
La Provisión de Energía Anaeróbica No Limita el Rendimiento durante el Test de Wingate en Ciclistas Entrenados en Resistencia
José Antonio López Calbet1, J. A De Paz2, S. Cabeza de Vaca3 y J. Chavarren1
1Department of Physical Education, University of Las Palmas de Gran Canaria, 35017 Las Palmas de Gran Canaria, Islas Canarias, España.
2Instituto Nacional de Educación Física de León, University of León, 24071 León, España.
3Centro de Tecnificación de Ciclismo, Chiclana de la Frontera, 11130 Cádiz, España.
Artículo publicado en el journal PubliCE, Volumen 0 del año 2003.
Publicado 8 de septiembre de 2004
Resumen
Palabras clave: fatiga, potencia anaeróbica, capacidad anaeróbica, lactato
INTRODUCCION
Aun se ignora el impacto que tiene la hipoxia severa aguda sobre la producción de energía aeróbica y anaeróbica durante el test de Wingate de 30s. Aunque la hipoxia moderada aguda [fracción inspirada de O2 (FIO2)=0.13] no tiene efectos sobre el consumo de O2 (VO2) o sobre el rendimiento en ejercicios supramáximos de hasta 30s de duración (39), se han reportado resultados conflictivos acerca de la influencia que podrían tener mayores niveles de hipoxia sobre el rendimiento y el metabolismo (22, 23). McLellan y cols. (23) no observaron cambios en la producción media de potencia (PMEDIA) ni en el VO2 durante el test de Wingate de 30s realizado con una FIO2 de ~0.11. La prolongación del test de Wingate hasta 45s ha resultado consistentemente en un menor VO2 bajo condiciones de hipoxia (22, 23) mientras que, en comparación con la normoxia, la PMEDIA durante el test de Wingate de 45s no fue afectada por la hipoxia aguda en un estudio (23) y fue ligeramente reducida (~3% menor) en otra investigación (23). Las pequeñas diferencias en la FIO2 y en el tiempo de exposición a la hipoxia antes del comienzo de los test de Wingate podrían explicar los resultados aparentemente contradictorios reportados por McLellan y cols. (22, 23). Se ha demostrado claramente, por otro lado, que durante ejercicios máximos de 30 a 45s de duración en condiciones de hipoxia aguda, la acumulación de lactato se incrementa marcadamente, indicando una mayor liberación de energía anaeróbica con la hipoxia aguda (23). Además, cierta evidencia sugiere que durante ejercicios supramáximos que provocan el agotamiento entre los 30 y 60 s, la contribución de las fuentes de energía anaeróbica se incrementa en condiciones de hipoxia moderada aguda (22, 39). No se sabe, si un mayor grado de hipoxia podría aumentar la liberación de energía anaeróbica durante el test de Wingate de 30 s.
Se ha estimado que, en general, las fuentes de energía anaeróbicas proveen el 70-80% de la energía utilizada a través del test de Wingate (6, 28, 33, 40). Sin embargo en comparación con los especialistas en velocidad, los atletas entrenados en resistencia tienen un mayor VO2 medio durante el test de Wingate (12) y obtienen una mayor fracción de energía a partir del metabolismo oxidativo. En cambio los atletas entrenados en velocidad obtienen una mayor fracción de energía a través de las vías anaeróbicas (6). De esta manera puede hipotetizarse que debido a que los atletas entrenados en resistencia dependen más de las fuentes de energía aeróbica para realizar ejercicios supramáximos, los mismos experimentarán una desmejora relativamente mayor del rendimiento, bajo condiciones de hipoxia aguda, que los atletas entrenados en velocidad, a menos que compensen la reducción en el VO2 por medio del aumento de la liberación de energía anaeróbica.
Por ello, el propósito principal de este estudio fue averiguar si la reducción en el suministro de O2 provocado por la hipoxia aguda severa puede ser contrarrestado por el aumento de la producción de energía anaeróbica durante el test de Wingate de 30 s. Otro propósito fue verificar si los ciclistas de pista de elite entrenados en resistencia experimentan una mayor desmejora del rendimiento que los ciclistas entrenados en velocidad, durante el test de Wingate de 30s bajo condiciones de hipoxia aguda. Para explicar más fácilmente los efectos de la hipoxia, la FIO2 fue reducida a 0.104, lo cual es equivalente a una altura de ~5300 m. Este nivel de hipoxia es cercano al límite que humanos sanos no aclimatados pueden tolerar agudamente durante ejercicios realizados en posición vertical (4, 32, 36).
METODOS
Sujetos
Este estudio fue aprobado por el comité de ética de la Universidad de las Palmas de Gran Canaria. Los sujetos fueron completamente informados sobre los propósitos, posibles riesgos y beneficios de su participación en el estudio, antes de dar su consentimiento escrito. Diez ciclistas de pista del Equipo Nacional Español participaron en este estudio. Todos ellos calificaron más de una vez entre los tres primeros en el Campeonato Nacional Español. El grupo incluyo varios campeones nacionales y a un medallista de plata en el último campeonato mundial, quien era un especialista en velocidad. Cinco de los ciclistas eran especialistas en velocidad, y los otros cinco eran entrenados en resistencia. Los valores medios±DE para la edad, la altura, el peso y el porcentaje de grasa corporal fueron, 19.0±0.7 vs. 18.8±0.4 años, 176±2 vs. 179± 0.7 cm, 74.7±3.1 vs. 65.8±1.0 kg, y 13.2±1.9 vs. 9.3±0.5%, para los ciclistas de velocidad y de resistencia respectivamente. Se les pidió a los sujetos que siguieran dietas similares y que disminuyeran su actividad física durante las 48 hs previas a las pruebas. También se les dio instrucciones para que no consumieran ninguna comida por al menos 4 hs antes de la sesión de evaluación.
Protocolos y Procedimientos Experimentales
El porcentaje de grasa corporal fue estimado antropométricamente por medio de la utilización de una ecuación específica para la población, la cual fue desarrollada y validada en nuestro laboratorio utilizando absorciometría dual por rayos X como patrón o gold standard (19). Los sujetos completaron cuatro sesiones de evaluaciones en días separados. Primero se evalúo el VO2 máximo (VO2 máx.) y la máxima producción de potencia (WMAX.) por medio de un test progresivo hasta el agotamiento (25 W/min a 90 rpm). Durante los siguientes dos días de evaluación, se determinaron las relaciones entre el VO2 y la intensidad (o la economía de pedaleo). Luego, durante la última sesión de evaluación, se llevaron a cabo los test de Wingate, mientras los sujetos respiraban el aire de la habitación (normoxia) o aire proveniente de bolsas de Douglas que contenían una fracción de O2 del 10.4% en N2 (hipoxia), equivalente a una altura de ~5300 m. Este nivel de hipoxia aguda fue elegida debido a que reduce el VO2 máx. aproximadamente en un 50% (10, 36), creando, por lo tanto, una condición en la cual se presentaría una limitación potencial al suministro de energía aeróbica durante los test de Wingate realizados en hipoxia. Los tests de Wingate en normoxia y en hipoxia se realizaron en orden aleatorio y separados por un período de recuperación de al menos 1 h, el cual es suficiente para permitir una recuperación completa (6).
