP es inducida por una contracción voluntaria [3,4] realizada normalmente a una intensidad máxima o cercana a la máxima. Para simplificar, este artículo se refiere a todas las respuestas de potenciación como la PAP, y se refiere a la actividad responsable de producir PAP como una contracción de acondicionamiento (CC).
La manifestación de PAP en el músculo esquelético ha sido registrada en muchos estudios, tanto en mamíferos como en humanos [5,17] provocando una discusión entre los artículos recientes de revisión sobre los mecanismos de la PAP [1,3] y su aplicación en el rendimiento deportivo [1-3,18]. Si se utiliza efectivamente, la PAP puede ser implementada en una rutina de entrenamiento de potencia para realzar el estímulo de entrenamiento de un ejercicio pliométrico [2, 18]. Ha sido probado que la inducción de PAP antes de la competencia podría ser mejor que las entradas en calor convencionales para el aumento del rendimiento en actividades deportivas explosivas que impliquen saltos, lanzamientos y carreras de velocidad [10]. Debido a la falta de consistencia en la literatura, las investigaciones permanecen inconclusas sobre los posibles beneficios de la PAP sobre el rendimiento y/o el entrenamiento en los deportes explosivos. La falta de consistencia de las investigaciones pasadas se debe probablemente a la interacción compleja de factores que influyen en el rendimiento agudo que sigue a una CC [1-3, 18]. Esta revisión analiza estos factores de confusión en mayor detalle, con el propósito de ayudar a informar y dirigir futuras investigaciones dirigidas a establecer la aplicación de la PAP (si la tiene) sobre el rendimiento y entrenamiento de las actividades deportivas explosivas.
MECANISMOS DE LA PAP
Ha sido propuesto que dos mecanismos principales son responsables de la PAP. Uno es la fosforilación de las cadenas livianas de miosina reguladoras (RLC), [1, 3, 4, 11, 12, 19, 20] y el otro es el incremento en el reclutamiento de unidades motoras de alto orden [1, 10, 20]. Existen también evidencias que sugieren que los cambios en el ángulo de peneación pueden contribuir a la PAP, y este mecanismo posible es brevemente introducido en este artículo.
FOSFORILACION DE LAS CADENAS LIVIANAS REGULADORAS
Una molécula de miosina es un complejo que contiene seis subunidades compuesta por dos cadenas pesadas (Figura 1) [21]. La terminación amino de cada cadena pesada, clasificada como cabeza de miosina, contiene dos RLCs, [9, 21] y cada RLC tiene un sitio vinculante específico para la incorporación de una molécula de fosfato. La fosforilación de la RLC es catalizada por la enzima quinasa de miosina de cadena liviana, la cual es activada cuando las moléculas de Ca2+ liberadas del retículo sarcoplásmico durante la contracción muscular, se unen a la calmodulina proteína reguladora de calcio [1, 5, 13, 21]. Se piensa que la fosforilación de las RLC potencia las contracciones subsiguientes por la alteración de la estructura de la cabeza de miosina y el alejamiento de su columna de filamento grueso [1, 21]. También ha sido demostrado que la fosforilación de las RLC deja más sensible la interacción actina-miosina al Ca2+ mioplásmico [13]. En consecuencia, la fosforilación de las RLC tiene su mayor efecto a una concentración relativamente baja de Ca2+, como es el caso de la contracción de las fibras o una contracción tetánica de baja frecuencia [1, 3, 4, 22, 23].
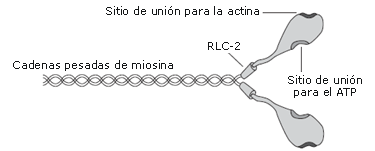
Figura 1. Molécula de miosina. Cada molécula está compuesta por dos cadenas pesadas de miosina. Las cadenas livianas reguladoras (RLC)-2 representan el par de RLCs ubicadas en el cuello de la cabeza de miosina. Cada RLC puede incorporar una molécula de fosfato, alterando la estructura de la cabeza de miosina. En cada cabeza de miosina hay un sitio de enlace para la actina y el adenosín trifostato (ATP).
Se ha reportado en muchos estudios en base a modelos animales con fibras aisladas [5, 7, 9, 13] (Figura 2), un incremento agudo en la fosforilación de las RLC, y una potenciación paralela de la tensión de la fibra luego de una estimulación tetánica de las fibras neurales eferentes específicas. Relativamente pocos estudios han intentado valorar una respuesta similar en el músculo esquelético humano. Stuart y col. [8] registraron un contenido de fosfato significativamente elevado en las RLC del músculo vasto lateral (p<0,01), y una potenciación significativa en la tensión de las fibras de los extensores de la rodilla, luego de una contracción isométrica máxima voluntaria de 10 segundos (MVC; p <0,05). Hubo también una correlación positiva aunque no significativa entre el alcance de la potenciación muscular y la cantidad de fosfato incorporado en las unidades individuales de RLC, y entre potenciación y porcentaje de fibras musculares de tipo II (p>0,05).
Smith y Fry [24] también tomaron muestras de biopsias musculares en el vasto lateral, y analizaron el rendimiento de la extensión dinámica de la pierna, antes y 7 minutos después de una MVC isométrica de 10 segundos. Los autores reportaron cambios no significativos en la fosforilación de las RLC y en el rendimiento de la extensión de la pierna para el total de la muestra (p>0,05). Los sujetos fueron entonces divididos en aquellos que respondieron al MVC con un incremento significativo en la fosforilación de las RLC, y aquellos que respondieron con una disminución significativa en la fosforilación de las RLC (p>0,05), pero entre los grupos no fueron encontradas diferencias significativas en el rendimiento en la extensión de pierna (p>0,05). Los factores metodológicos y diferencias en la distribución del tipo de fibra entre animales y humanos podrían explicar por qué el incremento observado en la fosforilación de las RLC luego de una CC no es tan consistente en humanos como en animales. No obstante, la significancia de la fosforilación de las RLC en el músculo esquelético humano sigue sin aclararse, y Stuart y col. [8] sugieren que otros factores podrían proporcionar el principal aporte a la PAP.
RECLUTAMIENTO AUMENTADO DE UNIDADES MOTORAS DE MAYOR JERARQUIA
Las investigaciones en animales han demostrado que una contracción tetánica isométrica inducida (causada por la estimulación de las fibras nerviosas aferentes específicas, las cuales activan alternadamente motoneuronas a adyacentes por medio de un impulso nervioso aferente; Figura 3) eleva la capacidad de transmisión del potenciales excitatorios a través del cruce sináptico en la médula espinal. Este estado beneficioso puede durar varios minutos luego de una contracción tetánica [10] y como resultado hay un aumento en los potenciales post-sinápticos, para el mismo potencial pre-sináptico durante la actividad subsiguiente [25, 26].
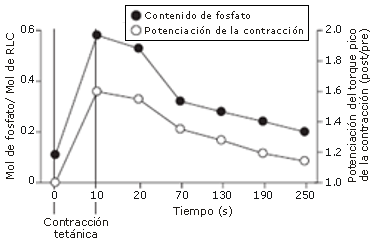
Figura 2. Transcurso de tiempo de la fosforilación de las cadenas livianas reguladoras (RLC) y la potenciación del torque pico de la contracción, luego de una contracción tetánica de preacondicionamiento de 10 segundos. La potenciación está representada como la proporción entre el valor del torque pico post-contracción voluntaria máxima (MVC) y el valor del torque pico pre-MVC (post/pre). Estos resultados indican la posible relación entre la fosforilación de las RLC y la potenciación de la tensión de la contracción (reproducido de Moore y Stull [7] con autorización).
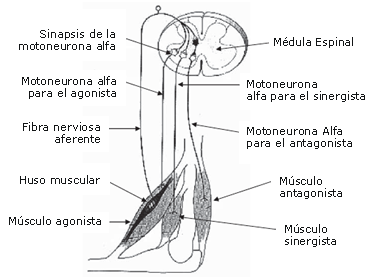
Figura 3. Impulsos nerviosos de una fibra aferente Ia. Un potencial de acción generado en la fibra aferente Ia viaja hasta la médula espinal, donde es transferido a las motoneuronas a adyacentes para el músculo agonista. El potencial de acción viaja luego directamente hacia el músculo agonista, iniciando el proceso de la contracción muscular.
Luscher y col.[26] propusieron un mecanismo posible como base de la elevada transmitancia de los potenciales de acción a través de conexiones sinápticas en la médula espinal. Para cada fibra nerviosa originaria (ej. fibra Ia), numerosas sinapsis se proyectan sobre cada motoneurona a. La activación de una motoneurona a trabaja de un modo todo o nada, donde la liberación del transmisor pre-sináptico debe coincidir con la sensibilidad del receptor post-sináptico. La falla de transmisión en diferentes conexiones sinápticas es un acontecimiento común durante la respuesta normal refleja o voluntaria, debido a una reserva de activación protegida de forma autónoma [26, 27]. Se sugiere la realización de una contracción tetánica inducida, de modo de disminuir la falla de transmición durante la actividad subsiguiente, por medio de una o la combinación de varias respuestas posibles. Estas incluyen un aumento en la cantidad de neurotransmisor liberado, un aumento en la eficacia del neurotransmisor, o una reducción en la falla de los puntos de bifurcación de los axones a lo largo de las fibras nerviosas aferentes [28].
Hirst y col. [27] aportaron evidencias que sostienen la disminución de la falla de transmisión monosináptica durante la actividad subsiguiente. Los investigadores estimularon fibras nerviosas aferentes de gato, y observaron un 54% de aumento en los potenciales post-sinápticos excitatorios (EPSPs) para el mismo estímulo pre-sináptico, luego de una contracción tetánica isométrica de 20 segundos. Mayores EPSPs representan mayor despolarización de la membrana de la motoneurona a, lo cual podría aumentar la probabilidad de la motoneurona a de alcanzar el umbral requerido para iniciar el potencial de acción y contraer posteriormente la fibra muscular de esa unidad motora.
