
Frecuencia de Zancada Preferida y Óptima, Rigidez y Economía: Cambios Asociados a la Fatiga Durante Una Carrera de Alta Intensidad de 1 Hora
Preferred and Optimal Stride Frequency, Stiffness and Economy: Changes with Fatigue During a 1-H High-Intensity Run
Iain Hunter1 y Gerald A. Smith2
1Department of Exercise Sciences, Brigham Young University, Provo, UT, USA
2Department of Physical Performance, Norwegian School of Sport Sciences, PO Box 4014, Ullevaal Stadion, 0806 Oslo, Norway
Artículo publicado en el journal Revista de Entrenamiento Deportivo, Volumen 33, Número 1 del año 2019.
Publicado 29 de abril de 2019
Resumen
Palabras clave: Frecuencia de zancada, economía de la carrera, optimización, rigidez
Abstract
Keywords: Stride frequency, Running economy, Optimization, Stiffness
INTRODUCCIÓN
La economía de la carrera es una característica que probablemente afecta el rendimiento en carreras que van desde medio fondo hasta ultra maratones. Se sabe que los patrones cinemáticos influyen en el costo metabólico de un corredor a una determinada velocidad de carrera, donde la longitud y la frecuencia de la zancada son dos de las principales características que afectan claramente la economía. Los cambios en la longitud y la frecuencia de la zancada que se alejan de los valores óptimos producen un aumento en el costo metabólico o una economía más pobre. La mayoría de los corredores eligen naturalmente una combinación longitud-frecuencia de zancada, que les permita minimizar el costo metabólico (Cavanagh y Williams, 1982). Esta combinación preferida de frecuencia y longitud de zancada da como resultado lo que se podría denominar auto-optimización.
La fatiga durante las carreras es una respuesta multidimensional que modifica las características fisiológicas y biomecánicas para producir cambios con respecto a las condiciones iniciales (Maclaren et al., 1989). Los cambios internos que se producen con la fatiga pueden trasladarse a cambios cinemáticos y cinéticos externos durante el transcurso de una carrera prolongada, que afectarán las características de la zancada a una velocidad de carrera dada y contribuirán con disminución de la economía de carrera. La observación de las respuestas de los corredores frente a la fatiga ha permitido detectar diversos cambios en las características cinemáticas, musculo-esqueléticas y fisiológicas después de un ejercicio de alta intensidad y/o de larga duración. Por ejemplo, en un recorrido relativamente rápido durante aproximadamente 12 minutos hasta el agotamiento voluntario, el consumo de oxígeno aumentó 5% (Candau et al., 1998). Durante una carrera de1 h inicialmente al 80% de VO2max, Sproule (1998) observó que la economía de la carrera se vio negativamente afectada con un aumento de aproximadamente el 9% en el VO2. En maratones, el costo metabólico aumentó más del 15% durante el transcurso de una carrera (Kyröläinen et al., 2000). En otro estudio se observó que la carrera en cinta rodante de baja intensidad, que inicialmente tenía un VO2max de 55% incrementó el costo en 18% después de 5 h de carrera (Place et al., 2004). Por el contrario, después de un ultramaratón con más de 6 h de carrera, Millet et al. (2000) no observaron cambios en el costo de carrera. Se han observado cambios cinemáticos asociados con la fatiga bajo algunas condiciones de carrera, pero no en todas las condiciones. Por ejemplo, la frecuencia de zancada disminuyó con la fatiga en algunos estudios (Candau y otros, 1998, Dutto y Smith, 2002, Siler y Martin, 1991), pero en otros estudios se observaron aumentos en la frecuencia de zancada (Kyröläinen et al. et al., 2004). En condiciones de fatiga se han observado otros cambios como el posicionamiento angular de la rodilla y del tobillo (Derrick et al., 2002). Estas diferentes, y a veces contradictorias, observaciones sobre las respuestas de los corredores quizás se deban a verdaderas diferencias asociadas a la fatiga entre las carreras de corta duración, de alta intensidad y las carreras de larga duración y menor intensidad, como las maratones o ultra maratones. Además se han observado considerables diferencias entre los individuos (Dutto y Smith 2002, Siler y Martin 1991), ya que algunos corredores son sensibles a la fatiga, mientras que otros mantienen características fisiológicas y mecánicas casi constantes.
Frecuentemente la carrera ha sido descripta como un patrón de movimiento con salto en el cual la pierna que corre se comporta como un resorte lineal. El modelo de masa-resorte introducido por McMahon y Cheng (1990) consiste en una adaptación relativamente simple de las características de la pierna que corre en función de la fuerza de reacción del suelo y del desplazamiento del centro de masa. A partir de esto, se puede determinar la rigidez de la pierna durante el posicionamiento como una característica global resultante del conjunto de rigidez muscular, tendinosa y articular. A pesar de su relativa simplicidad, el modelo ha permitido tener una mejor comprensión de la mecánica de la carrera, incluida la relación entre las características de rigidez y la frecuencia de zancada (Farley y González, 1996). Al igual que un resorte lineal simple en oscilación, la rigidez y la frecuencia aumentan proporcionalmente en los corredores. Sin embargo, la frecuencia de resonancia del sistema de resorte formado por la pierna (determinada por la rigidez y la masa) puede tener algunas diferencias con la frecuencia de la zancada con la que funciona una pierna. Existe cierta evidencia de que las grandes diferencias entre la frecuencia de resonancia y la de zancada provocan un mayor costo de carrera (Cavagna et al., 1997, Dalleau et al., 1998). Estas relaciones son de particular interés para el estudio actual que implica fatiga, porque se piensa que tanto las articulaciones como los tendones tienen una menor rigidez después del ejercicio fatigante (Kuitunen et al 2002, Kubo et al., 2001). Las activaciones excéntricas repetidas de los músculos que se producen durante la carrera provocan una reducción de la capacidad de estiramiento muscular y un cierto retraso en el ciclo de estiramiento-acortamiento que puede provocar cambios en la mecánica de la zancada que impliquen una mayor flexión de la rodilla (Noakes 2000). El almacenamiento elástico y el retorno de la energía, que es un componente integral del ciclo de estiramiento-acortamiento durante las carreras está directamente relacionado con la rigidez de la pierna que corre. Heise y Martin (1998) y Dalleau et al. (1998) observaron una relación inversa entre la rigidez y el costo de la carrera. Así, los cambios en la rigidez asociados con la fatiga pueden ayudar a explicar el aumento de los costos metabólicos que se observan típicamente durante las carreras de larga duración.
