
Metabolismo energético durante un ejercicio de fuerza: influencia de la carga
Publicado 11 de abril de 2013, 3:45

Queremos con esta nueva entrada de blog, iniciar una serie de publicaciones con una temática relacionada con el metabolismo muscular durante los esfuerzos de entrenamiento de la fuerza. La utilización de sustratos, metabolitos que se producen y la respuesta adaptativa a estos cambios en la fisiología muscular durante este tipo de esfuerzos. Esfuerzos que son realizados por el común de la gente que concurre a un gimnasio en búsqueda de una buena condición física y mejorar su salud en general.
El ejercicio de fuerza regular es un componente esencial de los programas de intervención eficaces para mejorar la fuerza tanto en atletas como en adultos con enfermedades crónicas e invalideces. La respuesta a un programa de entrenamiento de la fuerza depende finalmente de las adaptaciones metabólicas y morfológicas pronunciadas de las múltiples funciones celulares que dependen a su vez, de la gran magnitud de los cambios de una red de la señalización compleja que está envuelta durante cada sesión de entrenamiento en respuesta a la actividad contráctil.
Investigadores del Departamento de la Educación física de la Universidad de Ciencias Deportiva del País Vasco y el Departamento de Ciencias de la Salud de la Universidad de Navarra en España, han examinado esa cuestión tan recurrente del entrenamiento del a fuerza de entrenar al "fallo o no fallo", recientemente desde un lado diferente que la mayoría de los artículos antes vistos.
La respuesta adaptativa al entrenamiento de la fuerza puede ser diferente cuando el entrenamiento llevado hasta el fallo se compara a cuando no se lleva hasta el fallo, ya que diferentes grados de fatiga y acumulación de metabolitos musculares son provocados por el entrenamiento.
Estas dos sesiones de ejercicio se usan tradicionalmente para alcanzar los objetivos del entrenamiento específico. Así, el ejercicio 'llevado hasta el fallo' se caracteriza por una progresiva disminución en la carga y la potencia a lo largo de las series repetidas y se usa principalmente para aumentar la fuerza muscular e hipertrofia. El ejercicio 'llevado no hasta al fallo' se caracteriza por el mantenimiento de la carga y la potencia promedio a lo largo de las series y se usa principalmente para optimizar el desarrollo de la potencia muscular.
Así, analizando diferentes condiciones relacionadas a los cambios en la potencia pueden proveer algunos indicios para la comprensión de los mecanismos por los cuales el proceso de la contracción muscular intenta mantener una función adecuada durante el ejercicio de fuerza dinámico.
Seis voluntarios varones sanos participaron en el estudio. Sus valores promedios (±SD) de edad, altura, peso corporal, índice de masa corporal, consumo máximo de oxígeno estimado (VO2máx) en cicloergómetro y fuerza máxima (1RM) durante el ejercicio de press de piernas bilateral, eran de 34±6 años, 179±5 cm, 74.5±7.2 kg, 23.3±1.7 Kg·m-1, 57.1±4.9 ml·kg-1·min-1 y 199±43 kg, respectivamente. Todos eran atletas entrenados, principalmente en eventos de resistencia, pero ninguno se entrenaba para la competición. El porcentaje medio de fibras de contracción lenta (ST) para estos sujetos era de 65±12%.
Para eliminar cualquier posible efecto de factores que produzcan alguna confusión, distintas variables como la carga de inicio y el número total de repeticiones fueron controladas igualando sus valores entre las 2 sesiones de ejercicio. Cada sujeto participó, en días separados, de los dos experimentos en los que realizaron 50 repeticiones con la misma carga inicial. Esta carga inicial (es decir, 154+31 Kg u 83% 1RM, la carga más pesada que podría ser desplazada correctamente una sola vez, usando únicamente una técnica correcta) era el mayor peso que podía ser posible completar 10 repeticiones hasta el fallo (10RM) del ejercicio de press de piernas.
