
La Eficiencia en el Ciclismo Está Relacionada con el Porcentaje de Fibras Musculares de Tipo I
Edward F Coyle1, L. S Sidossis1, J. F Horowitz1 y John D Beltz1
The Human Performance Laboratory, Department of Kinesiology and Health Education, The University of Texas at Austin, Austin, TX 78712.
Artículo publicado en el journal PubliCE, Volumen 0 del año 1994.
Publicado 21 de abril de 2006
Resumen
Palabras clave: miosina, fibras rápidas, fibras lentas, energética muscular, contracción muscular, ejercicio físico
INTRODUCCION
Nosotros hemos observado recientemente que el consumo de oxígeno (VO2) durante el ejercicio submáximo en bicicleta a una tasa de esfuerzo determinada (por ej., economía de pedaleo), puede variar hasta en un 15% en un grupo de ciclistas competitivos bien entrenados (5). Estas diferencias podrían no ser explicadas por las diferencias en la técnica de ciclismo, ya que no hay relación entre la economía de pedaleo (por ej., W.l-1 VO2.min-1; ver Tabla 3 de la ref. 5) y los índices biomecánicos de efectividad, definidos como el porcentaje de fuerza aplicada al pedal, la que realmente crea una fuerza propulsiva a través de la revolución del pedal (ver IE 180 IE 360 en la Tabla 6 de la ref. 5). Si no es la técnica, pareciera entonces que las diferencias en la economía de pedaleo podrían estar relacionadas a factores musculares que influyen la eficiencia de la contracción. De hecho, la correlación de los datos presentados en nuestro estudio previo, sugiere que existe una relación entre economía de pedaleo y % de fibras musculares de Tipo I.
En un intento de determinar más directamente si el % de fibras de Tipo I influye en la eficiencia muscular en los deportistas, nosotros estudiamos otro grupo (n=19) de ciclistas altamente entrenados que tenían un máximo consumo de oxígeno (VO2 máx.) similar, y también eran homogéneos en la capacidad para pedalear en steady-state a altas tasas de esfuerzo. Sin embargo, estos sujetos tenían un amplio rango de % de fibras musculares de Tipo I en el vasto lateral (32-76%). Además de evaluar la eficiencia cuando estaban pedaleando (a 80 RPM), también se realizaron mediciones mientras realizaban una extensión de rodillas con las dos piernas juntas. Ambos tipos de ejercicios provocaban velocidades en la extensión de rodillas de aproximadamente 200º. s-1, que es una velocidad relativamente baja, con la cual nosotros pudimos predecir la velocidad de eficiencia pico de las fibras de Tipo I (p. ej., 1/3 Vmáx.) (8,15).
La clasificación de las fibras musculares lentas (Tipo I) o rápidas (Tipo II) está basada generalmente por el análisis químico de la actividad de la miosina adenosina trifosfatasa (ATPasa) (2), la que se vio que está relacionada con la composición de la cadena pesada de miosina. Esta composición de la cadena de miosina determina, en gran medida, las velocidades máximas del acortamiento del sarcómero (1,15), aunque las cadenas livianas 1 y 3 de miosina rápida también están relacionadas con la velocidad de acortamiento (20). La velocidad máxima de acortamiento del sarcómero (Vmáx.) de las fibras humanas de Tipo II, es aproximadamente, 3-5 veces mayor que las de Tipo I (8). La eficiencia muscular, definida como la cantidad de trabajo realizada por la fibra muscular en relación a su gasto energético, se ha observado que es la más alta a velocidades de acortamiento, de aproximadamente un tercio de Vmáx., tanto en fibras del Tipo I como II (11,15). Por lo tanto, cuando las contracciones son realizadas isométricamente o a velocidades relativamente bajas, las fibras musculares de Tipo I, en mamíferos, son más eficientes que las de Tipo II (11, 15, 22, 23).
A pesar de esta fuerte base teórica, pocos estudios han intentado determinar si las diferencias en el % de fibras musculares de Tipo I, observadas entre deportistas, influyen en la eficiencia muscular durante el ejercicio (19).
Para tratar de responder esta pregunta, nosotros estimamos la eficiencia a partir de la evaluación del VO2 y del cociente respiratorio (RER), durante esfuerzos en ciclismo en estado de equilibrio (steady-state), a intensidades conocidas. Esta medición de la eficiencia de ciclismo reflejará mejor la eficiencia muscular de ciclistas bien entrenados, capaces de ejercitar en steady-state a altas intensidades, que es cuando los músculos utilizan el mayor % del VO2 total del cuerpo.
METODOS
Diseño general
Los sujetos fueron 19 ciclistas competitivos. Primero se midió el VO2 máx. y la intensidad a la cual se provocaba el umbral de lactato (UL). Luego se determinó la eficiencia muscular durante las 2-3 semanas subsiguientes, como se describe a continuación. Las biopsias musculares fueron generalmente obtenidas dentro de las 3 semanas antes o después de los tests de esfuerzo. La eficiencia total (ET) y la eficiencia delta (ED), como se definen más adelante, fueron determinadas en los 19 sujetos. Además, se evaluó la ET durante la extensión de rodillas, en 13 de los 19 ciclistas. Este estudio fue aprobado por el Comité Institucional de Revisión de la Universidad de Texas, y los sujetos dieron su consentimiento por escrito.
