
Monohidrato de Creatina
Publicado 8 de julio de 2013, 3:03
La creatina (Cr) fue
descubierta en 1832 por el científico francés Michel Eugene Chevreul quien
extrajo un nuevo constituyente orgánico de la carne y lo denominó Creatina,
nombre proveniente del griego κρέας = kreas carne (Williams et al. 1999).
La Cr (CAS 57-00-1, 2006) es
un metabolito semejante en su dimensión molecular a un aminoácido (Figura 1), con una masa molecular de 131.13 g mol-1
y un punto de fusión es 303 °C (Pischel & Gastner 2007).

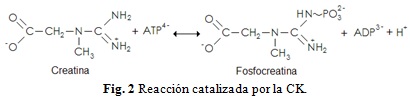
La síntesis endógena de Cr se fundamenta primero en la transferencia del grupo amidino de la arginina al grupo amino de la glicina, produciendo ornitina y guanidinoacetato, reacción catalizada por la enzima L-Arginina:Glicina Amidinotransferasa (AGAT – EC 2.1.4.1) (Brosnan & Brosnan 2007). El guanidinoacetato es metilado sobre el nitrógeno original de la glicina usando S-adenosilmetionina (AdoMet) como el donador del grupo metilo. Esta reacción produce Cr y S-adenosilhomocisteína (AdoHcy) y la cataliza la enzima Guanidinoacetato N-Metiltransferasa (GAMT – EC 2.1.1.2) (da Silva et al. 2009; Stout et al. 2008). La síntesis de Cr representa una considerable carga metabólica, en particular con respecto a la utilización de aminoácidos. Es relevante la relación de la síntesis de Cr con la ingesta diaria de glicina, arginina y metionina. La síntesis de 1.0 g Cr día-1 tiene un valor de 7.6 mmol, esto se compara a la ingesta diaria de cerca de 78 mmol de glicina, 37 mmol de arginina y 19 mmol de metionina (estos valores están basados en la ingesta de 80 g proteína día-1). Por consiguiente la síntesis de Cr consume cerca del 10% de la glicina dietaria ya que una molécula entera de glicina es incorporada a una molécula de Cr (Brosnan et al. 2007). En humanos, el hígado es el órgano más importante que contribuye a la biosíntesis de Cr mientras que el riñón juega solo un papel secundario. Ambos tejidos presentan gran actividad de AGAT y GAMT, aunque en el hígado la actividad es mayor (Brosnan & Brosnan 2007). Además, la actividad de AGAT ha sido detectada también en corazón, pulmón, bazo, músculo, cerebro, testículos y timo, donde se ha estimado que la cantidad total de AGAT en estos tejidos es cercana a las encontradas en riñón y páncreas (Wyss & Kaddurah-Daouk 2000; Stout et al. 2008). La Cr es degradada espontáneamente en una reacción monomolecular a creatinina (Crn, Mr 113.1), la cual es cuantitativamente excretada en la orina (Uzzan et al. 2009). La taza de excreción está estimada más o menos en el 1.7% del pool de Cr total (TCr) en el cuerpo por día. Como más del 90% de las moléculas de Cr y PCr se encuentran en el músculo esquelético, la excreción de Crn varía en función del género y la edad; así, la taza de excreción de Crn tiene su máximo nivel entre los 18 y 29 años, con una relación de 23.6 mg Crn por kilogramo de peso cada 24 horas (Brosnan & Brosnan 2007). De esta manera, el requerimiento diario de Cr ya sea por la dieta o por la síntesis endógena en un hombre de 70 Kg es aproximadamente de 2 g día-1 (Harris et al. 1992). Sin embargo, puesto que muchos atletas pesan más de 70 Kg y ya que el entrenamiento intenso promueve la degradación proteica, incrementando los niveles de Crn en suero y orina, atletas que llevan a cabo entrenamiento intenso tienen un mayor turnover y requerimiento diario de Cr (por ejemplo de 2 a 3 g día-1) (Williams et al. 1999). La Cr se encuentra primordialmente en los vertebrados, y puesto que se concentra principalmente en el tejido muscular, las fuentes primarias son el pescado y las carnes rojas, aunque trazas de Cr pueden ser encontradas en algunas plantas (Balsom et al. 1994). Ahora bien, la Cr es requerida principalmente por células con requerimientos energéticos altos y variables (Wallimann et al. 2011); sin embargo, los tejidos que almacenan las más