
Interacciones entre el Entrenamiento Concurrente de Fuerza y Resistencia
Duncan Mac Dougall1, Digby Sale1, S. Garner1 y I Jacobs1
Departamento de Educación Física y Medicina, Universidad de McMaster, Hamilton, ON; e Instituto de Defensa Civil de Medicina Ambiental, Downsview, ON, Canadá
Artículo publicado en el journal PubliCE, Volumen 0 del año 1999.
Publicado 18 de junio de 2007
Resumen
Palabras clave: músculo esquelético, adaptación, potencia aeróbica, actividad de las enzimas
El entrenamiento de la fuerza y la resistencia representan, en sus extremos, formas opuestas de entrenamiento. El entrenamiento de la fuerza consiste de un número relativamente pequeño de contracciones de fuerza máxima o casi máxima. El entrenamiento de la resistencia consiste de un gran número de contracciones submáximas. De acuerdo con esto, las respuestas adaptativas en el músculo esquelético al entrenamiento de la fuerza y de la resistencia son diferentes, y a veces opuestas. El entrenamiento de la fuerza provoca la hipertrofia en las fibras musculares (26) junto con un aumento en las proteínas contráctiles (28), lo que contribuye al incremento en la máxima fuerza de contracción. Por el contrario, el entrenamiento de la resistencia normalmente causa poca o ninguna hipertrofia en las fibras musculares (2, 7, 12, 20, 23), pero produce un aumento en las siguientes adaptaciones que se suponen mejoran el rendimiento de resistencia: densidad capilar (2, 7, 20, 37,39), densidad del volumen mitocondrial (17), y actividad de la enzimas oxidativas (12, 37). El entrenamiento de la fuerza y la resistencia, con frecuencia, es realizado en forma conjunta por los entusiastas del “fitness” y por los deportistas. Sin embargo debido a que las respuestas adaptativas al entrenamiento de fuerza y resistencia son diferentes, y algunas hasta podrían ser opuestas, es concebible que el músculo esquelético no pueda adaptarse de forma óptima a dos estímulos contrarios cuándo están simultáneamente superpuestos. Por ejemplo, el entrenamiento de fuerza podría causar una disminución en la densidad capilar (36, 41) y de la densidad mitocondrial (29), lo cual iría en contra del aumento en estas variables inducidas por el entrenamiento de resistencia. El entrenamiento de resistencia ha estado asociado con una pérdida en la fuerza (5, 31,33) y con un menor tamaño en las fibras musculares (22, 39), cambios obviamente antagonistas al desarrollo de fuerza.
Por el contrario, el entrenamiento concurrente de la fuerza y la resistencia podría interactuar para mejorar, más que para impedir el desarrollo de la fuerza y la resistencia.
Algunas formas de entrenamiento de resistencia han mostrado inducir incrementos en la fuerza (31, 34) y en el tamaño de las fibras musculares (2, 12). El entrenamiento de la fuerza ha mostrado inducir incrementos en la resistencia de corta (4-6 min) y larga (14-16) duración, en la potencia aeróbica máxima (14), y la actividad de las enzimas oxidativas (6).
Que la interacción entre entrenamiento concurrente de fuerza y potencia resulta en efectos “antagónicos” o una mejoría mutua de la respuesta al entrenamiento probablemente dependa de varios factores, incluyendo el nivel inicial de los deportistas; los modos de entrenamiento; la intensidad, volumen, y frecuencia de entrenamiento; y la forma en que se integran los dos tipos de entrenamientos. En sujetos previamente desentrenados, una combinación de entrenamiento de resistencia y fuerza, de intensidad y volumen moderados a altos, impidió el desarrollo de la fuerza pero no el incremento en la potencia aeróbica máxima o en la resistencia de corta duración (9, 14, 19). En sujetos previamente entrenados en resistencia, el agregado de entrenamiento de fuerza no provocó el desarrollo desigual de la fuerza como se observó en los sujetos desentrenados (19), y mejoró la resistencia sin provocar un aumento concomitante en la potencia aeróbica (15).
En el presente estudio, se examinó la interacción entre entrenamientos concurrentes de fuerza y resistencia de alta intensidad y volumen moderado en sujetos previamente desentrenados. Un aspecto singular del estudio fue que se realizaron varias mediciones relacionadas con la adaptación de los músculos esquelético para ayudar a explicar cualquier evidencia de antagonismo o mejoría mutua que pudieran ser encontradas.
METODOS
Sujetos
Los sujetos fueron ocho mujeres y ochos varones jóvenes sin experiencia previa en entrenamientos de fuerza o resistencia de alta intensidad. En concordancia con los Lineamientos Éticos para la Investigación con Sujetos Humanos de la Universidad McMaster, se les explicó a los sujetos los propósitos y los riesgos físicos, psicológicos, y/o sociales que implicaba la participación en la investigación, luego de los cual los estudiantes firmaron un consentimiento escrito. El proyecto contó con la aprobación del Comité de Ética de la Universidad McMaster.
Diseño
La mitad (4 mujeres, 4 varones) de los sujetos (grupo A) fueron asignados aleatoriamente a entrenar ambas extremidades inferiores en fuerza, y una extremidad en resistencia. La otra mitad de los sujetos (grupo B) entrenó ambas piernas en resistencia y una en fuerza. Por lo tanto, en el grupo A se evaluó la interacción entre el entrenamiento de fuerza y resistencia (F + E), en comparación con el entrenamiento de fuerza solamente (F), mientras que en el grupo B se evaluó la interacción entre entrenamiento de resistencia y fuerza (E + F), en comparación con entrenamiento sólo de resistencia (E). En la Tabla 1 se presentan las características de los sujetos de los dos grupos.
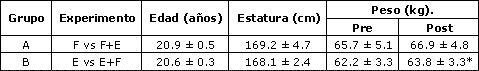
Tabla 1. Edad, estatura y peso corporal de los sujetos. Los valores
representan las medias ± EE; cada grupo estuvo formado por 4 mujeres y 4
varones. F, entrenamiento de fuerza; E, entrenamiento de resistencia, (*)
P<0.05, para el efecto principal en el peso corporal, pre-post entrenamiento.
Entrenamiento
El entrenamiento consistió en dos períodos de 11 semanas divididos por un período de 3 semanas, el último se planificó de manera tal que coincidiera con el receso de Navidad. Durante los períodos de entrenamiento los sujetos participaron en tres sesiones semanales de entrenamiento. Cada sesión duraba 1 hora, el tiempo que llevaba entrenar tanto fuerza y resistencia en una pierna, como la fuerza (Grupo A) o la resistencia (Grupo B) en la otra pierna. En los entrenamientos combinados de fuerza y resistencia, los ejercicios de resistencia precedieron a los de fuerza.