Luscher y col.[26] también valoraron los EPSPs de motoneuronas a de gato, en respuesta a estimulación eléctrica. Los autores encontraron una correlación positiva significativa entre las resistencias de entrada de la motoneurona y la amplitud del EPSP, para un estímulo estándar (r=0.77; p<0,01; Figura 4a), donde la resistencia de entrada estuvo asociada con el tamaño de la motoneurona a (con una menor resistencia de entrada que representa una motoneurona más grande). Esto sugiere que la falla de transmisión monosináptica es mayor en las motoneuronas más grandes (aquellas responsables de la activación de unidades motoras de primer orden o de contracción rápida). A la inversa, cuando una contracción fue estimulada siguiendo una contracción tetánica de 10 segundos, Luscher y col. [26] encontraron una correlación negativa significativa entre la potenciación del EPSP y la resistencia de entrada de la motoneurona (r=-0.92; p<0,001; Figura 4b). Esto demuestra que la contracción tetánica disminuye la falla de transmisión que ocurre principalmente en las motoneuronas más grandes, lo cual resulta en un efecto PAP considerable en estas motoneuronas. Si una CC puede inducir un aumento en el reclutamiento de motoneuronas de primer orden en humanos, este efecto podría teóricamente aumentar la contribución de las fibras de contracción rápida a la contracción muscular, y por lo tanto mejorar el rendimiento en la actividad explosiva subsiguiente [10].
Estudios anteriores han valorado la onda H en humanos para investigar el efecto de una CC en el reclutamiento de motoneuronas [10, 29]. La onda H (reflejo H) es registrada en las fibras musculares usando electromiografía, y es el resultado de una descarga neural aferente en respuesta a un pulso único de estimulación submáxima del haz nervioso pertinente (ver Figura 5 para más detalles). Por lo tanto, un aumento en la onda H luego de una CC puede representar una disminución en la falla de transmisión en las conexiones sinápticas, y el posterior incremento en el reclutamiento de unidades motoras de primer orden. Gullich y Schmidtbleicher [10] estimularon el nervio tibial y valoraron los cambios en la amplitud de la onda H en el gastrocnemio, antes y después de MVCs isométricas de 5 segundos de los flexores plantares. Los autores reportaron una depresión en la amplitud de la onda H, 1 minuto después de las MVCs (-24%; p<0,05), pero una potenciación de la amplitud de la onda H 5 a 13 minutos después de las MVCs (+20%; p<0,01). La onda H, sin embargo, no fue normalizada a onda M máxima (la onda M es el homólogo eléctrico de la activación de todas las unidades motoras) [30]. No obstante, otros factores no relacionados a la activación central, como la actividad aumentada de la bomba de Na+-K+ en las fibras musculares [12, 14, 28] puede ser responsable de los resultados que observaron Gullich y Schmidtbleicher [10]. Sin embargo, otros estudios han reportado una potenciación en la amplitud de la onda H normalizada de 3 a 10 minutos después de ocho series de MVCs dinámicas [29] y 5 a 11 minutos después de 10 segundos de MVC isométrica [31].
Conjuntamente, estos resultados sugieren que la PAP aumenta la amplitud de la onda H en humanos (aunque después de una recuperación suficiente), y esto puede ser el resultado del aumento en el reclutamiento de motoneuronas de primer orden en la médula espinal. Debe determinarse aún, si una CC puede elevar el reclutamiento de motoneuronas y el rendimiento durante una contracción voluntaria subsiguiente, o no.
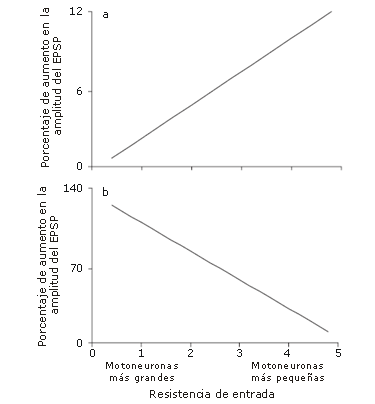
Figura 4. (a) Relación entre la resistencia de entrada de motoneuronas de gato, y la amplitud de sus potenciales excitatorios post-sinápticos (EPSP) en respuesta a la estimulación de las fibras nerviosas aferentes adyacentes. (b) Relación entre la resistencia de entrada de motoneuronas de gato, y el porcentaje de aumento (potenciación) en la amplitud de los EPSP, en respuesta a la estimulación de las fibras nerviosas aferentes adyacentes, luego de una tetania de 10 segundos. Aunque la amplitud de los EPSP sea mayor en las motoneuronas más pequeñas (aquellas con mayor resistencia de entrada), lo cual representa una mayor falla de transmisión en motoneuronas más grandes (a), la potenciación es mayor en las motoneuronas más grandes (aquellas con menor resistencia de entrada), lo que demuestra una disminución en la falla de transmisión en estas motoneuronas (b) [22].
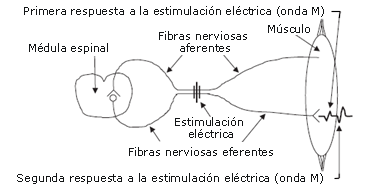
Figura 5. Producción de las ondas M y H. La estimulación de un nervio con un único impulso eléctrico submáximo evoca dos respuestas eléctricas en el músculo. La primer respuesta (onda M) es el resultado de un potencial de acción que viaja directamente a través de las fibras nerviosas eferentes (motoneuronas a). La segunda respuesta (onda H) es el resultado de un potencial de acción que viaja a través de la fibra nerviosa aferente hacia la médula espinal, donde es transmitido a la fibra nerviosa eferente adyacente, y hacia el músculo.
El efecto de las MVCs isometricas sobre el posterior reclutamiento voluntario de motoneuronas ha sido valorado usando la técnica de contracción interpolada (ITT). La ITT puede facilitar la medición de la activación de la motoneurona [32], comparando la amplitud de contracción máxima en reposo con aquella superpuesta sobre una MVC (para más detalles de la ITT por favor referirse a Folland y Williams [32] y Shield y Zhou [33]). Usando la ITT, Behm y col.[34] reportaron una disminución en la activación muscular voluntaria luego de 10 segundos de MVCs (p<0,05). Estos resultados están en contra de los mecanismos propuestos de PAP, pero pueden manifestar la preponderancia de fatiga central observada a través de este estudio (ver la sección volumen de la contracción de acondicionamiento).
No obstante, las investigaciones futuras deberían considerar el uso de ITT para investigar el mecanismo de PAP y su contribución al rendimiento subsiguiente.
CAMBIOS EN EL ANGULO DE PENEACION
El ángulo de peneación de un músculo (el ángulo formado por los fascículos y la aponeurosis interna) refleja la orientación de las fibras musculares en relación al tejido conectivo/tendón [35]. El ángulo de peneación afectará, por lo tanto, la transmisión de la fuerza a los tendones y huesos [35, 36]. La suma de las fuerzas de todas las fibras individuales que están siendo aplicadas al tendón pertinente durante la contracción muscular está reducida por un factor de cos (?) (donde ?=ángulo de peneación) [36]. Por consiguiente, los ángulos de peneación más pequeños tienen una ventaja mecánica respecto a la transmisión de fuerza al tendón [35, 36]. Usando ultrasonografía, Mahlfeld y col. [37] midieron ángulos de peneación en reposo del vasto lateral después de 3 segundos de MVCs isométricas. El ángulo de peneación inmediatamente después de las MVCs (15,7 º) no tuvo cambios respecto a los valores previos a la MVC (16,2 º); sin embargo, 3 a 6 minutos después de las MVCs, el ángulo de peneación disminuyó significativamente (14,4º; p<0,05). Este cambio podría solo ser equivalente al 0,9% de incremento en la transmisión de la fuerza a los tendones, pero es posible que este efecto contribuya a la PAP. Las contracciones de preacondicionamiento, sin embargo, probablemente también pueden incrementar el compliance del tejido conectivo/tendón [38] y esto puede contrarrestar cualquier aumento en la fuerza de transmisión causada por una disminución en el ángulo de peneación. Sin embargo, la posibilidad de que los cambios en la arquitectura del músculo contribuyan a la PAP justifica promover las investigacioes.
PAP Y POTENCIA MECANICA
El rendimiento en las actividades deportivas explosivas está en gran parte determinado por la potencia mecánica [10, 39-43]. La potencia mecánica puede definirse como la tasa a la cual la fuerza (F) es desarrollada sobre un intervalo de movimiento (d), en un período de tiempo específico (t) [P=F.d/t], o como fuerza multiplicada por velocidad (v) [P=F.v] [39, 40, 43]. Por consiguiente, aumentando el nivel de fuerza a una velocidad dada aumentará la potencia mecánica, y esto ha sido demostrado en modelos de fibras aisladas de rata/ratón [16, 17, 22].
De la misma manera, disminuyendo el tiempo sobre en el cual se aplica una fuerza específica, sin alterar la distancia sobre la cual dicha fuerza es aplicada, aumentará la velocidad, y en consecuencia la potencia mecánica. La PAP puede, por lo tanto, aumentar la fuerza y/o la velocidad de la contracción muscular, lo cual mejorará la potencia mecánica y el rendimiento deportivo asociado.
Hasta la actualidad, existe poca evidencia de que la PAP pueda aumentar la fuerza máxima. Esto es consecuente con la observación de que el aumento de la sensibilidad de la interacción miosina-actina al Ca2+ tiene poco o ningún efecto sobre el estado de la saturación del Ca2+, como el causado por altas frecuencias de estimulación (>20 Hz para la tetania, o 200 Hz para las contracciones voluntarias) [9, 22]. Stuart y col. [8] también encontraron que 10 segundos de MVC isométrica de los extensores de la rodilla no pudieron aumentar la velocidad de descarga máxima de las contracciones dinámicas subsiguientes. Aunque la PAP parece tener poco efecto en los extremos de la curva de fuerza-velocidad (Figura 6), se ha demostrado que aumenta la tasa de desarrollo de la fuerza (RFD) de las contracciones tetánicas obtenidas a cualquier frecuencia [9]. Un aumento en la RFD causa una curva de fuerza-velocidad menos cóncava (Figura 6), dando por resultado una mayor velocidad para una fuerza específica, o viceversa [3, 44]. Por lo tanto, la PAP puede mejorar el rendimiento en actividades que requieren producción de fuerza submáxima y velocidad [3, 11]. Normalmente, los atletas que participan en actividades deportivas explosivas no producirán fuerza máxima, porque la masa que intentan mover es a menudo relativamente pequeña (por ej. la masa corporal), pero deben aún superar esa masa, por lo que tampoco alcanzarán la máxima velocidad alcanzada sin carga [40]. En consecuencia, la PAP podría beneficiar el rendimiento de las actividades deportivas explosivas incrementando la RFD y así la potencia mecánica [3, 11].