Este estudio reúne diversas perspectivas de la carrera como la fatiga, la cinemática y el modelado para ayudar a explicar el control de la pierna que corre. Cuando no están fatigados, la mayoría de los corredores eligen una frecuencia de zancada que coincide estrechamente con la frecuencia que minimiza el costo metabólico (Cavanagh y Williams 1982); esta es la frecuencia óptima. Sobre la base del modelo masa-resorte, suponemos que esto se logra mediante el ajuste de la rigidez que permite lograr una activación muscular óptima y esto produce una frecuencia de zancada óptima. Cuando están fatigados, muchos corredores desplazan la frecuencia de la zancada lejos de su frecuencia óptima en ausencia de fatiga y, además, experimentan un aumento del costo metabólico. No se conoce con detalle cuales son las conexiones entre estas respuestas. Esta investigación analizará estas relaciones a través de la evaluación de las estrategias de frecuencia y la rigidez de los corredores inicialmente y al final de una carrera de 1 h de alta intensidad a velocidad constante. Los costos metabólicos medidos a través de una serie de manejos de frecuencia de zancada permitieron la determinación de la frecuencia de zancada óptima. A través de este proceso se abordaron varias preguntas de investigación: ¿Se producen cambios en el costo metabólico, en la frecuencia de zancada preferida y en la rigidez durante el transcurso de una carrera de una hora? ¿Se optimizan las frecuencias de zancada de los corredores para minimizar el costo metabólico a pesar de la fatiga al final de una carrera de una hora? Sobre la base de numerosos estudios previos, es esperable que el costo metabólico aumente durante la carrera. Se planteó la hipotesis que la frecuencia de la zancada disminuiría con la fatiga (Candau et al 1998, Dutto y Smith 2002, Siler y Martin 1991). Además, en base a la relación rigidez-frecuencia se planteó la hipótesis de que la rigidez disminuiría con la fatiga (Farley y Gonzales, 1996). Finalmente, similarmente a lo que ocurre en ausencia de fatiga, esperamos que los corredores auto-optimicen las características de la zancada seleccionando una frecuencia que minimice el costo metabólico.
MÉTODOS
En el estudio participaron 16 sujetos (5 mujeres, 11 varones) con una edad promedio de 28 (8) años, peso de 70,4 (10,5) kg, talla de 176 (10) cm y VO2máx de 59,1 (6,8) ml Kg-1 min-1 [media (SD)]. Todos los sujetos debían haber corrido por lo menos 20 millas (32 km) por semana durante al menos 4 semanas, antes de la recolección de los datos. Los sujetos no presentaban factores de riesgo de enfermedad cardiaca ni lesiones que pudieran afectar su desempeño. Antes de realizar en el estudio se explicaron los riesgos, beneficios y procedimientos a todos los sujeto. Los sujetos firmaron un documento de consentimiento informado antes de comenzar el estudio. El estudio fue aprobado por la Junta de Revisión Institucional de la Universidad Estatal de Oregón.
Todos los sujetos realizaron tres carreras. La primera carrera consistió en una prueba de esfuerzo progresiva para determinar el consumo de oxígeno máximo. Una semana más tarde, los sujetos realizaron una carrera preliminar para determinar una velocidad de carrera máxima individualizada que pudiera ser mantenida durante 1 h. La carrera comenzó a un ritmo estimado basado en el rendimiento de carrera de cada sujeto u en otra experiencia. Esto fue típicamente cerca de 96-99% de la velocidad de carrera del corredor en 10 kilómetros [basado en gráficos ritmo publicados en Gardner y Purdy (1970)]. Durante la carrera preliminar, se hicieron pequeños ajustes de velocidad para equiparar las percepciones de capacidad de cada corredor para completar la carrera de una hora con la velocidad más rápida posible. Las velocidades finales oscilaron entre 3,0 y 4,6 m/s, mientras que las tasas metabólicas correspondientes a la velocidad elegida tuvieron un valor promedio de 77,9 (6,8) % de VO2 máx.