Entrenamiento de fuerza. Para el entrenamiento de la fuerza se utilizó el ejercicio de prensa de piernas llevado a cabo en una máquina tipo Universal (Global Gym, Downsview, Ontario). El movimiento (fase de contracción concéntrica) incluía extensión simultánea de cadera y rodilla y la flexión plantar del tobillo. Cada repetición incluyó una fase de contracción concéntrica y una fase de contracción excéntrica. Los sujetos en el Grupo A realizaron seis series de 15-20 repeticiones máximas (RM), el mayor peso levantado estuvo asociado al número de repeticiones designado para cada pierna en el movimiento de prensa de piernas. Las series con cada pierna se realizaron en forma alternada, con 1 min de pausa entre las series, y hasta que cada pierna completara las seis series. Los sujetos en el Grupo B realizaron seis series de 15-20 RM, y la pierna con la que realizaron el ejercicio fue asignada aleatoriamente. Se realizaron pausas de 2 min entre series sucesivas.
Entrenamiento de resistencia. El entrenamiento de la resistencia se realizó en un cicloergómetro (Monark). El entrenamiento consistió de cinco series de 3 min a una potencia correspondiente al 90-100% del VO2 máx. En el Grupo A, que entrenó la resistencia con sólo una pierna, la cual fue asignada aleatoriamente, se permitió una pausa de 3 min entre series sucesivas. El Grupo B realizó las series de 3 min alternando las piernas y con 1 min de pausa entre series sucesivas. En la Tabla 2 se resume el programa de entrenamiento.

Tabla 2. Programa de entrenamiento. Cada grupo estuvo formado por 4
mujeres y 4 varones. El entrenamiento de resistencia se realizó en bicicleta
ergométrica a una potencia correspondiente al 90-100 % del VO2 máx.
El entrenamiento de fuerza consistió de 6 series de 15-20 repeticiones con el
mayor peso posible (RM) en un aparato de prensa de piernas. Cada sesión tuvo una
duración de 1 hora; se llevaron a cabo 3 sesiones semanales en días no
consecutivos; y los dos períodos de entrenamiento de 11 semanas fueron separados
por un período de descanso de 3 semanas.
Mediciones
Todas las mediciones fueron llevadas a cabo antes y después de las 22 semanas de entrenamiento.
Potencia aeróbica. La potencia aeróbica (VO2 máx., definido aquí como el consumo máximo de O2 alcanzado durante el test con una pierna) fue medida para cada pierna en forma separada. El test se llevó a cabo en una cicloergómetro con cupla electromagnética (Jaeger) utilizando un protocolo estándar con aumento progresivo y continuo de la carga. El test comenzaba a una potencia de 30 a 45 W, y la carga se incrementaba en 15 W cada 2 min, hasta el agotamiento. Se utilizó un monitor electrocardiográfico y un sistema computarizado de análisis de gases de circuito abierto para obtener los datos de la frecuencia cardíaca, flujo espirado, consumo de O2 , producción de CO2 , e índice de intercambio respiratorio cada 20 min.
Fuerza voluntaria. La fuerza voluntaria de cada pierna fue definida como el máximo peso que se podía levantar en una repetición (1 RM) en el aparato utilizado para el entrenamiento con sobrecarga. Las mediciones fueron realizadas con una precisión de 0.5 Kg utilizando pesas que podían ser colocados en la pila de pesas del aparato. Los sujetos utilizaron el mismo aparato y posición corporal para todos los tests y para el entrenamiento. Este test fue llevado a cabo en dos ocasiones separadas, antes del entrenamiento, y se tomó el mayor valor obtenido como valor pre-entrenamiento.
Resistencia relativa. La resistencia relativa fue definida para cada pierna como el número de repeticiones que podrían realizarse al 80 % de 1RM en el ejercicio de prensa.
Resistencia absoluta. Al final del programa de entrenamiento se midió la resistencia absoluta de cada pierna, definida como el mayor número de repeticiones realizadas con la carga de 1RM pre-entrenamiento en el ejercicio de prensa. Para ambos tests de resistencia se utilizó una tasa de repeticiones de 10 repeticiones/min la cual fue controlada mediante la utilización de un metrónomo.
Área de Sección Cruzada Muscular. El área de sección cruzada de los músculos extensores y flexores de la rodilla derecha e izquierda fue determinada a partir de imágenes registradas mediante tomografía computada (TC) obtenidas con un escáner para TC (modelo 20-30, Ohio-Nuclear). Los cortes de TC fueron realizados al nivel del muslo medio (entre el trocánter mayor y el epicóndilo lateral del fémur).
Perímetros y Pliegues del Muslo. Se midió el perímetro de las piernas derecha e izquierda con una cinta de acero al nivel del muslo medio. En este mismo sitio se midieron los pliegues anterior, posterior, lateral, y medial con calibres de tipo Harpenden.
Superficie, Composición, y Capilarización de las Fibras Musculares. Se obtuvieron biopsias musculares del vasto lateral de las piernas derecha e izquierda. Parte de una muestra fue montada en un medio fijo y congelada en isopentano con nitrógeno líquido para posteriores análisis histoquímicos, y la parte restante fue estudiada utilizando un microscopio de barrido de electrones. Las fibras musculares fueron clasificadas como lentas (ST o de Tipo I) o rápidas (FT o de Tipo II), luego del teñido para determinar la actividad de la adenosín trifosfatasa miofibrilar (ATPasa) a un pH de 9.4, luego de la preincubación a un pH 10.3 (13). La composición de las fibras musculares (% ST) fue determinada a partir de muestras que contenían un mínimo de 218 (máx.=1.392) fibras. Se utilizaron fotografías de secciones seriales teñidas para NADH-tetrazolio-reductasa (32) para determinar la superficie promedio de cada tipo de fibra usando planimetría computarizada. Las medias ± DE del número de fibras por muestra utilizado para calcular la superficie varió de 50 ± 23 a 83 ± 25 para ST, y de 49 ± 21 a 96 ± 60 para FT. Los capilares fueron visualizados mediante tinción con ácido periodico de schiff en amilasa (1). Los capilares por fibra y capilares por milímetro cuadrado fueron determinados a partir de campos de 50 fibras.
Actividades de las Enzimas Musculares. Una segunda biopsia fue congelada directamente en nitrógeno liquido y guardada a –70ºC hasta su análisis. En el momento del análisis, el tejido fue secado, y luego las impurezas visibles, el tejido conectivo, y los coágulos de sangre fueron disecados, antes de que el tejido fuera pesado en una balanza eléctrica (Cahn). El tejido era luego homogeneizado por sonicación en un medio con hielo y buffer de fosfato 0.1 M, pH 7.7, para determinar las actividades de la fosfofructoquinasa (PFK), lactato deshidrogenasa (LDH), y citrato-sintasa (CS) con métodos enzimáticos acoplados con NADH (11, 25).