Determinación del % de fibras musculares de Tipo I
Para mejorar la precisión en el cálculo del % de fibras de Tipo I (7), se realizaron varias biopsias en vasto lateral en ambos muslos, en la mayoría de los sujetos. A diez participantes se les realizaron biopsias en cuatro oportunidades, a cuatro sujetos en tres ocasiones, y a cinco, sólo una vez. El % de fibras de Tipo I se determinó sumando todas las fibras musculares de este Tipo, obtenidas en las múltiples biopsias, en relación con el número total de fibras obtenidas. Las incisiones en la piel se realizaron a 12-20 cm por encima de la rótula, y se trató, intencionalmente, de variar las zonas del vasto lateral, en las cuales se obtenía las biopsias. Esto se realizó insertando la aguja a distintas profundidades y alterando la posición medial/lateral de la aguja en el músculo. Las muestras musculares, preparadas para el análisis histoquímico, fueron seccionadas transversalmente (10ºm) a -20ºC, usando un micrótomo Cryostat. La distribución de las fibras de Tipo I y de Tipo IIa y IIb fue determinada a partir del análisis de adenosina trifosfatasa a un pH de 4,3; 4,55; y 10,3 (2).
Medición del VO2 máx., VO2, y RER
Para todos los tests se utilizó una bicicleta ergométrica Monark (modelo 819), equipada con un asiento de carrera, manubrio bajo, y pedales para zapatillas con tapones. El VO2 máx. fue medido durante ciclismo continuo, en pruebas de 8' a 10', con aumento de intensidad cada 2'. El UL fue medido como se describió en un estudio anterior (5), y se definió como la intensidad que provocaba un aumento de 1 mM en la concentración de lactato sanguíneo, por encima de la línea basal (por ej., concentración de lactato sanguíneo durante el ejercicio, al 50-60% del VO2 máx.).
Durante todos los tests, los sujetos respiraron a través de una válvula de Daniels; los gases espirados eran continuamente extraídos y analizados para determinar la concentración de O2 (Applied Electrochemistry S3A) y de CO2 (Beckman LB-2). Los volúmenes de aire inspirado también fueron medidos (Parkinson-Cowan CD4). Estos instrumentos fueron conectados a una computadora Apple IIe, que calculaba el VO2 y el RER (VCO2/VO2), cada 30 segundos.
Determinación de la eficiencia, cuando las tasas de esfuerzo eran menores a las del umbral de lactato
Los sujetos (n=19), para quienes el UL se producía a intensidades mayores al 70% del VO2 máx., pedalearon en forma continua a tasas de esfuerzos de aproximadamente el 50,60, y 70% del VO2 máx. durante 5 minutos cada una. Diez de los 19 sujetos, que tenían UL por encima del 80%, también fueron evaluados en el cuarto nivel de esfuerzo (75% del VO2 máx.), pero debajo de la intensidad que determinaba el umbral de lactato. Para establecer los niveles de esfuerzo, el sujeto debía mantener una velocidad de 80 RPM (siempre entre 78 y 82 RPM) y se fijaba la resistencia. La velocidad de extensión de rodillas cuando se pedalea a 80 RPM es de aproximadamente 200º. s-1 (74º de extensión de rodilla en 0,37 segundos). La tasa de esfuerzo fue calculada monitoreando las RPM (± 1 RPM en un contador electrónico) y las resistencias (± 1 NM de un potenciómetro calibrado adherido al péndulo).
Se calculó el promedio de VO2 y RER durante los 3-5 minutos de cada nivel. Simultáneamente se obtuvieron muestras de sangre durante el 5to minuto de ejercicio en cada nivel, a través de un catéter en la vena antecubital, y la concentración de lactato fue determinada, como se describió previamente (5).
La eficiencia total (ET) fue calculada como la proporción entre el trabajo realizado.min-1 (watts convertidos en Kcal.min-1) y la energía gastada.min-1 (kcal.min-1). El gasto energético.min-1 (kcal.min-1) fue calculado del VO2 y del RER, usando las tablas de Lusk (16). La ET de cada sujeto fue similar en todos los niveles de esfuerzo cuando se pedaleaba a 80 RPM. Por lo tanto, ET fue calculada como la ET promedio de tres de los cuatro niveles de esfuerzo.
La eficiencia delta (ED) se definió como la proporción entre el cambio en el trabajo realizado.min-1, y el cambio en la energía gastada.min-1. La ED fue calculada para cada sujeto a partir de una regresión lineal (y= mx + b) de la relación entre la energía gastada.min-1, como variable y: kcal.min-1 (ej. 3 o 4 puntos de datos, aproximadamente al 50, 60, 70 y 80% del VO2 máx..; considerar el último nivel de esfuerzo en sujetos con un alto UL) vs. trabajo realizado, (min-1, como variable x: kcal.min-1). La ED fue calculada a partir de la pendiente de la relación y fue igual al recíproco de m (l/m). A pesar de que tanto ET como ED miden la eficiencia, ED refleja de una manera más válida la eficiencia real de la contracción muscular (9).
