
Regulación Fisiológica del Rendimiento en el Maratón
University of Texas at Austin. Austin, Texas, Estados Unidos.
Artículo publicado en el journal PubliCE, Volumen 0 del año 2013.
Publicado 25 de octubre de 2013
Resumen
INTRODUCCION
La velocidad de carrera de un maratón está regulada por el metabolismo aeróbico en las fibras musculares reclutadas y por la conversión económica de esta energía en velocidad. Los aspectos de este concepto han sido reconocidos durante mucho tiempo. Hace aproximadamente 38 años Costill [1] escribió una monografía titulada "Lo que las investigaciones dicen a los entrenadores sobre las carreras de fondo" en la cual, el autor resume la literatura fisiológica en un paradigma intuitivo centrado en el máximo consumo de oxígeno y su utilización fraccionaria así como también en la economía de la carrera. La validez de estos conceptos para predecir el rendimiento de la maratón fue validada por Farrell et al. [2] en 1979 y la utilidad de la medición del lactato sanguíneo para identificar el ritmo de carrera en una maratón competitiva se fue consolidando posteriormente. El fenómeno de “golpear el muro” que se observa en una maratón debido a la oxidación inadecuada de carbohidratos fue ampliamente vinculado con el agotamiento del glucógeno muscular y la hipoglucemia. [3,4] Por consiguiente, en la actualidad el contexto dentro del cual se encuadran los reguladores fisiológicos de rendimiento de la maratón, es generalmente similar al que se discutió hace 30 años durante la reunión patrocinada por la Academia de Ciencias de Nueva York [5].
Los conceptos claves de esta regeneración oxidativa de trifosfato de adenosina (ATP) se presentan en la Figura 1. Los corredores de maratón corren a velocidad y tasa de consumo de oxígeno (es decir VO2 de maratón) superiores a las que pueden tolerar sus músculos en ejercicio sin experimentar fatiga, la cual progresa y los hace volverse más lentos durante los primeros 20 km. El VO2 en una maratón competitiva con ritmo adecuado es la tasa en estado estable más alta posible de regeneración oxidativa de ATP de todo el cuerpo que generalmente puede ser mantenida durante 42 km. Este VO2 de maratón es una función de, no sólo maximizar la tasa de producción de ATP en estado estable dentro de una fibra muscular dada, si no que también es una función de reclutar el mayor número de fibras musculares que puedan participar en la producción económica de velocidad. La tasa de producción de energía también variará con las condiciones medioambientales.
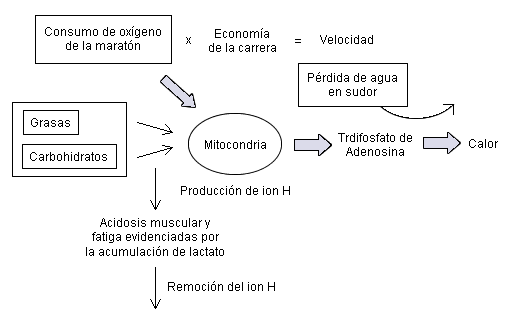
Figura 1. Concepto de los factores fisiológicos que regulan el rendimiento en maratón.
Una causa de fatiga muscular podría ser la acidosis progresiva y la alteración iónica. La velocidad de la maratón se aproxima a la intensidad en la cual, el lactato comienza a acumularse en la sangre (es decir el umbral del lactato sanguíneo) y en las fibras musculares. [1, 2] La medición del umbral del lactato sanguíneo para estimar el ritmo de una maratón competitiva se popularizó porque es práctica y teóricamente válida. El concepto no es que la molécula de lactato por si misma provoca fatiga, si no que su acumulación en la sangre refleja una alteración en la homeostasis de la célula muscular.