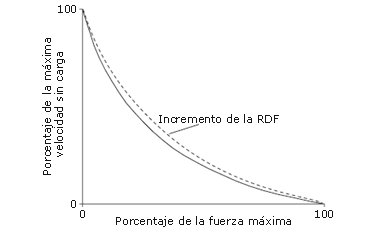
Figura 6. Relación entre fuerza y velocidad. La línea de puntos representa la curva de fuerza-velocidad menos cóncava debido al aumento en la tasa de desarrollo de la fuerza (RFD) [copiado de Sale [3] con autorización].
Hay consenso sobre la existencia de la PAP, pero para ser efectivamente utilizada en el rendimiento y/o entrenamiento, las investigaciones deben confirmar primero si la PAP puede ser inducida por una contracción voluntaria isométrica o dinámica, y luego demostrar que sus beneficios pueden conseguirse durante una actividad deportiva explosiva subsiguiente. Desafortunadamente, las mediciones de la PAP y de sus efectos sobre el rendimiento en una actividad deportiva explosiva subsiguiente en humanos es inconsistente. Además se conoce poco acerca del grado hasta el cual los mecanismos subyacentes propuestos de la PAP pueden jugar un rol en la inducción de una respuesta neuromuscular elevada.
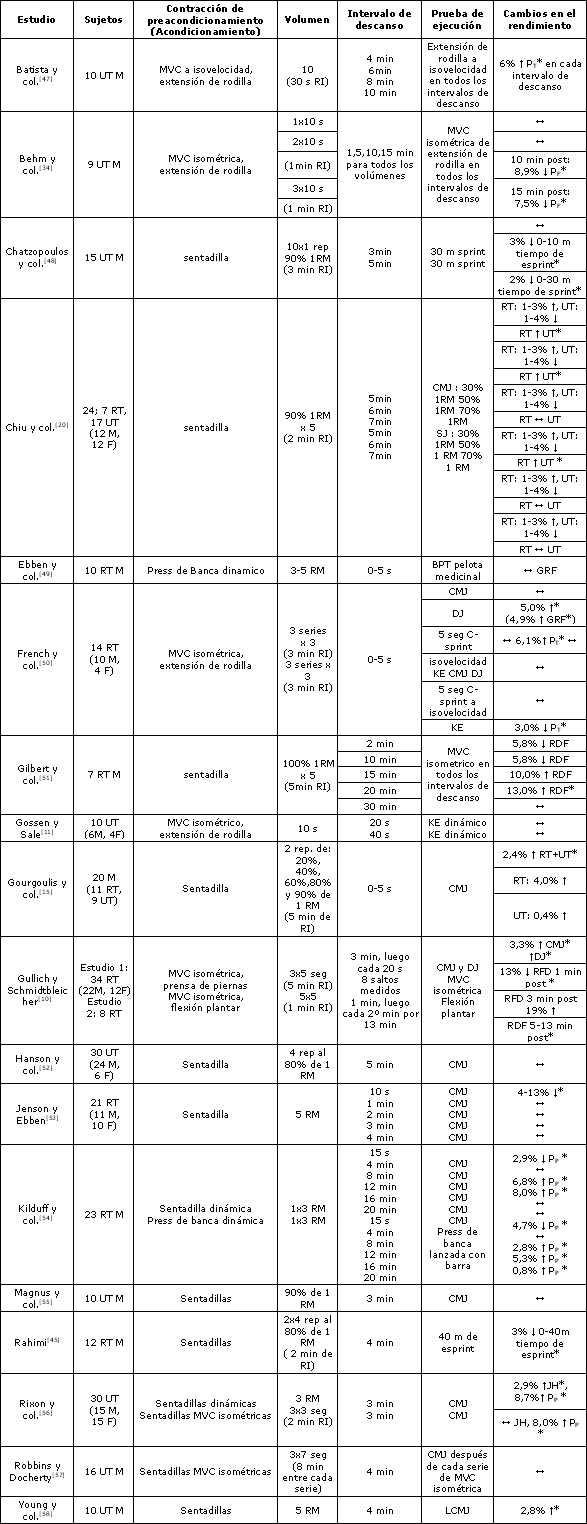
Tabla 1. Resumen de los estudios que han investigado los efectos de las contracciones de preacondicionamiento sobre una actividad subsiguiente. BPT=lanzamiento de press de banca; CMJ=salto con contramovimiento; C-esprint=esprint en bicicleta; DJ=salto en profundidad; F=mujeres; GRF=fuerza reactiva; JH= salto en alto; KE= extensiones de rodilla; LCMJ= salto con contramovimiento con carga; M= varones; MVC= contracciones voluntarias máximas, Pf= fuerza pico; PP= potencia pico; Pt= torque pico; RDF= tasa de desarrollo de la fuerza; RI= intervalo de descanso; RM= repetición máxima; RT= entrenado en resistencia; SJ= salto desde sentadilla; UT= no entrenado/recreacional; ? indica incremento; ? indica disminución; ? indica sin cambios; * indica diferencias significativas a un nivel p<0,05.
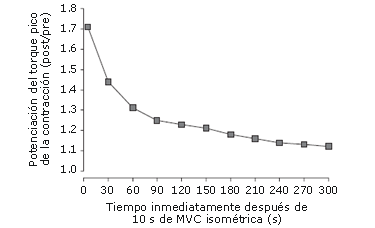
Figura 7. Transcurso de tiempo de la potenciación del torque pico de la contracción inmediatamente después de una contracción voluntaria máxima isométrica (MVC) [12]. La potenciación es representada como la proporción del valor de torque pico post-MVC y el valor de torque pico pre-MVC (post/pre).
EFECTOS AGUDOS DE LA PAP SOBRE LA ACTIVIDAD SUBSIGUIENTE
El rendimiento en las actividades deportivas explosivas depende predominantemente de la activación de grandes grupos musculares (ej. aductores/abductores del tobillo, rodilla, cadera y/o brazo). Por lo tanto, los estudios que evalúan los efectos de la PAP en grupos musculares pequeños han sido excluidos de las secciones siguientes. Es más, ha sido demostrado [45, 46] y está ampliamente aceptado que las contracciones de máxima intensidad o cercanas a la máxima intensidad (>80% de la MVC dinámica o isométrica) optimizan la PAP [4]. Por lo tanto, los estudios que evalúan los efectos de las contracciones submáximas sobre el rendimiento subsiguiente han sido también excluidos de las secciones posteriores. La Tabla 1 resume los estudios que han investigado los efectos de una CC voluntaria sobre la actividad voluntaria subsiguiente en humanos.
Coincidiendo con los resultados producidos por los estudios llevados a cabo sobre modelos de fibras aisladas de mamíferos, las investigaciones han reportado continuamente una respuesta contráctil mejorada luego de una CC en humanos. Hamada y col. [12] provocaron una contracción refleja en el nervio femoral antes de una MVC isométrica de 10 segundos de los extensores de la rodilla, 5 segundos después de la misma, y cada 30 segundos por 300 segundos después de dicha MVC. El PT (torque pico) de la contracción aumentó significativamente 5 segundos después de la MVC isométrica (+71%; p<0,01); sin embargo, 30 y 60 segundos después de la MVC, la potenciación de la contracción PT disminuyó +44% y +31%, respectivamente (p<0,01).
La potenciación siguió disminuyendo a una tasa más gradual por lo que restó del período de recuperación, pero estuvo aún en un +12% 300 segundos después de la MVC isométrica (p<0,01). Se han reportado en otros estudios hallazgos similares [6, 11, 59] que demuestran que la PAP pico es alcanzada inmediatamente después de un CC, pero al instante comienza a disminuir. La disminución de la PAP es rápida durante el primer minuto, pero luego se vuelve más gradual pareciéndose a una función exponencial (Figura 7).
Aunque se ha encontrado que una MVC isométrica eleva de manera consistente la tensión de la contracción subsiguiente, las evidencias que muestran que la PAP puede ser efectivamente utilizada para elevar el rendimiento en contracciones voluntarias subsiguientes, no es tan convincente.
Gossen y Sale [11] valoraron mecánicas de movimiento tanto de contracciones nerviosas como de contracciones voluntarias submáximas posteriores a una MVC isométrica de 10 segundos. Mientras que la MVC realzó el PT (p<0,01) de la contracción, el pico de velocidad de la extensión de rodilla posterior a una MVC fue significativamente más bajo que el ejecutado en una condición controlada. (326,7 vs. 341,6º.s-1; p<0,03). Estos resultados sugieren que aunque la MVC de 10 segundos induce PAP, también induce fatiga, y esta última es más dominante durante las contracciones voluntarias. Se ha propuesto, por lo tanto, que es el balance entre la PAP y la fatiga lo que determina si la respuesta contráctil subsiguiente es aumentada, disminuida o no presenta cambios [2].
PAP VERSUS FATIGA
El balance entre la PAP y la fatiga y sus efectos sobre las contracciones explosivas subsiguientes han sido observadas por varios estudios. Inmediatamente después de una CC, Gullich y Schmidtbleicher [10] y Gilbert y col. [51] reportaron una RDF isométrica disminuida o sin cambios, pero luego de una recuperación suficiente (4,5–12,5 minutos [10] y 15 minutos [51]) la RDF isométrica disminuyó significativamente (+10–24%; p<0,05). Se ha reportado también el mismo patrón sin cambios/disminución seguido por un aumento en el pico de potencia del salto con contramovimiento (CMJ) (+7–8%; p<0,05) [54] y esprint de 30 m (2–3%; p<0,05) [48] 8–12 minutos y 5 minutos después de una CC, respectivamente.
En conjunto, estos resultados sugieren que aunque los estudios sobre contracción han informado PAP máxima inmediatamente después de una CC (descripto en la sección 4, ver figura 7), la fatiga también se presenta en forma temprana. Es más, la fatiga parece ser más dominante en las etapas tempranas de la recuperación, y en consecuencia, el rendimiento en las actividades voluntarias subsiguientes está disminuido o no presenta cambios.
