
Inervación del Músculo Esquelético y Tipos de Fibras Musculares
Publicado 6 de febrero de 2014, 23:48

Continuando con el análisis fisiológico del músculo esquelético, he diseñado la siguiente entrada con el propósito de identificar los rasgos característicos del proceso de inervación de la fibra muscular. Los músculos
esqueléticos están controlados por motoneuronas α, neuronas colinérgicas cuyo
soma se encuentra en el asta anterior de la médula espinal y en los núcleos
motores de los pares craneales. El conjunto formado por una motoneurona α y las
fibras musculares esqueléticas que inerva constituyen una unidad funcional
llamada unidad motora (Figura 1). El axón de una motoneurona α se ramifica para
inervar varias fibras musculares, cada una de ellas inervada por una sola
motoneurona. Así cada vez que la motoneurona genera un potencial de acción
todas las fibras musculares que inerva se contraen [1].
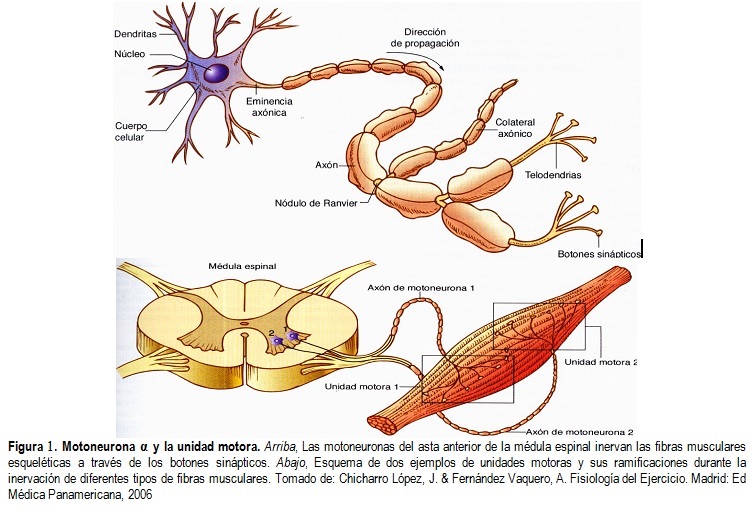
Las unidades motoras son heterogéneas desde el punto de vista morfológico, bioquímico y mecánico, difieren tanto en las características de las motoneuronas α como en las propiedades de las fibras musculares que las conforman. De esta manera, existen tres tipos distintos de unidades motoras:
- Tipo I, lentas y resistentes a la fatiga. Las fibras tipo I están acompañadas de producción energética aeróbica y son caracterizadas por una alta densidad mitocondrial, alta actividad de enzimas oxidativas, alta concentración de citocromo c, rico abastecimiento capilar, alta concentración de mioglobina y baja actividad de miosina-ATPasa [2]; así, las fibras musculares de este tipo son de tamaño pequeño, capaces de generar tensiones discretas durante periodos largos de tiempo sin fatigarse, además la fuerza que generan aumenta y disminuye de forma lenta. Las motoneuronas de estas unidades son de menor tamaño, menor velocidad de conducción y menor umbral de excitación con respecto a los otros tipos.
- Tipo IIX/IIB, rápidas y fatigables (las fibras tipo IIX son asignadas al hombre mientras que las fibras tipo IIB sólo existen en músculos de animales) están acompañadas de producción energética anaeróbica y son caracterizadas por una baja densidad mitocondrial, alta actividad de enzimas glucolíticas, alta actividad de CK, pobre abastecimiento capilar, baja concentración de mioglobina y alta actividad de miosina-ATPasa [2]. Poseen fibras musculares de tamaño grande que desarrollan mayor fuerza en cortos períodos de tiempo por emplear el metabolismo anaerobio. Sus motoneuronas son grandes, con elevadas velocidades de conducción y umbral de excitación [1].
- Tipo IIA, rápidas y resistentes a la fatiga. Combinan propiedades de I y IIX/IIB, ya que tienen capacidad aeróbica suficiente para resistir a la fatiga durante varios minutos.
Por otro lado, entre los axones de las motoneuronas y las fibras musculares esqueléticas se establece una sinapsis muy estudiada como modelo de transmisión química, que se llama unión neuromuscular o placa motora. Las fibras musculares están inervadas por fibras nerviosas grandes y mielinizadas. Cada fibra nerviosa se origina en las motoneuronas del asta anterior de la médula espinal, y tras penetrar el vientre muscular, se ramifica profundamente y estimula de tres a varios cientos de fibras musculares esqueléticas. A grandes rasgos, cada terminación nerviosa establece una sinapsis denominada unión neuromuscular con la fibra cerca de su punto medio, y el potencial de acción resultante viaja en ambas direcciones hacia los extremos de la fibra muscular y provoca su contracción.
Cerca de la unión neuromuscular, el nervio motor pierde su vaina de mielina y se divide en ramas finas que se denominan botones o pies terminales; estos contienen muchas vesículas pequeñas y claras albergando acetilcolina, el neurotransmisor de estas uniones. La acetilcolina, éster acético de la colina, es sintetizada en el citoplasma neuronal a partir de la unión de colina con acetato en presencia de acetil-CoA mediante la enzima acetilcolina transferasa y posteriormente es almacenada en las vesículas sinápticas, en las que se transporta a las terminaciones nerviosas donde se utiliza para la transmisión del impulso nervioso. Sin embargo, durante este proceso se requieren cuatro iones Ca2+ para abrir una vesícula colinérgica y además es imprescindible mantener una concentración mínima de Ca2+ extracelular de 10-4 M para que la conducción del impulso nervioso termine con la liberación de acetilcolina [3].