Evaluación de la Economía de Pedaleo
La economía de pedaleo se determino utilizando 12 cargas submáximas a intensidades de entre 60 y 90% del VO2 máx., a cuatro tasas de pedaleo diferentes: 60, 80, 100 y 120 rpm. Las intensidades de ejercicio y las tasas de pedaleo fueron administradas en orden aleatorio, separadas por períodos de 3-5 min. Para reducir el estrés térmico y minimizar las perdidas de agua debido a la sudoración, a los sujetos se les proporcionó ventilación e ingirieron en total 600 ml de agua durante los períodos de recuperación. Las evaluaciones fueron realizadas a 18-24ºC, 60-80% de humedad relativa y 750-770 mmHg de presión atmosférica. La duración de cada seria submáxima fue de 6 min. El VO2 medio registrado durante los últimos dos minutos fue tomado como representativo de cada intensidad submáxima de ejercicio. Para relacionar el VO2 a la potencia, se calcularon ecuaciones de regresión no lineales individuales por medio del ajuste lineal de cuadrados mínimos, aplicando el siguiente modelo no lineal: VO2 = a+b•W+c•rpm+d•rpm2, el cual arrojó un error estándar de estimación que siempre fue menor a 100 ml/min de O2, donde a, b, c, y d son constantes a ser determinadas por la ecuación de regresión no lineal; W es la intensidad de ejercicio y rpm es la tasa de pedaleo.
Tests de Wingate
Para determinar la concentración de lactato corporal total ([La]) en la vena femoral, se insertó percuténeamente un catéter con una aguja de 20 (Hydrocath, Ohmeda, Swindon, Reino Unido) en la vena femoral bajo anestesia local (2% de lidocaína), utilizando la técnica de Seldinger, como se reporto previamente (29). El catéter fue insertado a 2 cm por debajo del ligamento inguinal y se avanzó 12 cm hacia la rodilla para evitar la contaminación de la sangre proveniente de las venas profundas del cuadriceps con sangre de la vena safena. Una vez colocado, el catéter fue suturado a la piel para minimizar el riesgo de movimiento o de plegado, y la salida del catéter fue conectada a una llave de tres vías. Luego de un período de reposo de ~5 min, los sujetos llevaron a cabo una entrada en calor estandarizada que consistió en 10 min de pedaleo continuo a una intensidad cercana al 60% del VO2 máx. y luego realizaron cinco aceleraciones máximas de no más de 5-6s de duración. Luego de esto los sujetos descansaron por 10 minutos y fueron asignados aleatoriamente al test de Wingate en normoxia o en hipoxia. Para minimizar el riesgo de hipotensión y de alcalosis respiratoria durante los tests de Wingate en hipoxia, los sujetos comenzaron a respirar de las bolsas con gas hipóxico 3 minutos antes del comienzo del ejercicio (36). Durante los tests de Wingate, las muestras de sangre de la vena femoral fueron extraídas continuamente por períodos de 5 s, dando un total de seis muestras de sangre durante el ejercicio. Luego de ambos tests de Wingate, los sujetos se recuperaron en normoxia mientras estaban recostados en una cama, y se extrajeron muestras adicionales de sangre de la vena femoral a los 3, 5, 7 y 10 minutos después del comienzo del período de recuperación. En todos los ciclistas se utilizaron fuerzas de frenado equivalentes a 0.11 kp por kg de masa corporal.
Todos las evaluaciones fueron llevadas a cabo en un cicloergómetro Monark (Monark 8158E). Durante los tests de Wingate se utilizaron estribos punteras con correas dobles para fijar los pies a los pedales. Para registrar la velocidad de pedaleo cada 2 ms, se conectó un disco con 250 ranuras mediante un engranaje a la cadena (6). El disco con ranuras giraba en frente de una célula fotoeléctrica en interfase con una computadora. El trabajo realizado sobre el cicloergómetro Monark fue corregido por el trabajo requerido para acelerar la rueda, como se ha reportado anteriormente (6). Brevemente, la carga sobre el cicloergómetro fue considerada como la suma de la carga de fricción (fuerza de frenado aplicada a través de una cinta a la rueda) y la carga que sería requerida en cualquier instante para detener al sujeto, de seguir acelerando la rueda, lo cual depende del momento de inercia de la rueda y de otras fuerzas de fricción que actúan sobre la rueda. Para calcular esta carga, nosotros generamos una serie de curvas de desaceleración utilizando diferentes cargas de fricción. Primero, la rueda fue puesta en movimiento a una velocidad constante de 120 rpm por medio del pedaleo. Segundo, luego de alcanzar una velocidad estable, se detuvo el pedaleo súbitamente y el tiempo transcurrido entre las 105 y las 0 rpm fue utilizado para calcular la desaceleración para cada fuerza de frenado. Por último, se derivó una ecuación de regresión por medio del ajuste lineal de cuadrados mínimos entre la desaceleración de la rueda y la carga.
La energía anaeróbica producida durante los tests de Wingate fue calculada como la deuda de O2, como se reporto previamente (6). Primero, la demanda de O2 se estimó extrapolando la relación no lineal entre el VO2 y la producción de potencia, medida con cargas submáximas. Luego se computó la deuda de O2 como la diferencia entre la demanda de O2 y el O2 consumido durante las series supramáximas (25, 39).
Variables Respiratorias
El intercambio respiratorio de gases fue monitoreado respiración a respiración, a través de un sistema de circuito abierto (CPX, Medical Graphics, St. Paul, MN) y fue promediado cada 15s durante los test progresivos de ejercicio y cada 5s durante los test de Wingate. El sistema metabólico fue calibrado con mezclas de gases de calibración con concentraciones conocidas de O2 y CO2 (precisión 0.01%), las cuales fueron provistas por los fabricantes (CPX, Medical Graphics). En nuestro laboratorio, el VO2 y la producción de CO2 durante el ejercicio submáximo en bicicleta han sido valoradas con un coeficiente de variación menor del 5%, así como también con un coeficiente de confiabilidad intraclase mayor a 0.98, determinado en seis estudiantes de educación física a cuatro intensidades diferentes en 4 días diferentes. El mayor valor de VO2 alcanzado durante el test progresivo de ejercicio fue tomado como el VO2 máx., mientras que la intensidad alcanzada justo antes del agotamiento fue tomada como la WMAX. La WMAX fue ajustada por medio de una extrapolación dependiente de la duración de la última etapa (18). Para determinar la cinética de la respuesta del VO2, los datos respiración a respiración fueron promediados cada 5 segundos y ajustados a una curva utilizando un modelo exponencial, por medio del enfoque del error de los cuadrados mínimos. El procedimiento de ajuste de la curva fue reiterado hasta que cualquier cambio posterior en los parámetros para el modelo no resultara en una reducción en el error cuadrático medio entre la curva obtenida a partir del modelo y el conjunto original de datos. El modelo utilizado para ajustar los datos del VO2 tuvo una constante, la cual corresponde al VO2 de reposo, un término de amplitud (b) y una constante de tiempo (c) como son aquí presentados:
VO2(t) = a + b (1 – c-ct)
Donde t es el tiempo en segundos y VO2(t) es la variación en el VO2 dependiente del tiempo.