Sin embargo, la fatiga disminuye a una tasa más rápida que la PAP, y la potenciación del rendimiento puede notarse en algunos puntos durante el período de recuperación. La Figura 8 ilustra la relación PAP-fatiga y muestra como el efecto neto sobre las contracciones voluntarias subsiguientes podría ser muy diferente al efecto de las MVC sobre las contracciones nerviosas subsiguientes (representado en la Figura 7).
Existen también evidencias que indican que el período de recuperación puede no ser necesario para beneficiarse de la PAP, o que aún con un período de recuperación, el rendimiento en las actividades voluntarias subsiguientes puede permanecer sin cambios/disminuido. French y col. [50] no utilizaron período de recuperación, pero aún así observaron un aumento significativo tanto en la altura del salto en profundidad (DJ) como en PT de la extensión de la rodilla a isovelocidad (+5,0% y +6,1%, respectivamente; p<0,05), inmediatamente después de tres series de 3 segundos de MVC de extensiones isométricas de rodilla. De manera similar, Gourgoulis y col. [15] reportaron un incremento significativo en la altura del CMJ (+2.4%; p<0,05) inmediatamente después de la ejecución de dos repeticiones de sentadilla al 90% de una repetición máxima (1 RM).
Por el contrario, Chiu y col. [20] no pudieron detectar una mejora significativa en el pico de potencia de tres CMJ o tres saltos desde sentadilla (SJ) con carga (p>0,05), aún si fueron realizadas después de un período de recuperación de 5, 6 y 7 minutos, respectivamente, luego de 5 series de una repetición de sentadilla con el 90% de 1 RM. Los tres CMJ (5, 6 y 7 minutos post-activación), fueron ejecutados con diferentes cargas (30%, 50% y 70% de 1 RM, respectivamente), lo cual pudo haber afectado la producción de potencia pico, y haber hecho difícil establecer diferencias en el rendimientos en un transcurso de tiempo. Sin embargo, estos resultados fueron respaldados por los de Mangus y col. [55] quienes informaron ausencia de cambios en la altura del CMJ 3 minutos después de una sentadilla al 90% de 1 RM. Finalmente, Behm y col. [34] observaron ausencia de cambios en el pico de fuerza isométrica inmediatamente después de tres MVCs de 10 segundos; sin embargo, después de 10 a 15 minutos del período de recuperación, la fuerza máxima disminuyó (7–9%; p<0,05). Estos hallazgos contradictorios sugieren que la relación PAP-fatiga y sus efectos sobre actividades voluntarias subsiguientes son multifacéticos. En resumen, se ha sugerido que luego de una CC se requiere un tiempo de recuperación óptimo para disminuir la fatiga y aumentar la PAP; sin embargo, las evidencias son inconsistentes para respaldar esta teoría. Hay un número de posibles explicaciones para los resultados contrastantes producidos por los estudios antes mencionados. La relación entre PAP y fatiga, y los efectos en general de la historia contráctil en relación al rendimiento subsiguiente, está influenciada por una combinación de factores [2]. Estos incluyen: volumen de la CC (ej. series, repeticiones e intervalo de reposo entre numerosas series); intensidad de la CC (aunque hay consenso en que las contracciones de máxima intensidad optimizan la PAP), el tipo de CC realizada (ej. dinámica o isométrica); las características del sujeto (ej. fuerza muscular, distribución del tipo de fibras, nivel de entrenamiento o proporción de fuerza-potencia), y el tipo de actividad ejecutada después de la CC [1, 2]. La Figura 9 ilustra la integración de estos factores complejos y en la siguiente sección se discuten en más detalle.
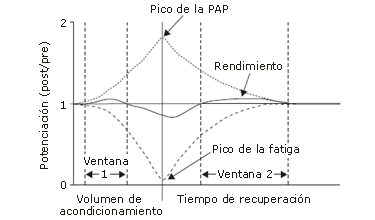
Figura 8. Modelo de la relación hipotética entre potenciación post-activación (PAP) y fatiga siguiendo un protocolo de contracción de preacondicionamiento (acondicionamiento) [3]. Cuando el volumen de acondicionamiento es bajo, la PAP es más dominante que la fatiga, e inmediatamente puede producirse una potenciación del rendimiento explosivo subsiguiente (post/pre) (ventana 1). A medida que el volumen de acondicionamiento aumenta, la fatiga se vuelve dominante, afectando negativamente al rendimiento subsiguiente. Luego del acondicionamiento, la fatiga se disipa a una tasa más rápida que la PAP, y la potenciación de un rendimiento explosivo subsiguiente puede elevarse en algún punto durante el período de recuperación (ventana 2).
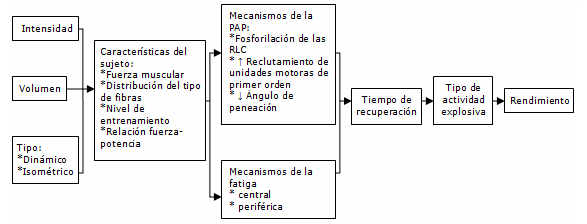
Figura 9. Factores complejos que influyen en el rendimiento de una actividad explosiva voluntaria luego de una contracción de acondicionamiento (acondicionamiento). La intensidad, volumen y tipo de acondicionamiento, afectarán a los individuos de manera diferente, dependiendo de las características de los sujetos. En conjunto, estos factores influirán en el grado hasta el cual son afectados los mecanismos de la potenciación post-activación (PAP) y la fatiga. La interacción entre los mecanismos de la PAP y la fatiga determinarán si la actividad subsiguiente es potenciada, y el período de recuperación requerido para alcanzar la potenciación. Sin embargo, sin tener en cuenta, las interacciones previas, la respuesta de algunas actividades explosivas al acondicionamiento puede ser diferente a la respuesta a otras actividades explosivas. RLC=cadenas livianas reguladoras.
VOLUMEN DE LA CONTRACCION DE ACONDICIONAMIENTO
El efecto del volumen de la CC sobre la interacción entre la PAP y la fatiga es resaltado en un estudio en particular. Hamada y col. [14] usaron un protocolo fatigante de 16 MVC isométricas de 5 segundos en un ejercicio de extensión de rodillas, con cada MVC separada por un intervalo de descanso de 3 segundos.
Una respuesta de contracción es simulada en el nervio femoral previo a las MVCs, entre cada MVC, 1 minuto después de las MVCs, y luego en cada segundo minuto después de las MVCs, por 13 minutos. El PT de la contracción aumentó gradualmente sobre las primeras tres MVCs, haciendo un pico de 127% de aumento sobre los valores de la línea de base (p<0,05). Esto demuestra que la PAP fue más dominante que la fatiga, después de los primeros tres MVCs cuando el volumen de MVC fue pequeño. Para el protocolo de fatiga restante, sin embargo, el PT de la contracción disminuyó progresivamente, en la MVC número 16 estuvo un 32% por debajo de los valores de condición inicial (p<0,05). Esto demuestra que así como el volumen de las MVC continúa aumentando, también lo hace la dominancia de la fatiga. Luego del protocolo de fatiga, el PT de la contracción aumentó progresivamente, y sobrepasó los valores de condición inicial después de 30 a 120 segundos de recuperación (+32%; p<0,05). Esto demuestra que la fatiga se disipó a una mayor tasa que la PAP y consecuentemente, hubo una potenciación en el PT de la contracción durante el período de recuperación. En la Figura 10 se presenta una adaptación de estos resultados. Estos hallazgos fueron respaldados en otros estudios [6]. Los autores registraron la tensión de la contracción en los dorsiflexores antes e inmediatamente después de la realización de protocolos de cinco MVC de dorsiflexión isométrica, donde cada protocolo se diferenció en la duración de las MVC (volumen). En consecuencia, cada protocolo indujo un nivel diferente de PAP, provocando la MVC isométrica de 10 segundos, la mayor potenciación (PT de la contracción: después de una MVC de 1 segundo=+43%; después de una MVC de 3 segundos=+130%; después de una MVC de 10 segundos =+142%; después de una MVC de 30 segundos=+65%; después de una MVC de 60 segundos=+14%). Nuevamente, la pregunta importante es: ¿ocurrirá o no un efecto similar durante la realización de actividades voluntarias explosivas?.
French y col. [50] midieron los efectos de CC de diferentes volúmenes sobre el rendimiento de actividades voluntarias explosivas subsiguientes. Los autores midieron un incremento significativo en el PT de la extensión de rodilla a isovelocidad inmediatamente después de 3 segundos de MVCs isométricas (+6,1%; p<0,05), pero repostaron una disminución significativa en el PT de la extensión de rodilla isocinética inmediatamente después de tres MVCs isométricas de 5 segundos (3%; p<0,05). En contraste, Behm y col. [34] midieron el pico de fuerza de la MVC isométrica después de una, dos y tres series de MVCs isométricas de 10 segundos, y el único efecto reportado fue una disminución del 8-9% de la fuerza pico 10 a 15 minutos después de las tres series de MVCs. Como se discutió en la sección 3, no se espera que la PAP aumente la fuerza pico isométrica (la cual representa la fuerza máxima), por lo que Behm y col. [34] podrían haber observado potenciación si hubieran medido la RDF voluntaria o el rendimiento dinámico. Asimismo, el volumen de CC más pequeño usado por Behm y col. [34] (MVC isométrica de 10 segundos) es probablemente más grande que el volumen más pequeño de CC usado por French y col. [50] (tres MVCs isométricas de 3 segundos separadas por 3 minutos), y puede haber inducido, por lo tanto, un mayor grado de fatiga. Es más, debido a las demás medidas diferentes tomadas por Behm y col. [34] durante el período de recuperación (incluyendo las contracciones tetánicas de alta frecuencia, las contracciones nerviosas, MVC isométricas al 30% y ITT), la fatiga pudo haber seguido acumulándose y reduciendo así, cualquier oportunidad para conseguir la PAP.