La carrera preliminar y la carrera de recolección de datos fueron realizadas con al menos una semana de separación. El día de la recolección de datos, los sujetos realizaron una entrada en calor de 5 minutos corriendo a una velocidad elegida libremente. Después de la entrada en calor, comenzó la carrera de 1 h con la cinta rodante fijada a una velocidad predeterminada y continuó con un período de carrera de 5 minutos sin monitoreo para que los corredores tuvieran tiempo suficiente para alcanzar un consumo de oxígeno estable y permitir la adecuación de los sujetos a la carrera en cinta rodante (Candau et al. 1998, Costill 1967, Powers et al., 1984). Después de los primeros 5 min, los corredores completaron una etapa que incluía intervalos de 2 min de carrera donde corrían con una frecuencia de zancada preferida (PSF), y luego 4 y 8% por encima y por debajo de esa PSF. Las últimas cuatro frecuencias de zancada se completaron en orden aleatorio. A partir de este período de 15 minutos, los corredores continuaron sin variar la frecuencia de zancada durante otros 35 minutos, es decir, hasta cumplir 50 minutos de carrera. Durante estos 35 min, los sujetos podían quitarse las máscaras para la medición de oxígeno y debían colocárselas nuevamente 2 min antes de las mediciones finales de VO2. Después de 50 minutos de carrera, los últimos 10 min fueron similares a la etapa de frecuencia de zancada manipulada que se realizó cerca del comienzo de la carrera con una nueva aleatorización de las frecuencias de zancada alteradas.
El proceso de cálculo de la frecuencia de zancada óptima (OSF) implicó medir el consumo de oxígeno a través de un intervalo de frecuencias de zancada. Cavanagh y Williams (1982) midieron el consumo de oxígeno en siete etapas, lo que permitió determinar la frecuencia de la zancada a través de un rango de aproximadamente -12 a + 16% de PSF. En este estudio elegimos ±4% y ±8% debido a las elevadas velocidades de carrera obtenidas (tan altas como 4,6 m/s), lo que aumentaba la dificultad para que los corredores pudieran apartarse de la frecuencia de zancada preferida especialmente al final de la carrera. Un metrónomo computarizado ayudó a los corredores a mantener las frecuencias de zancadas modificadas, pero este metrónomo no se utilizó durante la etapa de frecuencia preferida para que los corredores pudieran seleccionar naturalmente su frecuencia. Este segmento de PSF fue siempre la primera de las cinco condiciones, ya que se trataba simplemente de una continuación de la carrera libre previa. Se midieron las fuerzas de reacción del suelo verticales de la cinta rodante y se utilizaron para determinar la frecuencia real de zancada durante los cambios de frecuencia. En el análisis posterior se utilizaron estas frecuencias medidas y no las frecuencias fijadas, generadas por el metrónomo.
Para permitir el acostumbramiento a las nuevas frecuencias de zancada y al nuevo consumo de oxígeno en estado estacionario, las mediciones de consumo de oxígeno se realizaron durante el segundo minuto de los segmentos de frecuencia de zancada preferida y alterada. Aunque 1 minuto es relativamente poco tiempo para alcanzar un estado estacionario, los cambios en el consumo de oxígeno provocados por las nuevas frecuencias de zancada fueron bastante pequeños, por lo que 1 minuto fue suficiente tiempo para adaptarse. Un estudio piloto confirmó que el consumo de oxígeno se restableció en aproximadamente 60 s después de un cambio de frecuencia.
La fuerza vertical se midió usando una cinta rodante Quinton Q55 modificada para soportar la cinta de rodamiento por seis transductores de fuerza piezoeléctricos uniaxiales (modelos PCB Piezotronics 208A02 y 208A03). Con un motor de accionamiento aislado de las vibraciones de la cinta de rodamiento y de los rodillos, en las señales de fuerza sólo se visualizaban las vibraciones de baja amplitud del motor y de la cinta de rodamiento de aproximadamente 60-70 Hz. La frecuencia de resonancia de la cinta rodante fue de aproximadamente 100-200 Hz, dependiendo del punto de impacto. La validación estática y dinámica de los instrumentos confirmaron que las magnitudes de la fuerza durante la carrera se podían predecir con un porcentaje de error inferior al 5% (Fewster 1996). Los datos de fuerza se muestrearon a 500 Hz en períodos de 30 s durante cada condición de frecuencia de zancada y posteriormente se utilizaron en la determinación de las frecuencias de zancada y en el cálculo de la rigidez vertical (Kvert) y rigidez de las piernas (Kleg).
Se asumió que la pierna en carrera se comportaba como un resorte lineal, de tal manera que la fuerza de reacción se determinó a través del desplazamiento del centro de masa del cuerpo y de la rigidez del resorte. El modelo de masa-resorte descrito por McMahon y Cheng (1990) y aplicado en otros numerosos estudios, tales como el de Farley y González (1996), describe la semejanza de las características de pierna con las de un resorte en cuanto a la fuerza de reacción vertical del suelo y el desplazamiento vertical del centro de masa. Si la pierna se comporta como un resorte lineal ideal, el gráfico de fuerza versus desplazamiento debería ser lineal y la pendiente sería igual a la rigidez. Aunque no son perfectamente lineales, las respuestas típicas de desplazamiento de fuerza para corredores se aproximan a esta relación. La rigidez del resorte puede ser estimada como la relación entre la fuerza máxima y el desplazamiento máximo. McMahon y Cheng (1990) propusieron dos relaciones de este tipo: rigidez vertical (Kvert) y rigidez de la pierna (Kleg) utilizando la fuerza de reacción vertical máxima del suelo divididas por desplazamiento vertical (ΔY) o por desplazamiento (compresión) a lo largo de la longitud de la pierna (ΔL) respectivamente. ΔY se determinó utilizando doble integración de las curvas de aceleración vertical-tiempo (Cavagna 1975), mientras que ΔL fue estimado siguiendo la metodología sugerida por McMahon y Cheng sobre la base de ΔY, longitud de la pierna, tiempo de posicionamiento y velocidad de carrera. En este estudio, en base a estudios antropométricos se estableció que la longitud de la pierna era 53% de la altura del cuerpo (Morin et al., 2005, Winter 1990). Sobre la base de la relación masa-resorte, se determinó la frecuencia de resonancia (RF) de la pierna en carrera usando la rigidez vertical e ignorando cualquier amortiguación presente en tales oscilaciones (Cavagna et al., 1997):
![]()
donde m es masa corporal.