Las maratones se corren a intensidades bastante por debajo del consumo de oxígeno máximo (por ejemplo 65-85% VO2max), y dado que la fatiga se asocia con una glucógenolisis acelerada y no con eventos anaeróbicos en el músculo, podríamos plantear la hipótesis que el aumento en la entrega de oxígeno a los músculos que realizan ejercicio (por ejemplo flujo de sangre y contenido de oxígeno en la sangre) aumentaría menos la velocidad de la maratón, en comparación con lo que ocurre en las carreras realizadas a velocidades que producen el consumo de oxígeno máximo (por ejemplo 1500-5000m). Sin embargo, es posible que un aumento en la entrega de oxígeno que incremente la presión de oxígeno muscular pueda producir un mejor estado redox (por ejemplo relación difosfato de adenosina /ATP) que haga mas lenta la glucógenolisis para una cierta velocidad de regeneración oxidativo de ATP o una velocidad de carrera dada.
Los corredores de maratón de primer nivel llegan a la final en aprox.2:30:00 o más rápidamente, mientras que la mayoría de los finalistas de maratón son más lentos y los corredores de carreras de caridad son considerablemente más lentos. Debido a la relación inversa entre duración del ejercicio e intensidad, junto con el hecho que los mejores atletas de resistencia pueden realizar ejercicio en VO2 más altos de manera sostenida antes de experimentar fatiga, existe una amplio intervalo de valores de porcentaje de VO2max durante la competencia de maratón entre los diferentes individuos. En un extremo, un corredor lento puede promediar 50-60% de su VO2max mientras que un corredor de máximo nivel puede alcanzar un promedio >80% VO2max. Si la economía de carrera no es diferente al comparar los corredores lentos y los corredores rápidos, por definición, el gasto calórico total debería ser el mismo, aunque la tasa de intercambio respiratorio y cantidad total de carbohidratos oxidados debería ser más alta en los corredores más rápidos. El autor no cuenta con datos que consideren la posibilidad que los corredores de maratón lentos y rápidos presenten diferencias en cuanto al nivel en el cual el agotamiento de los carbohidratos provoca fatiga.
Los corredores de maratón de máximo nivel obtienen más de dos tercios de su energía de carbohidratos provenientes del glucógeno muscular y en menor grado de la oxidación de glucosa sanguínea [1]. El ejercicio a 70-85% de VO2max no puede ser mantenido sin una oxidación de carbohidratos suficiente y así la severa disminución de glucógeno muscular, a menudo junto con hipoglucemia, provocan la necesidad de reducir la intensidad a aprox. 40-60% VO2max. Este fenómeno ha sido llamado “golpear el muro” y la velocidad subsiguiente sería la que es posible mantener principalmente por la oxidación de grasas, glucosa sanguínea y lactato. El lactato se generaría a partir del glucógeno en las fibras musculares inactivas [6].
Loa atletas de resistencia altamente entrenados poseen más mitocondrias musculares y así tienen una mayor capacidad de oxidar tanto glucógeno como triacilglicéridos (es decir triacilglicéridos intramusculares o específicamente triacilglicéridos intramiocelulares [IMTG]). En comparación con los individuos desentrenados, los individuos entrenados típicamente se comparan mientras corren a un cierto VO2 absoluto (mL/kg/min) durante el cual los sujetos entrenados presentan una elevada oxidación de grasas derivada de IMTG. Esto se asocia con, y puede ser una consecuencia de, una disminución en la oxidación de glucógeno muscular [7]. Así, el entrenamiento de resistencia aumenta la capacidad de oxidar grasas, y esto es más evidente en las intensidades inferiores a las de una maratón de competición [7]. El mayor VO2 que los corredores de maratón de máximo nivel pueden mantener durante una carrera se alimenta por una mayor oxidación de glucógeno muscular y en menor grado IMTG, ambos sustratos localizados dentro del músculo. [8]