Análisis Morfométrico. El tejido estudiado en el microscopio de barrido de electrones fue inmediatamente fijado en glutaraldeído, secado en etanol, y embebido en Epon, usando técnicas estándar. Secciones levemente oblicuas fueron luego fotografiadas con una amplificación de ~ × 50.000, bajo un Phillips EM200. Un promedio de 52 fibras (rango, 37-60) fueron aleatoriamente seleccionadas por biopsia, y para cada biopsia se seleccionó aleatoriamente y se fotografió un campo fotográfico para el interior de cada fibra. En cada micrografía se realizó un análisis estereológico por medio de un sistema de test de líneas cortas de 168 puntos (42) de acuerdo al método descrito por Hoppeler et al (18). Se calcularon las densidades volumétricas de miofibrillas (Vvmiof), mitocondria interior (Vvmit), lípidos (Vvlip), y citoplasma (Vvcit).
Análisis Estadísticos
Los datos fueron analizados utilizando el análisis de variancia de dos factores (pre-post-entrenamiento y condición de entrenamiento), con mediciones de un factor (pre-post-entrenamiento). El análisis indicaría antagonismo o “adición” en el entrenamiento concurrente de fuerza y resistencia como una interacción significativa entre los dos factores (pre-post-entrenamiento × condición de entrenamiento). Cuando se observaba una interacción significativa, se llevaba a cabo un test post-hoc (Tukey A) para identificar diferencias estadísticas ente los valores medios. Se aceptó una significancia estadística de p<0.05. La estadística descriptiva incluyó las medias ± EE.
RESULTADOS
El mismo patrón de resultados se observó en el pequeño número de varones y mujeres en cada grupo; por lo tanto, los resultados tanto de varones como de mujeres fueron combinados para su análisis.
Efecto De Entrenamiento Concurrente de Fuerza y la resistencia sobre el Desarrollo de la Fuerza (Grupo A).
Fuerza voluntaria. La fuerza en 1RM en el ejercicio de prensa de piernas aumentó (p=0.006) tanto en la pierna entrenada en fuerza (F) como en la entrenada en fuerza-resistencia (F+E) (Figura 1) . A pesar de que la pierna F tuvo un mayor aumento (30.5 kg, 30.2 %) que la pierna F+E (21.2 kg, 20.4%), la interacción entre pre-post-entrenamiento y la condición de entrenamiento no fue significativa (p=0.18).
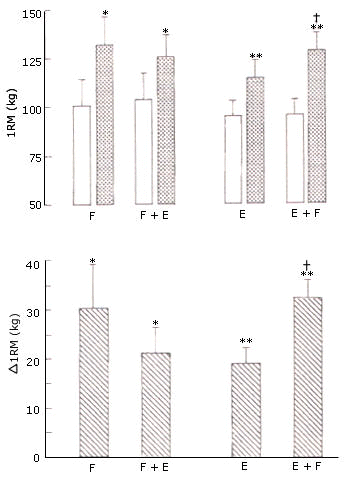
Figura 1. Fuerza en una repetición máxima (1RM) en el ejercicio de prensa
de piernas en el grupo A (arriba izquierda), que entrenó la fuerza con una
pierna (F), y la fuerza + la resistencia con la otra (F+E); y en el grupo B
(arriba derecha), que entrenó la resistencia con una pierna (E), y la
resistencia + fuerza en la otra (E+F). Arriba: valores pre (barras blancas) y
post (barras con puntos) entrenamiento. Abajo: aumentos después del
entrenamiento. Los valores representas las medias ± EE. Efecto principal pre vs
post entrenamiento:(*) p=0.006, (**) P< 0.001; Interacción (E+F > E): (†)
P<0.001.
Área de Sección Cruzada Muscular. El área de sección cruzada de los extensores aumentó (p<0.001) de manera similar en las piernas F (12.9%) y F+E (12.2%) (Figura 2). El área de sección cruzada de los flexores y el cociente entre las áreas de sección cruzada de los extensores y los flexores no cambió después del entrenamiento.
Perímetro del Muslo, Pliegues Cutáneos, y Peso Corporal. En general el perímetro del muslo aumentó leve pero significativamente (p=0.0008) (Figura 3). La pierna F+E no mostró un aumento significativamente mayor que la pierna F (interacción p=0.171). La suma de cuatro pliegues cutáneos no cambió significativamente luego del entrenamiento (Figura 3). El peso corporal no cambió significativamente después del entrenamiento (Tabla 1).
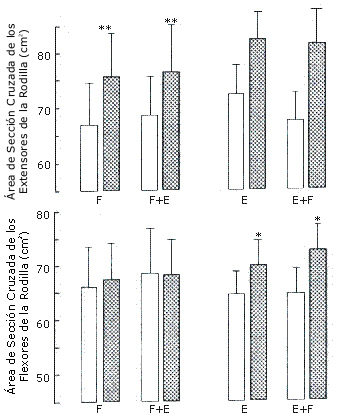
Figura 2. Área de sección cruzada de los músculos extensores (arriba) y
flexores (abajo) de la rodilla en el grupo A (izquierda) que realizó los
protocolos de entrenamiento F y F+E; y en el grupo B (derecha) que realizó los
protocolos de entrenamiento E y E+F. Los valores representan las medias ± EE,
pre (barras blancas) y post (barras con puntos) entrenamiento. Efecto principal
pre vs post entrenamiento:(*) p= 0.012, (**) p<0.001.
Figura 3. Perímetro del muslo (arriba) y suma de cuatro pliegues cutáneos
(anterior, posterior, medial, lateral) (abajo) en el grupo A (izquierda), que
realizó los protocolos de entrenamiento F y F+E; y en el grupo B (derecha) que
realizó los protocolos de entrenamiento E y E+F. Los valores representan las
medias ± EE, pre (barras blancas) y post (barras con puntos) entrenamiento.
Efecto principal pre vs post entrenamiento (*), p<0.01 (**) p<0.001; Interacción
(E±F > E): (†) P<0.05.
Superficie de las Fibras Musculares. No hubo cambios significativos en la superficie de las fibras ST (F, 3946 ± 538 a 4675 ± 537; F+E, 4481 ± 491 a 5412 ± 1196) o FT (F, 4693 ± 619 a 5378 ± 783; F±E 4884 ± 685 a 5738 ± 1209) (medias ± EE, um2). La relación del área FT/ST no cambió (F, 1.22 ± 0.12 a 1.15 ± 0.10; F+E, 1.09 ± 0.09 a 1.08 ± 0.08).