Durante cada test de carrera de una hora, se controló periódicamente la velocidad de la cinta rodante con un tacómetro digital de mano (Modelo 21C13, Kernco Instruments), para asegurar una velocidad constante durante toda la carrera. Después de la recopilación de datos, se calcularon las frecuencias de zancada iniciales y finales preferidas y alteradas utilizando las muestras de 30 s de los datos de fuerza a partir de las cuales se extrajeron 20 pasos (10 zancadas) para el análisis. Los pasos fueron definidos desde el momento en que se apoyaba el talón con un pie hasta el momento en que se apoyaba el talón del otro pie; y el tiempo de apoyo de talón se definió como el primer punto de datos mayor que 20 N posterior a la fase de despegue donde las fuerzas son iguales a cero. El tiempo de zancada, se determinó a partir de dos pasos consecutivos. La frecuencia de zancada promedio se calculó obteniendo el promedio de diez zancadas.
El consumo de oxígeno se midió durante el segundo minuto de cada frecuencia de zancada preferida y alterada con un dispositivo de medición del metabolismo (metabolic cart) SensorMedics (Modelo 2900). El dispositivo se calibró con gases de concentración conocida antes de que cada sujeto corriera. El consumo de oxígeno se muestreó cada 20 s. Dado que la recolección durante cada segmento de la carrera demoraba un minuto completo, se promediaron los tres valores medidos en ese período para caracterizar el consumo de oxígeno para cada frecuencia de zancada. Se creó un polinomio de segundo grado con las cinco muestras de consumo de oxígeno en función de la frecuencia de zancada. La frecuencia de zancada correspondiente al valor mínimo de esta función representaba la frecuencia de zancada óptima (OSF), tal como se muestra en la Figura 1.
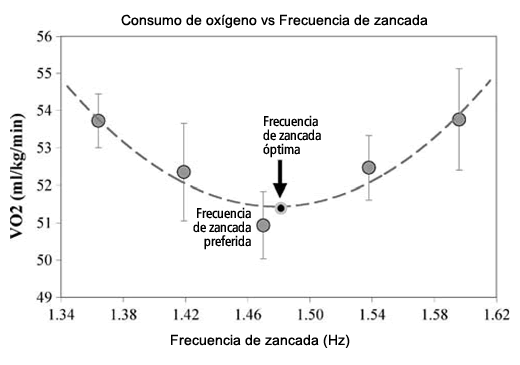
Figura 1. El método para determinar la frecuencia de zancada óptima (OSF) implicó la obtención de la curva de mejor ajuste de una ecuación de segundo grado con cinco puntos de datos (observados aquí con los datos obtenidos en un corredor). La OSF es el menor VO2 de la curva. La frecuencia de zancada preferida (PSF) fue la base a partir de la cual, se determinaron las otras condiciones de zancada (± 4 y ± 8% de PSF). Las frecuencias reales de cada corredor difieren ligeramente de las frecuencias objetivo marcadas con el metrónomo. Los valores medios de R2 para las ecuaciones de mejor ajuste fueron 0,73 y 0,71 (inicial y final, respectivamente)
Las comparaciones estadísticas utilizadas para contrastar las hipótesis de investigación de este estudio se llevaron a cabo de la siguiente manera: se utilizó el análisis de varianza de medidas repetidas para comparar las frecuencias de zancada preferidas y óptimas al comienzo y al final de la carrera. Además se utilizó el Test t de muestras apareadas para comparar los valores de las frecuencias de zancada, de paso y de resonancia, la rigidez y el VO2 obtenidos al principio con los valores correspondientes obtenidos al final de la carrera. Se plantearon comparaciones intra-sujeto de las características obtenidas al inicio versus las obtenidas al final usando los 20 pasos de cada corredor durante la condición de PSF. Se utilizaron test t de muestras no apareadas para las comparaciones de frecuencia de pasos intra-sujetos. El análisis de la covarianza (ANCOVA) se utilizó para analizar las comparaciones de rigidez intra-sujeto utilizando la frecuencia de paso como covariable. Las diferencias se consideraron significativas cuando a partir de las respectivas comparaciones, se obtenían probabilidades inferiores a 0,05. Además, para comprender mejor las relaciones entre frecuencia de zancada, rigidez y VO2, se calcularon las correlaciones de Pearson entre estas variables y también entre los cambios respectivos entre las condiciones iniciales y las finales de la carrera.
RESULTADOS
Una hora de carrera a ritmo cercano al ritmo de carrera es una tarea física exigente, que para los sujetos bien entrenados de este estudio demandó una distancia de casi 14 km. Cerca del inicio de la carrera, el costo metabólico fue de 45,9 (7,2) ml kg-1 min-1 y aumentó cerca de 3% hasta 47,4 (6,7) ml kg-1 min-1 en los últimos minutos de la carrera (P = 0,004). A medida que VO2 aumentaba, se observó una disminución de la frecuencia de la zancada cercana a 1-2% mientras los corredores se cansaban (P=0,026). No se observaron cambios significativos en frecuencia de resonancia, en la rigidez vertical ni en la rigidez de las piernas durante la carrera entre los sujetos, sin embargo, algunos corredores individualmente experimentaron cambios significativos en estas variables (Tabla 2). En la Figura 2 se presentan los valores de VO2, de frecuencia de zancada y de rigidez de los corredores desde el principio hasta el final de la carrera de una hora.