altas concentraciones de este metabolito no sintetizan su propia Cr sino que la obtienen desde el torrente sanguíneo por un específico transportador denominado CreaT, hoy en día conocido como SLC6A8 ya que mediante clonación molecular ha sido caracterizado por ser parte de una amplia familia de transportadores de neurotransmisores dependientes de Na+ y Cl-, conocida como la familia 6 de transportadores de solutos (SLC6) (Snow & Murphy 2001; Peral et al. 2002; Christie 2007). Cerca del 95% de las reservas de Cr en el cuerpo se hallan al interior del músculo esquelético, el otro 5% restante se encuentra en el corazón, cerebro, hígado, riñón, testículos y células fotorreceptoras de la retina (Snow & Murphy 2001; Speer et al. 2004; Wallimann & Hemmer 1994; Wallimman et al. 2011). La concentración promedio de Cr en el tejido muscular es de 120 mmol Kg-1 de masa muscular (Stout et al. 2008). En el músculo, la Cr se encuentra en un 33% en forma aislada y el 67% restante en forma de fosfocreatina (PCr), la forma cargada energéticamente vía creatina cinasa (CK - EC 2.7.3.2) (Figura 2). En las fibras musculares tipo I la concentración de PCr es ligeramente más baja (entre un 15 y 20%) en comparación con las fibras tipo II (Chicharro & Fernández 2006). Las concentraciones citosólicas de Cr y PCr en el miocito son de 13 - 27 mM, respectivamente (Brosnan et al. 2007). La PCr y Cr en relación al ATP y ADP son moléculas más pequeñas y menos cargadas negativamente que pueden encontrarse en concentraciones mucho mayores en muchas de las células y tejidos que contienen CK, en relación con eso permiten un mayor flujo intracelular de fosfatos de alta energía. Además, el delta de energía libre de Gibbs (∆G°’) a pH 7.0 para la hidrólisis de PCr es de 245.0 kJ mol-1 comparado con los 231.8 kJ mol-1 para el ATP, implicando que en tejidos con un sistema activo de CK, la fosforilación potencial pueda ser regulada a un nivel mayor que en tejidos desprovistos del sistema CK (Brosnan & Brosnan 2007). Este factor es esencial para el funcionamiento óptimo de algunas ATPasas celulares, por ejemplo las ATPasas miofibrilares (Wallimann 2009). Durante el ejercicio intenso, la Cr en su forma energéticamente cargada, la PCr, está disponible en las fibras musculares para la regeneración inmediata de ATP mediante una reacción catalizada por la CK en un proceso rápido y anaeróbico (Figura 2) (Chicharro & Fernández 2006).
El metabolismo de los fosfágenos o fosfatos de alta energía, como la PCr, proporciona la energía necesaria para la contracción muscular al inicio de la actividad y durante ejercicios explosivos, muy breves y de elevada intensidad. En los periodos de inactividad muscular la PCr es rápidamente resintetizada, paradójicamente la síntesis de PCr a partir de Pi y Cr precisa la energía procedente de la hidrólisis del ATP (Bessman & Carpenterm 1985). Como se está en fase de recuperación el ATP se está sintetizando por los sistemas aeróbicos y parte de este ATP irá destinado a ceder su energía para reponer las concentraciones basales de PCr (Chicharro & Fernández 2006).
La Cr cristaliza con agua en un prisma monoclínico reteniendo una
molécula de agua de cristalización por molécula de Cr (Pischel & Gastner 2007),
formando el popular suplemento dietario Monohidrato de Cr, el cual hace parte de los
suplementos nutricionales frecuentemente consumidos por atletas de élite y
personas del común debido a su ratificados efectos ergogénicos (Hespel 1999;
Barbero 1999; Casey
& Greenhaff 2000; González et al. 2003;
Maughan et al. 2004; Kreider et al. 2003b, 2010;
Wallimann 2008; Stec & Rawson 2010; Kreider & Jung 2011;
Del Favero et al. 2011; Kim et al. 2011; Carrillo & Gilli 2011).
Fragmento tomado del artículo: Bonilla DA. (2013). Principios Metabólicos de los Efectos de la Suplementación con Creatina sobre el Rendimiento Deportivo. PubliCE Standard
Diego A. Bonilla