Cuando se corren 42,2 km en comparación con 10km, es importante destacar que a pesar de correr más de cuatro veces, la velocidad típica de carrera de la maratón solo se reduce aprox. 10% en comparación con una carrera de 10 km. Dicho de otra manera, si los corredores de maratón fijaran un ritmo ligeramente más rápido (por ejemplo 5-10%) que el ideal (e.g. la velocidad del umbral de lactato está cerca de la ideal) durante una distancia de 42,2km, se fatigarían prematuramente (es decir después de 5-10km), probablemente debido a una glucógenolisis acelerada, tal como se ha observado en ciclistas [9]. Esta fatiga podría manifestarse por acidosis y un agotamiento eventual del glucógeno en las unidades motoras que se reclutan con mayor facilidad en los músculos que participan en la carrera. Incluso cuando el ritmo es ideal y constante durante la maratón, aumenta la sensación de esfuerzo necesaria para el reclutamiento de suficientes unidades motoras, sobre todo después de aprox. 25-35 km de carrera. En ese punto de la carrera, el glucógeno muscular es bajo en muchas fibras musculares, particularmente en las fibras de tipo I de fácil reclutamiento. Si la oxidación, principalmente de carbohidratos, no se mantiene en una tasa lo suficientemente alta en suficientes fibras musculares, es necesario aminorar el paso. La ingesta de carbohidratos durante los ejercicios retrasa el tiempo de fatiga porque la glucosa exógena llega a la sangre y ayuda a mantener la tasa de oxidación de carbohidratos [4,10]. El mantenimiento de la concentración de glucosa sanguínea por la ingesta de carbohidratos y la consiguiente prevención de hipoglucemia, evita la neuroglucopenia (es decir el dañó en la función neuronal por falta de glucosa) y los síntomas del sistema nerviosos central de fatiga que a veces se manifiestan con una gran respuesta de catecolaminas y palidez de piel subsiguiente asociada con irritabilidad, confusión y letargo.
Las velocidades de carrera más rápidas requieren aumentos en la frecuencia de zancada y en la longitud de la misma y por lo tanto van acompañadas de una activación más frecuente de un número mayor de unidades motoras. Esto implica que cuando se corre en la velocidad de maratón, que por supuesto está muy por debajo de la velocidad de esprint, no se reclutan de manera simultánea todas las unidades motoras. Es difícil obtener la estimación de la cantidad total (por ejemplo kg) de músculo que se recluta al correr así como el ciclo de trabajo de reclutamiento de unidades motoras (por ejemplo cada zancada vesus zancada de por medio versus cada tres zancadas, en promedio). Sin embargo, en los ciclistas de resistencia altamente entrenados, se ha observado que aquéllos que tienen un umbral de lactato sanguíneo más alto pueden mantener una producción de potencia dada por un tiempo dos veces más largo (por ejemplo 30 minutos contra 60 minutos) y que esto sería el resultado de su capacidad para reclutar 22% más de masa muscular para convertir en producción de potencia [9, 10]. El concepto es que la capacidad de reclutar una mayor cantidad de masa muscular para compartir en el trabajo, durante un período de 10 minutos, reduce la tasa de trabajo relativa en las fibras musculares reclutadas durante cada pedaleo porque más fibras intervienen en el trabajo en una cierta pedaleada y así se reduce el ciclo de trabajo promedio de las fibras musculares reclutadas. Como resultado, las fibras individuales experimentan menos fatiga debido a una menor tasa de regeneración oxidativa de ATP dentro de sus mitocondrias. Como resultado, disminuye la perturbación en la homeostasis cuando más masa muscular puede participar en el trabajo. Esto reduce la glucógenolisis muscular tal como se observa en la producción de lactato y le permite al atleta de resistencia realizar ejercicio a una intensidad alta durante períodos de tiempo significativamente mayores. Esencialmente, el concepto es que, con el desarrollo de la capacidad de distribuir el trabajo en una masa muscular mayor, mas mitocondrias pueden compartir el trabajo de metabolismo oxidativo reforzando el número potencial de unidades motoras que podrían reclutarse en un ciclo de trabajo menor cuando se corre a una velocidad dada [9, 11]. De hecho, el reclutamiento de unidades motoras puede regular el estrés mitocondrial.
