
Rendimiento Muscular y Adaptaciones Enzimáticas al Entrenamiento Intervalado de Sprint
Duncan Mac Dougall1, J. R Macdonald1, H. J Green1, Audrey L Hicks1, Robert S McKelvie1 y Kelly M Smith1
Departamento de Kinesiología. McMaster University. Hamilton, Ontario Canadá.
Artículo publicado en el journal PubliCE, Volumen 0 del año 1998.
Publicado 28 de enero de 2004
Resumen
Palabras clave: protocolo de wingate, biopsia muscular, enzimas glucolíticas, enzimas oxidativas
INTRODUCCION
Es bien sabido que un programa de entrenamiento de resistencia puede resultar en incrementos significativos en la densidad mitocondrial (14, 15) y la actividad enzimática oxidativa (13, 24), pero que tiene efectos mínimos sobre las enzimas glucolíticas (12). En estudios con animales y humanos, los cambios en la actividad enzimática oxidativa son frecuentemente de 3 a 5 veces mayores que los incrementos observados en el máximo consumo de oxígeno (VO2 máx.) (5, 6, 8) y presentan una estrecha correlación con los incrementos observados en la capacidad de resistencia (6). El hecho de si un programa de entrenamiento anaeróbico o de sprint puede resultar en un incremento en la actividad máxima de, ya sea, las enzimas glucolíticas u oxidativas, es en cierta forma más controversial. Mientras que la mayoría de los investigadores ha señalado incrementos en la actividad enzimática glucolítica después del entrenamiento de sprint (2, 4, 16, 25), algunos no lo han señalado (9, 11). Además, hay reportes en la literatura acerca de que el entrenamiento de sprint tiene, ya sea, ningún efecto (4, 17) o un efecto menor (27) sobre la actividad enzimática mitocondrial, de lo que tiene el entrenamiento de resistencia. Es posible que muchas de estas disparidades (ver Tabla 1) puede ser causadas por las diferentes intensidades y duraciones del ejercicio llamado “entrenamiento de sprint”, así como los problemas en la simulación de esfuerzos máximos de sprint con ciertos modelos animales.
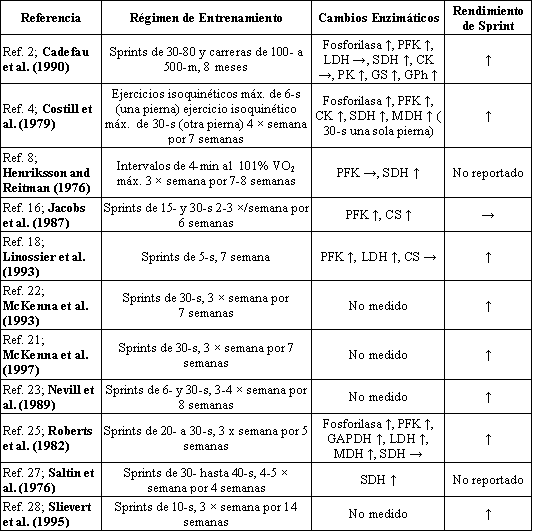
Tabla 1. Efectos del entrenamiento de sprint sobre la actividad enzimática
muscular y el rendimiento de sprint en hombres. VO2 máx., máximo
consumo de oxígeno; PFK, fosfofructo quinasa; LDH, lactato dehidrogenasa; SDH,
succinato dehidrogenasa; CK, caseín quinasa; PK, proteín quinasa; GS, glucógeno
sintetasa; GPh, glucógeno fosforilasa; MDH, malato dehidrogenasa; CS, citrato
sintetasa; GADPH, gliceraldehído-3-fosfato dehidrogenasa. ↑, Incremento; →,
ningún cambio.
Las sugerencias acerca de que el entrenamiento de sprint no induce incrementos en la actividad enzimática muscular son en cierto modo poco claras, ya que en la mayoría de los estudios en los cuales fue incluida una medición de rendimiento, se ha demostrado que tal entrenamiento resulta en una mejora en la producción de potencia de corta duración (4, 5, 23, 28). De este modo, nosotros decidimos reexaminar este aspecto investigando la actividad enzimática glucolítica y oxidativa en un grupo de adultos jóvenes físicamente activos y sanos, antes y después de que ellos llevaran a cabo un programa de entrenamiento intervalado e intenso de sprint, similar a los entrenamientos en los que previamente se encontró que resultaban en una mejora de la producción de potencia de corta duración (21, 22).