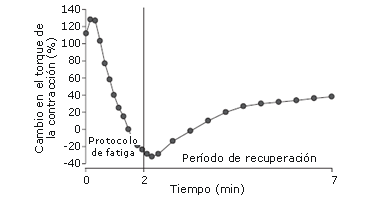
Figura 10. Transcurso del tiempo en el torque de la contracción del extensor de rodilla durante un protocolo de fatiga y a lo largo de un período de recuperación subsiguiente de 5 minutos. El protocolo de fatiga consistió en 16 MVCs de 5 segundos separadas por 3 segundos de recuperación. Se registró una contracción nerviosa antes del protocolo de fatiga, entre cada MVC, y 5 segundos después del protocolo de fatiga, y luego, cada 30 segundos durante el período de recuperación. El torque de la contracción es presentado como porcentaje de los valores previos al protocolo de fatiga [14].
Los resultados de los cuatro estudios antes mencionados [6, 14, 34, 50] demuestran la influencia del volumen de la CC sobre la relación PAP-fatiga.
También presentan la posibilidad de que la PAP se desarrolle más rápido que la fatiga y pueda, por lo tanto, ser utilizada inmediatamente después de un volumen de CC relativamente bajo (ventana 1 en la Figura 8). Por lo contrario, a medida que el volumen de la CC aumenta, también lo hace la fatiga y su dominancia en la relación PAP-fatiga, y por lo tanto se requiere un período de recuperación antes que la PAP se produzca (ventana 2 en la Figura 8). Aún debe determinarse el período de recuperación específico requerido para diferentes volúmenes de CC, y es difícil comparar los resultados de estudios individuales porque las metodologías no han sido estandarizadas. Si las investigaciones futuras intentan hacer inferencias sobre el tipo de calentamiento o protocolo de entrenamiento ideal que optimicen la PAP, el volumen de la CC y la recuperación entre la CC y la actividad subsiguiente deberían ser valorados juntos.
TIPO DE CONTRACCION DE PREACONDICIONAMIENTO
Aunque, en distintos grados, cualquier tipo de contracción es factible de activar los mecanismos de la PAP [4], el grado de potenciación alcanzada está probablemente relacionada al tipo de contracción. En consecuencia, el uso de diferentes tipos de CC ha contribuido posiblemente a la obtención de resultados inconsecuentes, los cuales ya han sido discutidos.
Como las investigaciones pasadas han usado normalmente tanto CC isométricas como dinámicas, este artículo solo discutirá las diferencias entre estos dos tipos de contracciones.
Varios estudios han investigado los efectos de las MVCs isométricas sobre las actividades explosivas subsiguientes, y mientras que dos estudios reportaron un incremento [10,50], otros reportaron ausencia de cambios en el rendimiento [11, 34, 57]. Estudios anteriores han usado también contracciones dinámicas voluntarias máximas o cercanas al máximo para inducir PAP, y nuevamente algunos registraron potenciación de la actividad explosiva subsiguiente [15, 45-48, 54, 58] y otros no [20, 49, 52, 53, 55].
Estos resultados conflictivos (ver Tabla 1 para los resultados) presentan una relación confusa entre el tipo de contracción (isométrica vs. dinámica) y la respuesta de la PAP, y solo un estudio (para nuestro conocimiento) ha comparado directamente la CC isométrica y dinámica con respecto a sus efectos sobre el rendimiento en las actividades explosivas subsiguientes [56]. Este estudio informó que mientras se observó un aumento significativo en la altura del CMJ (2,9%; p<0,01) y el pico de potencia (8,7%; p<0,001) 3 minutos después de tres MVC isométricas de 3 s de sentadilla, no se presentaron cambios en la altura del CMJ (p>0,05), pero se midió un aumento significativo en el pico de potencia del CMJ (8,0%; p< 0,001) 3 minutos después de una serie de 3 RM de sentadilla dinámica. Los autores concluyen que los acondicionamientos isométricos inducen una mayor respuesta de PAP que los dinámicos. Sin embargo, los dos tipos de acondicionamiento no fueron equiparados respecto al volumen y la frecuencia, y como resultado, es difícil hacer una comparación directa entre sus efectos.
Teóricamente, los diferentes tipos de contracción tendrían diferentes efectos sobre la fatiga neuromuscular [60, 61]. Babault y col. [60] midieron la fatiga neuromuscular durante un protocolo fatigante de contracción dinámica y un protocolo fatigante de contracción isométrica, donde ambos protocolos fueron equiparados en términos de la disminución del PT. Los autores reportaron que la fatiga temprana presentada durante el protocolo dinámico fue preferentemente de origen periférico (fatiga periférica definida como la disminución de la capacidad para generar fuerza debido al fallo en el potencial de acción, al fallo del acoplamiento excitación-contracción, o al daño en la secuencia de los puentes cruzados en presencia de conducción nerviosa incrementada o sin cambios [61]), mientras que la fatiga central (definida como una reducción en la conducción nerviosa al músculo [61]) se desarrolló hacia el final del protocolo fatigante dinámico. El protocolo isométrico, sin embargo, produjo el perfil opuesto, por el cual la fatiga fue en principio de origen central y luego periférico.
Babault y col. [60] propusieron que la diferencia en el desarrollo de la fatiga entre las contracciones isométricas y concéntricas podría estar asociada con la acumulación de metabolitos en el músculo, los cuales podrían activar y/o sensibilizar los grupos de fibras nerviosas aferentes de menor diámetro (III y IV) [60, 62, 63]. En orden regular, esto causaría fatiga central por la inhibición de la activación de la motoneurona a, y/o reducción de la conducción descendente supraespinal [60, 63] y/o disminución de la tasa de disparo de la motoneurona [64]. La naturaleza intermitente de las contracciones dinámicas podría favorecer el flujo sanguíneo, ayudando, por consiguiente, a la remoción de productos metabólicos. Por lo tanto, la acumulación metabólica sería mayor durante las contracciones isométricas, dando por resultado una mayor fatiga central. Por el contrario, se ha informado que la acumulación de lactato alivia la fatiga periférica [65].
Esto podría describir el desarrollo más lento de fatiga periférica durante las contracciones isométricas cuando son comparadas con las contracciones dinámicas [60].
Si las contracciones isométricas y dinámicas pueden inducir distintos tipos de respuesta a la fatiga, entonces es válido asumir que también tendrían distintos efectos sobre el mecanismo de la PAP. Por ejemplo, el movimiento excéntrico de las contracciones dinámicas (pero no de las contracciones isométricas) aumenta los disparos de los husos musculares, activando fibras nerviosas del grupo Ia [63]. Esto puede realzar las descargas nerviosas aferentes a la médula espinal. En consecuencia, la disminución en el fallo de transmisión de las fibras nerviosas Ia hacia las unidades motoras a adyacentes, que resulta en un aumento en la activación de unidades motoras de primer orden durante la actividad subsiguiente, podría ser mayor después de contracciones dinámicas. Por otro lado, las contracciones isométricas activan un mayor número de unidades motoras que las contracciones dinámicas [66]. En consecuencia, más fibras musculares podrían estar involucradas durante una contracción isométrica, y esto podría resultar en un mayor porcentaje de fosforilación de RLC, y mayores cambios en la arquitectura del músculo.
En resumen, la evidencia preliminar sugiere que las CC isométricas pueden inducir mayor fatiga central, pero probablemente activen en mayor medida los mecanismos periféricos de la PAP. En contraste, las CCs dinámicas pueden inducir mayor fatiga periférica, pero probablemente activen en mayor medida los mecanismos centrales de la PAP (Tabla 2). El modo en el que estos mecanismos interactúan no ha sido determinado aún, pero es razonable asumir que las contracciones isométricas y dinámicas tendrán efectos diferentes sobre las actividades explosivas subsiguientes. Las diferencias entre las contracciones isométricas y dinámicas van a influenciar también el volumen y período de recuperación requeridos para potenciar las actividad explosiva subsiguiente. Las investigaciones futuras deberían investigar los efectos del tipo de contracción sobre los mecanismos de la PAP y la fatiga, estandarizando, al mismo tiempo, el volumen y período de recuperación de la CC. Tampoco se sabe aún si una CC de cualquier tipo es más beneficiosa que los métodos convencionales de calentamiento [18] y aunque un estudio sugirió que es así [46] sus resultados fueron específicos para los individuos y protocolos valorados. Las investigaciones futuras deberían comparar los efectos potenciadores de la CC con las técnicas de calentamiento convencionales.
CARACTERISTICAS DE LOS SUJETOS
Ha sido sugerido que las características de los sujetos afectan la respuesta PAP-fatiga de un individuo, estas características incluyen a la fuerza muscular, la distribución del tipo de fibras, el nivel de entrenamiento y la relación fuerza-potencia. Estos factores son discutidos en más detalle en las secciones siguientes.
Fuerza Muscular
Hay evidencias que sugieren que la fuerza muscular de un individuo podría determinar en parte su respuesta a la PAP luego de una CC. Gourgoulis y col. [15] observaron un incremento del 4% en la altura del CMJ (p<0,05), luego de cinco series de sentadilla, en las cuales los sujetos pudieron cargar más de 160 kg. Por el contrario, aquellos sujetos incapaces de realizar la sentadilla con cargas >160 kg, solo registraron un incremento del 0,4% en la altura del CMJ (p>0,05). De forma similar, Kilduff y col. [54] reportaron una correlación entre la fuerza muscular (absoluta y relativa) y la potenciación del pico de potencia del CMJ, 12 minutos después de una serie de 3 RM de sentadilla (r=0,63; p<0,01). Una posible explicación para estos hallazgos podría asociarse a la distribución del tipo de fibras. La relación positiva y lineal entre fuerza muscular y porcentaje de fibras musculares tipo II está bien documentada (r=0,5-0,93; p<0,05),[67-69] y las fibras musculares tipo II muestran el mayor aumento en la fosforilación de las RLC luego de una CC [7].
Es más, los sujetos con un mayor porcentaje de fibras musculares tipo II presumiblemente tengan un mayor número de unidades motoras de alto orden en reserva, las cuales podrían ser activadas por medio de una disminución en la falla de transmisión, luego de una CC. El efecto combinado de una mayor fosforilación de las RLC y un mayor aumento en el reclutamiento de unidades motoras de alto orden predispondría teóricamente a los individuos con un mayor porcentaje de fibras musculares de tipo II a una mayor respuesta de la PAP. En consecuencia, podría especularse que los sujetos más fuertes en los dos estudios discutidos anteriormente [15, 54] tendrían un mayor porcentaje de fibras musculares de contracción rápida, y así alcanzaron una mayor respuesta de PAP.