Lactato Sanguíneo
La [La] se determinó en todo el cuerpo utilizando un analizador de lactato (YSI 1500 Sport, Yellow Springs, CO) provisto con un agente de hemólisis (Triton X-100). Con este instrumento, hemos obtenido un coeficiente de variación para la valoración el lactato sanguíneo [La] menor a 1% para la [La] entre 1 y 26 mM. Las curvas de recuperación de [La] sanguíneo fueron integradas a través del tiempo y expresadas como milimoles • segundos. Los valores reportados corresponden al área bajo la curva, i.e., el área bajo la curva luego de considerar la concentración de lactato sanguíneo de reposo inmediatamente antes del comienzo de los tests de Wingate.
Análisis Estadísticos
La estadística descriptiva fue realizada para cada variable para confirmar la suposición de normalidad y homocedasticidad. El efecto de la fracción de O2 inspirado sobre el lactato sanguíneo femoral durante los tests de Wingate fue determinado utilizando análisis de varianza de dos vías para medidas repetidas, siendo la especialidad de los ciclistas un factor de segundo nivel. En el caso de un valor F significativo, las comparaciones planeadas se llevaron a cabo utilizando las pruebas t de Student para datos apareados o no apareados (según lo que fuera apropiado) con la corrección de Bonferroni para las comparaciones múltiples. El nivel de significancia fue establecido a p<0.05. Los datos fueron expresados como medias±DE.
RESULTADOS
Diferencias Entre los Ciclistas Entrenados en Resistencia y Entrenados en Velocidad
Los ciclistas de resistencia mostraron mayores valores de VO2 máx. que los ciclistas entrenados en velocidad (72±1 y 62±2 ml.kg-1.min-1, p<0.05). Como se muestra en la Figura 1A y en la Tabla 1, el pico de producción de potencia (PMAX) y la PMEDIA, tanto en términos absolutos como relativos, fue mayor en los ciclistas de velocidad que en los ciclistas de resistencia. De la misma manera, los ciclistas de velocidad alcanzaron mayores velocidades máximas y medias de pedaleo que los ciclistas entrenados en resistencia (Figura 1B). Sin embargo, los ciclistas de velocidad, se agotaron más rápido que los ciclistas de resistencia, como lo indica el índice de fatiga durante el test de Wingate, el cual fue 0.46±0.12 y 0.32±0.12 W.s-1.kg-1 de masa corporal, para los ciclistas de velocidad y de resistencia respectivamente (Figura 2A, p<0.05). En consecuencia, la superioridad de los ciclistas de velocidad en la producción de potencia se redujo progresivamente durante la segunda mitad de los test de Wingate, desarrollando en ambos grupos valores casi similares de producción de potencia por kilogramo de masa corporal durante los últimos 15s de los tests (Figura 1A). Los ciclistas de velocidad incurrieron en una mayor demanda de O2 durante los primeros 10-15 segundos del test de Wingate (Figuras 1 C y D). Probablemente debido a su mayor VO2 máx., los ciclistas de resistencia fueron capaces de consumir un 26% más de O2 por kilogramo de masa corporal durante el test de Wingate que los ciclistas de velocidad (p<0.05), ya que ambos grupos utilizaron un porcentaje similar de su VO2 máx. durante los tests de Wingate en normoxia (Figura 2B). De hecho, se observó una estrecha correlación entre el VO2 medio durante el test de Wingate y el VO2 máx. (Figura 2C; r=0.86, p<0.001). Con una mayor demanda de O2 y un menor VO2, la deuda de O2 por kilogramo de masa corporal resultó un 33% mayor en los ciclistas de velocidad de que en los ciclistas de resistencia (Figura 1H). La diferencia entre el 26% mayor de energía aeróbica producida por los ciclistas de resistencia y la deuda de O2 33% mayor en los ciclistas de velocidad derivó en el desarrollo de una PMEDIA ~8% mayor, desarrollada durante el test de Wingate por los especialistas en velocidad. A pesar de estas notables diferencias en la producción de energía anaeróbica durante los tests de Wingate, la tasa de acumulación de [La] sanguíneo durante el ejercicio fue similar en los ciclistas de resistencia y de velocidad. En ambos grupos la [La] sanguínea no cambio durante los primeros 15s de ejercicio, pero luego de esto aumentó, describiendo una parábola a medida que pasaba el tiempo de ejercicio (en todas las condiciones, r=0.99, p<0.001; Figura 1G). Durante los primeros 10 minutos del período de recuperación, los ciclistas de velocidad acumularon un 27% más de lactato en la vena femoral que los ciclistas de resistencia (p<0.05, Figura 3).
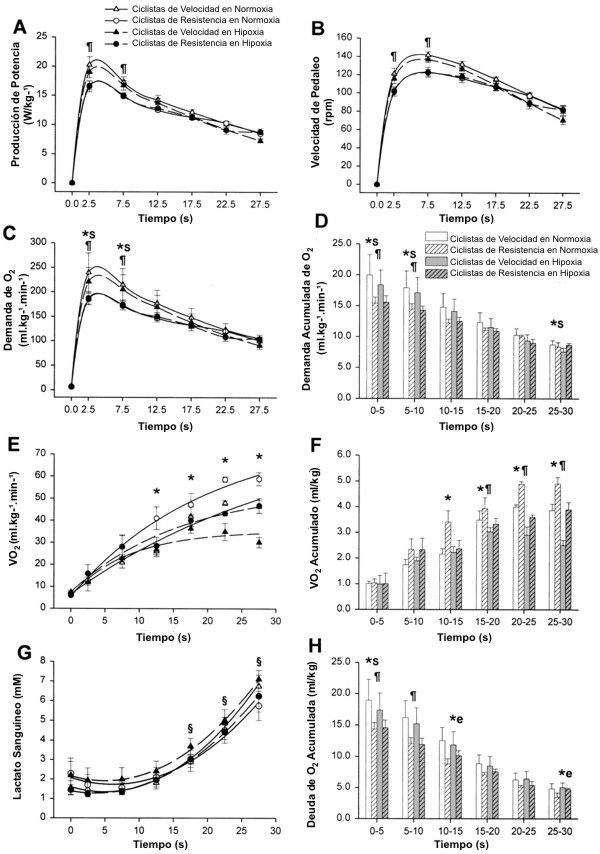
Figura 1. Lapso de tiempo para la producción de potencia (A), velocidad de
pedaleo (B), demanda de O2 (C), demanda acumulada de O2
(D), consumo de O2 (VO2; E), VO2 acumulado
(F), concentración sanguínea de lactato en la vena femoral (G) y deuda de O2
(H) durante los tests de Wingate de 30 segundos en condiciones de normoxia y
de hipoxia severa aguda. * p<0.05 normoxia vs. hipoxia; p<0.05 ciclistas
entrenados en velocidad vs. ciclistas entrenados en resistencia; § p<0.05 en
comparación con el minuto 0; *S p<0.05 normoxia vs. hipoxia, pero solamente
para los ciclistas entrenados en velocidad; *e p<0.05 normoxia vs hipoxia,
pero solamente para los ciclistas entrenados en resistencia.