En la Tabla 1 se observan los valores medios de frecuencia de zancada y VO2 correspondientes todas las frecuencias de zancada utilizadas. Los cambios sistemáticos a través de las cinco condiciones demostraron que los corredores pudieron ajustar la frecuencia de zancada para satisfacer los requisitos del estudio. La Figura 2b presenta la amplia gama de frecuencias de zancada preferidas utilizadas por los corredores. A partir del ANOVA de medidas repetidas, se estableció la ausencia de diferencias entre la frecuencias de zancada preferidas y las óptimas ni al inicio ni cerca del final (P=0,978). Por lo tanto los corredores de este estudio fueron capaces de auto-optimizar sus frecuencias de zancada tanto al principio, cuando no estaban cansados como más tarde cuando si estaban cansados, después de casi una hora de carrera donde los valores de consumo de oxígeno y de frecuencia de zancada habían cambiado con respecto a los valores previos.
Tabla 1. Características medidas y obtenidas mediante un modelo de las condiciones de frecuencia de zancada (Media±SD). PSF sentó la base para determinar las condiciones de frecuencia. Las características de OSF fueron calculadas a partir del punto mínimo observado en el modelo de ecuación cuadrática en función del rango de frecuencias. La comparación estadística de los valores medios iniciales y finales se realizó mediante un test t de muestras apareadas. La comparación PSF versus OSF se realizó mediante un ANOVA de medidas repetidas (n=16 en cada grupo)
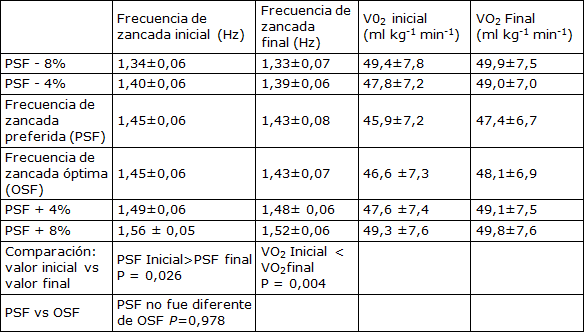
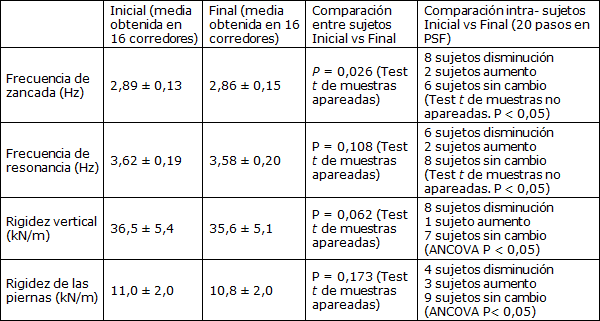
Tabla 2. Comparaciones de frecuencia y rigidez iniciales y finales, intra-sujeto e inter-sujetos (Media±SD).
La relación entre rigidez y frecuencia se puede observar en la Figura 3, donde una frecuencia de zancada mas alta se asoció con una mayor rigidez. Los datos medios de rigidez-frecuencia están casi alineados tanto en las condiciones iniciales como en las finales. Las pendientes similares de la relación rigidez-frecuencia en condiciones iniciales y finales demuestran una conexión mecánica invariable con comportamiento similar al de un resorte. Sin embargo, cuando los corredores se fatigan, se desplazan más hacia la izquierda de la relación con una frecuencia de zancada mas baja en comparación con la condición de ausencia de fatiga.
La Figura 2 presenta la considerable variabilidad de la respuesta a la fatiga que se observó entre los individuos. Para explorar estas diferencias intra-sujetos se realizaron comparaciones de la frecuencia de paso inicial y final, la frecuencia de resonancia, y la rigidez vertical y de las piernas a través de 20 pasos realizados con frecuencia de zancada preferida (Tabla 2). La variabilidad típica de la frecuencia de pasos fue de aproximadamente 2% (coeficiente de variación), mientras que las características de rigidez variaron aproximadamente en 5-7% a lo largo de los 20 pasos. Por lo tanto, la variabilidad intra-sujeto de la rigidez fue considerablemente mayor que las diferencias medias de rigidez observadas entre las condiciones iniciales y las finales. Las comparaciones de las frecuencias de resonancia intra-sujetos fueron consistentemente aproximadamente 20% mayores que la frecuencia de paso tanto al inicio como al final de la carrera y en las cinco condiciones de frecuencia.
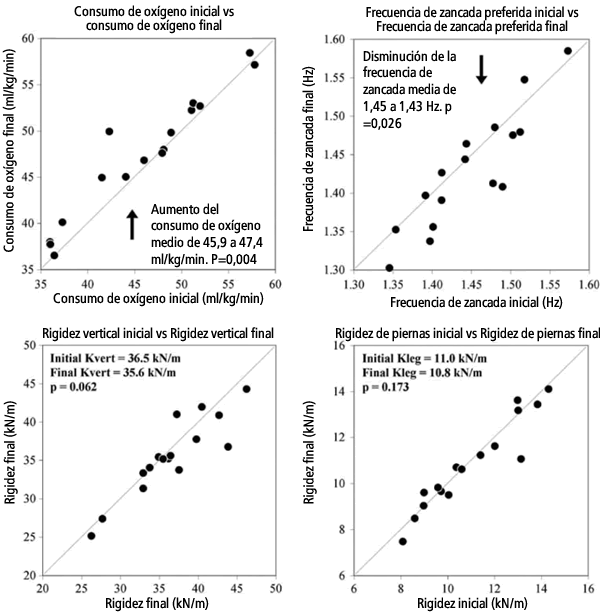
Figura 2. a-d Comparación entre los valores de consumo de oxígeno, frecuencia de zancada, rigidez vertical y rigidez de las piernas determinados al inicio y al final. En cada gráfico se dibujó una línea de identidad donde los valores iniciales y finales serían iguales, algo que sería equivalente a ausencia de cambios. La mayoría de los corredores experimentaron un mayor consumo de oxígeno (por encima de la línea), pero disminuyeron la frecuencia de zancada (por debajo de la línea) desde el principio hacia el final de la carrera de una hora. Aunque se observaron disminuciones significativas en la rigidez vertical y de las piernas en algunos sujetos, en general los cambios no fueron significativos.