En un día experimental (protocolo 'hasta el fallo'), los sujetos realizaron 5 series 10 repeticiones (10REP) hasta el fallo, separadas por 2 minutos de pausa entre cada serie. En este día experimental, los sujetos pudieron terminar todas las 10 repeticiones con la carga inicial durante la primer serie. Sin embargo, a veces, los sujetos no podían levantar la carga inicial durante las siguientes series debido a la fatiga. Siempre que los sujetos no pudieran levantar la carga, se disminuía por 15 Kg, permitiéndoles así completar el experimento (50 repeticiones). En el otro día experimental (protocolo 'no hasta el fallo'), los sujetos realizaron 10 series de 5 repeticiones (5REP) no hasta el fallo con la misma carga inicial como en el protocolo (10REP), separados por 2 minutos de pausa entre cada serie. Cinco repeticiones por serie fueron escogidas durante este día experimental 'no al fallo' porque ése es el número de repeticiones máximo en el que la producción de potencia máxima puede mantenerse o ligeramente (10–15%) cae durante una serie de 10 repeticiones hasta el fallo en el ejercicio de extensión de piernas. En el formato 5REP todos los sujetos pudieron terminar el protocolo completamente con la carga inicial y mantener la producción de potencia promedio a lo largo de las series repetidas. En el formato 10 REPS, sin embargo, todos los sujetos disminuyeron la producción de la potencia promedio a lo largo de las series.
Los experimentos se llevaron a cabo en el mismo momento del día separadamente de uno a dos meses. Ningún cambio se observó en los sujetos en la fuerza del press de piernas máximo (1RM) entre el primero (194+25 Kg) y el segundo (185+32 Kg) día experimental. Para evitar la perturbación de los sujetos, se les dijo que registraran su dieta normal las 48 horas anteriores al primer día experimental y repetir la misma dieta para el segundo día experimental. La duración de cada repetición en sus fases concéntricas y excéntricas fue registrada. Las repeticiones eran separadas por pausas de 1 segundo para prevenir la mejora del rendimiento del ciclo de estiramiento-acortamiento.
Se tomaron biopsias musculares del muslo para el análisis de sustratos (Cr, PCr, IMP ATP), muestras de sangre para el análisis de metabolitos y enzimas (lactato y CK), se tomaron medidas de potencia y de producción de fuerza. El cargo de energía celular (una medida de la magnitud en la cual, el 'pool' de nucleótido de adenina total de la célula (ATP, ADP y AMP) es el fosforalizada) era calculado como el cociente de (ATP + 1/2 ADP)/(ATP + ADP + AMP) y así se cuantifica la proporción de energía utilizable a energía utilizada en las muestras musculares.
Resultados
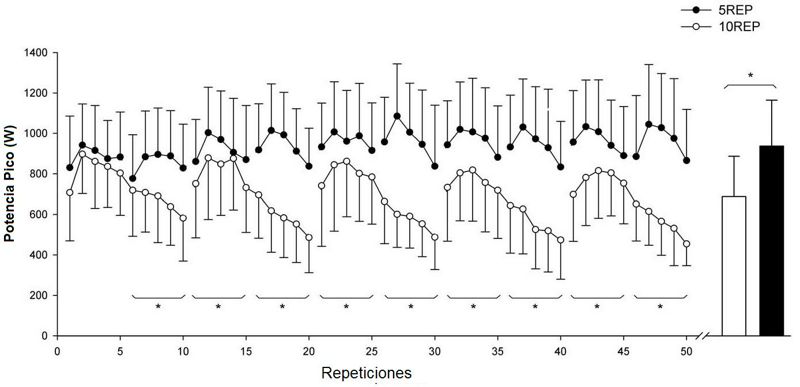 Figura 1. Perfiles de la
producción de la potencia pico (promedio para n = 6 sujetos) para cada
ejercicio durante las dos condiciones experimentales: cuando el ejercicio era 5
series de 10 repeticiones hasta el fallo (10REP; círculos blancos), y cuando el
ejercicio era 10 series de 5 repeticiones no hasta (5REP; círculos negros). Las
barras representan el promedio de la producción de la potencia pico a lo largo
de las 50 repeticiones para 10REP y 5REP. * diferencia significativa
(P<0.05) entre 10REP y 5REP (agrupados de 5 a 5 repeticiones). Los valores
son promedios ±SD.
Figura 1. Perfiles de la
producción de la potencia pico (promedio para n = 6 sujetos) para cada
ejercicio durante las dos condiciones experimentales: cuando el ejercicio era 5
series de 10 repeticiones hasta el fallo (10REP; círculos blancos), y cuando el
ejercicio era 10 series de 5 repeticiones no hasta (5REP; círculos negros). Las
barras representan el promedio de la producción de la potencia pico a lo largo
de las 50 repeticiones para 10REP y 5REP. * diferencia significativa
(P<0.05) entre 10REP y 5REP (agrupados de 5 a 5 repeticiones). Los valores
son promedios ±SD.