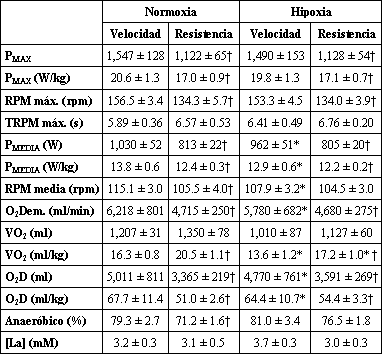
Tabla 1. Diferencias entre los ciclistas entrenados en velocidad y los
ciclistas entrenados en resistencia. Los valores son presentados como
medias±DE. PMAX., potencia pico; RPM máx., máxima velocidad de
pedaleo; TRPM máx., tiempo para alcanzar la tasa máxima de pedaleo; PMEDIA,
producción de potencia media; RPM media, velocidad de pedaleo media; O2
Dem., demanda de O2; VO2, consumo de oxígeno; O2D,
deuda de O2; Anaeróbico, porcentaje de producción de energía
provista por el metabolismo anaeróbico; [La], concentración de lactato
sanguíneo en la vena femoral. * diferencia significativa desde la normoxia a
la hipoxia; † diferencia significativa entre los ciclistas entrenados en
velocidad y los entrenados en resistencia.
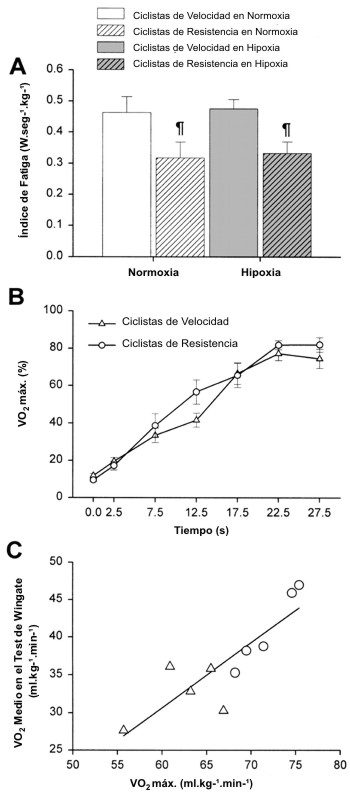
Figura 2. A: Indice de fatiga durante los tests de Wingate de 30 s. B:
porcentaje de utilización del VO2 máximo (VO2 máx.)
durante los tests de Wingate realizados en condiciones de normoxia. C:
correlación entre el VO2 medio durante los tests de Wingate en
normoxia y el VO2 máx. p<0.05 ciclistas entrenados en velocidad vs.
ciclistas entrenados en resistencia.
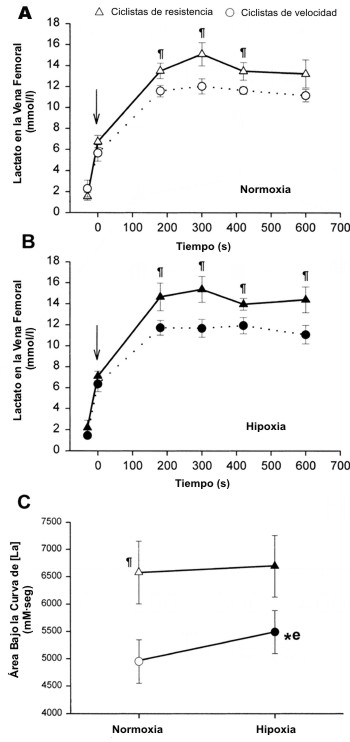
Figura 3. A: Concentración de lactato sanguíneo ([La]) en la vena
femoral durante el período de recuperación después del test de Wingate en
normoxia. B: Concentración de lactato sanguíneo ([La]) en la vena femoral
durante el período de recuperación después del test de Wingate en hipoxia. C:
área bajo la curva de [La] en la vena femoral durante los primeros 10 minutos
del período de recuperación (los valores basales están descontados). p<0.05
ciclistas de velocidad vs. ciclistas de resistencia * e p<0.05 normoxia vs.
hipoxia, pero solo para los ciclistas de resistencia. La flecha vertical
indica el momento correspondiente al final del test de Wingate (minuto 0 de la
curva de recuperación).
Como se muestra en la Figura 4 la contribución de energía anaeróbica al gasto energético total durante el test de Wingate se redujo siguiendo un patrón parabólico (r>0.98, p<0.01). Los ciclistas de velocidad obtuvieron una proporción de energía ligeramente mayor a través de las vías anaeróbicas (Tabla 1). La extrapolación de las curvas representadas en la Figura 3 al 0% de la contribución de energía anaeróbica nos permitió estimar la duración que un test máximo debería tener para posibilitar la completa utilización de todo el potencial energético anaeróbico (Figura 4). Debido a que los ciclistas de velocidad tuvieron una mayor deuda de O2 y obtuvieron una mayor fracción de energía a través de las vías anaeróbicas, la tasa de liberación de energía anaeróbica fue mucho más alta en los ciclistas de velocidad que en los ciclistas de resistencia. A pesar del más rápido reclutamiento de su capacidad anaeróbica, los ciclistas de velocidad necesitaron 4-5 segundos más que los ciclistas de resistencia para expresar completamente su capacidad anaeróbica.
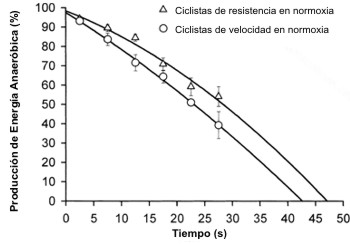
Figura 4. Fracción de energía provista por las fuentes anaeróbicas durante
los tests de Wingate de 30s realizados en normoxia. La parábola fue ajustada
para cada conjunto de datos (r=0.99, p<0.01) y extrapolada al 0% de la
contribución anaeróbica para estimar el tiempo requerido para utilizar
totalmente la capacidad anaeróbica durante un tests máximo realizado en
normoxia.
Efectos de la Hipoxia Severa Aguda
El paso del tiempo para la producción de potencia, la demanda de O2, el VO2, la deuda de O2 y la [La] sanguínea durante los tests de Wingate en condiciones de normoxia y de hipoxia se muestran en la Figura 1. Aunque la PMAX no fue afectada por la hipoxia en ninguno de los grupos, la PMEDIA y la tasa de pedaleo se redujeron en un 6-7% en los ciclistas de velocidad (p<0.05). Sin embargo, en los ciclistas de resistencia la PMEDIA y la tasa de pedaleo se mantuvieron al mismo nivel que en la normoxia. Con una menor PMEDIA, la demanda de O2 disminuyó en los ciclistas de velocidad cuando se ejercitaron en hipoxia (p<0.05). En ambos grupos, la hipoxia resultó en un VO2 un 16% menor (p<0.01; Figura 1E). La divergencia entre el VO2 bajo condiciones normóxicas e hipóxicas durante los tests de Wingate comenzó 10 segundos después del comienzo del test y se volvió más acentuada a mediada que el ejercicio progresaba (p<0.05).