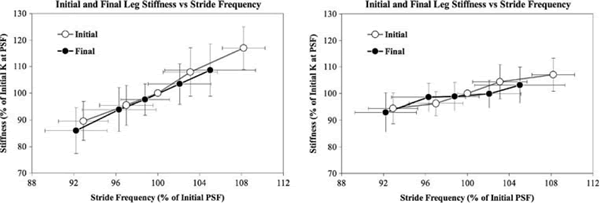
Figura 3. a, b: Valores medios de la rigidez vertical y rigidez de la pierna en función de la frecuencia de zancada. En el gráfico los valores de rigidez y frecuencia fueron normalizados en forma de porcentaje del valor de la frecuencia de zancada preferida. Los puntos de datos corresponden a valores medios obtenidos en cada una de las cinco frecuencias de zancada. Las barras de error corresponden a la desviación estándar entre sujetos para cada condición de frecuencia de zancada. Aunque se observó una relación de rigidez-frecuencia en condiciones de fatiga similar a la observada inicialmente, los corredores se desplazaron mas hacia la izquierda de la curva en las frecuencias más bajas. Aunque la frecuencia de zancada preferida final fue significativamente menor que la inicial, esto no puede ser atribuido a una incapacidad para mantener la rigidez de las piernas, ya que los corredores demostraron claramente que tenían esta capacidad en las condiciones finales +4 y + 8%. Es importante destacar que, a pesar de que en los gráficos es evidente la relación lineal de la rigidez vertical y la rigidez de las piernas con respecto a la frecuencia de zancada, las pendientes de la relación rigidez-frecuencia son considerablemente diferentes para las dos características. Frente a un cambio dado en la frecuencia de la zancada el cambio en la rigidez vertical es sustancialmente mayor al de la rigidez de las piernas
DISCUSIÓN
Uno de los supuestos de este estudio fue que una carrera de una hora realizada en una intensidad casi máxima provocaría una fatiga suficientemente alta en los sistemas fisiológicos y estructurales relevantes para inducir cambios medibles en el consumo de oxígeno, y quizás en las características de la zancada. Sobre la base de estudios previos realizados a 80% del VO2max (Sproule 1998, Xu y Montgomery 1995), era esperable que una carrera de una hora provocara aumentos porcentuales bajos en el consumo de oxígeno. Por lo tanto, el aumento de alrededor del 78-80% del VO2máximo observado aquí fue anticipado, aunque más pequeño que el observado en estudios previos, debido a diferencias en el tiempo de carrera, en los protocolos de evaluación o a las características de los sujetos.
Sobre la base del modelo masa-resorte y a lo observado en la Figura 3, existe una clara relación entre la frecuencia de zancada y la rigidez (Farley y González 1996). Esperábamos que la frecuencia de zancada, la rigidez vertical y de las piernas disminuyeran con la fatiga en función de observaciones previas y a respuestas conocidas de los músculos y tendones. En conjunto, la frecuencia de zancada disminuyó significativamente al final de la carrera de una hora, pero individualmente sólo la mitad de los corredores demostró este cambio. En general entre los sujetos, el stiffness no presentó cambios entre las condiciones iniciales y las finales (P = 0,062 y 0,173, respectivamente), pero individualmente la mitad de los corredores presentaron cambios significativos de stiffness, confirmando la tendencia del grupo. Por el contrario, los cambios de rigidez de las piernas para los corredores individuales fueron principalmente no significativos. Estas respuestas algo diferentes entre la rigidez vertical y la rigidez de las piernas probablemente se deban a cambios relativamente pequeños en la rigidez de las piernas en comparación con la rigidez vertical frente a variaciones equivalentes en la frecuencia de zancada (Figura 3).
En los corredores de este estudio se observaron considerables diferencias inter-individuales en respuesta a la fatiga (Figura 2). Algunos corredores presentaron consumo de oxígeno, frecuencias y características de rigidez vertical casi constantes, mientras que otros experimentaron cambios sustanciales. En otros estudios previos se observó la naturaleza individual de tales respuestas (por ejemplo, Dutto y Smith 2002, Siler y Martin 1991), pero no ha sido explicado con detalle.
A pesar de que los sujetos de este estudio tuvieron marcadas variaciones en VO2máximo (45-68 ml kg-1 min-1), en la velocidad de carrera para el test de una hora (3,0-4,6 m/s) y en la frecuencia de zancada preferida (1,35-1,57 Hz) , ninguna de estas características iniciales se asoció con la respuesta de un corredor frente a la fatiga. Como era de esperar, los sujetos que corrieron a mayor velocidad tuvieron un VO2 mayor (r=0,87), pero la velocidad de carrera sólo se relacionó débilmente con los cambios ocasionados por la fatiga en VO2 (r=-0,22), en la frecuencia de zancada (r =-0,37), en la rigidez vertical (r=0,29) o en la rigidez de las piernas (r = 0,39).