Eficiencia durante la extensión de rodillas con las dos piernas
En 13 ciclistas también se midió la ET mientras realizaban una extensión de rodillas con las dos piernas. El propósito de este procedimiento era determinar si la eficiencia estaba relacionada con el % de fibras musculares de Tipo I, durante un ejercicio que principalmente recluta sólo a los cuádriceps, ya que el % de fibras de Tipo I fue determinado en el vasto lateral. Nosotros pensamos que si el % de fibras de Tipo I influyen en la eficiencia muscular, esto no debería ser sólo específico del ciclismo.
Los sujetos se sentaron con un ergómetro para extensión de rodillas (Eagle Cybex Inc.). Los movimientos del tronco y la cadera fueron bloqueados utilizando un cinturón fuertemente ajustado a través de las crestas ilíacas. La extensión de rodillas se realizó simultáneamente con las 2 piernas. Los sujetos debían levantar un brazo del ergómetro que hacía contacto con ellos en la parte media de la tibia. El eje rotativo de las rodillas fue posicionado al punto de pivote del brazo ergométrico. El otro extremo del mismo estaba adherido a un cable que estaba directamente alrededor de una roldana y atado a una carga de pesas. El hecho de levantar el brazo del ergómetro con la extensión de rodillas, provocaba que las pesas se eleven una distancia vertical. El trabajo realizado.min-1, fue calculado como el producto del peso (fuerza) y la distancia total hacia arriba, recorrida en 60 segundos (sumando las distancias de todas las repeticiones en ese tiempo). El desplazamiento vertical fue calculado con una presición de ± 1%, usando un potenciómetro adherido al centro del brazo de palanca y conectado a una computadora IBM AT. También se determinaron las velocidades angulares del brazo ergométrico.
El ángulo de la rodilla, al comienzo de la extensión, era de 75º (ángulo aproximado entre la tibia y el fémur). La velocidad de la extensión aumentaba constantemente durante los primeros 20-25º hasta alcanzar una velocidad pico promedio de 249 ± 7º. s-1. La velocidad promedio de la extensión de rodilla fue 177±6º. s-1, a la cual nosotros consideramos aproximadamente 200º. s-1. Los sujetos detenían el movimiento entre un 10 al 15º antes de la extensión completa, y dejaban flexionar pasivamente sus piernas para volver al punto de partida, antes de realizar otra extensión. Ellos mantuvieron una frecuencia de 50 extensiones.min-1 y un desplazamiento vertical bastante uniforme (50 cm).
Se determinó el pico de VO2 durante este test de extensión de rodilla, utilizando un protocolo de incremento discontinuo de los niveles de esfuerzo. En otra ocasión, los sujetos realizaron este test con niveles de esfuerzo calculados para provocar, aproximadamente, entre el 50 y el 70% del pico de VO2. La concentración de lactato sanguíneo a estas intensidades, no aumentó significativamente, por encima de los niveles de reposo. Cada tasa de esfuerzo fue realizada durante 5 minutos, y al menos con 10 minutos de descanso entre cada serie. Se levantaron pesos de aproximadamente 16-22 kg. Esto representó el 15±1% y el 19±1%, respectivamente, de una repetición máxima medida durante la extensión de rodilla con dos piernas. Se sacó un promedio del VO2 y del RER durante el período de 3-5 minutos, y se calculó el gasto energético.min-1. La ET fue también calculada, como se describió anteriormente.
Análisis Estadísticos
Se utilizó la fórmula de momento.producto para calcular los coeficientes de correlación y se estableció una significancia estadística de P<0,05 (17). También se realizaron correlaciones parciales entre los años de entrenamiento de resistencia (variable x) y ET y ED (variable y), considerando los efectos de la influencia del % de fibras de Tipo I, manteniéndose constante los ejes de x e y (17; p.116).
RESULTADOS
Sujetos y Entrenamiento
Las características físicas de los 19 ciclistas competitivos se presentan en la Tabla 1. Estos hombres han estado entrenando la resistencia por un promedio (± SE) de 5,2 ± 0,8 años (rango 2-12 años), y han practicado ciclismo por 3,5 ± 0,4 años (rango 1,5-7 años). Tuvieron un VO2 máx. promedio de 4.9±0.11.min-1. Un promedio de 3,2 ± 0,4 biopsias musculares fueron realizadas y se obtuvieron 1860 ± 366 fibras para determinar el % de fibras musculares de Tipo I.

Tabla 1. Tipos de fibra muscular y características físicas de los 19
ciclistas. Los valores son presentados como medias±SE.
Niveles de esfuerzo en steady-state en ciclismo
El UL se produjo a un promedio del 77.6º±0.3% VO2 máx. (rango 73-84% VO2 máx.). El gasto calórico fue medido durante el pedaleo en steady-state a intensidades por debajo del UL, las que tuvieron un promedio de 52.3±0,7%, 61.2±0.8%; y 71.0±0.7% del VO2 máx. para los 19 sujetos; y durante una 4ta. intensidad que provocó el 79.6 ± 0.5% del VO2 máx., en 10 sujetos con un alto UL. Con relación al promedio de los niveles de esfuerzo usados durante estos tests, el 50% del VO2 máx. fue alcanzado pedaleando a una intensidad de aproximadamente 180 W, mientras que el 80% del VO2 máx. requirió un esfuerzo cercano a los 300 W, teniendo en cuenta, obviamente, que estas relaciones variaron con las diferencias en la eficiencia. Además, la concentración de lactato sanguíneo estuvo en steady-state y no se elevó por encima de los valores de reposo, durante la determinación de la eficiencia de ciclismo. Por otro lado, el VO2 alcanzó un steady-state durante el período de 3-5 minutos de ejercicio en estos deportistas de resistencia, y basados en una observación nuestra no publicada, creemos que extender el período a 10 minutos en cada intensidad, no provocará mayores aumentos en el VO2. Esto está de acuerdo con un informe de Weltman y cols. (21).