La hipoxia tuvo efectos opuestos sobre la deuda de O2 en los ciclistas de velocidad y los ciclistas de resistencia, ya que, en comparación con la normoxia, la hipoxia resultó en una deuda de O2 un 5% menor en los ciclistas de velocidad (p<0.05), mientras que promovió un valor un 7% mayor en los ciclistas de resistencia (p<0.05). Este efecto diferente fue adicionalmente sostenido por un significativo efecto de interacción en el análisis con ANOVA. En concordancia, los ciclistas de resistencia tuvieron un área bajo la curva de [La] sanguíneo un 11% mayor durante el período de recuperación luego de los tests de Wingate en condiciones de hipoxia (p<0.05; Figura 3), mientras que la hipoxia no afecto la [La] en la recuperación en los ciclistas de velocidad. Durante el período de ejercicio de 30 s, sin embargo, la tasa de acumulación de lactato fue similar en normoxia y en hipoxia (Figura 1G). El índice de fatiga fue también similar en normoxia y en hipoxia (Figura 2A). La contribución anaeróbica al gasto energético en la condición de hipoxia fue mayor que en la condición de normoxia. Sin embargo, este efecto fue significativo solamente en los ciclistas de resistencia (p<0.05).
DISCUSION
El efecto de la hipoxia severa aguda, equivalente a una altura de ~5300 m, sobre el metabolismo del ejercicio y el rendimiento ha sido estudiado en ciclistas de pista de elite entrenados en resistencia y en velocidad. Este estudio demuestra que la hipoxia severa aguda no altera el pico de potencia y el índice de fatiga durante el test de Wingate de 30 s. En contraste, la PMEDIA es reducida con la hipoxia severa aguda en los ciclistas entrenados en velocidad, pero no en ciclistas entrenados en resistencia, a pesar del hecho de que en ambos grupos el VO2 medio se redujo en un 16% durante los tests de Wingate en hipoxia. Además, se ha mostrado que, en comparación con los ciclistas de velocidad, los ciclistas entrenados en resistencia muestran una potencia aeróbica máxima superior, alcanzan menores niveles de PPICO y PMEDIA, tienen una menor deuda de O2, tienen un menor índice de fatiga, obtienen una mayor fracción del gasto energético a través de las vías oxidativas durante el test de Wingate de 30 s, y muestran menores [La] sanguíneo en la vena femoral después del test de Wingate.
Efectos de la Hipoxia Aguda Severa
En contraste con nuestra hipótesis, el rendimiento en el test de Wingate se redujo solamente con la hipoxia aguda severa en los ciclistas de velocidad, mientras que se mantuvo constante en los ciclistas de resistencia. En ambos grupos, el VO2 se redujo en la misma proporción (i.e., 16%), pero debido a que el VO2 por kilogramo de masa corporal fue mayor en los ciclistas de resistencia, la reducción absoluta de VO2 fue ligeramente más grande en los ciclistas resistencia que en los ciclistas de velocidad. Si asumimos que la eficiencia de pedaleo es similar en la normoxia y en la hipoxia aguda, la única explicación posible para estos hallazgos es una mayor liberación de energía anaeróbica bajo condiciones de hipoxia (39). El hecho de que la deuda de O2 se incremento con la hipoxia en los ciclistas de resistencia, pero no en los ciclistas de velocidad respalda este concepto. Cuando el metabolismo aeróbico no es capaz de equiparar la producción aeróbica de ATP con el consumo, varias vías metabólicas son estimuladas para suplir la producción de energía, especialmente la hidrólisis de fosfocreatina y la glucólisis. Para nuestro conocimiento, solamente McLellan y cols. (23) han investigado el efecto de la hipoxia aguda moderada sobre el metabolismo muscular durante el test de Wingate. Estos autores reportaron que, en comparación con la normoxia, la [La] muscular se duplica cuando el test de Wingate es realizado en condiciones de hipoxia aguda. Debido a que la vía glucolítica del metabolismo anaeróbico representa ~3/4 de la capacidad anaeróbica (1, 3, 33), un incremento en la acumulación de lactato de la magnitud de la reportada por McLellan y cols. (23) explicaría la mayoría de la reducción en la generación aeróbica de ATP con la hipoxia en el presente estudio.
Se ha demostrado que durante el ciclismo a intensidad máxima, la producción de lactato comienza casi con el comienzo de las contracciones musculares alcanzando concentraciones de hasta ~7 (11), ~11 (16) y ~17-29 mM (3, 16, 23, 28) en 6, 10, y 30 segundos respectivamente. A pesar de la mayor acumulación de lactato con la hipoxia, se halló que la [La] (medida en la vena femoral) durante la recuperación fue ligeramente menor bajo condiciones de hipoxia en un estudio (23) y similar a la condición de normoxia en otro estudio con el mismo grupo (22). Con un procedimiento novedoso, en el presente estudio la sangre fue recolectada de la vecindad de los músculos en contracción, debido a que el metabolismo sistémico y local del brazo puede interferir con la [La] del antebrazo. Además, por medio de la recolección de sangre cada 5 segundos hemos sido capaces de medir por primera vez la cinética de la acumulación de lactato sanguíneo en la vena femoral durante los tests de Wingate. Nuestros datos demuestran que la cinética de la acumulación de lactato sanguíneo en la vena femoral es independiente de la FIO2 durante ejercicios máximos de 30 segundos de duración. En ambas condiciones hubo un retraso de 15 segundos entre el comienzo del ejercicio y el comienzo de la acumulación de lactato sanguíneo, indicando que la mayor parte del lactato producido al comienzo del ejercicio es retenido dentro del músculo. No tenemos pistas por medio del presente estudio para explicar porque el lactato no es liberado durante los primeros 15 segundos de ejercicio. Si el factor principal que afecta la liberación neta de lactato es el pH muscular (30), el hecho de que el pH muscular apenas sufre cambios al comienzo del ejercicio podría facilitar la acumulación de lactato muscular, o quizás los transportadores musculares de lactato deban primero ser activados.