La hipertermia puede limitar el rendimiento en maratón porque estresa los sistemas cardiovascular, nervioso central y muscular. [10, 12, 13] El nivel de hipertermia corporal experimentado durante una maratón refleja el equilibrio entre la producción de calor y la dispersión del mismo. El calor se produce por la hidrólisis de ATP y los procesos metabólicos necesarios para la regeneración oxidativa de ATP (Figura 1). Cuándo se corre por un terreno llano, se realiza poco trabajo físico y la gran mayoría de la energía metabólica, calculada mediante calorimetría indirecta (es decir con el VO2 y la tasa de intercambio respiratorio), se transfiere al calor y se libera dentro del cuerpo [14]. La implicancia importante de esto es que los individuos que tienen una economía de carrera superior, o sea un menor VO2 para una determinada velocidad de carrera, también generaran proporcionalmente menos calor. Ésta podría ser una ventaja distinta al competir en ambientes calurosos que limitan la cantidad de dispersión de calor, como ha sido típicamente el caso durante las competencias de maratón olímpicas.
El mecanismo principal para la dispersión de calor durante una maratón, sobre todo en ambientes calurosos, es el enfriamiento a través de la evaporación de sudor [13]. La pérdida de sudor que no se compensa con consumo de fluidos producirá deshidratación. El mayor problema con la deshidratación es que reduce la dispersión de calor debido al menor flujo de sangre superficial y a la disminución en la tasa de sudoración [15]. La cantidad de deshidratación que probablemente puede ser tolerada sin desarrollar una hipertermia perjudicial depende del ambiente y de la tasa de producción de calor del individuo [10, 13]. Cuando el ambiente está frío (por ejemplo 5-10°C) o templado y seco (por ejemplo 21-22°C), se ha planteado la hipótesis que una pérdida de agua de aprox. 2% del peso corporal podría ser tolerada sin riesgo para el bienestar y el rendimiento [10]. Sin embargo, cuando la maratón se corre en un ambiente caluroso y/o húmedo, se supone que la deshidratación de 2% del peso corporal aumenta la probabilidad de afectar el rendimiento y de sufrir hipertermia y golpe de calor.
Un determinante importante de rendimiento de maratón es la velocidad de carrera que se puede mantener en un cierto VO2 generado (i.e mL/kg/min) [1, 2]. Esta relación se denomina “economía de la carrera”. Una población de corredores presenta un intervalo de 25-30% en la economía de carrera [2]. Por ejemplo, si un corredor con una economía de carrera promedio fuera capaz de mantener un VO2 de 50 mL/kg/min durante la maratón, esta tasa de gasto de energía y velocidad de carrera deberían arrojar un tiempo de maratón de 2:40:00. Sin embargo, se estima que los corredores más económicos finalizarán en un tiempo <2:20:00 y los menos económicos en aprox. 3:00:00. No están claro cuales son los factores que determinan la economía de carrera y hasta que punto la misma puede ser mejorada con el entrenamiento. Notablemente, Jones [16] informó que la economía de carrera mejoraba sustancialmente en corredoras de maratón de elite durante un período de 5 años. Sin embargo, dado que se han realizado pocos estudios a largo plazo, no se sabe si esta mejora es típica o única.
Correr una maratón a la velocidad más rápida posible estaría regulado por la tasa de metabolismo aeróbico (es decir VO2 de la maratón) de una cantidad limitada de energía proveniente de carbohidratos (es decir glucógeno muscular y glucosa sanguínea) y por la velocidad que puede mantenerse sin que se desarrolle hipertermia.
Records Mundiales Actuales
El record mundial actual para varones es 2:04:55, y fue establecido por Paul Tergat en 2003 en Berlín. En las mujeres, Paula Radcliff alcanzó el record impresionante de 2:15:25 en el año 2003 en Londres. Este asciende a tiempos de carrera por kilómetro de 2 minutos 58 segundos para varones y 3 minutos 23 segundos para mujeres. Expresado como tiempo promedio de carrera por milla, esto se traduce a 4 minutos 46 segundos para varones y 5 minutos 9 segundos para mujeres. De hecho, el record mundial de Paula Radcliff es una marca excepcional dado que bajó el record de maratón 3-4 minutos en el transcurso de 1 año. Además, su ritmo de maratón sólo es ligeramente más bajo que el record mundial de ritmo de media maratón que se ubicó en 1:06:44 (Elana Meyer, 1999).