Inesperadamente, ninguno de los cambios en VO2, en la frecuencia o en la rigidez se asociaron claramente entre sí (todos presentaron correlaciones no significativas, r<0,50). Esto no coincide con los cambios significativos de rigidez y los cambios en la relación de frecuencia de zancada observados en el estudio de Dutto y Smith (2002), que también utilizó una carrera prolongada extenuante. Podría ser que en el presente estudio, las características de la zancada fueran quizás más variables debido a la necesidad de que durante las etapas de la prueba que se corrieron con frecuencias alternativas se respirara a través de una máscara con tuberías para determinar el consumo de oxígeno, lo que probablemente haya dificultado la detección de relaciones significativas. Sin embargo, observamos que la variabilidad del tiempo de zancada en los sujetos de este estudio (aproximadamente 1,0%) fue similar a la observada por Belli et al. (1995) (1,8% a una velocidad similar). Lamentablemente, las conexiones detrás de las diferencias interindividuales en respuesta a la fatiga siguen siendo poco claras. En estudios futuros sería útil incluir factores tales como patrones de actividad muscular y composición muscular para detallar más las características de respuesta individual durante una carrera exhaustiva.
La frecuencia de zancada óptima fue casi idéntica a la frecuencia de zancada preferida tanto al principio como al final de la carrera de una hora (P = 0,978). Sobre la base de trabajos previos de Cavanagh y Williams (1982), Cavagna et al. (1997) y Kaneko et al. (1987), era esperable la auto-optimización en la condición de ausencia de fatiga. Los desplazamientos hacia abajo de la frecuencia de zancada que se produjeron con la fatiga introdujeron un factor potencialmente perturbador en el proceso de optimización. El protocolo del estudio requería que los corredores alcanzaran una frecuencia de zancada que se ubicara por encima de la frecuencia preferida en la condición de fatiga. Así, los corredores serían claramente capaces de desarrollar mayor rigidez y, por tanto, mayor frecuencia (Figura 3). Pero esto sólo se lograría con un mayor costo metabólico por encima de la condición preferida. Mantener constantes la frecuencia y la rigidez durante una hora habría aumentado el costo de la carrera por encima del 3% observado. Sólo podemos especular acerca de cuales podrían ser las conexiones en esta relación; quizás la rigidez principal del complejo músculo-tendón haya provocado que los corredores recluten fibras musculares rápidas adicionales en comparación con el estado de activación inicial. Krustrup et al. (2004) observaron un aumento en el reclutamiento de fibras rápidas a lo largo del tiempo en carreras realizadas a 80% del VO2máximo. Esto puede aportar una explicación para el aumento del costo metabólico observado durante carreras extenuantes de larga duración. Además, la menor rigidez observada en algunos de los corredores podría influir en la magnitud del almacenamiento elástico de energía durante el posicionamiento. Un ajuste de este tipo que afecta el ciclo de estiramiento-acortamiento, también puede afectar negativamente el costo de la carrera. En un estudio se observó una correlación negativa entre la rigidez vertical y de las piernas y el costo de la carrera en un grupo de corredores (Dalleau et al., 1998, Heise y Martin, 1998). Sin embargo, tal como lo demuestran claramente los datos de este estudio (Figura 1), esto no implica que en un corredor individual el aumento de la rigidez (y por lo tanto de la frecuencia) pueda mejorar la economía. Cavagna et al. (1997) y Dalleau et al. (1998) sugirieron que las diferencias entre resonancia y frecuencia de zancada favorecen el costo de la carrera. Sin embargo, en este estudio, la diferencia entre frecuencia de resonancia y de zancada no varió con la fatiga y no permite explicar el aumento en el consumo de oxígeno. La optimización de la frecuencia de zancada probablemente se relacione con equilibrar el almacenamiento elástico de la energía frente al costo de acelerar las piernas. Lamentablemente no se conoce detalladamente como sería el control de estos componentes competitivos del proceso de optimización. Cualquiera sea el mecanismo, este experimento demuestra que la optimización sigue funcionando cuando un corredor está fatigado, a pesar de los cambios en la rigidez y la frecuencia de zancada que podrían producirse.
Finalmente, a partir de las cinco frecuencias de zancada en las que se midió el VO2, se observó un patrón consistente al determinar la frecuencia de zancada óptima. La frecuencia de zancada preferida se midió bajo condiciones de carrera libre (sin metrónomo) y produjo un consumo de oxígeno consistentemente más bajo que el estimado por la ecuación de regresión de segundo grado para esa frecuencia (es decir, el punto de datos de PSF estaba por debajo de la curva, Figura 1). Sospechamos que existe algún pequeño costo metabólico asociado con el ajuste de la frecuencia de zancada al metronomo, que tiende a elevar ligeramente los valores relacionados con ese control de frecuencia. Quizás se habría obtenido un conjunto de datos VO2-frecuencia más consistente si se hubiera medido primero la frecuencia de zancada preferida, y luego se hubiera incluido como una de las condiciones controladas por metrónomo para la determinación de la frecuencia optima de zancada. Sugerimos este enfoque para cualquier estudio de frecuencia de zancada controlada con metrónomo que se realice en el futuro.
En resumen, la carrera en cinta rodante cercana al 80% del VO2máx durante 1 h provocó cambios en las características fisiológicas y mecánicas. El consumo de oxígeno se desplazó hacia arriba alrededor del 3% durante la carrera, mientras que la frecuencia de la zancada disminuyó 1-2%. Las respuestas a la fatiga en términos de cambios en la frecuencia de la zancada y cambios en la rigidez vertical y de las piernas, fueron bastante variables entre los sujetos. Además, los corredores pudieron ajustar las características de rigidez-frecuencia para lograr minimizar el costo metabólico. En cada corredor, las frecuencias de zancada preferida y óptima fueron similares a pesar de los cambios provocados en la frecuencia óptima con la fatiga.