Tabla 2. Ilustración de los efectos hipotéticos de las contracciones de preacondicionamiento isométricas o dinámicas sobre los mecanismos centrales y periféricos de la potenciación post-activación (PAP) y la fatiga.
Distribución del Tipo de Fibras
Hamada y col. [14] proporcionaron evidencias que apoyaban la relación entre la distribución del tipo de fibraS y la PAP. Los autores separaron sus sujetos en dos grupos: uno con predominancia de fibras musculares de contracción rápida (tipo II) (T-II; n=4), y el segundo, con predominancia de fibras musculares de contracción lenta (tipo I) (T-I; n=4).
Reportaron una mayor respuesta del PT en el grupo T-II durante una MVC isométrica de 3 segundos (250,0 vs. 171,0 N.m; p<0,01). Es más, en respuesta a un protocolo de fatiga de 16 MVCs isométricas de 5 segundos de extensores de rodilla, el grupo T-II mostró una potenciación en la tensión de la contracción significativamente mayor durante las primeras etapas del protocolo de fatiga (+127% vs. +40% de incremento en el PT después de la tercera MVC; p<0,05). Sin embargo, el grupo T-II también tubo una mayor disminución tanto en el PT de la contracción como en el PT de la MVC durante las últimas etapas del protocolo de fatiga (p<0,05). Por lo tanto, aunque los sujetos con un mayor porcentaje de fibras musculares tipo II provocaron una mayor respuesta de PAP, también provocaron una mayor respuesta de la fatiga luego de un protocolo de CC de alto volumen.
Hay un número de posibles razones por las cuales Hamada y col. [14] observaron una mayor respuesta de fatiga en el grupo T-II. Como se expresó, Hamada y col. [14] reportaron una mayor producción de PT en el grupo T-II durante las primeras etapas del protocolo de fatiga. Por lo tanto, de acuerdo con la relación fuerza-fatiga [70], se esperaría una mayor respuesta de fatiga en el grupo T-II. Además se ha reportado una correlación negativa entre la tasa glucolítica inicial y la fatiga durante el ejercicio intermitente [71]. La tarea específica empleada por Hamada y col. [14] (16 MVCs isométricas de 5 segundos con 3 segundos de reposo entre MVCs) dependería predominantemente de una alta tasa de producción anaeróbica de adenosín trifosfato (ATP), especialmente en sujetos con un gran porcentaje de fibras musculares tipo II [72, 73]. Por lo tanto, aunque se espera que los sujetos con un alto porcentaje de fibras musculares tipo II produzcan un mayor PT de la MVC, debido a que se produce una más alta tasa inicial de producción anaeróbica de ATP, también mostrarán probablemente disminuciones del PT mayores, debido a una mayor utilización de las reservas de energía anaeróbica y la producción de metabolitos asociados a la fatiga [74, 75].
Nivel de Entrenamiento
El nivel de entrenamiento de un individuo puede también influenciar las respuestas de la PAP y la fatiga luego de una CC. Chiu y col. [20] separaron una muestra de 24 sujetos en atletas que estaban entrenando y participando en un deporte a nivel nacional o internacional (RT; n=7), y en aquellos que participaban en entrenamiento de resistencia recreativo (UT; n=17). Cinco series de una sentadilla con el 90% de 1 RM y 5-7 minutos de recuperación subsiguiente indujeron un 1-3% de aumento en la altura del CMJ y SJ en el grupo RT. De manera contraria, el grupo UT reaccionó a al mismo acondicionamiento con una disminución de 1-4% en la altura del CMJ y SJ. Chiu y col. [20] sugirieron que aquellos sujetos que entrenaban a un nivel más alto de sobrecarga desarrollarían resistencia a la fatiga como una adaptación de sus regimenes de entrenamiento intenso y tendrían más posibilidades de producir PAP. Chiu y col. [20] sin embargo, no midieron la distribución de tipo de fibras, por lo que es posible que un mayor porcentaje de fibras musculares de contracción rápida en el grupo RT haya contribuido también a los efectos observados en este estudio.
Relación Fuerza-Potencia
Existe también evidencia que sugiere que la relación fuerza-potencia del sujeto influenciará su respuesta de PAP a una CC. Schneiker y col. [76] reportaron una correlación negativa significativa entre la relación fuerza-potencia y la potenciación del pico de potencia durante un CMJ con carga, ejecutado 2-4 minutos después de una serie de 6 RM de sentadilla (r2=0,65; p<0,05). Es más, cuando la muestra de sujetos entrenados en fuerza fue separada en sujetos con una relación fuerza-potencia de <19 W.kg-1 (grupo 1), y sujetos con una relación fuerza-potencia de >19 W.kg -1 (grupo 2), el grupo 1 tuvo una correlación negativa significativa entre la relación fuerza-potencia y la potenciación del pico de potencia (r2=0,91; p<0,05). En contraste, el grupo 2 mostró ausencia de relación entre la relación fuerza-potencia y la potenciación del pico de potencia (p>0,05). Estos resultados sugieren que aquellos sujetos menos capaces para convertir efectivamente su fuerza en potencia probablemente se beneficiarán más de la PAP que aquellos sujetos capaces de hacerlo. Además parece que puede haber una relación fuerza-potencia umbral sobre la cual los sujetos no se benefician de la PAP.
En resumen, se ha sugerido que diferentes características de los sujetos afectan la respuesta PAP-fatiga de un individuo, y esto puede explicar en parte las contradicciones de las investigaciones del pasado. Las evidencias sugieren que los individuos que posiblemente obtengan mayores beneficios de la PAP son aquellos con una mayor fuerza muscular, con un mayor porcentaje de fibras tipo II (auque la fatiga puede ser mayor también en estos individuos), un alto nivel de entrenamiento de la resistencia, y una menor relación fuerza-potencia. Sin embargo son necesarios estudios adicionales para validar estos hallazgos, así como también para determinar los posibles efectos de otras características de los sujetos como la longitud de músculos y/o palancas. Para que los entrenadores consideren la implementación de CC previa a actividades explosivas (en entrenamiento o rendimiento) puede ser pertinente medir primero la susceptibilidad de cada atleta a la PAP durante el período fuera de temporada.
TIPO DE ACTIVIDAD SUBSIGUIENTE
Una explicación adicional para los resultados contradictorios de las investigaciones del pasado se basa en los diferentes tipos de actividades explosivas subsiguientes usadas para determinar los efectos agudos de la PAP. Los tipos de actividades subsiguientes empleados por estudios anteriores han incluido las MVCs isométricas [10, 34, 51] contracciones dinámicas aisladas (por ej., extensiones de rodilla a isovelocidad) [11, 47, 50] y actividades balísticas compuestas (ej. CMJ y DJ) [10, 15, 46, 49, 52-58]. Es posible que una CC específica no tenga el mismo efecto en diferentes actividades explosivas.
Con respecto a las diferencias entre las contracciones explosivas isométricas o dinámicas, estudios anteriores han reportado correlaciones moderadas a fuertes entre la RDF isométrica y dinámica (r=0,65-0,75) [77] y correlaciones moderadas a fuertes entre el pico de fuerza isométrico y dinámico (r=0,66–0,77) [77,78]. Estos resultados indican una clara relación entre los tests que miden la fuerza y potencia isométrica y dinámica. Hay, sin embargo. Una serie de diferencias en los procesos nerviosos y mecánicos involucrados en las actividades isométricas y dinámicas. Por ejemplo, el reclutamiento de unidades motoras y la frecuencia de disparo para una contracción isométrica, estará probablemente regulado por el principio de tamaño [79], en el cual las unidades motoras son reclutadas en un orden jerárquico de unidades de orden bajo a más alto. Por otro lado, las contracciones dinámicas podrían mostrar un patrón específico de reclutamiento de unidades motoras pertinente al ángulo de la articulación y posición durante el intervalo de movimiento [80]. Además, el movimiento excéntrico involucrado en las contracciones dinámicas, pero no en las contracciones isométricas, podría resultar en una mayor entrada aferente (grupo de fibras nerviosas Ia) desde los husos musculares [61, 81]. Como resultado, la respuesta de activación de la motoneurona a para las contracciones isométricas y dinámicas sería diferente. [82] Es más, la utilización de la energía elástica (ciclo estiramiento-acortamiento), acumulada en el músculo durante una contracción excéntrica, brinda una contribución importante para el rendimiento total de los movimientos dinámicos [83, 85]. El ciclo estiramiento-acortamiento, sin embargo, no se utiliza durante una contracción isométrica y en consecuencia, las contracciones isométricas pueden no reflejar las capacidades musculares para las situaciones dinámicas [82]. Por último, la PAP es mayor mientras el músculo está acortado [86] y se extiende a mayores frecuencias de estimulación en las contracciones concéntricas en comparación con las isométricas [22]. Esto sugiere que la PAP puede tener un efecto de mejora del rendimiento más allá de lo que podría esperarse en las contracciones isométricas.