Afortunadamente, el Dr Andrew Jones ha realizado evaluaciones detalladas y periódicas en el laboratorio a Paula Radcliff desde los 17 años (en 1991) hasta el momento en que estableció el record mundial a los 29 años de edad (en 2003) [17]. Durante el período de 1991-5, se informó que la velocidad de carrera que permitía alcanzar su umbral de lactato aumentó dramáticamente de 6 minutos 25 segundos a 5 minutos 20 segundos por milla [16]. Jones [16] declaró que "Esto representa una mejora en la velocidad de carrera de 20% en el umbral de lactato en 4 años y es indicativo de una capacidad de resistencia mucho mayor”. Notablemente, el VO2max no aumentó durante este período, aunque la intensidad del ejercicio que permitió alcanzar el umbral de lactato sanguíneo aumentó de 80 a 88% VO2max. Más notablemente, la economía de carrera aumentó sustancialmente, tal como lo reflejó una reducción de 53 a 48 mL/kg/min en el costo del oxígeno por correr a 16,0 km/h (o 6 minutos por milla). Esto asciende a una mejora de aprox. 9% en la economía de la carrera durante este período de 4 años. Parece que la mejora continua en la economía de carrera durante el período de 1995-2003 jugó un papel significativo en su desarrollo fisiológico continuo y en el establecimiento del record mundial actual [17]. También se observaron mejoras en la economía de carrera durante un período largo de entrenamiento en un corredor varón, de medio fondo que alcanzó la categoría de élite [18]. Por consiguiente, los datos sobre hasta que punto la economía de la carrera puede mejorar con el entrenamiento continuado se limitan a algunos informes de casos que han efectuado el seguimiento de atletas durante varios años.
El rendimiento en ciclismo de resistencia también está influenciado por la eficiencia mecánica mientras se pedalea y, tal como ocurre con la economía de la carrera, existe controversia sobre hasta que punto la eficiencia mecánica puede ser mejorada con el entrenamiento. Recientemente se informó en un atleta que se transformó en un ganador múltiple del Tour de Francia, que la eficiencia mecánica durante ciclismo mejoró 8% durante un período de 7 años de entrenamiento [19]. Por consiguiente, parecería que si la economía de carrera o eficiencia de pedaleo efectivamente responden al entrenamiento, tal como se sugiere en estos informes de casos de atletas individuales, las mejoras se producen solamente aprox. 1-2% por año. Aunque este índice de mejora es significativo y se puede medir con confiabilidad anualmente en lo que respecta al rendimiento deportivo en carreras, podría ser demasiado pequeño para ser detectado en evaluaciones en el laboratorio porque se encuentra dentro del intervalo de error de la medición.
Por consiguiente, parecería que las mediciones de eficiencia o economía deberían ser realizadas durante varios años y estos estudios deberían ser realizados en grupos de atletas para determinar si estos informes de caso pueden ser generalizados a personas con capacidad de entrenar de una manera extremadamente intensa durante años.
¿Cuán Rápido Pueden Correr los Maratonistas de Élite?