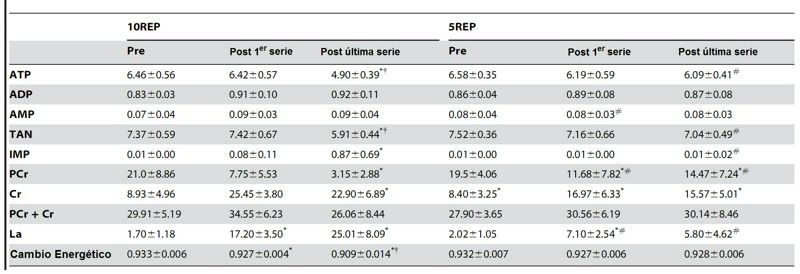 Tabla 1. Efectos del ejercicio de press de
piernas sobre los nucleótidos de adenina del músculo, IMP, PCr, Cr, lactato y
cargo de energía antes de la primer serie, después de la última repetición de
la primer serie y después de la última repetición de la última serie, durante
10REP y 5REPS. Los valores se expresan
como promedios SD en mmol·kg-1 de músculo húmedo, excepto el cargo de energía;
n = 426. TAN, nucleótidos de adenina totales (ATP + ADP + AMP); IMP, Inosina
5'-monofosfato; PCr, fosfocreatina; Cr, creatina; La, lactato. * diferencia significativa (P<0.05) con el
valor de pre-ejercicio. †diferencia significativa (P<0.05) con el
valor post de la primer serie. # diferencia
significativa (P<0.01-0.05) con el ejercicio 10REP.
Tabla 1. Efectos del ejercicio de press de
piernas sobre los nucleótidos de adenina del músculo, IMP, PCr, Cr, lactato y
cargo de energía antes de la primer serie, después de la última repetición de
la primer serie y después de la última repetición de la última serie, durante
10REP y 5REPS. Los valores se expresan
como promedios SD en mmol·kg-1 de músculo húmedo, excepto el cargo de energía;
n = 426. TAN, nucleótidos de adenina totales (ATP + ADP + AMP); IMP, Inosina
5'-monofosfato; PCr, fosfocreatina; Cr, creatina; La, lactato. * diferencia significativa (P<0.05) con el
valor de pre-ejercicio. †diferencia significativa (P<0.05) con el
valor post de la primer serie. # diferencia
significativa (P<0.01-0.05) con el ejercicio 10REP.
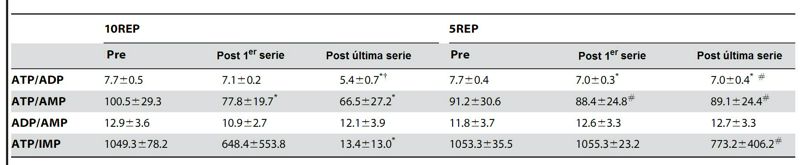 Tabla 2. Efectos del ejercicio de press de
piernas sobre las proporciones de metabolitos de los nucleótidos antes de la
primer serie y durante los protocolos 10REP y 5REP. Los valores se expresan como promedios SD. * diferencia significativa (P<0.05) con el
valor de pre-ejercicio. † diferencia significativa (P<0.05) con el
valor post de la primer serie. # diferencia
significativa (P<0.01-0.05) con un ejercicio de 10REP.
Tabla 2. Efectos del ejercicio de press de
piernas sobre las proporciones de metabolitos de los nucleótidos antes de la
primer serie y durante los protocolos 10REP y 5REP. Los valores se expresan como promedios SD. * diferencia significativa (P<0.05) con el
valor de pre-ejercicio. † diferencia significativa (P<0.05) con el
valor post de la primer serie. # diferencia
significativa (P<0.01-0.05) con un ejercicio de 10REP.