Es también probable que mientras una CC específica pueda mejorar el rendimiento de una actividad dinámica particular, pueda disminuir o no tener efectos sobre el rendimiento de una actividad dinámica diferente. French y col. [50] analizaron la extensión de rodilla a isovelocidad, CMJ, DJ, y el rendimiento de 5 segundos de esprinten bicicleta, antes e inmediatamente después de tres extensiones de rodilla de 3 segundos. Los autores reportaron mejoras significativas en la altura en el DJ, RFD del DJ, y PT de la extensión de rodilla (+5,0%, +9,5% y +6,1%, respectivamente; p<0,05) después de las MVCs, pero hallaron efectos no significativos en cualquiera de las otras actividades (p>0,05). French y col.[50] usaron el análisis del tiempo de movimiento para explicar sus resultados. Reportaron que el DJ y la MVC de extensión de rodilla tuvieron un período de activación muscular de =0,25 segundos. Por el contrario, el CMJ y el esprint de 5 s en bicicleta tuvieron un periodo de activación muscular de =0,25 segundos. Las acciones musculares explosivas fueron definidas previamente como aquellas que tenían un periodo de activación =0,25 segundos [77]. French y col. [50] concluyeron, por lo tanto, que la PAP fue solamente efectiva en tareas definidas como acciones musculares explosivas. Sin embargo, las conclusiones de French y col. [50], deben interpretarse con precaución. Algunos estudios han registrado un efecto de potenciación en el rendimiento del CMJ, como en otras actividades que de lo contrario pueden no caer bajo la definición de acción muscular explosiva [10, 15, 46, 51, 54, 56, 58]. Además French y col. [50] solo midieron ejercicios realizados inmediatamente después de la CC, y puede haberse necesitado un intervalo de descanso para que se produzca un efecto de potenciación. Por último, el ejercicio de CC fue un ejercicio aislado que apuntó solo a los extensores de rodilla. El DJ puede cargar los extensores de rodilla hasta un punto mayor que el CMJ y el esprint de 5 segundos en bicicleta, lo que explicaría el aumento en la altura del DJ y RFD. Sin embargo, el CMJ y el esprint de 5 segundos en bicicleta pueden depender de la contribución de otros grupos musculares grandes distintos, lo cual debido a la cinética de la CC, no habían sido potenciados. Por lo tanto, estos resultados resaltan la importancia de equiparar las condiciones de cinética de las CC con las actividades explosivas subsiguientes. Haciendo esto, un individuo activa más probablemente las unidades motoras de primer orden, produce fosforilación en las RLC y cambia la arquitectura de aquellas fibras musculares específicamente asociadas con la actividad subsiguiente.
El propósito de la siguiente investigación ha sido establecer la aplicación de la PAP para actividades deportivas explosivas específicas. Las actividades deportivas explosivas son dinámicas por naturaleza así que, por las razones discutidas anteriormente, la respuesta isométrica a una CC no debería usarse para inferir efectos de la misma CC sobre las actividades deportivas subsiguientes. Si los investigadores están investigando la aplicación de la PAP al escenario del entrenamiento, los efectos reportados de una CC sobre actividades balísticas subsiguientes (ej. CMJ y DJ) puede ser útil, ya que los ejercicios balísticos son usados en los programas de entrenamiento de la potencia. Por otro lado, mientras que la PAP puede algunas veces ser efectiva para mejorar el rendimiento de un ejercicio balístico, puede no tener el mismo efecto ergogénico sobre el rendimiento de una actividad deportiva explosiva específica (por ej., carrera de velocidad, salto en largo). Si la PAP va a ser utilizada en competición, las investigaciones deben determinar primero sus efectos más allá de aquellos reportados para los ejercicios balísticos de entrenamiento. Dos estudios recientes han mostrado que la PAP puede mejorar el rendimiento de una actividad deportiva explosiva específica, reportando una disminución en el tiempo de esprint (-3% en 10 m [48] -2% en 30 m [48] y -3% en 40 m [45], p<0,05) 4 a 5 minutos después de la ejecución cercana a la máxima (>80% 1RM) de sentadilla. No obstante, se requieren estudios adicionales para establecer la aplicación de la PAP a muchas actividades deportivas explosivas diferentes. Es más, aún si la PAP muestra consecuentemente que realza el rendimiento de diferentes actividades deportivas explosivas, serán necesarias varias sugerencias prácticas para aplicar efectivamente la PAP en un escenario competitivo (como la necesidad de un posible equipamiento para el área de calentamiento y los requerimientos para realizar dentro de un período de recuperación optimo la posterior activación).
Como resultado de estas cuestiones poco prácticas, la aplicación de la PAP en el rendimiento ha sido cuestionada [18], pero con los aumentos en el rendimiento reportados >3%, las investigaciones adicionales están justificada.
CONCLUSION
La utilización de la PAP es posible para realzar la potencia mecánica y por lo tanto el rendimiento y /o el estímulo de entrenamiento de una actividad deportiva explosiva. Las evidencias sobre la aplicación práctica de la PAP sobre las actividades explosivas están aún inconclusas. Los resultados inconsistentes de las investigaciones parecen deberse a la compleja interacción de los diferentes factores que determinan el grado hasta el cual son afectados los diferentes mecanismos de la PAP y de la fatiga.
Se espera que mayores volúmenes e intensidades de CC induzcan mayores niveles tanto de PAP como de fatiga. Sin embargo, las tasas en la cual la PAP y la fatiga se desarrollan y disipan pueden diferenciarse, dando por resultado dos períodos de oportunidades para potenciar el rendimiento; inmediatamente después de una CC de bajo volumen, o después de un período de recuperación específico luego de una CC de alto volumen. El tipo de CC puede tener diferentes efectos sobre los mecanismos de la PAP y la fatiga.
Por ejemplo, las MVCs isométricas pueden inducir fatiga central, pero PAP periférica, mientras que las MVCs dinámicas pueden inducir la respuesta contraria. La interacción de estos mecanismos diferent
Referencias
1. Hodgson M., Docherty D., Robbins D (2005). Post-activation potentiation: underlying physiology and implications for motor performance. Sports Med 35 (7): 585-95
2. Robbins D. W (2005). Postactivation potentiation and its practical applicability: a brief review. J Strength Cond Res May; 19 (2): 453-8
3. Sale D. G (2002). Postactivation potentiation: role in human performance. Exerc Sport Sci Rev Jul; 30 (3): 138-43
4. Sale D. G (2004). Postactivation potentiation: role in performance. Br J Sports Med Aug; 38 (4): 386-7
5. Manning D. R., Stull J. T (1982). Myosin light chain phosphorylationdephosphorylation in mammalian skeletal muscle. Am J Physiol Mar; 242 (3): C234-41
6. Vandervoort A. A., Quinlan J., McComas A. J (1983). Twitch potentiation after voluntary contraction. Exp Neurol Jul; 81 (1): 141-52
7. Moore R. L., Stull J. T (1984). Myosin light chain phosphorylation in fast and slow skeletal muscles in situ. Am J Physiol Nov; 247 (5 Pt 1): C462-71
8. Stuart D. S., Lingley M. D., Grange R. W., et al (1988). Myosin light chain phosphorylation and contractile performance of human skeletal muscle. Can J Physiol Pharmacol Jan; 66 (1): 49-54
9. Vandenboom R., Grange R. W., Houston M. E (1993). Threshold for force potentiation associated with skeletal myosin phosphorylation. Am J Physiol Dec; 265 (6 Pt 1): C1456-62
10. Gullich A., Schmidtbleicher D (1996). MVC-induced short-term potentiation of explosive force. New Studies in Athletics 11 (4): 67-81
11. Gossen E. R., Sale D. G (2000). Effect of postactivation potentiation on dynamic knee extension performance. Eur J Appl Physiol Dec; 83 (6): 524-30
12. Hamada T., Sale D. G., MacDougall J. D., et al (2000). Postactivation potentiation, fiber type, and twitch contraction time in human knee extensor muscles. J Appl Physiol Jun; 88 (6): 2131-7
13. Szczesna D., Zhao J., Jones M., et al (2002). Phosphorylation of the regulatory light chains of myosin affects Ca2+ sensitivity of skeletal muscle contraction. J Appl Physiol Apr; 92 (4): 1661-70
14. Hamada T., Sale D. G., MacDougall J. D., et al (2003). Interaction of fibre type, potentiation and fatigue in human knee extensor muscles. Acta Physiol Scand 178 (2): 165-73
15. Gourgoulis V., Aggeloussis N., Kasimatis P., et al (2003). Effect of a submaximal half-squats warm-up program on vertical jumping ability. J Strength Cond Res May; 17 (2): 342-4
16. Grange R. W., Cory C. R., Vandenboom R., et al (1995). Myosin phosphorylation augments force- displacement and forcevelocity relationships of mouse fast muscle. Am J Physiol Sep; 269 (3 Pt 1): C713-24
17. Grange R. W., Vandenboom R., Xeni J., et al (1998). Potentiation of in vitro concentric work in mouse fast muscle. J Appl Physiol Jan; 84 (1): 236-43
18. Docherty D., Hodgson M (2007). The application of postactivation potentiation to elite sport. Int J Sports Physiol Perf 2 (4): 439-44
19. Baudry S., Duchateau J (2007). Postactivation potentiation in a human muscle: effect on the rate of torque development of tetanic and voluntary isometric contractions. J Appl Physiol Apr; 102 (4): 1394-401
20. Chiu L. Z., Fry A. C., Weiss L. W., et al (2003). Postactivation potentiation response in athletic and recreationally trained individuals. J Strength Cond Res Nov; 17 (4): 671-7
21. Szczesna D (2003). Regulatory light chains of striated muscle myosin. Structure, function and malfunction. Curr Drug Targets Cardiovasc Haematol Disord Jun; 3 (2): 187-97
22. Abbate F., Sargeant A. J., Verdijk P. W., et al (2000). Effects of highfrequency initial pulses and posttetanic potentiation on power output of skeletal muscle. J Appl Physiol Jan; 88 (1): 35-40
23. Baudry S., Klass M., Duchateau J (2008). Postactivation potentiation of short tetanic contractions is differently influenced by stimulation frequency in young and elderly adults. Eur J Appl Physiol 103 (4): 449-59
24. Smith J. C., Fry A. C (2007). Effects of a ten-second maximum voluntary contraction on regulatory myosin light-chain phosphorylation and dynamic performance measures. J Strength Cond Res Feb; 21 (1): 73-6