Eficiencia total del ciclismo y % de fibras musculares de Tipo I
Estos ciclistas altamente entrenados tenían un promedio de 56±3% de fibras musculares Tipo I. La mayoría tenía <2% de fibras del Tipo IIb; sólo tres ciclistas tenían 10% o más de estas fibras. El % de fibras de Tipo I varió entre 32 y 76%, y la ET cuando pedaleaban a 80 RPM, varió desde 18.3 a 22.6% (Tabla 2). Como se muestra en la Figura 1 A, la ET estuvo significativamente correlacionada con el % de fibras musculares de Tipo I (r= 0.65; p<0.001).
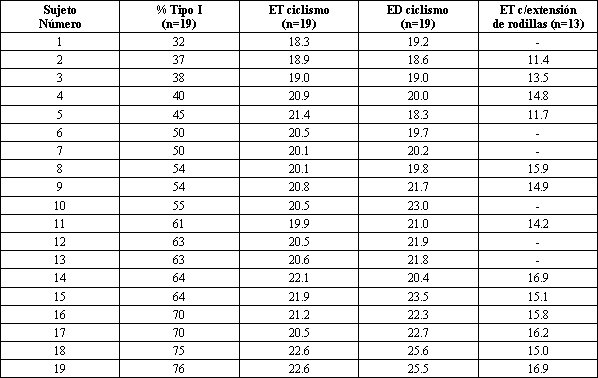
Tabla 2. Datos de cada sujeto en relación al porcentaje de fibras de
Tipo I, a la eficiencia total (ET), y a la eficiencia delta (ED) en ciclismo,
así como a la eficiencia total (ET), durante el test de extensión de rodillas.
Eficiencia delta en ciclismo y % de fibras musculares de Tipo I
La ED varió desde 18.3 a 25.6% en estos sujetos (Tabla 2). Como se muestra en la Figura 1 B, la ED también estuvo altamente correlacionada con el % de fibras Tipo I (r= 0.85; p<0,001). La extrapolación de esta relación predice que una persona con todas las del Tipo II (0% de Tipo I), tendría una ED de 13,5%. Por el contrario, una persona con el 100% de fibras de Tipo I tendría una ED de 27,4%, siendo aproximadamente el doble.
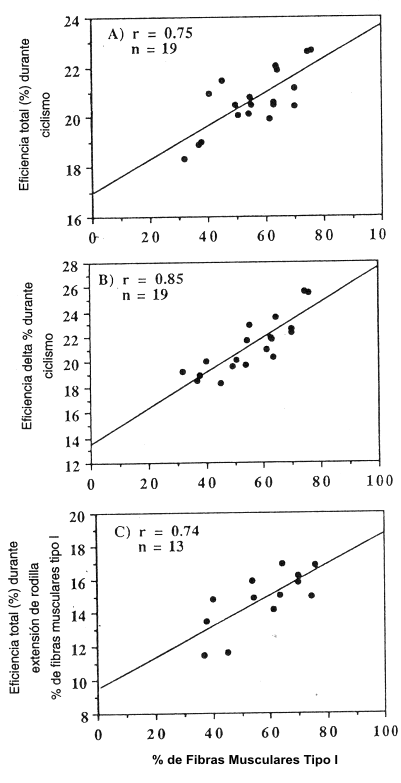
Figura 1. La relación entre el % de fibras musculares de Tipo I y la
eficiencia en ciclismo, medida de distintas formas. A) Eficiencia total (ET) con
una velocidad de pedaleo de 80 RPM, determinada como el cociente entre el
trabajo realizado.min-1 y la energía gastada.min-1.
Ecuación: ET= 0.0671 * (% Tipo I) + 16.91; r=0.75; n=19; p<0,001. B)
Eficiencia delta (ED) con una velocidad de pedaleo de 80 RPM, determinada como
el cociente entre el cambio en el trabajo realizado.min-1 y el cambio
en la energía gastada.min-1. Ecuación: ED= 0.1393 * (% Tipo I) +
13.49; r=0,85; n=19; p<0.001. C) Eficiencia total (ET) con el test de
extensión de rodilla con las dos piernas, determinada como el cociente entre el
trabajo realizado.min-1 y la energía gastada.min-1.
Ecuación: ET= 0.0920 * (% Tipo I) + 9.489; r= 0.75; n=13; p<0.005.