Debajo de la terminación nerviosa, la membrana muscular se engrosa e invagina formando la fosa o depresión sináptica; en esta zona la membrana de la célula muscular se resuelve en numerosos pliegues sinápticos que incrementan el área en la que puede actuar la acetilcolina (Figura 2). Esta estructura se denomina placa motora terminal, y está revestida por una o más células de Schwann que la aíslan de los líquidos del entorno debido a la vaina de mielina formada alrededor del axón.
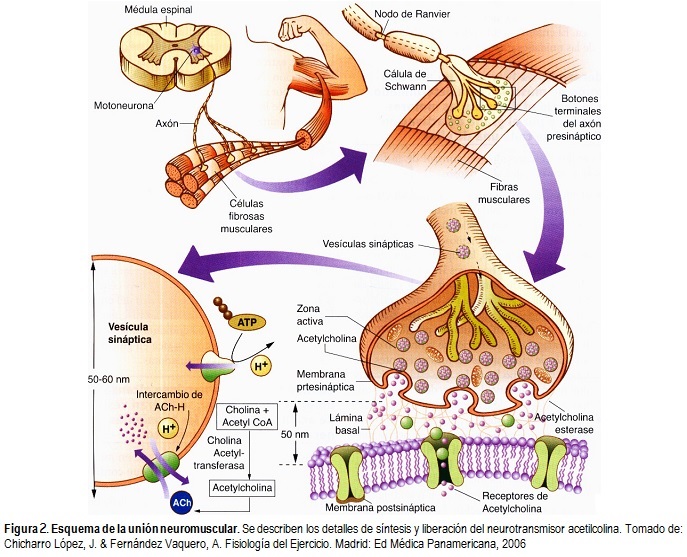
Existen tres tipos de
receptores nicotínicos en la unión neuromuscular, dos situados en la superficie
muscular y uno en la terminación del nervio parasimpático. Al liberarse las
moléculas de acetilcolina a partir de la terminación nerviosa presináptica,
éstas cruzan el espacio sináptico y se unen a los receptores postsinápticos
permitiendo el flujo de iones a través de ellos y despolarizando la placa
terminal. Los receptores postsinápticos están situados justo al lado opuesto de
donde se liberan las moléculas de acetilcolina y están compuestos por las
subunidades α, β, γ y δ
(Figura 3). En estos receptores existen dos subunidades α, las
cuales presentan los sitios de unión para la acetilcolina; así, cuando dos
moléculas de acetilcolina estimulan simultáneamente a las dos unidades α, se abre un canal en el receptor permitiendo el paso de Na+
y Ca2+ hacia el miocito y la salida de K+, se ha estimado
que 400.000 receptores se abren para crear el estímulo suficiente para generar
el potencial de acción que desencadena la contracción muscular [1].
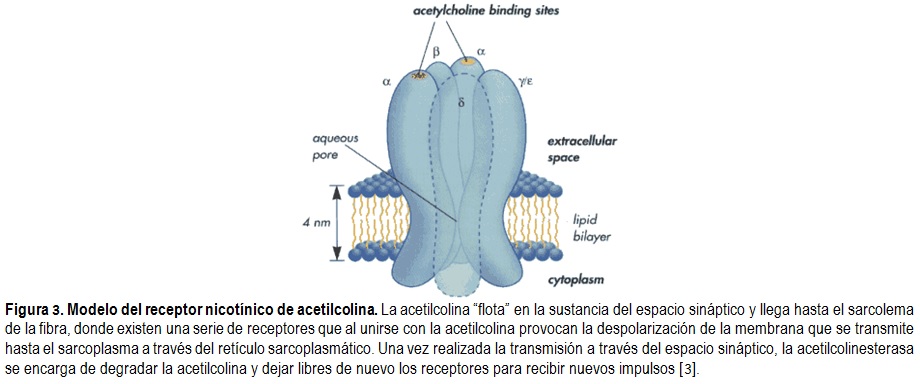
La acetilcolina se une durante un tiempo muy corto a sus receptores postsinápticos antes de ser degradada en acetato y colina por la acetilcolinesterasa que está concentrada en la hendidura de los pliegues sinápticos [3]. La denervación de un músculo rápido y su reinervación en un músculo lento puede hacer variar las propiedades de sus unidades motoras: las unidades motoras rápidas adquieren tras denervación/reinervación las propiedades de las unidades motoras lentas o viceversa. Esto se atribuye al cambio de la actividad fásica a la nueva actividad tónica impuesta por las motoneuronas que las reinervan, así el patrón de actividad sináptica podría variar los tipos de proteínas expresadas por las fibras musculares. Esto tiene gran importancia en fisiología del deporte ya que atletas entrenados en ejercicios que requieren alta aceleración y potencia muestran nervios motores con mayor velocidad de conducción y mayor expresión de la isoforma rápida de la ATPasa de la miosina [1].
REFERENCIAS
[1] Chicharro J & Fernández A. (2006). Fisiología del Ejercicio. Madrid: Ed Médica Panamericana.
[2] Casey A. & Greenhaff, PL. (2000). Does Dietary Creatine Supplementation Play a Role in Skeletal Muscle Metabolism and Performance? Am J Clin Nutr 72 (suppl); 607S–17S
[3] http://www.javeriana.edu.co/Facultades/Ciencias/neurobioquimica/libros/neurobioquimica