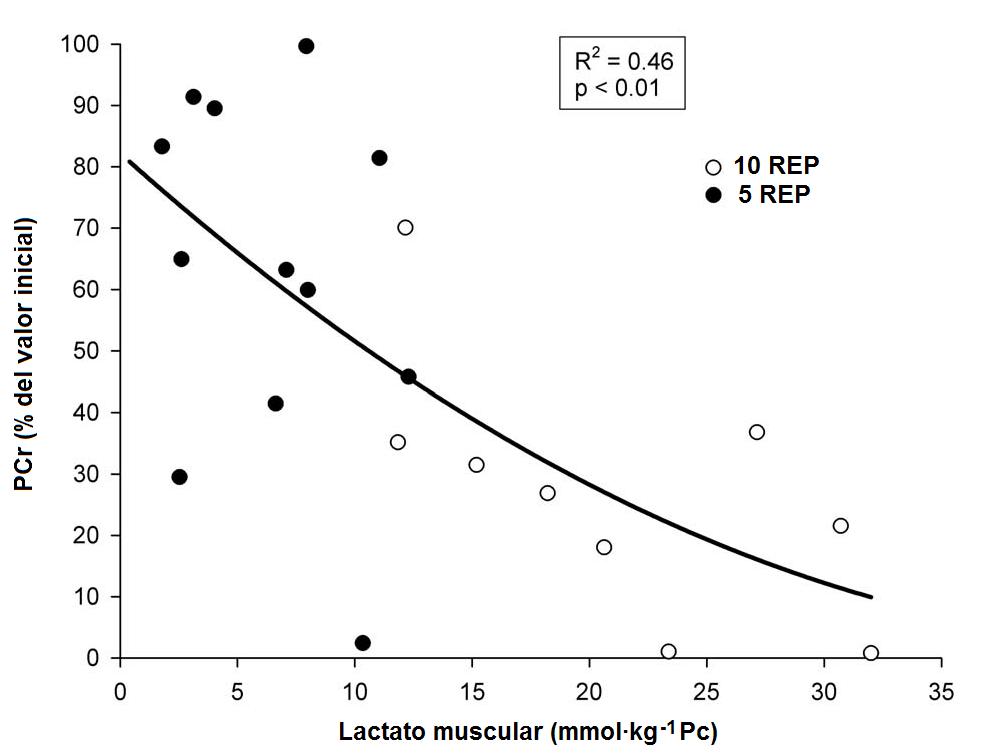 Figura 2. Relación individual entre las
concentraciones del lactato muscular y las concentraciones de PCr muscular
(expresado en porcentaje del valor inicial).
Figura 2. Relación individual entre las
concentraciones del lactato muscular y las concentraciones de PCr muscular
(expresado en porcentaje del valor inicial).
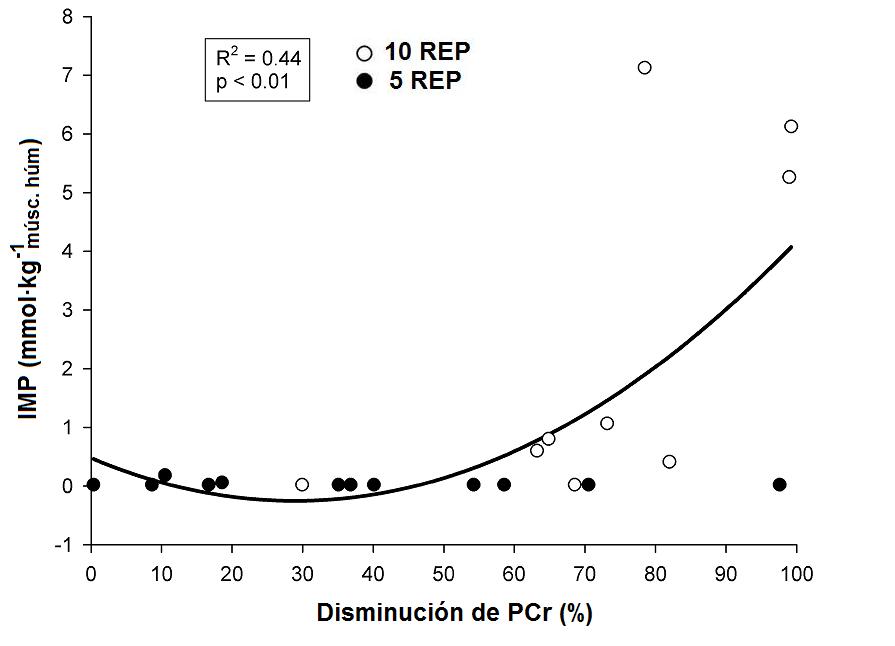 Figura 3. Relación individual entre las disminuciones
de PCr (expresado en porcentaje del valor inicial) y las concentraciones de IMP
del músculo, durante 10REP (círculos blancos), y 5REP (círculos negros).
Figura 3. Relación individual entre las disminuciones
de PCr (expresado en porcentaje del valor inicial) y las concentraciones de IMP
del músculo, durante 10REP (círculos blancos), y 5REP (círculos negros).