25. Gossard J. P., Floeter M. K., Kawai Y., et al (1994). Fluctuations of excitability in the monosynaptic reflex pathway to lumbar motoneurons in the cat. J Neurophysiol Sep; 72 (3): 1227-39
26. Luscher H. R., Ruenzel P., Henneman E (1983). Composite EPSPs in motoneurons of different sizes before and during PTP: implications for transmission failure and its relief in Ia projections. J Neurophysiol Jan; 49 (1): 269-89
27. Hirst G. D. S., Redman S. J., Wong K (1981). Post-tetanic potentiation and facilitation of synaptic potentials evoked in cat spinal motoneurons. J Physiol 321: 97-109
28. Enoka R (2002). Neuromechanics of human movement. 3rd ed. Champaign (IL): Human Kinetics
29. Trimble M. H., Harp S. S (1998). Postexercise potentiation of the H-reflex in humans. Med Sci Sports Exerc Jun; 30 (6): 933-41
30. Maffiuletti N. A., Martin A., Babault N., et al (2001). Electrical and mechanical H (max)-to-M(max) ratio in power- and endurance-trained athletes. J Appl Physiol Jan; 90 (1): 3-9
31. Folland J. P., Wakamatsu T., Finland M. S (2008). The influence of maximal isometric activity on twitch and H-reflex potentiation, and quadriceps femoris performance. Eur J Appl Physiol 104 (4): 739-48
32. Folland J. P., Williams A. G (2007). Methodological issues with the interpolated twitch technique. J Electromyogr Kinesiol Jun; 17 (3): 317-27
33. Shield A., Zhou S (2004). Assessing voluntary muscle activation with the twitch interpolation technique. Sports Med 34 (4): 253-67
34. Behm D. G., Button D. C., Barbour G., et al (2004). Conflicting effects of fatigue and potentiation on voluntary force. J Strength Cond Res May; 18 (2): 365-72
35. Folland J. P., Williams A. G (2007). The adaptations to strength training: morphological and neurological contributions to increased strength. Sports Med 37 (2): 145-68
36. Fukunaga T., Ichinose Y., Ito M., et al (1997). Determination of fascicle length and pennation in a contracting human muscle in vivo. J Appl Physiol Jan; 82 (1): 354-8
37. Mahlfeld K., Franke J., Awiszus F (2004). Postcontraction changes of muscle architecture in human quadriceps muscle. Muscle Nerve Apr; 29 (4): 597-600
38. Kubo K., Kanehisa H., Kawakami Y., et al (2001). Effects of repeated muscle contractions on the tendon structures in humans. Eur J Appl Physiol Jan-Feb; 84 (1-2): 162-6
39. Newton R. U., Kraemer W. J (1994). Developing explosive muscular power: implications for a mixed methods training strategy. Natl Strength Cond Assoc J 16 (5): 20-9
40. Baker D., Nance S (1999). The relation between strength and power in professional rugby league players. J Strength Cond Res 13 (3): 224-9
41. Potteiger J. A., Lockwood R. H., Haub M. D., et al (1999). Muscle power and fibre characteristics following 8 weeks of plyometric training. J Strength Cond Res 13 (3): 275-9
42. Stone M. H (1993). Literature review: explosive exercises and training. Natl Strength Cond Assoc J 15 (3): 6-19
43. Rahimi R (2007). The acute effect of heavy versus light-load squats on sprint performance. Phy Educ Sport 5 (2): 163-9
44. Saez Saez de Villarreal E., Gonzalez-Badillo J. J., Izquierdo M (2007). Optimal warm-up stimuli of muscle activation to enhance short and long-term acute jumping performance. Eur J Appl Physiol Jul; 100 (4): 393-401
45. Batista M. A., Ugrinowitsch C., Roschel H., et al (2007). Intermittent exercise as a conditioning activity to induce postactivation potentiation. J Strength Cond Res Aug; 21 (3): 837-40
46. Chatzopoulos D. E., Michailidis C. J., Giannakos A. K., et al (2007). Postactivation potentiation effects after heavy resistance exercise on running speed. J Strength Cond Res Nov; 21 (4): 1278-81
47. Ebben W. P., Jenson R. L., Blackard D. O (2000). Electromyographic and kinetic analysis of complex training variables. J Strength Cond Res 14 (4): 451-6
48. French D. N., Kraemer W. J., Cooke C. B (2003). Changes in dynamic exercise performance following a sequence of preconditioning isometric muscle actions. J Strength Cond Res Nov; 17 (4): 678-85
49. Gilbert G., Lees A., Graham-Smith P (2001). Temporal profile of post-tetanic potentiation of muscle force characteristics after repeated maximal exercise. J Sports Sci 19: 6
50. Hanson E. D., Leigh S., Mynark R. G (2007). Acute effects of heavyand light-load squat exercise on the kinetic measures of vertical jumping. J Strength Cond Res Nov; 21 (4): 1012-7
51. Jensen R. L., Ebben W. P (2003). Kinetic analysis of complex training rest interval effect on vertical jump performance. J Strength Cond Res May; 17 (2): 345-9
52. Kilduff L. P., Bevan H. R., Kingsley M. I., et al (2007). Postactivation potentiation in professional rugby players: optimal recovery. J Strength Cond Res Nov; 21 (4): 1134-8
53. Mangus B. C., Takahashi M., Mercer J. A., et al (2006). Investigation of vertical jump performance after completing heavy squat exercises. J Strength Cond Res Aug; 20 (3): 597-600
54. Rixon K. P., Lamont H. S., Bemben M. G (2007). Influence of type of muscle contraction, gender, and lifting experience on postactivation potentiation performance. J Strength Cond Res May; 21 (2): 500-5
55. Robbins D. W., Docherty D (2005). Effect of loading on enhancement of power performance over three consecutive trials. J Strength Cond Res Nov; 19 (4): 898-902
56. Young W. B., Jenner A., Griffiths K (1998). Acute enhancement of power performance from heavy load squats. J Strength Cond Res 12 (2): 82-4
57. Baudry S., Duchateau J (2004). Postactivation potentiation in human muscle is not related to the type of maximal conditioning contraction. Muscle Nerve Sep; 30 (3): 328-36
58. Babault N, Desbrosses K, Fabre MS, et al (2006). Neuromuscular fatigue development during maximal concentric and isometric knee extensions. J Appl Physiol Mar; 100 (3): 780-5
59. Kay D., St Clair Gibson A., Mitchell M. J., et al (2000). Different neuromuscular recruitment patterns during eccentric, concentric and isometric contractions. J Electromyogr Kinesiol Dec; 10 (6): 425-
60. Gandevia S. C (2001). Spinal and supraspinal factors in human muscle fatigue. Physiol Rev Oct; 81 (4): 1725-89
61. Taylor J. L., Butler J. E., Gandevia S. C (2000). Changes in muscle afferents, motoneurons and motor drive during muscle fatigue. Eur J Appl Physiol Oct; 83 (2-3): 106-15
62. Linnamo V., Hakkinen K., Komi P. V (1998). Neuromuscular fatigue and recovery in maximal compared to explosive strength loading. Eur J Appl Physiol Occup Physiol 77 (1-2): 176-81
63. Karelis A. D., Marcil M., Peronnet F., et al (2004). Effect of lactate Infusion on M-wave characteristics and force in the rat plantaris muscle during repeated stimulation in situ. J Appl Physiol Jun; 96 (6): 2133-8
64. Duchateau J., Hainaut K (1984). Isometric or dynamic training: differential effects on mechanical properties of a human muscle. J Appl Physiol Feb; 56 (2): 296-301
65. Thorstensson A., Grimby G., Karlsson J (1976). Force-velocity relations and fibre composition in human knee extensor muscles. J Appl Physiol 40 (1): 12-6
66. Maughan R. J., Watson J. S., Weir J (1983). Relationships between muscle strength and muscle cross-sectional area in male sprinters and endurance runners. Eur J Appl Physiol Occup Physiol 50 (3): 309-18
67. Aagaard P., Andersen J. L (1998). Correlation between contractile strength and myosin heavy chain isoform composition in human skeletal muscle. Med Sci Sports Exerc Aug; 30 (8): 1217-22
68. Enoka R. M., Stuart D. G (1992). Neurobiology of muscle fatigue. J Appl Physiol May; 72 (5): 1631-48
69. Gaitanos G. C., Williams C., Boobis L. H., et al (1993). Human muscle metabolism during intermittent maximal exercise. J Appl Physiol Aug; 75 (2): 712-9
70. Katz A., Sahlin K., Henriksson J (1986). Muscle ATP turnover rate during isometric contraction in humans. J Appl Physiol Jun; 60 (6): 1839-42
71. Greenhaff P. L., Nevill M. E., Soderlund K., et al (1994). The metabolic responses of human type I and II muscle fibres during maximal treadmill sprinting. J Physiol Jul 1; 478 (Pt 1): 149-55
72. Fabiato A., Fabiato F (1978). Effects of pH on the myofilaments and the sarcoplasmic reticulum of skinned cells from cardiace and skeletal muscles. J Physiol March 1; 276 (1): 233-55
73. Chasiotis D., Hultman E., Sahlin K (1983). Acidotic depression of cyclic AMP accumulation and phosphorylase b to a transformation in skeletal muscle of man. J Physiol Feb 1; 335 (1): 197-204
74. Schneiker K., Billaut F., Bishop D (2006). The effects of preloading using heavy resistance exercise on acute power output during lower-body complex training [abstract]. Book of Abstracts of the 11th Annual Congress, European College of Sports Science, Jul 5-8, Lausanne, 89
75. Blazevich A. J., Gill N., Newton R. U (2002). Reliability and validity of two isometric squat tests. J Strength Cond Res May; 16 (2): 298-304
76. Henneman E., Somjen G., Carpenter D. O (1965). Functional significance of cell size in spinal motoneurons. J Nneurophysiol May; 28: 560-80
77. Ter Haar Romeny B. M., Denier Van der Gon J. J., Gielen C. C (1982). Changes in recruitment order of motor units in the human biceps muscle. Exp Neurol Nov; 78 (2): 360-8
78. McComas A. J (1996). Skeletal muscle: form and function. Champaign (IL): Human Kinetics
79. Baker D., Wilson G., Carlyon B (1994). Generality versus specificity: a comparison of dynamic and isometric measures of strength and speed-strength. Eur J Appl Physiol Occup Physiol; 68 (4): 350-5
80. Newton R. U., Murphy A. J., Humphries B. J., et al (1997). Influence of load and stretch shortening cycle on the kinematics, kinetics and muscle activation that occurs during explosive upper-body movements. Eur J Appl Physiol Occup Physiol; 75 (4): 333-42
81. Babault N., Maffiuletti N., Pousson M (2008). Postactivation potentiation in human knee extensors during dynamic passive movements. Med Sci Sports Exerc; 40 (4): 735-43
82. Wilson G. J., Elliot B. C., Wood G. A (1991). The effect on performance of imposing a delay during a stretch-shorten cycle movement. Med Sci Sports Exerc 23 (3): 364-70
83. Walshe A. D., Wilson G. J., Ettema G. J (1998). Stretch-shorten cycle compared with isometric preload: contributions to enhanced muscular performance. J Appl Physiol Jan; 84 (1): 97-106