Es interesante y revelador especular cuán rápido pueden correr los corredores de maratón de élite. Una metodología utilizada ha sido simplemente graficar los tiempos de records mundiales o velocidad durante los años y estimar matemáticamente la velocidad en la cual ésta relación curvilínea alcanzará una asíntota, supuestamente un reflejo del potencial humano final. Sin embargo, esta metodología se basa solamente en la historia del pasado y por lo tanto está muy influenciada por fenómenos sociológicos que determinaron si los individuos con el mayor potencial biológico innato tuvieron la oportunidad de cultivar su talento para correr maratones. Por lo tanto, este registro histórico representa la progresión de factores sociológicos y factores fisiológicos. Este sería el caso entre las mujeres, dado que los records mundiales mejoraron rápidamente durante el período de 1970-90 a medida que aumentaron las oportunidades de las mujeres para correr. Por consiguiente, en la opinión de este autor sería prematuro, predecir cual podría ser el límite final para el rendimiento en maratón de varones y mujeres utilizando registros históricos porque es solamente durante los últimos 30-50 años que algunos corredores de distancia han contado con el apoyo adecuado y los medios para destinar su energía completamente al entrenamiento para una maratón. Parecería que a medida que mayor cantidad de corredores de medio fondo de élite (por ejemplo 5km y 10km) “se pasan” al maratón, que el conjunto de talento en bruto continuará como mínimo, mejorando y alimentará la progresión de los records mundiales de las próximas décadas.
Otra metodología para estimar los records mundiales futuros y el rendimiento final en maratón es desarrollar un modelo biológico que tome todos los mejores atributos fisiológicos observados en los individuos por separado y los combine, teóricamente, en uno individual. Esta metodología fue adoptada por Joyner [20] quién presentó un modelo para el rendimiento óptimo en maratón basado en las medidas documentadas de VO2max, umbral de lactato y economía de carrera entre los corredores de élite. El tiempo de maratón más rápido en varones estimado por el modelo de Joyner [20] fue 1:57:58. Este modelo es para un varón hipotético con un VO2max de 84 mL/kg/min, un umbral de lactato de 85% de VO2max y con una economía de carrera excepcional. En la opinión del autor, esta estimación es conservadora porque existe un potencial para tiempos aun más rápidos, especialmente a partir de una economía de carrera mejorada.
El record mundial masculino actual para media maratón (21,1km) es 58 minutos 55 segundos, obtenido por Haile Gebrselassie en 2006. Los factores exactos que le impiden a un corredor de maratón mantener su mejor ritmo de media maratón, durante una maratón completa, no están completamente claros. Los corredores, por supuesto, deben evitar el agotamiento de carbohidratos o “golpear el muro”. Para ser económica, la carrera a una velocidad de 5 minutos por milla o mayor requiere una forma de carrera precisa y coordinada. Los corredores tienen opciones limitadas para cambiar la longitud de la zancada, frecuencia de la zancada o posición del cuerpo para compensar o atenuar la fatiga muscular local. Es probable que la fatiga muscular local, incluso en una cantidad pequeña de músculo, sea suficiente para alterar la forma de correr y elevar el costo de oxígeno de la carrera y acelerar el agotamiento de glucógeno, precipitando así el fenómeno conocido como “golpear el muro”.
En la opinión de este autor, la mejor manera de alcanzar mejoras significativas en el record mundial de rendimiento de maratón será mejorando la resistencia a la fatiga dentro de los grupos musculares y en la actividad de las articulaciones que experimentan agotamiento de glucógeno o fatiga neurológica. Esta fatiga podría manifestarse en un determinado individuo por cinemáticas alteradas (i.e alteración de la forma) después de 60-90 minutos de correr al paso de maratón fijado como meta. Podrían obtenerse mejoras a través de entrenamiento centrado muy específicamente en el mantenimiento de una forma de correr económica. Además, el entrenamiento que ponga en juego más unidades motoras de la musculatura que participa en la carrera para compartir el trabajo, tal como discutimos previamente, debería reducir el trabajo por unidad motora y por lo tanto debería reducir la capacidad de fatiga o la necesidad de reducir significativamente el ritmo en la maratón, hasta un valor por debajo del ritmo de media maratón. Obviamente, el CNS y las vías neuronales periféricas aferentes y eferentes son fundamentales para las adaptaciones inducidas por el entrenamiento y finalmente para el rendimiento.