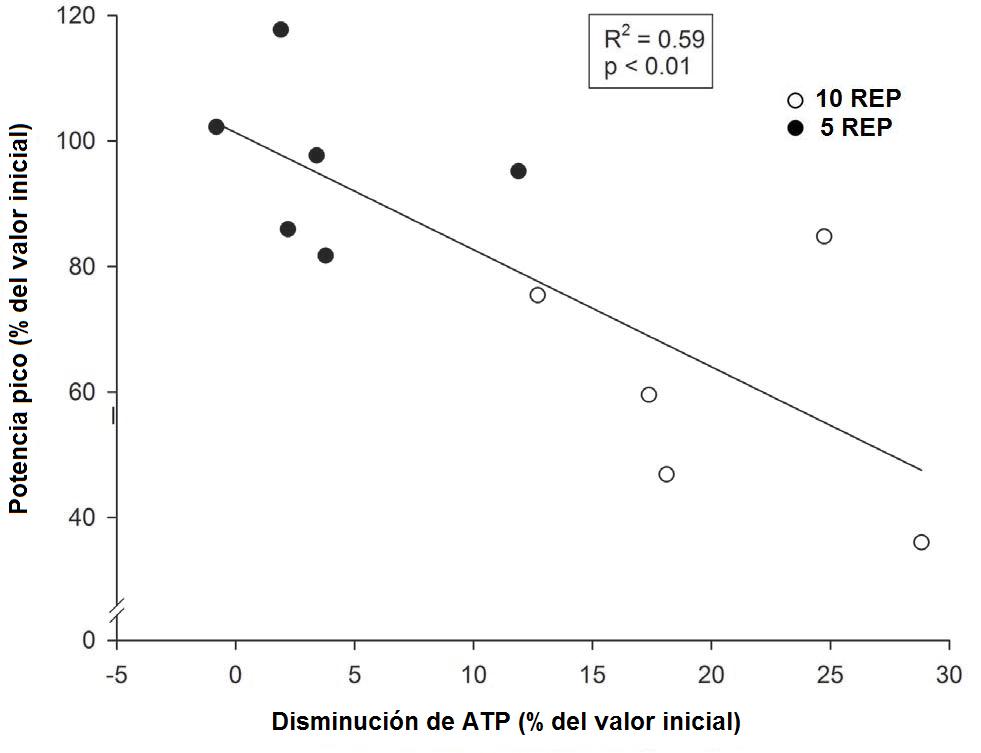 Figura 4. Relaciones individuales entre los cambios de la producción
promedio de la potencia pico relativa (expresados en porcentaje del valor
inicial) entre la primera y las últimas dos repeticiones de la primer serie y
entre la primera y las últimas dos repeticiones del ejercicio, y las
concentraciones de lactato muscular, durante 10REP (círculos blancos), y 5REP
(círculos negros).
Figura 4. Relaciones individuales entre los cambios de la producción
promedio de la potencia pico relativa (expresados en porcentaje del valor
inicial) entre la primera y las últimas dos repeticiones de la primer serie y
entre la primera y las últimas dos repeticiones del ejercicio, y las
concentraciones de lactato muscular, durante 10REP (círculos blancos), y 5REP
(círculos negros).
Por lo tanto, tenemos que correlaciones significativas (P<0.01) entre
los cambios en la PCr del músculo y el lactato muscular (R2 = 0.46),
entre los cambios en la PCr del músculo y el IMP (R2 = 0.44), así
como entre los cambios en la potencia y los cambios en los niveles del ATP del
músculo (R2 = 0.59) y del lactato muscular (R2 = 0.64).
El ejercicio de press de piernas 10REP exigió un esfuerzo máximo por parte de los sujetos, como se ve reflejado por la marcada disminución en la potencia que tuvo lugar durante las últimas 5 repeticiones de cada serie. El uso de este modelo de 10REP causó un estado de deficiencia de energía y una disminución en el fosfato, ocasionando un vaciamiento casi completo de las reservas de PCr, una reducción significativa en el ATP (21%) y del tamaño del 'pool' de nucleótido de adenina total muscular (TAN), y en marcados aumentos en la acumulación de lactato y del IMP en el músculo así como los altos niveles de ácido úrico en la sangre. Los niveles elevados de lactato muscular indican que la glucólisis anaeróbica es extensivamente activada durante este tipo de ejercicio. El IMP del músculo mayor se origina de la degradación del ATP y parece reflejar el fallo de la resíntesis de ATP para coincidir las tasas de hidrólisis de ATP [13]. La acumulación elevada de ácido úrico en plasma después de las 10RP indica que el IMP no volvió a deaminarse a AMP después del ejercicio, y que había un desfosforilación del IMP en cambio a inosina, y por consiguiente, a hipoxantina y ácido úrico [Dado que la desaminación del AMP no es reversible en condiciones fisiológicas, la recuperación del contenido de los nucleótidos de adenina requiere el proceso de reaminación de IMP a AMP, catalizado por las enzimas adenilosuccinato sintetasa y adenilosuccinato liasa. En el proceso de reaminación, el grupo amino lo aporta el aspartato, consumíendose energñia a partir del GTP (trifosfato de guanosina), por lo que un ciclo completo conlleva la desaminación de una molécula de aspartato y la producción de amonio y fumarato. Estos proceso de desaminación y reaminación no son simultáneos durante la contracción muscular, ya que la desaminación del AMP se produce durante el ejercicio y la reaminación durante el período de recuperación (López Chicharro)].