Potencia aeróbica máxima (VO2 máx). Tanto las extremidades que entrenaron la fuerza (7.9 %) como las que entrenaron la resistencia + la fuerza (8.3%) exhibieron incrementos similares en el VO2 máx (p=0.006.Figura 4).
Resistencia durante el Levantamiento de Pesas. La resistencia relativa (repeticiones al 80 % de 1RM) aumentó luego del entrenamiento (p=0.013, Figura 5). Hubo una interacción significativa (p=0.013), indicando que la pierna con la cual se realizaron los entrenamientos F+E (152 %) mejoró más en ésta variable luego del entrenamiento que la pierna con la cual se entrenó la F (81 %). Las extremidades con las cuales se realizaron los entrenamientos F (26.8) y F+E (24.4) mostraron resultados similares respecto del número de repeticiones que se pudieron hacer con la carga de 1RM pre-entrenamiento después del mismo (resistencia absoluta, Figura 6).
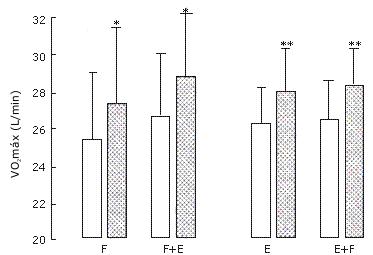
Figura 4. Potencia aeróbica máxima en el grupo A (izquierda) que realizó
los protocolos de entrenamiento F y F+E; y en el grupo B (derecha) que realizó
los protocolos de entrenamiento E y E+F. Los valores representan las medias ±
EE, pre (barras blancas) y post (barras con puntos) entrenamiento. Efecto
principal pre vs post entrenamiento: (*) p=0.006,(**) p<0.001.
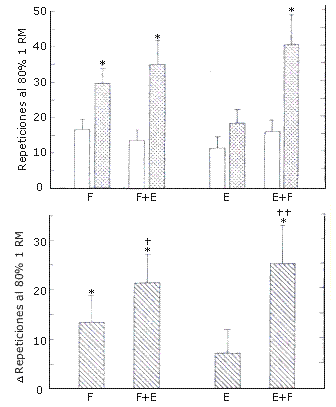
Figura 5. Repeticiones al 80 % de 1RM en el grupo A (izquierda) que
realizó los protocolos de entrenamiento F y F+E; y en el grupo B (derecha) que
realizó los protocolos de entrenamiento E y E+F. Arriba: valores pre (barras
blancas) y post (barras con puntos) entrenamiento. Abajo: aumento en las
repeticiones luego del entrenamiento. Los valores representan las medias ± EE.
Efecto principal pre vs post entrenamiento: (*) p<0.05; interacción: (†) p =
0.013(F+E>F), (††) p<0.001 (E+F>E).
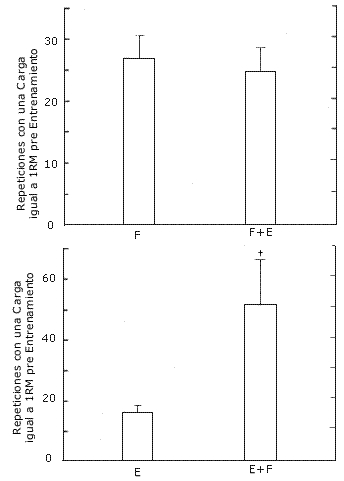
Figura 6. Repeticiones realizadas, después del entrenamiento, con la carga
de 1RM pre-entrenamiento en el grupo A (arriba) que realizó los protocolos de
entrenamiento F y F+E; y en el grupo B (abajo) que realizó los protocolos de
entrenamiento E y E+F. Los valores Representan las medias ± EE. Interacción (E+F>E):
p<0.05.
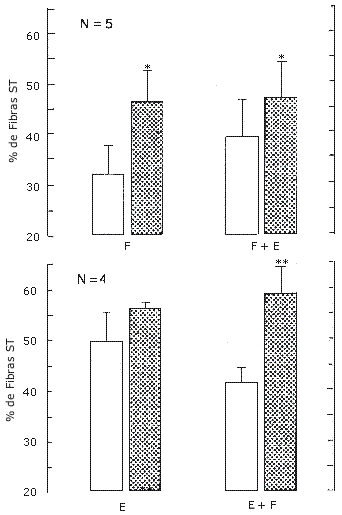
Figura 7. Porcentaje de fibras lentas (ST) del vasto lateral en el grupo A
(arriba) que realizó los protocolos de entrenamiento F y F+E; y en el grupo B
(abajo) que realizó los protocolos de entrenamiento E y E+F. Los valores
representan las medias ± EE, pre (barras blancas) y post (barras con puntos)
entrenamiento. Efecto principal pre vs post entrenamiento (*) p=0.007, (**)
p<0.001.
Porcentaje de Fibras ST y Capilarización. Hubo un aumento significativo (P=0.007) en el % de fibras ST tanto en la pierna F (31.8 - 46.5) como en la pierna F+E (39.2 - 47.0) (Figura 7). Los capilares por fibra (F, 1.92 ± 0.27 a 1.76 ± 0.26; F+E, 2.15 ± 0.32 a 2.26 ± 0.39) y los capilares por milímetro cuadrado de superficie muscular (F, 260 ± 15 a 247 ± 37; F+E, 256 ± 32 a 245 ± 39) no cambiaron significativamente luego del entrenamiento.
Actividades de las Enzimas Musculares. Las actividades (mkatal/kg de peso seco) de la PFK (F, 0.095 ± 0.015 a 0.098 ± 0.014; F±E, 0.087 ± 0.013 a 0.091 ± 0.012) y de la LDH (F, 5.03 ± 0.46 a 5.12 ± 0.61; F+E, 4.54 ± 0.41 a 4.59 ± 0.33) no cambiaron significativamente luego del entrenamiento. Por el contrario la actividad de la CS aumento significativamente (p<0.001, Figura 8). También hubo una interacción significativa (p = 0.005) para esta enzima. La pierna F+E (51.2 %) mostró un mayor incremento que la pierna F (22.1 %).