Varios estudios han mostrado que en 30 segundos no puede utilizarse la capacidad anaeróbica total (6, 7, 25, 26, 31, 33, 35). La hipoxia parece estimular la utilización adicional de la capacidad anaeróbica para compensar la reducción en la producción aeróbica de ATP. Este efecto fue claro en los ciclistas de resistencia, mientras que los ciclistas de velocidad mostraron un pequeña reducción en la deuda de O2, lo cual, en conjunto con un menor VO2, derivó en la disminución de la PMEDIA. La cuestión crítica es ¿qué limita el rendimiento durante el test de Wingate? Nuestros hallazgos están en contraste con el paradigma del predominio, el cual propone que el rendimiento durante el test de Wingate está limitado por la tasa de liberación de energía anaeróbica (2, 7), lo cual, a su vez, depende de la disponibilidad de sustratos y del control enzimático (7, 28, 35). Es sabido que el rendimiento durante el test de Wingate no está limitado por la disponibilidad de glucógeno (16, 17, 23, 26). Nuestro estudio, novedosamente, muestra que ni la disponibilidad de energía anaeróbica ni la tasa de liberación de energía anaeróbica limita el rendimiento durante el test de Wingate en ciclistas de elite entrenados en resistencia. Nuestros resultados señalan a la tasa de utilización de ATP como el factor limitante del rendimiento durante el test de Wingate tradicional. De acuerdo con nuestros resultados, los factores que reducen la tasa de hidrólisis de ATP deberían ser comunes a las vías aeróbicas y anaeróbicas. Este requerimiento es cumplido por medio del ADP y del Pi los cuales se acumulan durante ejercicios máximos y de esta manera podrían establecer el límite superior de la utilización de ATP a través de sus efectos inhibitorios sobre la contracción muscular (9, 38). En contraste, de acuerdo con nuestros resultados, el rendimiento en los ciclistas de velocidad parece estar limitado por los sustratos en condiciones de hipoxia, como se reportó en normoxia en sujetos desentrenados (35), y en sujetos físicamente activos (7).
Debería destacarse que el nivel de hipoxia utilizado en nuestro estudio está muy cercano al límite que los humanos pueden tolerar agudamente. Durante el ejercicio progresivo hasta el agotamiento bajo estas condiciones, los valores arteriales de PO2 en el momento del agotamiento se acercan a 30-35 mm Hg (36). Este nivel de hipoxia es muy cercano al límite que puede ser tolerado por un corto tiempo por humanos no aclimatados y es similar al reportado en sujetos aclimatados a la altura en la cima de Monte Everest simulada en el estudio Operación Everest II (34). La hipoxemia severa, a su vez, podría haber provocado fatiga central (27) o haber reducido el pico de flujo sanguíneo a la pierna (5). Aunque no podemos descartar un componente central en la aparición de la fatiga, de acuerdo con investigaciones previas realizadas con menores niveles de hipoxia (22, 23), en este estudio no se hallaron efectos de la hipoxia severa aguda sobre el índice de fatiga o sobre el pico de potencia. Además, los signos clínicos de fatiga y de la respuesta ventilatoria durante los tests de Wingate en hipoxia (datos no mostrados) fueron similares en ambos grupos de ciclistas. Por lo tanto, no parece que las diferencias en los mecanismos de fatiga central podrían explicar las reducciones en el rendimiento con la hipoxia en los ciclistas de velocidad.
La deuda de oxígeno, como se midió en este estudio, no es una pura estimación de la energía anaeróbica utilizada, debido a que el O2 consumido desde las reservas de O2 [especialmente el O2 ligado a la mioglobina, el cual se estimó que era de 2 mmol/kg de peso húmedo (14), y el O2 ligado a la hemoglobina] al comienzo del ejercicio es computado como deuda de O2, llevando a una sobreestimación de la producción de energía anaeróbica, y, a la inversa, a una subestimación del VO2 real (25). Este error intrínseco del método para medir la deuda de O2 es, sin embargo, bastante pequeño debido a la comparativamente baja cantidad de O2 almacenada en los músculos esqueléticos en relación con la magnitud de la deuda de O2. Si las series de ejercicio son realizadas en normoxia e hipoxia, las reservas de O2 al comienzo del ejercicio serían menores en la condición hipóxica, reduciendo la sobreestimación de la deuda de O2. La saturación de la mioglobina en reposo, sin embargo, fue probablemente muy similar en la normoxia y en la hipoxia, debido a las características especiales de la curva de disociación de la mioglobina, la cual a una presión de O2 de cercana a 3 mmHg presenta una saturación de la mioglobina del 50% (30). Con el nivel de hipoxia utilizado en este estudio, se ha reportado una PO2 arterial de reposo de 45-50 mmHg y una saturación de ~80% (32). Consecuentemente, la cantidad de O2 almacenado como O2 ligado a la hemoglobina fue probablemente un 20% menor al comienzo del test de Wingate en condiciones de hipoxia. Si este fue realmente el caso, la diferencia de la deuda de O2 entre la hipoxia y la normoxia podría haber sido todavía mayor que el 7% calculado para los ciclistas de resistencia, y quizás no se habría observado una reducción en la deuda de O2 en la condición hipoxia en los ciclistas de velocidad. Asumiendo que ambos grupos tuvieron una reducción similar en sus reservas de O2 con la hipoxia, nuestros datos demuestran que los ciclistas de resistencia tienen una mayor capacidad para aumentar la producción de energía anaeróbica en respuesta a la hipoxia que los ciclistas de velocidad. Quizás los ciclistas de velocidad ya estaban utilizando su energía anaeróbica a una tasa cercana a la máxima en normoxia, y por ello, no fueron capaces de compensar la reducción en la producción aeróbica de ATP en la condición de hipoxia por medio del aumento en la tasa de liberación de energía anaeróbica.
Diferencias entre los Especialistas en Resistencia y los Especialistas en Velocidad
Una particularidad de este estudio fue el nivel sobresaliente de los ciclistas examinados, quienes estaban entre los mejores de España con varios logros en competiciones internacionales en ambos grupos. En el caso de los ciclistas de velocidad, la PPICO y la PMEDIA estuvieron entre las más altas reportadas en la literatura (8, 12, 22, 23, 40). Debería mencionarse que la PMAX. obtenida en el cicloergómetro Monark fue similar y estuvo estrechamente correlacionada con la PMEDIA medida durante los 30s de un sprint de 500 m en pista (medida en 10 ciclistas estudiados aquí con un ergómetro SRM, datos no mostrados; Ref. 8). La PMEDIA desarrollada por estos ciclistas fue mucho más allá de la producción de la potencia aeróbica máxima, y en consecuencia, la deuda de O2 fue también más alta que la reportada en estudios previos para ciclistas (8, 13, 40). Como se esperaba, los ciclistas de velocidad tuvieron menores valores de VO2 máx., pero una duda de O2 mucho más grande que los especialistas de resistencia (8, 24). Asumiendo que los especialistas en velocidad utilizan el 80-90% de su capacidad anaeróbica en los 30s (6, 25, 31, 33), puede estimarse que su máxima deuda acumulada de O2 real está entre 75 y 85 ml/kg. De hecho, se ha sugerido que los atletas de velocidad de clase mundial pueden alcanzar deudas de O2 todavía mayores (8, 25, 31). La superioridad de los ciclistas de velocidad en la potencia y capacidad anaeróbica es probablemente el resultado de un porcentaje incrementado en las fibras tipo II, una maquinaria enzimática más apropiada para producir ATP, a través de las vías anaeróbicas, y una mayor capacidad de amortiguación (8, 13, 20, 21). Sin embargo, el índice de fatiga fue marcadamente mayor en los ciclistas de velocidad que en los ciclistas de resistencia. Debido a que los ciclistas de velocidad tienen una mayor proporción de fibras tipo II que los ciclistas de resistencia (21), los ciclistas de velocidad deberían elegir una tasa de pedaleo más rápida, ya que las fibras tipo II son más eficientes a altas velocidades de contracción (15). La elección de altas frecuencias de pedaleo puede ser ventajosa para las competiciones de velocidad en pista (37). En un interesante experimento utilizando un cicloergómetro isoquinético, Jones y cols. (17) demostraron que con 60 y 140 rpm se desarrolla la misma PMEDIA, pero el pico de potencia es considerablemente mayor a 140 rpm, con el precio, sin embargo, del incremento en el índice de fatiga.