El resultado neto habría sido una pérdida antieconómica de purinas del músculo que podría requerir el reemplazo por el ciclo de los nucleótidos purinas o la síntesis de nuevo. La magnitud de la producción de energía anaeróbica y la caída en la producción de la potencia pico con 10REP son más altas que los cambios encontrados previamente en culturistas usuarios de esteroides anabólicos que siguen un régimen de ejercicio que comprende 5 series de 10RM cada una de sentadillas frontal, sentadillas con barra atrás, press de piernas y extensiones de rodillas (como en un estudio de Tesch, 1986).
Sin embargo, es cualitativa y cuantitativamente similar a lo que ha sido reportado previamente por otros y después del pedaleo de alta intensidad intermitente, ejercicio isométrico de rodilla, o isocinético que llevan al agotamiento entre 6" y 60". Estos cambios también han sido asociados con reducciones grandes en el glucógeno muscular y ATP, particularmente en las fibras Tipo II, con cambios más pronunciados localizándose en el tipo de fibras IIx, mientras que el pH sanguíneo y muscular pueden alcanzar valores tan bajos como 7.1 y 6.6, respectivamente. Sin embargo, el protocolo experimental en los estudios previos no fue igual que en el presente estudio de Gorostiaga, ya que se usaron contracciones isoinerciales repetitivas en las acciones del press de piernas. Así, parece ser que 10REP causa una marcada ruptura en el balance de energía en el músculo.
En contraste a 10 repeticiones con una carga de 10MR (10REP), el balance de energía se mantuvo durante 5 repeticiones con una carga de 10MR (5REP), a pesar de la acumulación de series. Así, comparado con 10REP, el formato 5REP resultó en una notablemente disminución mucho menor en el contenido de PCr muscular (,15% vs 80% de caída) con solamente aumentos modestos en el lactato muscular y ningún cambio mensurable en los niveles musculares de ATP y IMP y en los niveles sanguíneos de ácido úrico. Estos cambios menores de los metabolitos observados en 5REP, a pesar de la alta tasa de recambio de ATP, indican que la síntesis de ATP igualó la utilización de ATP y esa homeostasis celular se mantuvo, demostrando, por consiguiente, que la tasa de desaminación de AMP fue baja.
Por lo tanto, la producción de la potencia pico sólo se mantuvo durante el formato 5REP. La ausencia de aumento del ácido úrico sanguíneo durante 5REP, está en línea con el hallazgo de los niveles de ATP inalterados. Estos resultados indican que reduciendo el número de repeticiones por serie al 50%, mantiene la potencia y el balance de energía en el músculo a lo largo de las series.
De importancia son las relaciones entre ATP/ADP y ATP/AMP, ya que los niveles de estos nucleótidos son importantes reguladores del metabolismo muscular. Al final de 5REP, la proporción de ATP/ADP calculada disminuyó un 9% comparado con los valores iniciales, y fue coincidente sin cambios en el cargo de energía calculada, relaciones de AMP, ATP/AMP y ATP/IMP, con una disminución de 7–20% en la potencia pico promedio entre la segunda y la quinta repetición de cada serie, y con disminuciones moderadas (30%) en los niveles de PCr. La dicotomía entre los cambios en la proporción de ATP/ADP y cambios en la proporción de ATP/AMP durante el ejercicio no tiene precedentes. Así, se han observado en otros estudios, aumentos en el contenido muscular de ADP libre estimado, sin ningún cambio en el AMP libre estimado, durante 15 segundos de sprint en un pedaleo isocinético o después de 10 minutos de pedalear al 65% VO2máx, indicando que un aumento inicial ligero en la disponibilidad del ADP no cambia de sitio a la adenilato kinasa, favoreciendo la formación de AMP (ADP + ADP<>AMP + ATP), cuando la disminución en la producción de la potencia y los cambios en PCr del músculo o los niveles de Pi, son moderados. La actividad de la adenilato kinasa fue baja aún después de 5REP, probablemente porque la desfosforilación del ADP al AMP es 'tamponada' por la PCr. En esta situación la proteína kinasa AMP-activada (AMPK) sólo puede activarse ligeramente por la mayor disponibilidad del ADP. Al final de 10REP, sin embargo, una disminución mayor (30%) en la proporción de ATP/ADP, más alta que la vista después de 5REPS, fue acompañada por las disminuciones en las proporciones de ATP/AMP y de ATP/IMP, en paralelo con mayores disminuciones (33%) en la producción de la potencia y con los niveles de PCr casi depletadas. Las menores proporciones de ATP/AMP y de ATP/IMP indican que, opuesto a lo ocurrido en 5REP, durante el formato 10REP, la adenilato kinasa y la deaminasa de AMP fueron significativamente activadas. Los niveles muy bajos de PCr y la activación de la adenilato kinasa podrían amplificar la activación de la AMPK, que actúa como enzima 'sensora' de combustible que supervisa el nivel de energía celular para prevenir las grandes consecuencias de las disminuciones mayores en el estado de energía. Este mecanismo dual (disminuciones iniciales en la proporción de ATP/ADP seguida por disminuciones tardías en las proporciones de ATP/AMP y de ATP/IMP) permitiría a la AMPK dar cuenta del déficit de energía progresivamente en una amplia gama de disponibilidad de energía.