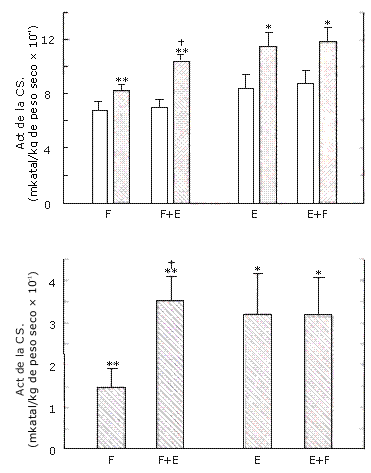
Figura 8. Actividad de la citrato-sintasa (CS) en el grupo A (izquierda)
que realizó los protocolos de entrenamiento F y F+E; y en el grupo B (derecha)
que realizó los protocolos de entrenamiento E y E+F. Arriba: valores pre (barras
blancas) y post (barras con puntos) entrenamiento. Abajo: aumentos después del
entrenamiento. Los valores representan las medias ± EE. Efecto principal pre vs
post entrenamiento:(*) p = 0.003, (**) P<0.001; interacción (F+E> F): (†) p =
0.005.
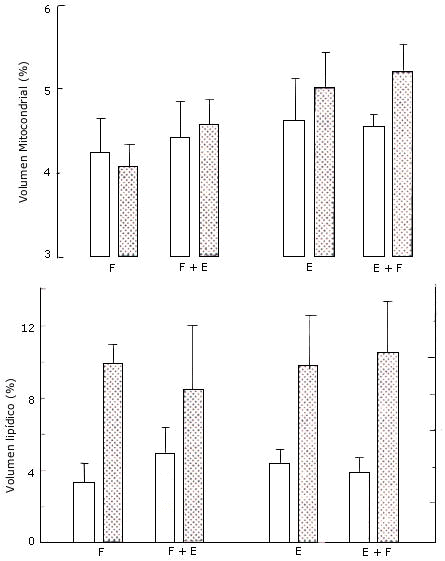
Figura 9. Densidad del volumen mitocondrial (arriba) y lipídico (abajo)
en el vasto lateral en el grupo A (izquierda) para que realizó los protocolos de
entrenamiento F y F+E; y en el grupo B (derecha) que realizó los protocolos de
entrenamiento E y E+F. Los valores representan las medias ± EE, pre (barras
blancas) y post (barras con puntos) entrenamiento.
Densidad del Volumen Mitocondrial y Lipídico. La densidad del volumen mitocondrial pareció levemente mayor en la pierna F+E luego del entrenamiento, pero el cambio no fue estadísticamente significativo (Figura 9). Además, hubo grandes aumentos (F, 194 %; F+E, 70 %) en la densidad del volumen de lípidos, aunque tampoco se alcanzó significancia estadística (p = 0.085, Figura 9).
Efecto del Entrenamiento Concurrente de Resistencia y de Fuerza sobre el Desarrollo de la Resistencia (Grupo B)
Potencia aeróbica máxima (VO2 máx). Tanto la extremidad con la que se realizó el protocolo E (6.9 %) como con la que se realizó el protocolo E+F (7.2 %) mostraron aumentos similares en el VO2 máx después del entrenamiento (p<0.001, Figura 4).
Resistencia durante el Levantamiento de Pesas. La resistencia relativa (repeticiones al 80 % de 1RM) aumentó luego del entrenamiento (p = 0.022, Figura 5), observándose una interacción significativa (p = 0.030). En la pierna con la cual se realizó el protocolo E+F (157 %), se observó un incremento significativo de la resistencia relativa, lo cual no ocurrió con la pierna con la que se realizó el protocolo E (60 %). Luego del entrenamiento, la pierna E+F (51) pudo hacer más repeticiones con la carga igual a 1RM pre-entrenamiento (resistencia absoluta) (p=0.032) que la pierna E (16) (Figura 6).
Porcentaje de Fibras ST y Capilarización. Hubo un aumento significativo (p<0.001) en el % de fibras ST tanto en la pierna E (49.9 - 56.3) como en la pierna E+F (41.6 - 58.8) (Figura 7). Los capilares por fibra (E, 2.38 ± 0.22 a 2.19 ± 0.22; E+F, 2.48 ± 0.26 a 2.49 ± 0.26) y los capilares por milímetro cuadrado de superficie muscular (E, 308 ± 16 a 267 ± 13; E+F, 309 ± 22 a 265 ± 24 no cambiaron significativamente luego del entrenamiento.
Actividades de las Enzimas Musculares. Las actividades de la PFK (E, 0.096 ± 0.011 a 0.101 ± 0.012; E+F, 0.093 ± 0.005 a 0.098 ± 0.007) y la LDH (E, 4.46 ± 0.77 a 5.09 ± 0.77; E±F 5.28 ± 0.77 a 5.28 ± 0.63) no cambiaron significativamente luego del entrenamiento. Por el contrario, la actividad de la CS aumentó significativamente (p = 0.003), y de manera similar en las piernas E (38,1 %) y E+F (36,3 %) (Figura 8).
Densidad del Volumen Mitocondrial y Lipídico. Después del entrenamiento, la densidad del volumen mitocondrial fue un 9% mayor en la pierna E y un 15% mayor en la pierna E±F, pero los cambios no fueron estadísticamente significativos (Figura 9). Los grandes aumentos (E, 120%; E+F, 169%) en la densidad del volumen lipídico tampoco alcanzaron significancia estadística (p = 0.063) (Figura 9).
Fuerza Voluntaria. La fuerza en 1RM en el ejercicio de prensa de piernas aumentó significativamente (p<0.001) después del entrenamiento (Figura 1), observándose una interacción significativa (p<0.001). A pesar de que tanto la pierna E (20.3 %) como la pierna E+F (34.1 %) aumentaron significativamente su fuerza en 1RM, el aumento en la pierna E+F fue significativamente mayor.
Área de Sección Cruzada Muscular. El área de sección cruzada de los extensores de la rodilla aumentó significativamente (p<0.001) tanto en la pierna E (14.3%) como en la pierna E+F (20.9%) (Figura 2). El mayor aumento en la pierna E+F no fue significativo (interacción p = 0.13). El área de sección cruzada de los flexores de la rodilla aumentó (p = 0.012) tanto en E (8.8%) como en E+F (12.4%). El mayor aumento en la pierna E+F no fue significativo (interacción p = 0.126). El cociente entre las áreas de los extensores/flexores no cambió significativamente después del entrenamiento.
Perímetro del Muslo, Pliegues Cutáneos, y Peso Corporal. El perímetro del muslo aumentó leve pero significativamente (p = 0.001) después del entrenamiento. El incremento en la pierna E+F (4.8%) fue significativamente mayor (interacción, p=0.037) que el aumento en E (3.4%) (Figura 3). La suma de cuatro pliegues cutáneos no cambió significativamente luego del entrenamiento (Figura 3). Hubo un pequeño (2.6%) pero significativo aumento (p<0.05) en el peso corporal después del entrenamiento (Tabla 1).