METODOS
Sujetos
12 hombres jóvenes sanos graduados y estudiantes de pregrado en Educación Física (edad 22.7±2 años, talla 175±6 cm, masa corporal 73.4±6.2 kg) se ofrecieron para participar en la investigación. Todos eran entusiastas de la aptitud física, físicamente activos que practicaban pedestrismo, entrenamiento de la fuerza y otros deportes, pero ninguno era atleta universitario al momento del estudio. A los sujetos se les informo los propósitos del estudio y los riesgos asociados y ellos dieron un informe de consentimiento por escrito. A los sujetos se les proporcionó una remuneración por participar y completar el estudio, y la adherencia al entrenamiento fue de 100%. El proyecto fue aprobado por el Comité de Ética Humana de la Universidad de McMaster.
Programa de Entrenamiento
Los sujetos entrenaron 3 veces por semana en días alternados por un período total de 7 semanas. El entrenamiento consistió de intervalos de esfuerzos máximos de 30 s en una bicicleta ergométrica Monach frenada mecánicamente, en la cual fue usado el protocolo de Wingate (1). El programa comenzó con 4 intervalos, con 4 min de recuperación por sesión en la semana 1, y con un incremento de los intervalos de dos cada semana hasta la semana 4, luego fueron realizados 10 intervalos por sesión. Los intervalos de recuperación fueron de una duración de 4 min durante las semanas 1-4 y fueron subsecuentemente disminuidos en 30 s cada semana durante las 3 semanas restantes.
Durante los períodos de recuperación entre intervalos, los sujetos fueron alentados a mantener algún grado de rotación de los pedales contra, ya sea, ninguna carga o una carga de 0.5 kg. Esto fue necesario en las primeras etapas del programa de entrenamiento para prevenir mareos y nauseas, después de los intervalos de ejercicio. La velocidad de rotación del pedal no fue registrada durante la recuperación, pero en ninguna instancia la producción de potencia excedió los 25 W. Todas las sesiones de entrenamiento fueron supervisadas por un asistente de investigación, el cual ajustaba la carga, tomaba los tiempos de los intervalos de recuperación, y proporcionaba aliento verbal durante las series de ejercicio. Los sujetos fueron instruidos para no hacer ningún entrenamiento adicional a través de la duración del estudio, y la adherencia a esto fue subsecuentemente confirmada mediante un cuestionario post-estudio.
Mediciones
Todas las mediciones fueron hechas antes y después del programa de entrenamiento de 7 semanas.
Potencia Anaeróbica
La potencia y la capacidad aeróbica máxima fueron evaluadas a través de 4 esfuerzos repetidos de 30 s (Wingate test) sobre una bicicleta ergométrica con 4 min de recuperación entre cada esfuerzo. La carga fue de 0.075 kg/kg de masa corporal y el cicloergómetro fue precargado con peso para la aplicación inmediata del mismo al comienzo de la evaluación. Los pies de los sujetos fueron firmemente sujetados a los pedales, y la altura del asiento fue ajustada para una comodidad y eficiencia de pedaleo, óptimas. Los sujetos trataban de alcanzar una velocidad de pedaleo máxima contra la inercia impuesta solo por el ergómetro a través de 2 s, luego de lo cual era aplicada la carga completa y el contador electrónico de revoluciones era activado. La producción de potencia para cada segundo, la producción de potencia pico, la producción de potencia durante 30 s promedio (trabajo total) y el porcentaje de fatiga fueron calculados y desplegados para cada evaluación (SMI Opto-Sensor and Software System, St. Cloud, MN). La evaluación post-entrenamiento fue administrada 2 días después de la última sesión de entrenamiento.
Las concentraciones de lactato y potasio de la arteria braquial y la vena femoral fueron también monitoreadas continuamente por medio de catéteres, a través de este período.
Potencia Aeróbica
La potencia aeróbica máxima (VO2 máx.) fue medida en una bicicleta ergométrica frenada eléctricamente (Jaeger), usando un protocolo continuo estándar de carga progresiva. Fue usado un sistema de análisis de gases de circuito abierto para calcular el consumo de oxígeno cada 20 s, durante la evaluación hasta el agotamiento. El consumo de oxígeno más alto (promediado cada 1 min), alcanzado durante la evaluación, fue seleccionado como el VO2 máx. del sujeto.