Las diferencias descriptas relacionadas con las
repeticiones sobre la respuesta metabólica aguda a series repetidas del press
de piernas pueden reflejar dos estímulos diferentes para adaptaciones inducidas
por el entrenamiento que ocurren después del entrenamiento pesado de fuerza.
Así, algunos estudios han demostrado que el entrenamiento de la fuerza de alta
intensidad no hasta el fallo de los músculos extensores de la rodilla produce
un ambiente favorable para lograr mayores mejoras en la producción de fuerza y
la potencia máxima comparado al entrenamiento hasta el fallo. Tomado juntos,
los resultados de esos estudios y el presente indican que un programa de
ejercicio de fuerza de extensión de rodillas dinámico caracterizado por una
baja acumulación de metabolitos y un mantenimiento de la homeostasis celular y
un balance de energía, puede ser una opción más eficaz y segura, comparada
hasta al entrenamiento hasta el fallo, diseñado para maximizar la acumulación
de fatiga/metabolitos . Bajo esta suposición, sería bueno de reemplazar la
clásica filosofía del entrenamiento de 'sin dolor, no hay ninguna ganancia', a
una más racional y más basada en 'ningún dolor, más ganancia'.
Relaciones entre los
metabolitos musculares. Una significativa relación curvilínea negativa se
observó a lo largo de los ejercicios entre la disminución porcentual de la PCr
del músculo y el contenido de lactato muscular correspondiente. Es más, indica
que los cambios entre la PCr y el lactato ocurren principalmente en simultáneo
durante el ejercicio, apoyando las observaciones de otros autores donde la
glucólisis anaeróbica se inicia en el músculo al comienzo del ejercicio pesado.
En acuerdo con nuestros resultados, la acumulación de IMP igualada
cuantitativamente por una disminución en el ATP intramuscular, ha sido
reportada durante el ejercicio de pedaleo submáximo y máximo cuando los niveles
de PCr se caen por debajo del 40% de los niveles de reposo.
¿Qué causa la fatiga
durante el ejercicio de press de piernas de series consecutivas? Varios autores han indicado que la capacidad para regenerar ATP a las
tasas requeridas y así, disminuir la producción de fuerza y potencia durante el
ejercicio máximo de corta duración, puede estar relacionado a una incapacidad
para mantener la tasa requerida de producción anaeróbica de ATP a partir de la
degradación de PCr y del glucógeno, principalmente en las fibras tipo II, un
aumento correspondiente en el fosfato inorgánico (Pi) y sus formas
diprotonadas, H2PO4-, aumentos en [H+], alteraciones en el transporte de Ca2+,
flujo de K+ desde el músculo o transmisión neuromuscular anormal o fallo de
excitación de la membrana. En el presente estudio de Gorostiaga, la caída en la
producción de la potencia durante ambos ejercicios en conjunto se correlacionó
fuertemente a la caída en las reservas de ATP y a los niveles de lactato en
músculos mixtos homogeneizados. La asociación entre los cambios en la potencia
y los cambios en la acumulación de lactato muscular está conforme con estudios
anteriores. La asociación observada entre la pérdida de las reservas de ATP y
la disminución relativa en la potencia, apoya la idea de que el vaciamiento de
ATP en un porcentaje pequeño de fibras puede llevar a su fallo para la
producción de potencia muscular. Esta idea está en coincidencia con algunos
estudios que reportan niveles de ATP post-ejercicio en fibras individuales, tan
bajos como 1 a 2.4 mmol·Kg-1 (de músculo húmedo) después de un ejercicio de
extensión de rodillas máxima, o de pedaleo isocinético. De allí, no es
irrazonable indicar que algunos cambios bioquímicos, como una disminución en
las reservas de ATP y aumentos en el lactato y derivados del ATP (H+, Pi, ADP)
de fibras musculares individuales puedan contribuir a la fatiga durante el
ejercicio de press de piernas de series sucesivas.