Withers y cols. (40) reportaron que los ciclistas entrenados en resistencia pueden utilizar el 94% de su capacidad anaeróbica en evaluaciones máximas de 45 segundos. Utilizando un enfoque diferente, nosotros hemos estimado que los ciclistas de resistencia podrían utilizar la totalidad de su capacidad anaeróbica en 43 segundos mientras que los ciclistas de velocidad necesitarían 47 segundos ( ver Figura 3).
Limitaciones de la Deuda de O2 como Medición de la Producción de Energía Anaeróbica
A pesar de las limitaciones de la deuda de O2 como método para estimar la producción de energía anaeróbica, deberíamos enfatizar que nuestras estimaciones sobre la división entre las fuentes de energía aeróbica y anaeróbica durante el test de Wingate concuerdan asombrosamente bien con las tasas de turnoter o recambio de ATP reportadas por Parolin y cols. (28). Estos autores midieron las tasas aeróbicas y anaeróbicas de turnover de ATP en biopsias musculares obtenidas en tres momentos diferentes durante un test de Wingate isoquinético a 100 rpm. Durante los últimos 15 segundos del test de Wingate, la contribución aeróbica media al gasto energético fue del 54% en el estudio de Parolin y cols. y del 48% en nuestros ciclistas de resistencia. La pequeña diferencia entre nuestras estimaciones y los datos obtenidos a partir de las biopsias por Parolin y cols. reflejan probablemente el hecho de que la liberación de lactato desde el músculo no fue considerada en el trabajo de Parolin y cols. Por lo cual ellos probablemente subestimaron ligeramente la contribución anaeróbica durante los últimos 15 segundos del test de Wingate, período en el cual hemos mostrado un incremento en la concentración de lactato en la vena femoral.
Los valores reales de la deuda de O2 podrían ser menores o mayores que los reportados aquí, pero si asumimos que la eficiencia mecánica durante el test de Wingate fue similar en normoxia y en hipoxia aguda, entonces nuestra conclusión de que la producción de energía anaeróbica se incremente durante ejercicios máximos en condiciones de hipoxia para suplir la reducción en el VO2 es irrefutable.
En resumen, fueron estudiados los efectos de la hipoxia severa aguda, equivalente a una altura de ~5300 m, sobre el metabolismo del ejercicio y el rendimiento en ciclistas de pista de elite entrenados en resistencia y en velocidad. Hemos demostrado que el pico de potencia y el índice de fatiga no son alterados por la hipoxia severa aguda, mientras que el VO2 medio se redujo en un 16%, tanto en los ciclistas de resistencia como en los ciclistas de velocidad. Interesantemente, a pesar de esta marcada reducción en el VO2, solamente los ciclistas entrenados en resistencia fueron capaces de mantener la PMEDIA a través del incremento de la producción de energía anaeróbica, lo cual muestra que ni la capacidad anaeróbica ni la tasa de liberación de energía anaeróbica, limitan el rendimiento en el test de Wingate en ciclistas de resistencia. A la inversa, en los ciclistas entrenados en velocidad se observo una pequeña disminución en la PMEDIA con la hipoxia, inferior a la esperada a partir de la reducción en el VO2. Los ciclistas entrenados en resistencia, por otro lado, poseen mayores potencias aeróbicas máximas y obtienen una mayor fracción de la energía gastada durante el test de Wingate a través de las vías oxidativas, que los ciclistas entrenados en velocidad. A su vez, los ciclistas entrenados en velocidad muestran una mayor deuda de O2 (+33%), dependen más de las fuentes de energía anaeróbica, y alcanzan mayores picos de potencia y mayores PMEDIA durante los tests de Wingate. Sin embargo, los ciclistas entrenados en velocidad, desarrollan la fatiga a una tasa más rápida que los ciclistas entrenados en resistencia durante ejercicios supramáximos. En contraste con el paradigma de la predominancia, este estudio muestra por primera vez que el rendimiento durante el test de Wingate tradicional no está limitado por el suministro de energía anaeróbica en los ciclistas de resistencia.
Agradecimientos
Quisiéramos agradecer a José Navarro de Tuero, Angel Vivas y a Mecánico Mei por su excelente asistencia técnica.
Notas al Pie
Este estudio estuvo respaldado por un subsidio (08/UNI01/99) del Centro de Alto Rendimiento y de Investigación en Ciencias del Deporte, Consejo Superior de Deportes de España y por la Universidad de Las Palmas de Gran Canaria (proyectos de Infraestructura 2000).
Dirección para el pedido de reimpresiones y otra correspondencia
J. A. L. Calbet, Departamento de Educación Física, Universidad de Las Palmas de Gran Canaria, Campus Universitario de Tafira, 35017 Las Palmas de Gran Canaria, Islas Canarias, España (Correo electrónico: lopezcalbet@terra.es).