Por lo tanto, según Joyner, [20] las personas poseen la capacidad fisiológica para correr una maratón en 1:58:00. El autor no se sorprendería si esta predicción se cumple en la próxima década. Esto se logrará si el ritmo del record mundial actual para la media maratón se mantiene en una maratón completa. Paula Radcliff prácticamente lo ha logrado al establecer el record mundial femenino actual. El límite final para el rendimiento de maratón podría ser establecido por los límites de la economía de la carrera y la resistencia a la fatiga del músculo local en individuos que poseen la capacidad de correr 26,2 millas con gastos de energía superiores a 70 mL/kg/min sin sufrir hipertermia debilitante.
Agradecimientos
Edward F. Coyle es consultor del Gatorade Sport Science Institute e investigador contratado de Quaquer Oats-Gatorade y POMS Wonderful.
Dirección de Contacto
Professor Edward F. Coyle, Department of Kinesiology and Health Education, University of Texas at Austin, Bellmont 820, Austin, TX 78712, USA. E-mail: coyle@mail.utexas.eduReferencias
1. Costill DL (1968). What research tells the coach about distance running. Washington, DC: American Association for Health, Physical Education and Recreation
2. Farrell PA, Wilmore JH, Coyle EF, et al (1979). Plasma lactate accumulation and distance running performance. Med Sci Sports 11: 338-44
3. Bergstrom J, Hermansen L, Hultman E, et al (1967). Diet, muscle glycogen and physical performance. Acta Physiol Scand; 71: 140-50
4. Levine SA, Gordon B, Derick CL (1924). Some changes in the chemical constituents of the blood following a marathon race. J Am Med Assoc; 82 (22): 1778-9
5. Milvy P (1977). The marathon: physiological, medical, epidemiological, and psychological studies. Ann N Y Acad Sci; 301: 1-1090
6. Ahlborg G, Felig P (1982). Lactate and glucose exchange across the forearm, legs and splanchnic bed during and after prolonged leg exercise. J Clin Invest; 69: 45-54
7. Phillips SM, Green HJ, Tarnopolsky MA, et al (1996). Effects of training duration on substrate turnover and oxidation during exercise. J Appl Physiol; 81 (5): 2182-91
8. Romijn JA, Coyle EF, Sidossis LS, et al (2000). Substrate metabolism during different exercise intensities in endurance-trained women. J Appl Physiol; 88: 1707-14
9. Coyle EF, Coggan AR, Hopper MK, et al (1988). Determinants of endurance in well-trained cyclists. J Appl Physiol; 64: 2622-30
10. Coyle E (2005). Fluid and fuel intake during exercise. J Sport Sci; 22: 39-55
11. Coyle EF (1995). Integration of the physiological factors determining endurance performance ability. Exerc Sport Sci Rev; 23: 25-63
12. Gonzalez-Alonso J, Teller C, Andersen SL, et al (1999). Influence of body temperature on the development of fatigue during prolonged exercise in the heat. J Appl Physiol; 86: 1032-9
13. Sawka MN, Young A (2006). Physiological systems and their responses to conditions of heat and cold. In: Tipton CM, editor. American College of Sports Medicine’s Advanced exercise physiology. Philadelphia (PA): Lippincott Williams and Wil-kins: 535-63
14. Webb P, Annis J, Troutman SJ (1972). Human calorimetry with a water-cooled garment. J Appl Physiol; 32: 412-9
15. Coyle EF, Gonzalez-Alonso J (2001). Cardiovascular drift during prolonged exercise: new perspectives. Exerc Sport Sci Rev; 29: 88-92
16. Jones AM (1998). A five year physiological case study of an Olympic runner. Br J Sports Med; 32: 39-43
17. Jones AM (2006). The physiology of the world record holder for the women’s marathon. Intern J Sports Sci Coaching; 1 (2): 101-16
18. Conley DL, Krahenbuhl GS, Burkett LN, et al (1984). Following Steve Scott: physiological changes accompanying training. Phys Sportsmed; 12: 103-6
19. Coyle EF (2005). Improved muscular efficiency displayed as ‘Tour de France’ champion matures. J Appl Physiol; 98: 2191-6
20. Joyner MJ (1991). Modeling optimal marathon performance on the basis of physiological factors. J Appl Physiol; 71: 683-7