Aunque el estudio es limitado por el número de sujetos examinados, las relaciones entre los metabolitos es muy fuerte. También, puede argumentarse que el diseño experimental pudo fortalecerse coincidiendo la misma carga a lo largo de las series y el período de pausa total entre los protocolos (por ejemplo, la pausa de 1 minuto entre las series en el formato 5REP y pausa de 2 minutos entre las series en el formato 10REP). Esto habilitaría comparaciones entre dos tipos de ejercicio de igual volumen. Es obvio que, en este caso, la magnitud de las diferencias entre los ejercicios sería inferior que lo observado en el presente estudio. Sin embargo, tiene que ser recalcado que el objetivo principal era comparar los cambios metabólicos y cambios de la potencia muscular durante dos tipos de series de un ejercicio de fuerza tradicionalmente usados para alcanzar los resultados del entrenamiento específico ('hipertrofia' versus 'mejora de fuerza óptima'), pero no para comparar dos ejercicios de igual volumen. Finalmente, este estudio se realizó en hombres entrenados en resistencia con una proporción alta (65%) de fibras tipo I. Debe tenerse en cuenta al generalizar los resultados de este estudio a otras poblaciones (por ejemplo, atletas de fuerza con alta proporción de fibras tipo II). A pesar de estas limitaciones, los resultados proveen importante y nueva información sobre las características metabólicas de dos formas de entrenar la fuerza de alta intensidad, un tipo de diseño muy popular para aumentar la fuerza, potencia e hipertrofia muscular en atletas y adultos con enfermedades crónicas e invalideces.
En resumen, podemos decir que los resultados principales de este estudio fueron, primero, que reduciendo el número de repeticiones de diez a cinco durante las series, manteniendo la misma carga inicial y períodos de recuperación entre ellas, indujeron demandas notablemente menores sobre el sistema de los fosfatos altos en energía y sobre el aporte de energía glucolítica, permitió la síntesis de ATP igualar la utilización de ATP, mantener la potencia y el balance de energía a lo largo de las series y experimentar mucho menos fatiga e incomodidad durante el ejercicio de press de piernas bilateral. Segundo, en músculos homogeneizados mixtos, los cambios en la PCr muscular durante ambas series de ejercicio, se correlacionó con el lactato muscular y el contenido de IMP. Estas correlaciones indican que los cambios entre la PCr y el lactato muscular ocurren principalmente en forma simultánea, mientras que el IMP solamente se acumula cuando los niveles de PCr son bajos. Finalmente, la caída en la producción de la potencia se correlacionó fuertemente a la caída en las reservas de ATP y a los niveles del lactato muscular. Esto indica que una disminución en las reservas de ATP y aumentos en el lactato y derivados del ATP (H+, Pi, ADP) de fibras musculares individuales podrían contribuir a la fatiga durante el ejercicio de press de piernas de series sucesivas.
Para tener en cuenta. A primera vista se puede decir ciertamente que el estudio no nos da mucha nueva información. Si se comparan los resultados principales con la visión común acerca de varios regímenes del entrenamiento de la fuerza, el vaciamiento total de las reservas de PCr en el grupo de las repeticiones más altas y el aumento en los niveles de IMP, de lo que Gorostiaga y cols. debidamente argumentan que dichos resultados reflejan el fallo de la resíntesis de ATP para coincidir con las tasas de hidrólisis de ATP y eventualmente alimentar el ciclo del ácido úrico (como se refleja por el 19% de aumento en el grupo que entrenó hasta el fallo), pone un énfasis sobre la demanda energética y metabólica a menudo subestimada del entrenamiento hasta el fallo.
Bibliografía:
José López Chicharro, Almudena Fernández Vaquero. Fisiología del ejercicio. Editorial Médica panamericana, (3ª ed.) 2006.
Gaitanos GC, Williams C, Boobis LH, Brooks S. Human muscle metabolism during intermittent maximal exercise. J Appl Physiol. 1993;75(2):712-9.
Bogdanis GC, Nevill ME,
Boobis LH, Lakomy HKA. Contribution of phosphocreatine and aerobic metabolism to
energy supply during repeated sprint exercise. J Appl Physiol. 1996;80:876-84.