Referencias
1. Belli A., Lacour J.R., Komi P.V., Candau R., Denis C. (1995). Mechanical step variability during treadmill running. Eur. J. Appl. Physiol. 70:510–517
2. Candau R., Belli A., Millet G.Y., Georges D., Barbier B., Rouillon J.D. (1998). Energy cost and running mechanics during a treadmill run to voluntary exhaustion in humans. Eur. J. Appl. Physiol. 77:479–485
3. Cavagna G.A (1975). Force platforms as ergometers. J. Appl. Physiol. 39:174–179
4. Cavagna G.A., Mantovani M., Willems P.A., Musch G. (1997). The resonant step frequency in human running. Pflügers. Arch. Eur. J. Physiol. 434:678–684
5. Cavanagh P.R., Williams KR (1982). The effect of stride length variation on oxygen uptake during distance running. Med. Sci. Sports Exerc. 14:30–35
6. Costill D.L (1967). The relationship between selected physiological variables and distance running performance. J. Sports Med. Phys. Fitness. 7:61–66
7. Dalleau G., Belli A., Bourdin M., Lacour J.R (1998). The spring–mass model and the energy cost of treadmill running. Eur. J. Appl. Physiol. 77: 257–263
8. Derrick T.R., Dereu D., Mclean S.P (2002). Impacts and kinematic adjustments during an exhaustive run. Med. Sci. Sports Exerc. 34:998-1002.
9. Dutto D.J., Smith G.A (2002). Changes in spring–mass characteristics during treadmill running to exhaustion. Med. Sci. Sports Exerc. 34:1324–1331
10. Farley C.T., Gonzalez O. (1996). Leg stiffness and stride frequency in human running. J. Biomech. 29:181–186
11. Fewster J. (1996). The role of musculoskeletal forces in the human walk–run transition. Unpublished master’s thesis, Oregon State University, Corvallis, Oregon Gardner JB, Purdy JG (1970). Computerized running training programs. Track & Field News, Los Altos Heise GD, Martin PE (1998). ‘‘Leg spring’’ characteristics and the aerobic demand of running. Med. Sci. Sports Exerc. 30: 750–754
12. Kaneko M., Matsumoto M., Ito A., Fuchimoto T. (1987). Optimal step frequency in constant speed running. In: Jonsson B (ed). Biomechanics X–B. Human Kinetics, Champaign, pp 803–807
13. Krustrup P., Söderlund K., Mohr M., Bangsbo J. (2004). The slow component of oxygen uptake during intense, sub-maximal exercise in man is associated with additional fibre recruitment. Pflugers Arch. Eur. J. Physiol 447:855–866
14. Kubo K., Kanehisa H., Kawakami Y., Fukunaga T. (2001). Effects of repeated muscle contractions on the tendon structures in humans. Eur. J. Appl. Physiol. 84:162–166
15. Kuitunen S., Avela J., Kyröläinen H., Nicol C., Komi P.V. (2002). Acute and prolonged reduction in joint stiffness in humans after exhausting stretch–shortening cycle exercise. Eur. J. Appl. Physiol. 88:107–116
16. Kyröläinen H., Pullinen T., Candau R., Avela J., Huttunen P., Komi P.V (2000). Effects of marathon running on running economy and kinematics. Eur. J. Appl. Physiol. 82:297–304
17. Maclaren D.P.M., Gibson H., Parry-Billings M., Edwards RHT (1989). A review of metabolic and physiological factors in fatigue. In: Pandolf KB (ed). Exercise and sport sciences reviews. Williams & Wilkins, Baltimore, pp 29–66
18. McMahon T.A., Cheng G.C (1990). The mechanics of running: how does stiffness couple with speed? J. Biomech. 23:65–78
19. Millet G., Lepers R., Lattier G., Martin V., Babault N., Maffiuletti N. (2000). Influence of ultra-long-term fatigue on the oxygen cost of two types of locomotion. Eur. J. Appl. Physiol. 83:376–380
20. Morin J-B, Dalleau G., Kyröläinen H., Jeannin T., Belli A. (2005). A simple method for measuring stiffness during running. J. Appl. Biomech. 21:167–180
21. Noakes T.D. (2000). Physiological models to understand exercise fatigue and the adaptations that predict or enhance athletic performance. Scand. J. Med. Sci. Sports. 10:123–145
22. Place N., Lepers R., Deley G., Millet G.Y. (2004). Time course of neuromuscular alterations during a prolonged running exercise. Med. Sci. Sports Exerc. 36:1347–1356
23. Powers S.K., Dodd S., Garner R. (1984). Precision of ventilatory and gas exchange alterations as a predictor of the anaerobic threshold. Eur. J. Appl. Physiol. 52:173–177
24. Siler W.L., Martin P.E (1991). Changes in running pattern during a treadmill run to volitional exhaustion: fast versus slower runners. Int. J. Sport. Biomech. 7:12–28
25. Sproule J. (1998). Running economy deteriorates following 60 min of exercise at 80% VO2max. Eur. J. Appl. Physiol. 77:366–371
26. Winter D.A. (1990). Biomechanics and motor control of human movement. Wiley, New York
27. Xu F., Montgomery DL (1995). Effect of prolonged exercise at 65 and 80% of VO2max on running economy. Int. J. Sports Med. 16:309– 315