Superficie de las Fibras Musculares. No hubo cambios significativos en la superficie de las fibras ST (E, 4864 ± 1318 a 4786 ± 375; E+F, 5274 ± 695 a 5347 ± 367) o FT (E, 5590 ± 687 a 5543 ± 291; E+F, 5690 ± 555 a 6299 ± 457). La relación del área FT/ST no cambió (E, 1.14 ± 0.08 a 1.17 ± 0.07; E+F, 1.12 ± 0.06 a 1.18 ± 0.05).
DISCUSION
El entrenamiento nominalmente designado como de fuerza (F) y resistencia (E) utilizado en el presente estudio tuvo más estímulos en común que diferencias; por lo tanto, en el grupo A, la pierna F tuvo incrementos similares a la pierna F+E en las mediciones de resistencia relacionadas con el VO2 máx, en el número de repeticiones con la carga igual a 1RM pre-entrenamiento en el ejercicio de prensa de piernas, y en el % de fibras ST; y F tuvo incrementos significativos aunque menores (vs F+E) en las repeticiones al 80 % de 1RM, y en la actividad de la CS. De manera similar, en el grupo B la pierna E exhibió un incremento en el área de sección cruzada de los extensores de la rodilla, que fue similar al de la pierna E+F, y tuvo aumentos significativos, aunque menores, en la fuerza en 1RM en el ejercicio de prensa de piernas y en el perímetro de muslo. Por lo tanto, el entrenamiento de F y E utilizado en el presente estudio podría ser considerado como un híbrido más que formas extremas o “puras” de entrenamiento de fuerza y resistencia.
La naturaleza híbrida del entrenamiento de E probablemente se debió en parte a la alta intensidad de las series de (3 min al 90-100 % del VO2 máx) utilizadas; dichas series podrían asegurar adaptaciones características del entrenamiento de la resistencia, en las unidades motoras lentas (8, 27), pero también indujeron incrementos en el tamaño y en la fuerza muscular. Además, el entrenamiento de E en ciclismo (necesario en el presente estudio debido al modelo de entrenamiento con una pierna) podría probablemente incrementar más la fuerza (31, 34) y el tamaño muscular (2, 12; ver sin embargo, 39) que, por ejemplo, el pedestrismo. El entrenamiento de F comprendió series de 15-20 repeticiones con un peso que varió desde entre el 75-80% de 1RM al comienzo del entrenamiento y el 85-90 % de 1RM hacia el final [por comparación solo se pueden realizar 5-10 repeticiones con el 75-90 % de 1 RM en ejercicios para el tronco superior tales como el press de banca (35) y flexión de codos (39)]. A pesar de que la intensidad (% 1RM) fue adecuada para un entrenamiento efectivo de la fuerza, el número de repeticiones por serie (15-20) y por sesión (90-120) podría ser considerado elevado (hubo 900 contracciones en cada sesión de entrenamiento de sobrecarga). El programa de F fue adecuado para aumentar la fuerza y el tamaño muscular, pero también aumentó el VO2 máx (quizás debido a un efecto “central” a partir del entrenamiento de resistencia de la otra pierna), la resistencia durante el levantamiento de pesas, y la actividad de la CS.
Cuando se realizan en forma concurrente el entrenamiento de F y E estos podrían no interactuar del todo, por ejemplo, el entrenamiento concurrente podría causar las mismas adaptaciones en fuerza y resistencia de lo que lo harían el entrenamiento de F o de E si se realizaran por separado. Asimismo estos tipos de entrenamiento podrían interactuar de forma antagónica; con lo cual, las adaptaciones al entrenamiento de la fuerza y/o de la resistencia serían menores que en respuesta al entrenamiento solo de F o de E. O podrían interactuar en forma aditiva, con lo cual, las adaptaciones en fuerza y resistencia serían mayores que en respuesta al entrenamiento solo de F o de E. La interacción que ocurra y la forma que tome probablemente dependa de varios factores: intensidad, volumen, y frecuencia de los dos tipos de entrenamiento; modos de entrenamiento, nivel de entrenamiento de los sujetos; y la forma en que los dos entrenamientos se integran. Además, los resultados y conclusiones a las que hemos arribado estarán afectadas por las mediciones seleccionadas del desarrollo de la fuerza y la resistencia. Estos factores deben ser considerados cuando se interpreta el presente estudio y los realizados previamente (3, 9, 10, 14, 15, 19).
En el presente estudio no se observaron efectos antagónicos, probablemente debido a la naturaleza híbrida del entrenamiento de F y E, y a que el volumen total fue moderado. Sin embargo, estudios previos han mostrado efectos antagónicos en la forma de desarrollo desigual de la fuerza (9, 14, 19). En estos estudios el antagonismo podría haber ocurrido debido al mayor volumen de entrenamiento utilizado (14), o a la variable de adaptación utilizada [e. g., desarrollo desigual de la fuerza isocinética de alta velocidad pero no de baja velocidad (9)]. Por lo general, es más probable que ocurra un antagonismo cuando se realizan en forma concurrente formas extremas de gran volumen de entrenamiento de F y E (e.g., entrenamiento para maratón y levantamiento competitivo de pesas).
Con respecto a un posible efecto aditivo del entrenamiento combinado de F y E, se observó un patrón interesante. Cuando un estímulo presuntamente más fuerte para una adaptación determinada fue agregado a un estímulo más débil, en ocasiones pudo observarse un efecto aditivo, pero cuando un estímulo más débil fue agregado a uno más fuerte, no se observó el efecto de adición. Por lo tanto, en el grupo A (F vs F±E) el agregado del entrenamiento de E fue aditivo para respecto del incremento en la resistencia (repeticiones al 80% de 1RM) y en la actividad de la CS, a pesar de que el entrenamiento solo de F causó incrementos significativos en estas variables. De forma similar, en el grupo B (E vs E+F) el agregado del entrenamiento de F fue aditivo para incrementar la fuerza y una de las variables del tamaño muscular (perímetro de muslo), a pesar de que el entrenamiento solo de E provocó aumentos significativos en estas variables. Por el contrario, el entrenamiento de E no fue aditivo para aumentar la fuerza y el tamaño muscular (F vs F+E), si bien el entrenamiento solo de E causó incrementos en estas variables, ni el trabajo de F fue aditivo para aumentar el VO2 máx y la actividad de la CS (E vs E+F), aunque el trabajo solo de F produjo aumentos en estas variables. Una excepción fue que el entrenamiento de F fue aditivo para la resistencia de corta duración. Los tests de resistencia fueron realizados en la máquina en la cual se realizó el entrenamiento de F; por lo tanto, la mayor resistencia probablemente estuvo relacionada con un aumento en la fuerza, la destreza, y la eficiencia en el movimiento del ejercicio durante el transcurso del entrenamiento de F.