Actividad Enzimática Muscular
En 9 de los 12 sujetos, fueron tomadas biopsias percutaneas a partir del vasto lateral, bajo anestesia local, por medio de succión manual. Fueron obtenidos aproximadamente 100-200 mg de tejido húmedo por muestra. Una parte de cada muestra fue usada para la determinación del sitio de binding de la oubaina [3H], como un indicador de la concentración de Na+-K+-ATPasa y la parte restante fue utilizada para los análisis de enzimas musculares.
Para los análisis enzimáticos, el tejido fue congelado-secado, disecado libre de sangre y tejido conectivo y homogeneizado en glicerol 50%, 20 mM de buffer de fosfato de sodio (pH=7.4), β-mercaptofenol 5 mM, EDTA 0.5 mM, y albumina sérica bovina 2%. La actividad de la glucógeno fosforilasa total, hexoquinasa (Hex), fosfofructo quinasa (PFK), lactato dehidrogenasa (LDH), citrato sintetasa (CS), succinato dehidrogenasa (SDH), malato dehidrogenasa (MDH), y 3-hidroxiacil-CoA dehidrogenasa fueron determinadas fluorometricamente, de acuerdo a los procedimientos descritos por Henriksson y colegas (7). Todos los ensayos fueron realizados en duplicado, y para un sujeto dado, fueron completados en el mismo día de análisis. Los datos fueron expresados en moles por kilogramo de proteína por hora.
Análisis Estadísticos
Todos los datos fueron analizados con ANOVA a una vía para mediciones repetidas. Luego fueron analizados los efectos principales usando un test post hoc Tukey para diferencias significativas. Los valores son presentados como medias±DS.
RESULTADOS
Mediciones de Rendimiento
Las mediciones, antes y después del entrenamiento, de producción de potencia anaeróbica pico y promedio (trabajo total) para las cuatro series de ejercicio están ilustradas en la Figura 1. Aunque las diferencias en la producción de potencia no fueron estadísticamente significativas para la primera serie de ejercicio, en cada una de las siguientes tres series, tanto la producción de potencia pico y el trabajo total en 30 s, fueron significativamente mayores después del entrenamiento (p<0.05). El efecto del programa de entrenamiento sobre el VO2 máx. está ilustrado en la Figura 2. El VO2 máx. se incremento desde 3.73±0.13 hasta 4.01±0.08 l/min (p<0.05). Debido a que no hubo cambios significativos en la masa corporal a través del programa del entrenamiento, el VO2 máx. relativo a la masa corporal también se incrementó significativamente desde 51.0±1.8 hasta 54.5 ml.kg-1.min-1.
Actividad Enzimática Muscular
Los efectos del programa de entrenamiento sobre la actividad enzimática glucolítica máxima estan resumidos en la Figura 3 y la Tabla 2. Después del entrenamiento, la actividad de la Hex fue un 56% más alta (p<0.05) y la de la PFK fue ≈49% más alta (p<0.05), que antes del entrenamiento. El cambio total de 9% en la actividad de la fosforilasa y el cambio de 7% en la LDH (Tabla 2) no fueron significativamente diferentes. La actividad de las enzimas oxidativas esta resumida en la Figura 4 y la Tabla 2. El entrenamiento resultó en incrementos significativos (p<0.05) en la actividad de la CS (en ≈36%), MDH (en ≈29%), y SDH (en ≈65%). El cambio de 39% en la 3-hidroxiacil-CoA dehidrogenasa (Tabla 2) no fue significativo.
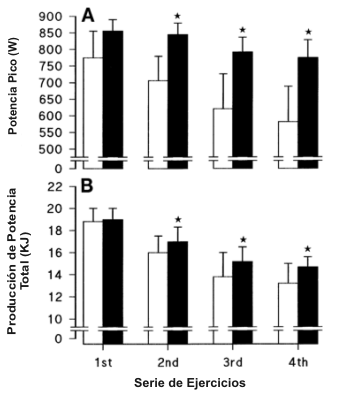
Figura 1. A: producción de potencia pico alcanzada durante 4 esfuerzos
máximos sucesivos de 30 s (protocolo de Wingate). Los histogramas abiertos
indican los valores pre-entrenamiento y los histogramas cerrados indican los
valores post-entrenamiento. B: trabajo total a través de un esfuerzo de 30 s.