Eficiencia durante el test de extensión de rodilla y % de fibras musculares de Tipo I
El VO2 pico durante la extensión de rodilla con las dos piernas, tuvo un promedio de 2.5±6.81.min-1. Como se muestra en la Figura 1 C, la ET durante la extensión de rodilla, también estuvo significativamente correlacionada con el % de fibras de Tipo I (r= 0.74; p<0.05). Los valores absolutos de ET durante este test de extensión fueron menores que durante los tests de ciclismo (Tabla 2), posiblemente debido a las menores tasas de gasto energético, en acuerdo a lo que se discutió previamente (9). Se debe notar, sin embargo, que nuestras mediciones del gasto energético durante el test de extensión de rodilla, no parecieron ser lo suficientemente exactas como para permitir la precisión requerida para calcular la ED, y como se relaciona la misma con las diferencias en el Tipo de fibras musculares. Una cantidad relativamente pequeña de músculo estuvo activa durante esta prueba y, por lo tanto, el VO2 aumentó solamente 0.51.min-1 (1.25±0.07 vs. 1.75 ±0.06 l.min-l), mientras que la intensidad del trabajo aumentaba, aproximadamente, del 50 al 70% del pico de VO2.
DISCUSION
Nosotros observamos previamente, que la economía de ciclismo (W.l-1, VO2.min-1; ver Tabla 3 de la ref. 5), puede variar enormemente en un grupo de ciclistas bien entrenados. Sin embargo, se vio que estas diferencias no fueron debidas a diferencias en la técnica de ciclismo, evidenciadas por varios datos biomecánicos derivados del uso de un dinamómetro en el pedal (5). Las variaciones entre sujetos en los índices biomecánicos de efectividad, definida como el porcentaje de fuerza aplicada al pedal que, actualmente crea un torque propulsivo a través de la revolución del pedal (ver IE 180 IE 360 en la Tabla 6 de la ref. 5), no estuvieron correlacionadas con la economía de ciclismo. En cambio, esta variable estuvo relacionada con el % de fibras musculares de Tipo I en el vasto lateral (r= 0.76; p<0,001; n=15). Esta correlación puede ser calculada a partir de los datos de las Tablas 4 y 5 de nuestro trabajo previo (5). El propósito del presente estudio era evaluar, más a fondo, la hipótesis que el % de fibras musculares de Tipo I influyen en la eficiencia de ciclismo.
En el presente trabajo, hemos estimado la eficiencia muscular a través de mediciones corporales del VO2, mientras se realizaba ejercicio. Es importante discutir las presunciones y limitaciones de este enfoque. En primer lugar, el trabajo físico realizado.min-1, fue medido con exactitud (± 1%) y, por lo tanto, la cuantificación de la eficiencia (cociente proporcional entre el trabajo realizado.min-1 y energía gastada.min-1) fue dependiente de la validación de la estimación del gasto energético (kcal.min-1), a partir e la evaluación del VO2 y del RER durante el ejercicio en steady-state. A pesar de que VO2 y RER fueron medidos en una forma muy confiable, es difícil determinar que % del gasto energético total contribuye al trabajo físico realizado, en oposición con el gasto energético para otros procesos orgánicos, como por ejemplo, la estabilización y el movimiento de piernas. Nosotros intentamos reducir la variabilidad en la eficiencia total (ET), examinando solamente a ciclistas experimentados altamente entrenados, durante ejercicios en steady-state a intensidades por debajo del UL. Sin embargo, y más importante, nosotros también calculamos la eficiencia delta (ED) durante el ciclismo, en un amplio rango de intensidades. ED provee la más válida estimación de la eficiencia muscular porque las diferencias en la energía gastada.min-1 son calculadas, relativo solamente al cambio en el trabajo real realizado.min-1, lo que elimina la influencia de procesos metabólicos que no contribuyan al verdadero trabajo llevado a cabo (9,24). Por lo tanto, la evidencia más sólida de la relación directa entre la eficiencia muscular y el tipo de fibra se muestra en la Figura 1 B (ED vs. % Tipo I; r=0.85; P<0,001).
También observamos que la influencia del tipo de fibra muscular sobre la eficiencia no es específica del ciclismo, y que hay una relación positiva entre % Tipo I y ET, durante la extensión de rodilla con las dos piernas. Esta observación brinda un apoyo adicional a la idea de que las variaciones en la eficiencia son realmente un fenómeno muscular y que las fibras de Tipo I son más eficientes que las de Tipo II, durante la acción general de extensión de rodilla, realizada a relativamente bajas velocidades (200º. s-1).