Referencias
1. Bangsbo, J, Gollnick PD, Graham TE, Juel C, Kiens B, Mizuno M, and Saltin B (1990). Anaerobic energy production and O2 deficit-debt relationship during exhaustive exercise in humans. J Physiol 422: 539-559
2. Bar-Or, O (1987). The Wingate anaerobic test. An update on methodology, reliability and validity. Sports Med 4: 381-394
3. Bogdanis, GC, Nevill ME, Boobis LH, Lakomy HK, and Nevill AM (1995). Recovery of power output and muscle metabolites following 30s of maximal sprint cycling in man. J Physiol 482: 467-480
4. Boushel, R, Calbet JA, Radegran G, Sondergaard H, Wagner PD, and Saltin B (2001). Parasympathetic neural activity accounts for the lowering of exercise heart rate at high altitude. Circulation 104: 1785-1791
5. Calbet, JA (2000). Oxygen tension and content in the regulation of limb blood flow. Acta Physiol Scand 168: 465-472
6. Calbet, JA, Chavarren J, and Dorado C (1997). Fractional use of anaerobic capacity during a 30- and a 45-s Wingate test. Eur J Appl Physiol 76: 308-313
7. Casey, A, Constantin-Teodosiu D, Howell S, Hultman E, and Greenhaff PL (1996). Creatine ingestion favorably affects performance and muscle metabolism during maximal exercise in humans. Am J Physiol Endocrinol Metab 271: E31-E37
8. Craig, NP, and Norton KI (2001). Characteristics of track cycling. Sports Med 31: 457-468
9. Dahlstedt, AJ, Katz A, and Westerblad H (2001). Role of myoplasmic phosphate in contractile function of skeletal muscle: studies on creatine kinase-deficient mice. J Physiol 533: 379-388
10. Fulco, CS, Rock PB, and Cymerman A (1998). Maximal and submaximal exercise performance at altitude. Aviat Space Environ Med 69: 793-801
11. Gaitanos, GC, Williams C, Boobis LH, and Brooks S (1993). Human muscle metabolism during intermittent maximal exercise. J Appl Physiol 75: 712-719
12. Granier, P, Mercier B, Mercier J, Anselme F, and Prefaut C (1995). Aerobic and anaerobic contribution to Wingate test performance in sprint and middle-distance runners. Eur J Appl Physiol 70: 58-65
13. Green, S, Dawson BT, Goodman C, and Carey MF (1996). Anaerobic ATP production and accumulated O2 deficit in cyclists. Med Sci Sports Exerc 28: 315-321
14. Harris, RC, Hultman E, Kaijser L, and Nordesjo LO (1975). The effect of circulatory occlusion on isometric exercise capacity and energy metabolism of the quadriceps muscle in man. Scand J Clin Lab Invest 35: 87-95
15. He, ZH, Bottinelli R, Pellegrino MA, Ferenczi MA, and Reggiani C (2000). ATP consumption and efficiency of human single muscle fibers with different myosin isoform composition. Biophys J 79: 945-961
16. Jacobs, I, Tesch PA, Bar-Or O, Karlsson J, and Dotan R (1983). Lactate in human skeletal muscle after 10 and 30s of supramaximal exercise. J Appl Physiol 55: 365-367
17. Jones, NL, McCartney N, Graham T, Spriet LL, Kowalchuk JM, Heigenhauser GJ, and Sutton JR (1985). Muscle performance and metabolism in maximal isokinetic cycling at slow and fast speeds. J Appl Physiol 59: 132-136
18. Kuipers, H, Verstappen FT, Keizer HA, Geurten P, and van Kranenburg G (1985). Variability of aerobic performance in the laboratory and its physiologic correlates. Int J Sports Med 6: 197-201
19. Lopez Calbet, JA, Armengol Ramos O, Chavarren Cabrero J, and Dorado Garcia C (1997). An anthropometric equation for the determination of the percentage of body fat in young men from the Canarian population. Med Clin (Barc) 108: 207-213
20. MacDougall, JD, Hicks AL, MacDonald JR, McKelvie RS, Green HJ, and Smith KM (1998). Muscle performance and enzymatic adaptations to sprint interval training. J Appl Physiol 84: 2138-2142
21. Mackova, E, Melichna J, Havlickova L, Placheta Z, Blahova D, and Semiginovsky B (1986). Skeletal muscle characteristics of sprint cyclists and nonathletes. Int J Sports Med 7: 295-297
22. McLellan, TM, Cheung SS, and Meunier MR (1993). The effect of normocapnic hypoxia and the duration of exposure to hypoxia on supramaximal exercise performance. Eur J Appl Physiol 66: 409-414
23. McLellan, TM, Kavanagh MF, and Jacobs I (1990). The effect of hypoxia on performance during 30s or 45s of supramaximal exercise. Eur J Appl Physiol 60: 155-161
24. Medbo, JI, and Burgers S (1990). Effect of training on the anaerobic capacity. Med Sci Sports Exerc 22: 501-507
25. Medbo, JI, Mohn AC, Tabata I, Bahr R, Vaage O, and Sejersted OM (1988). Anaerobic capacity determined by maximal accumulated O2 deficit. J Appl Physiol 64: 50-60
26. Medbo, JI, and Tabata I (1989). Relative importance of aerobic and anaerobic energy release during short-lasting exhausting bicycle exercise. J Appl Physiol 67: 1881-1886
27. Noakes, TD (2000). Physiological models to understand exercise fatigue and the adaptations that predict or enhance athletic performance. Scand J Med Sci Sports 10: 123-145
28. Parolin, ML, Chesley A, Matsos MP, Spriet LL, Jones NL, and Heigenhauser GJ (1999). Regulation of skeletal muscle glycogen phosphorylase and PDH during maximal intermittent exercise. Am J Physiol Endocrinol Metab 277: E890-E900
29. Radegran, G, and Calbet JA (2001). Role of adenosine in exercise-induced human skeletal muscle vasodilatation. Acta Physiol Scand 171: 177-185
30. Richardson, RS, Noyszewski EA, Leigh JS, and Wagner PD (1998). Lactate efflux from exercising human skeletal muscle: role of intracellular PO2. J Appl Physiol 85: 627-634
31. Saltin, B (1990). Anaerobic capacity: past, present and prospective. In: Biochemistry of Exercise VII, edited by Taylor, Gollnick, Green, Ianuzzo, Noble, Métivier, and Sutton. Champaign, IL: Human Kinetics, p. 387-412
32. Spriet, LL (1995). Anaerobic metabolism during high-intensity exercise. In: Exercise Metabolism, edited by Haargreaves M. Champaign, IL: Human Kinetics, p. 1-39
33. Sutton, JR, Reeves JT, Wagner PD, Groves BM, Cymerman A, Malconian MK, Rock PB, Young PM, Walter SD, and Houston CS (1988). Operation Everest II: oxygen transport during exercise at extreme simulated altitude. J Appl Physiol 64: 1309-1321
34. Trump, ME, Heigenhauser GJ, Putman CT, and Spriet LL (1996). Importance of muscle phosphocreatine during intermittent maximal cycling. J Appl Physiol 80: 1574-1580
35. Van Hall, G, Calbet JA, Sondergaard H, and Saltin B (2001). The reestablishment of the normal blood lactate response to exercise in humans after prolonged acclimatization to altitude. J Physiol 536: 963-975
36. Van Ingen Schenau, GJ, de Koning JJ, and de Groot G (1992). The distribution of anaerobic energy in 1000 and 4000 metre cycling bouts. Int J Sports Med 13: 447-451
37. Westerblad, H, Dahlstedt AJ, and Lannergren J (1998). Mechanisms underlying reduced maximum shortening velocity during fatigue of intact, single fibres of mouse muscle. J Physiol 510: 269-277
38. Weyand, PG, Lee CS, Martinez-Ruiz R, Bundle MW, Bellizzi MJ, and Wright S (1999). High-speed running performance is largely unaffected by hypoxic reductions in aerobic power. J Appl Physiol 86: 2059-2064
39. Withers, RT, Sherman WM, Clark DG, Esselbach PC, Nolan SR, Mackay MH, and Brinkman M (1991). Muscle metabolism during 30, 60 and 90s of maximal cycling on an air-braked ergometer. Eur J Appl Physiol 63: 354-362