Los valores están expresados como medias±DS; n=12 hombres. * p<0.05.

Tabla 2. Actividad enzimática de la TPhos, LDH, y 3-HAD, antes y después del
entrenamiento. Los valores son presentados como medias±DS, expresados en mol •
kg proteina 1 • h 1; n = 9 hombres. TPhos, glucógeno
fosforilasa total; LDH, lactato dehidrogenasa; 3-HAD, 3-hidroxiacil-CoA
dehidrogenasa. Todos los cambios fueron no significativos.
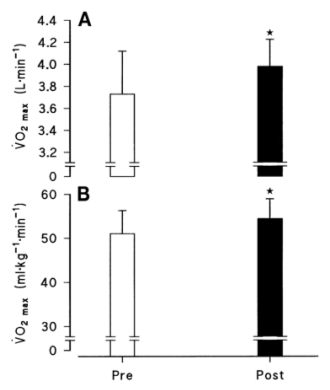
Figura 2. Máximo consumo de oxígeno (VO2 máx.), antes (Pre) y
después (Post) del entrenamiento. Los valores están expresados en unidades
absolutas (l/min) y relativas a la masa corporal (ml.kg-1.min-1)
en B. Los valores están expresados como medias±DS; n=12 hombres. * p<0.05.
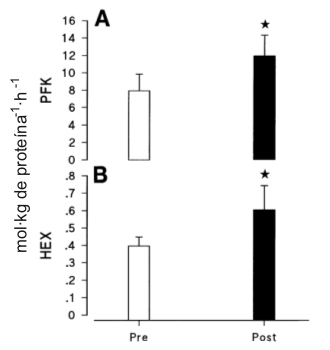
Figura 3. Actividad enzimática máxima para la fosfofructo quinasa (PFK; A) y
hexoquinasa (Hex; B), antes y depués de entrenamiento. Los valores están
expresados como medias±DS; n=12 hombres. * p<0.05.
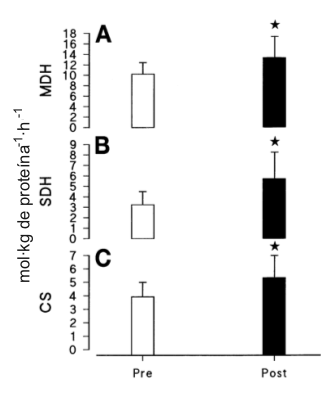
Figura 4. Actividad enzimática máxima para la malato dehidrogenasa (MDH; A),
succinato dehidrogenasa (SDH; B), y citrato sintetasa (CS; C), antes y depués
del entrenamiento. Los valores están expresados como medias±DS; n=12 hombres. *
p<0.05.
DISCUSION
A través de las 7 semanas de entrenamiento, los intervalos de ejercicio fueron realizados a la máxima intensidad. Durante la última semana de entrenamiento, las muestras de sangre capilar de la yema de los dedos en una submuestra de 5 sujetos indicaron concentraciones de lactato de 29-32 mmol/l después de los 10 intervalos de ejercicio. A través de los 4 intervalos de evaluación para medir los cambios en la producción de potencia máxima, las concentraciones de lactato arterial alcanzaron ≈25 mmol/l y las concentraciones de la vena femoral 27 mmol/l. Aunque esta intensidad de entrenamiento puede ser considerada como muy estresante e incómoda, la duración total del ejercicio fue extremadamente corta, alcanzando solo 5 min por sesión durante las últimas semanas de entrenamiento. A pesar del estímulo de entrenamiento relativamente corto, el programa resultó en mejoras significativas en el VO2 máx., producción de potencia máxima de corta duración, y en incrementos en la actividad máxima de tanto, las enzimas marcadoras glucolíticas, como oxidativas.