Sin embargo, no podemos descartar completamente la posibilidad que la eficiencia muscular pueda estar determinada por alguna otra variable que no sea el % de fibras musculares de Tipo I. Por ejemplo, nosotros observamos que hay una relación significativa entre el número de años de entrenamiento en resistencia y el % de fibras musculares de Tipo I (r=0.60; p<0.01). También observamos lo mismo en un estudio previo (r=0,65; P<0,001; Fig. 8 de la ref. 5). Por lo tanto, es posible que la relación observada entre la eficiencia y el % de fibras de Tipo I se deba más a la habilidad desarrollada por el ciclista que ha entrenado por más tiempo, que al tipo de fibra muscular en sí. De hecho, el número de años que los sujetos del presente estudio han estado entrenando en resistencia estuvo significativamente correlacionado con ED (r= 0.73; p<0,001) y con ET (r=0.63; p<0,01) mientras realizaban la prueba en ciclismo. Sin embargo, si los años de entrenamiento de resistencia fuera el principal factor para determinar ET o ED, nosotros pensamos que habríamos observado, previamente, alguna manifestación biomecánica de una mejor habilidad, tal como una efectividad superior en el ciclismo, que estuviera relacionada con la economía del mismo. Sin embargo, este no fue el caso. Además, a pesar de que los años de entrenamiento en resistencia estuvo correlacionado, tanto con ET como con ED en el presente estudio, no hemos observado una relación significativa entre los años de entrenamiento en ciclismo y estas variables (ET y ED; r= 0,10-0,25; NS). En realidad, las correlaciones significativas entre los años de entrenamiento en resistencia con ED y ET, parecen estar influenciadas en gran medida por la asociación entre el % de fibras musculares de Tipo I y los años de entrenamiento en resistencia. Esto está basado en nuestras observaciones, ya que cuando la relación entre los años de entrenamiento en resistencia y el % de fibras de Tipo I es excluida del análisis, las correlaciones entre esta primer variable, con ED y ET, se reducen a niveles no significativos (r=0,52 y 0.35; p>0,05, respectivamente). Por estas razones, nosotros pensamos que es probable que el % de fibras musculares de Tipo I sea, en verdad, el principal factor que influye sobre la eficiencia muscular.
También se debe notar que el % de fibras del Tipo I en el vasto lateral, no necesariamente son las mismas en toda la musculatura que es utilizada en ciclismo, a pesar de que este músculo brinda una medición razonable del % de estas fibras en los músculos extensores de la rodilla (13). Nosotros presumimos que el % de fibras de Tipo I del vasto lateral brinda una estimación del "pool" de fibras, de toda la musculatura, que participa en el ciclismo. Por lo tanto, los resultados dan una estimación de la tendencia general, y no necesariamente una cuantificación exacta de la eficiencia absoluta de las fibras de Tipo I y II.
Las observaciones en este estudio, de que los sujetos con un alto % de fibras de Tipo I son más eficientes que los sujetos con un bajo % de estas fibras, deben ser interpretadas a la luz de las velocidades de contracción empleadas. La eficiencia muscular pico, generalmente, ocurre a una velocidad de aproximadamente un tercio de la velocidad máxima de acortamiento (Vmáx.), tanto en fibras musculares de Tipo I como II (11,15). La Vmáx. de las fibras musculares de Tipo II es 3-5 veces mayor que las de Tipo I (8) y, por lo tanto, la velocidad de eficiencia pico en las fibras de Tipo II también es 3-5 veces mayor (11,15). Es probable que las fibras musculares de Tipo I sean más eficientes en los sujetos del presente estudio, debido que las fibras en el vasto lateral se hayan contraído más cerca de la velocidad de eficiencia pico de las fibras de Tipo I, comparadas con las fibras de Tipo II, mientras se realizaban extensiones de rodilla a velocidades cercanas a los 200º. s-1.
Esta idea se puede ilustrar mejor y fundamentar teóricamente, en principio, estimando las velocidades que se predicen para provocar la eficiencia pico en las fibras de Tipo I, comparadas con las de Tipo II, y la relativa velocidad de acortamiento del vasto lateral en el presente estudio. Sin embargo, se debe notar que estos puntos son especulativos y sirven, principalmente, para presentar una hipótesis de trabajos, ya que hay probablemente, limitaciones en la extrapolación de las observaciones "in Vitro" al modelo presente. Sin embargo, Fitts y cols. (8) han determinado, directamente, la Vmáx. de las fibras I y II, en forma aislada, en el músculo deltoides de nadadores de resistencia con características similares a los sujetos de nuestro trabajo. Cuando los valores de la Vmáx. en el estudio de Fitts y cols. (8), medidos a 15ºC, fueron recalculados para una Vmáx. esperada a 37ºC (comunicación personal con el Dr. Robert Fitts), nosotros estimamos que un tercio de la Vmáx. en las fibras de Tipo I (la velocidad de eficiencia pico), corresponde a una velocidad de acortamiento de, aproximadamente, de una longitud de fibras. s-1. Goldspink (11) también observó que las fibras de Tipo I trabajan más eficientemente, a una velocidad de acortamiento cercana a una longitud de fibra. s-1 (ver Fig. 2a de la ref. 11). De acuerdo a Wickiewicz y cols. (25), la extensión de rodilla a 200º.s-1 requeriría que el vasto lateral se acortara aproximadamente 90 mm.s-1. Debido a que el promedio en la longitud de las fibras en el vasto lateral es 72 mm, más o menos, la velocidad de acortamiento sería aproximadamente de 1,2 longitud. s-1 (90 mm. s-1x 72mm. s-1) (25). Por lo tanto, la velocidad estimada de acortamiento de las fibras musculares en el vasto lateral bajo estas condiciones (1,2 longitud de fibras. s-1) es cercana a la velocidad estimada de eficiencia pico en las fibras de Tipo I (10,15). Por los datos de Fitts y cols. (8), las fibras de Tipo II en atletas de resistencia, estarían en su velocidad de eficiencia pico cuando se contraen a aproximadamente 3 longitud. s-1, mientras que estas fibras en personas no entrenadas y en otros mamíferos, parece que trabajan a una eficiencia pico cuando se contraen a 5 longitud. s-1 (8,11). Nuevamente, nuestro principal punto ilustrativo es que, es probable que las velocidades de contracción usadas en el presente estudio, fueran más cercanas a la velocidad de eficiencia pico de las fibras de Tipo I que las de Tipo II, razón por la cual las de Tipo I parecen más eficientes que las de Tipo II.