El nivel de entrenamiento de los sujetos puede determinar si va a producir un efecto antagónico o un efecto aditivo con el entrenamiento concurrente de F y E. En sujetos previamente desentrenados, el desarrollo de la resistencia no se ve perjudicado (9, 14, 19, y el presente estudio) e incluso podría provocar una mejora (presente estudio); en cambio el desarrollo de la fuerza podría (9, 14, 19) o no (presente estudio) verse perjudicado. En sujetos previamente entrenados en resistencia, que siguen con el entrenamiento pero que adicionan entrenamientos de fuerza, el desarrollo de la fuerza (19) y la resistencia (15) puede aumentar.
Para el presente estudio elegimos un modelo de entrenamiento que permitió que cada sujeto actuara a la vez como control, lo que al mismo tiempo permitió la evaluación de la interacción entre el entrenamiento concurrente de la fuerza y la resistencia. Por lo tanto, el grupo A, entrenó tanto la fuerza como en resistencia con una de las piernas, y en fuerza solamente con la otra, mientras que el grupo B entrenó la fuerza y resistencia con una de las piernas y la resistencia solamente con la otra. Este modelo evitó la variabilidad en la respuesta al entrenamiento entre grupos separados formados por un pequeño número de sujetos. Por ejemplo, un grupo particular, a pesar de la asignación aleatoria de sujetos, podría por casualidad contener más sujetos entrenables. Este grupo tendría una mayor respuesta al entrenamiento, independientemente del método de entrenamiento. Este problema se puede minimizar con grupos más grandes. El modelo utilizado en el presente estudio tuvo al menos una limitación. El modelo fue elegido con la suposición de que la mayor parte de cualquier interacción (antagonismo o adición) causada por el entrenamiento concurrente de fuerza y resistencia debía expresarse al nivel periférico en variables tales como el área de sección cruzada muscular y la actividad enzimática. Si ocurriese antagonismo o adición pero se expresaran enteramente como un fenómeno “central”, entonces en nuestros sujetos aún las piernas que solo hicieron entrenamiento de fuerza o de resistencia habrían sido afectadas de la misma manera que las piernas que combinaron entrenamiento de F y E. El antagonismo o la adición no se hubiesen revelado. En un estudio llevado a cabo por Hickson (14), hubo datos que indicaron que el antagonismo podría en algunos casos ser enteramente central (e.g., fatiga del sistema nervioso central); el desarrollo de la fuerza voluntaria se vio perjudicado por el entrenamiento concurrente de F y E, pero los incrementos en el tamaño muscular (indicados por las mediciones de los perímetros del muslo) no se vieron afectados. En comparación, nosotros no observamos un impedimento en el desarrollo de la fuerza o del tamaño muscular, pero no podemos excluir la posibilidad de que haya ocurrido en nuestro estudio algún tipo de efecto antagónico mediado enteramente por el sistema central. Sin embargo, se puede concluir que no ocurrió ningún efecto antagónico a nivel periférico (muscular).
En conclusión, un resultado del presente estudio que no concuerda directamente con el propósito principal merece, sin embargo, un comentario. El porcentaje de fibras ST en el vasto lateral aumentó después de cinco meses de entrenamiento. El aumento o el cambio en el % de fibras ST después del entrenamiento de F o E en los seres humanos es poco común, pero tiene precedentes (21, 38, 40). El desentrenamiento ha sido asociado con una disminución en el % de fibras ST (23). Cualquier cambio observado en la composición del tipo de fibra sugiere el tema de errores metodológicos asociados con la determinación del % de fibras ST a partir de biopsias de grandes músculos. Estos errores (referencias 4, 24) y sus consecuencias en la interpretación de los resultados, acerca de los cambios en el % de fibras ST, han sido discutidos recientemente en otros trabajos (23, 38). En el presente estudio las biopsias pre y post-entrenamiento no fueron analizadas de una sola vez sino con 5 meses de separación, sugiriendo la posibilidad de que una diferencia en el pH o en la temperatura del medio pre-incubación en los dos análisis hubiese podido causar un error sistemático en el teñido y la determinación posterior del % de fibras ST. Sin embargo, esta posibilidad está disminuida por la consideración de que sería necesario una diferencia de al menos varias décimas de unidad de pH para afectar la diferenciación de los principales tipos de fibras (ST vs FT). De cualquier manera, los cambios que observamos en el % de fibras ST deberían ser considerado con cuidado.
Referencias
1. ANDERSEN, P (1975). Capillary density in skeletal muscle of man. Acta Physiol. Scand. 95: 203-205
2. ANDERSEN, P. and J. HENRIKSSON (1977). Capillary supply to the quadriceps femoris muscle of man. Adaptive response to exercise. J. Physiol Lond 270:677
3. BELL, G.J., S.R. PETERSEN H.A. QUINNEY, and H.A. WENGER (1988). Sequencing of endurance and high-velocity strength training. Can. J.Sport Sci. 12:214-219
4. BLOMSTRAND, E., and B. EKBLOM (1982). The needle biopsy technique for fiber type determination in human skeletal muscle-a methodological study. Acta Physiol. Scand. 116: 437-442
5. COSTILL, D.L (1967). The relationship between selected physiological variables and distance running performance. J. Sports Med. Phys. Fitness 7:61-6
6. COSTILL, D.L., E.F. COYLE, W. F.FINK, G.R.LESMES, and F. A WITZMANN (1997). Adaptations in skeletal muscle following strength training. J.Appl Physiol 4G:96-99
7. DENIS. C., J.,-C. CHATARD, D. DORMOIS, E.T. LINOSSIER, A. GEYSSANT, and J.R. LACOUR (1986). Effects of endurance training on capillary supply of human skeletal muscle on two age groups (20 and 60 years). J. Physiol. París 81: 379-383
8. DUDLEY, G. A., W. M. ABRAHAM, and R. L. TERJUNG (1982). Influence of exercise intensity and duration on biochemical adaptations in skeletal muscle. J. Apple Physial.53: 844-850
9. DUDLEY, G. A., and R. DJAMIL (1985). Incompatibility of endurance- and strength- training modes of exercise. J. Apply Physial.59:1446-1451