Nuestro hallazgo de incremento de la actividad de la PFK después del entrenamiento de sprint es consistente con varios estudios previos en sujetos humanos (2, 16, 25). El incremento de la actividad de estas enzimas reguladas alostericamente puede haber resultado en una aceleración de la velocidad del flujo glucolítico durante los esfuerzos máximos y así, al menos parcialmente, explico tanto la mayor producción de potencia pico y promedio de corta duración que fue encontrada en las series 2, 3 y 4, después del entrenamiento. No es conocido el porque el incremento de la actividad de la PFK no resultó en un incremento de la producción de potencia pico en la primera serie de ejercicio, como fue hallado por McKenna et al. (22) después de un protocolo de entrenamiento similar, pero este resultado es consistente con hallazgos de Jacobs et al. (16). El efecto inhibidor de la [H+] sobre la actividad de la PFK es bien conocido (3), y es posible que las diferencias de rendimiento asociadas con una mayor actividad de la PFK, podría ser esperado que se convirtieran en incrementalmente aparentes, a medida que el músculo se vuelve más acidótico. Además, la mejora de la capacidad de la bomba Na+-K+ y un posible incremento en la tolerancia a los H+ puede también contribuir al incremento en el trabajo total realizado después del entrenamiento. La significancia del gran incremento en la Hex es de algún modo más difícil de interpretar. No es sabido, sin embargo, que el ejercicio intervalado intenso, como en el presente estudio, sea acompañado por grandes incrementos en la concentración de glucosa en el plasma (20) y, presumiblemente, el músculo. Así un incremento en la actividad de la Hex incrementaría el potencial para una mayor velocidad de utilización del glucógeno durante el ejercicio y los intervalos de recuperación.
El incremento significativo en el VO2 máx. y los grandes incrementos en la actividad de las enzimas oxidativas fueron de algún modo inesperados, dada la naturaleza del estímulo de entrenamiento y a su brevedad. Los cambios en esta magnitud están usualmente asociados con programas de entrenamiento que implican varias horas por semana a una intensidad de ejercicio submáxima (7, 26, 30). Aunque el (los) mecanismo (s) mediante el cual tal entrenamiento de resistencia mejora la actividad de las enzimas oxidativas es desconocido, generalmente se pensó que tanto la intensidad de ejercicio (aquella en donde hay una gran implicancia de las vías metabólicas oxidativas) y la duración del estímulo (volumen del entrenamiento) son componentes importantes. En el presente estudio, el metabolismo oxidativo puede ser considerado en tener solo una contribución mínima al aporte de energía durante cada intervalo de ejercicio. Aunque esta contribución relativa probablemente se incrementa con los sucesivos intervalos de ejercicio (29), el consumo de oxígeno respiración a respiración, con un ejercicio similar, indico valores pico al final de los 30 s que son <60% del VO2 máx. y una declinación rápida después del período de recuperación (17). Además, la duración del estímulo de entrenamiento fue extremadamente corta en el presente estudio, ascendiendo a un total de 6 min/semana en la semana 1 e incrementándose a un total de 15 min/semana en la semana 7.
En comparación con la literatura sobre entrenamiento de resistencia, ha habido relativamente pocas investigaciones acerca de los efectos del entrenamiento de sprint sobre las enzimas mitocondriales y la potencia aeróbica en humanos. Entre estos, sin embargo, hay reportes de incrementos en el VO2 máx. (8, 27) e incrementos en la actividad de la CS muscular (16), después del entrenamiento. Ha sido encontrado que el entrenamiento de sprint incrementa la actividad de la CS en el músculo rojo vasto lateral de ratas (19). En base a esta literatura y a los hallazgos del presente estudio, parece que el entrenamiento a una intensidad que exceda el VO2 máx. puede ser un componente más importante que el volumen del entrenamiento, para estimular un incremento en el potencial oxidativo muscular. La inspección de la producción de potencia individual durante 30 s alcanzada durante las sesiones de entrenamiento de sprint indica que las mismas variaron desde un equivalente a ≈ 210% VO2 máx. en el primer intervalo hasta ≈ 140% VO2 máx. en el intervalo 9 y 10. A tales intensidades, la velocidad de producción del piruvato puede ser considerada como casi máxima, y uno esperaría grandes incrementos en la velocidad de la actividad catalítica de las enzimas competitivas piruvato dehidrogenasa (PDH) y LDH. Aunque el incremento de 7% en la actividad de la LDH después del entrenamiento no fue estadísticamente significativo, nosotros no medimos la actividad de la PDH. Uno puede especular, sin embargo, que se hubiera incrementado la actividad de la PDH, esto hubiera resultado en un incremento en la velocidad de entrada del piruvato en la mitocondria y quizás, ese fue el estímulo para el aumento en la regulación de las enzimas mitocondriales.