Como se discutió previamente, la relación entre ED pedaleando a 80 RPM, y el % de fibras de Tipo I (Fig. 1 B), probablemente brinda la estimación más válida de las diferencias relativas en la eficiencia muscular de las fibras de Tipo I, en comparación con las fibras de Tipo II. Cuando esta relación es extrapolada (a 0% de Tipo I y a 100% de Tipo II), se predice que la eficiencia es, aproximadamente, dos veces mayor en el Tipo I que en el Tipo II (27% vs. 13%, respectivamente), cuando se pedalea a 80 RPM. Este reporte concuerda con las observaciones que afirman que las fibras musculares de Tipo I son 2-3 veces más eficientes que las fibras de Tipo II, cuando son evaluadas durante la contracción isotónica a velocidades relativamente bajas (10, 11, 22, 23).
El reclutamiento de fibras musculares cuando se pedalea a intensidades entre el 50 y el 80% del VO2 máx. (aproximadamente 180-300 W), en estos ciclistas bien entrenados (VO2 máx.: 4,9º 0,1 l.min-1), es una consideración importante. Estas tasas de esfuerzo fueron elegidas en base a la evidencia de que la mayoría de las fibras en el vasto lateral, sea tanto las de Tipo I como las Tipo II, son reclutadas bajo las condiciones del presente experimento (4,12). Si este no fuera el caso, y sólo una pequeña parte de las fibras de Tipo I fuera reclutada, obviamente no sería posible estimar la eficiencia de las fibras de Tipo II. Ivy y cols. (12), evaluaron los cambios de metabolitos en las fibras musculares individuales, con el fin de determinar el perfil de reclutamiento de estas fibras cuando los ciclistas (VO2 máx. de 4,55 l.min-1) ejercitaban al 52% y 72% del VO2 máx.. Basados en los cambios en la glucosa-6-fosfato y en la concentración de malato dentro de las fibras musculares, concluyeron que cuando los ciclistas pedaleaban al 52% del VO2 máx. "casi todas las fibras, sin importar el tipo, habían sido reclutadas" y que "todas las fibras participan en la contracción en un grado sustancial, aún con cargas de trabajo moderadas" (52-72% del VO2 máx..). Esto concuerda con nuestra observación, de que el ciclismo prolongado, al 71% del VO2 máx.., resulta en un vaciamiento de glucógeno en casi todas las fibras del vasto lateral (92% de las fibras, Fig. 2 de la ref. 4).
Nuestra observación sobre la correlación positiva entre el % de fibras de Tipo I y la ED, cuando se pedalea a 80 RPM, no concuerda con el informe de Suzuki (19). El observó que la ED, pedaleando a 60 RPM, era similar en un grupo de ciclistas entrenados (N= 3) con un predominio de fibras de Tipo I (78%), en comparación con individuos no entrenados (N= 3), con relativamente pocas fibras de Tipo I (24%). Es probable que el pequeño tamaño de la muestra y las grandes diferencias en el nivel de entrenamiento de los dos grupos estudiados por Suzuki (19), hayan influido en la interpretación de sus datos.
El % de fibras musculares de Tipo I en el presente estudio se determinó a través de múltiples biopsias en el vasto lateral. Las fibras fueron clasificadas como de Tipo I o Tipo II en base al clásico análisis químico de la actividad de la miosina ATPasa (2,18). Casi todas las fibras de Tipo II en estos sujetos fueron de Tipo IIa, como es de esperar en atletas de resistencia bien entrenados (6,14). Es posibles que algunas de las fibras de estos deportistas tengan la coexistencia de cadenas de miosina, tanto de Tipo I como de Tipo IIa (14). Sin embargo, parece ser que a pesar de esta posible coexistencia, las propiedades contráctiles de estas fibras identificadas histoquímicamente como de Tipo I y II son bien distintivas, y tienen marcadas diferencias en la eficiencia.
Nosotros concluimos que la eficiencia muscular varía enormemente en ciclistas altamente entrenados en resistencia, y que la mayor parte de esta variabilidad está relacionada con diferencias en el % de fibras musculares de Tipo I. Por lo tanto, cuando los sujetos que entrenan la resistencia realizan una extensión de rodilla a una velocidad de aproximadamente 200º. s-1, las fibras de Tipo I parecen ser sustancialmente más eficientes que las de Tipo II, posiblemente como resultado de una menor tasa de consumo y reposición de ATP (turnover), reflejada por un menor VO2, mientras se realizan ejercicios a una intensidad dada.
Agradecimientos
Apreciamos enormemente la cooperación e interés de los ciclistas que participaron en este estudio. Agradecemos al Dr. John Ivy por realizar algunas de las biopsias musculares. También apreciamos los útiles comentarios del Dr. Roger Farrar.
Este proyecto se realizó, en parte, con una beca del Louise Spence Griffeth Fellowship and Dr. E.F. Coyle, L.S. Sidossis era becario de la Fundación Onassis.