10. DUDLEY, G. A., and S.J. FLECK (1987). Strength and endurance training. Are thy mutually exclusive?. Sports Med. 4: 79-85
11. ESSEN, B., A. LINDHOLM, and G. THORTON (1980). Histochemical properties of muscle fibre types and enzyme activities in skeletal muscle of standard- bred trotters of different ages. Equine Vet. J. 12: 175-180
12. GOLLNICK, P. D., R. B. ARMSTRONG, B. SALTIN C., W. SAUBERT. IV, W. L. SEMBROWICH, and R. E. SHEPHERD (1973). Effects of training on enzyme activity and fiber composition of human skeletal muscle. J. Apple Physial.34:107-111
13. GUTH, L., and F.J. SAMAHA (1969). Qualitative differences between actomyosin ATPase of slow and fast mammalian muscle. Exp. Neurol.25:138-152
14. HICHSON. R.C (1980). Interference of strength development by simultaneously training for strength and endurance. Eur J. Appl. Physiol.Occup.Physiol.45:255-263
15. HICKSON, R.C., B.A. DVORAK, E. M. GOROSTIAGA, T.T. KUROWSKI, and C. FOSTER (1988). Potential for strength and endurance training to amplify endurance performance. J Appl Physiol.,65:22852290
16. HICKSON, R.C., M.A. ROSENKOETTER, and M.M BROWN (1980). Strength training effects on aerobic power and short term endurance. Med. Sci. Sports Exercise 12:336-339
17. HOPPELER, H (1986). Exercise-induced ultrastructural changes in skeletal muscle. Int J. Sports Med.7:187-204
18. HOPPELER, H., P LUTHI, H. CLASSEN, E.R. WEIBEL, AND H. HOWALD (1973). The ultraestructure of the normal skeletal muscle: a morphometric analysis of untrained men, women and well-trained orinteers. Pfluegers Arch 344:217-232
19. HUNTER, G., R. DEMMENT, and D. MILLER (1987). Development of strength and maximum oxygen uptake during simultaneous training for strength and endurance and endurance. J. Sports Med. Phys. Fitness 27:269-275
20. INGJER, F (1979). Effects of endurance training on muscle fibre ATP-ase activity, capillary supply and mitochondrial content in man. J. Physiol.Lond. 294:419-432
21. JANSSON, E., B. SJODIN, and P. TESCH (1978). Changes in muscle fibre type distribution after physical training. A sign of fibre type transformation?. Acta Physiol. Scand. 104:235-237
22. KLAUSEN, K., L.B. ANDERSON, and I. PELLE (1981). Adaptive changes in work capacity, skeletal muscle capillarization and enzyme levels during training and detraining. Acta Physiol. Scand 113:9-16
23. LARSSON, L., and T. ANSVED (1985). Effects of long-term physical training and detraining on enzyme histochemical and functional skeletal muscle characteristics in man. Muscle Nerve 8:714-722
24. LEXELL, J., K.HENRIKSON-LARSEN, and M. SJOSTROM (1983). Distribution of different fibre types in human skeletal muscles. A study of cross-sections of whole m. vastus lateralis. Acta Physiol. Scand. 117: 115-122
25. LOWRY, O. H., and J. V. PASSONNEAU (1972). A flexible System of enzymatic analysis. New York: Academic
26. MAC DOUGALL, J.D., G.C.B. ELDER, D.G. SALE, J. R. MOROZ, and J. R. SUTTON (1980). Effects of strength training and immobilization on human muscle fibres. Eur. J. Apple Physiol. Occup Physiol.43:25.34
27. MAC DOUGALL, D., and D. SALE (1981). Continuous vs. Interval training: a review for the coach and athlete. Can. j. Appl. Sport Sci. 6:93-97
28. MAC DOUGALL, J.D., G. C. B. ELDER, D.G. SALE, and J. R. SUTTON (1982). Muscle ultrastuctural characteristics of elite power lifters and body-builders. Eur. J. Appl Physiol. Occup Physiol. 48: 117-126
29. MACDOUGAL, J. D., D. G. SALE, J. R. MOROZ, G.C.B. ELDER, J. R. SUTTON, and H. HOWALD (1979). Mitochondrial volume density in human skeletal muscle following heavy resistance training. Med. Sci. Sports 11:164-166
30. MAUGHAN, R. J. HARMON, S. B. LEIPER, D. SALE, and A. DELMAN (1986). Endurance capacity of untrained males and females in isometric and dynamic muscular contractions. Eur. J. Appl Physiol.Occup.Physiol. 55:395-400
31. MOROZ, D. E. And M.E. HUSTON (1987). The effects of replacing endurance running training with cycling in female runners. Can.J. Sport Sci. 12:131-135
32. NOVIKOFF, A.B., W.Y. SHIN, and J. DRUCKER (1961). Mitochondrial localization of oxidation enzymes: staining results with two tetrazolium salts. J. Biophys, Biochem.Cytol. 9: 47-61
33. ONO, M., M. MIYASHITA, and T. ASAMI (1976). Inhibitory effect of long distance running training on the vertical jump and other performance among aged males. n: Biomechanics V-B, edited by P.V. Komi, Baltimore, MD: University Park, p. 94-100
34. ROSLER, K., K. E. CONLEV, H HOWALD, C.GERBER, and H.HOPPELER (1986). Specificity of leg power changes to velocities used in bicycle endurance training. J. Appl Physiol. 61:30-36
35. SALE, D., and D. MACDOUGALL (1981). Specificity in strength training: a review for the coach and athlete. Can. J. Appl. Sport Sci. 6:87-92
36. SCHANTZ, P (1983). Capillary supply in heavy-resistance trained non postural human skeletal muscle. Acta thysiol. Scand. 117: 153-155
37. SCHANTZ, P. G (1986). Plasticity of human skeletal muscle with special reference to effects of physical training on enzyme levels of the NADH shuttles and phenotypic expression of slow and fast isoforms of miofibrilar proteins (Abstract). Acta Physiol. Scand. Suppl. 128:558
38. SIMONEAU, J.-A., G. LORTIE, M. R. BOWLAY, M. MARCOTTE, M.C.THIBAULT, and C.BOUCHARD (1985). Human skeletal muscle fiber type alteration with high-intensity intermittent training. Eur. J. Appl.Physiol. Occup. Physiol 54:250-253
39. TERRADOS, N., J. MELICHNA, C. SYLVEN, and E. JANSSON (1986). Decrease in skeletal muscle myoglobin with intensive training in man. Acta Physiol. Scan. 128:651-652
40. TESCH, P. A., and J. KARLSSON (1985). Muscle fiber types and size in trained and untrained muscle of elite athletes. J. Appl. Physiol 59:1716-1720
41. TESCH, P.A., A. THORSSON, and P. KAISER (1984). Muscle capillary supply and fiber type characteristics in weight and power lifters. J. Appl. Physiol. 56:35-38
42. WEIBEL, E. B (1979). Stereological methods. In: Practical Methods for Biological Morphometry. London: Academic, vol.1