Referencias
1. Barany, M (1967). ATPase activity of myosin correlated with speed of muscle shortening. J. Gen. Physiol. 50: 197-216
2. Brooke, M.H. and K.K. Kaiser (1970). Three myosin ATPase systems: the nature of their pH lability and sulfhydryl dependence. J. Histochem. Cytochem. 18: 670-672
3. Cavanaugh, P.R. and D.J. Sanderson (1986). The biomechanics of cycling: studies of the pedaling mechanics of elite pursuit riders. In: Science of Cycling, E.R. Burke (Ed.). Champaign, IL: Human Kinetics Publishers, Inc. pp. 91-122
4. Coggan, A.C. and E.F. Coyle (1987). Reversal of fatigue during prolonged exercise by carbohydrate infusion or ingestion. J. Appl. Physiol. 63: 2388-2395
5. Coyle, E.F., M.E. Feltngr, S.A. Kautz, et al (1991). Physiological and biomechanical factors associated with elite endurance cycling performance. Med. Sci. Sports Exerc. 23: 93-107
6. Coyle, E.F., W.H. Martin Iii, S.A. Bloomfield, O.H. Lowry, and J.O. Holloszy (1985). Effects of detraining on responses to submaximal exercise. J. Appl. Physiol. 59: 853-859
7. Elder, G.C.B., K. Bradbury, and R. Roberts (1982). Variability of fiber type distributions within human muscles. . Appl. Physiol. 53: 1473-1480
8. Fitts, R.H., D.L. Costill, and P.R. Gardetto (1989). Effects of swim exercise training on human muscle fiber function. J. Appl. Physiol. 66: 465-475
9. Gaesser, G.A. and G.A. Brooks (1975). Muscular efficiency during steady-state exercise: effects of speed and work rate. J. Appl. Physiol. 38: 1132-1139
10. Gibbs, C.L. and W.R. Gibson (1972). Energy production of rat soleus muscle. Am. J. Physiol. 223: 864-871
11. Goldspink, G (1978). Energy turnover during contraction of different types of muscle. In: Biomechanics VI-A, E. Asmussen and K. Jorgensen (Eds.). Baltimore: University Park Press, pp. 27-39
12. Ivy, J.L., M. M-Y.Chi, C.S. Hintz, W.M. Sherman, R.P. Hellendall and O.H. Lowry (1987). Progressive metabolite changes in individual human muscle fibers with increasing work rates. Am. J. Physiol. 252 (Cell Physiol. 21). C 630-C 639
13. Johnson, M.A., J. Polgar, D. Weightman and D. Appleton (1973). Data on the distribution of fiber types in 36 human muscles. J. Neurol. Sci. 18: 111-129
14. Klitgaard, H., O. Bergman, R. Betto, G. Salbiati, S. Schiaffino, T. Clausen and B. Saltin (1990). Coexistence of myosin heavy chain I and IIa isoforms in human skeletal muscle muscle fibres with endurance training. Eur. J. Physiol. (Pflugers Arch.) 416: 470-472
15. Kushmerick, M.J (1983). Energetics of muscle contraction. In: Handbook of Physiology, Sec. 10: Skeletal Muscle. L.E. Peachey, R. H., and S.R. Geiger (Eds.). Bethesda, MD: A. Physiologycal Society, pp. 189-236
16. Lusk, G (1928). The Science of Nutrition. Philadelphia: Saunders
17. Peatman, J.G (1963). Introduction to Applied Statistics. New York: Harper and Row
18. Staron, R.S. and D. Pette (1986). Correlation between myofibrillar ATPase activity and myosin heavy chain composition in rabbit muscle fibers. Histochemistry 86: 19-23
19. Suzuky, Y (1979). Mechanical efficiency of fast-and slow-twitch muscle fibers in man during cycling. J. Appl. Physiol.: Respirat. Environ. Exerc. Physiol. 47: 263-267
20. Sweeney, H.L., M.J. Kushmerick, K. Mabuchi, F. Sreter and J. Gergely (1988). Myosin alkali light chain and heavy chain variations correlate with altered shortening velocity of isolated skeletal muscle fibers. J. Biol. Chem. 263: 9034-9039
21. Welltman, A., D. Snead, P. Stein, R. Schurrer, R. Ruth and J. Weltman (1990). Reliability and validity of a continuous incremental treadmill protocol for the determination of lactate threshold, fixed blood lactate concentrations, and VO2max. Int. J. Sports med. 11: 26-32
22. Wendt, I.R. and C.L. Gibbs (1973). Energy production of mammalian fast-and slow-twitch muscles during development. Am. J. Physiol. 226: 642-647
23. Wendt, I.R. and C.L. Gibbs (1974). Energy production of mammalian fast-and slow-twitch muscles during development. J. Physiol. 226: 642-647
24. Whipp, B.J. and K. Wasserman (1969). Efficiency of muscular work. J. Appl. Physiol. 26: 644-648
25. Wickiewicz, T.L., R.R. Roy, P.L.Powell, J.J. Perrine, and V.R. Edgerton (1984). Muscle architecture and force-velocity relationships in human. . Appl. Physiol. 57: 435-443