En conclusión, nosotros concluimos que el entrenamiento intervalado de sprint relativamente intenso, pero corto puede resultar en un incremento en tanto la actividad enzimática muscular oxidativa y glucolítica, producción de potencia máxima de corta duración, y VO2 máx. El incremento en la producción de potencia puede haber sido un resultado del incremento en la actividad enzimática glucolítica máxima y en la capacidad de la bomba Na+-K+, mientras que el incremento en la actividad enzimática mitocondrial puede haber sido el resultado del incremento de la velocidad del flujo del piruvato durante tal entrenamiento.
Agradecimientos
Los autores agradecen la ayuda de Jennifer O’Brien y Alex Lauzier en la realización de este estudio.
Notas al Pie de Página
Los subsidios para este estudio fueron proporcionados por el Natural Science and Engineering Research Council de Canadá.
Dirección para Correspondencia
J. D. MacDougall, Dept. of Kinesiology, McMaster University, Hamilton, Ontario, Canada L8S 4K1.
Referencias
1. Bar-Or, O (1987). The Wingate anaerobic test: an update on methodology, reliability, and validity. Sports Med. 4: 381-394
2. Cadefau, J., J. Casademont, J. M. Grau, J. Fernandez, A. Balaguer, M. Vernet, R. Cusso, and A. Urbano-Marquez (1990). Biochemical and histochemical adaptation to sprint training in young athletes. Acta Physiol. Scand. 140: 341-351
3. Chasiotis, D., K. Sahlin, and E. Hultman (1982). Regulation of glycogenolysis in human muscle at rest and during exercise. J. Appl. Physiol. 53: 708-715
4. Costill, D. L., E. F. Coyle, W. F. Fink, G. R. Lesmes, and F. A. Witzmann (1979). Adaptations in skeletal muscle following strength training. J. Appl. Physiol. 46: 96-99
5. Davies, K., J. A. L. Packer, and G. A. Brooks (1982). Exercise bioenergetics following sprint training. Arch. Biochem. Biophys. 215: 260-265
6. Davies, K., J. A. L. Packer, and G. A. Brooks (1981). Biochemical adaptation of mitochondria, muscle and whole animal respiration to endurance training. Arch. Biochem. Biophys. 209: 538-553
7. Henriksson, J., M. M.-Y. Chi, C. S. Hintz, D. A. Young, K. K. Kaiser, S. Salmons, and O. H. Lowry (1986). Chronic stimulation of mammalian muscle: changes in enzymes of six metabolic pathways. Am. J. Physiol. 251 (Cell Physiol. 20): C614-C632
8. Henriksson, J., and J. S. Reitman (1977). Time course of changes in human skeletal muscle succinate dehydrogenase and cytochrome oxidase activities and maximal oxygen uptake with physical activity and inactivity. Acta Physiol. Scand. 99: 91-97
9. Henriksson, J., and J. S. Reitman (1976). Quantitative measures of enzyme activities in type I and type II muscle fibres of man after training. Acta Physiol. Scand. 97: 392-397
10. Hickson, R. C., W. W. Heusner, and W. D. Van Huss (1976). Skeletal muscle enzyme alterations after sprint and endurance training. J. Appl. Physiol. 40: 868-872
11. Holloszy, J. O (1975). Adaptation of skeletal muscle to endurance exercise. Med. Sci. Sports Exerc. 7: 155-164
12. Holloszy, J. O. and F. W. Booth (1976). Biochemical adaptation to endurance exercise in muscle. Annu. Rev. Physiol. 38: 273-291
13. Hoppeler, H (1986). Exercise-induced changes in skeletal muscle. Int. J. Sports Med. 7: 187-204
14. Hoppeler, H., H. Howald, K. Conley, S. L. Lindstedt, H. Claasen, P. Vock, and E. R. Weibel (1985). Endurance training in humans; aerobic capacity and structure of skeletal muscle. J. Appl. Physiol. 59: 320-327
15. Kowalchuk, J. M., G. J. F. Heigenhauser, M. I. Lindinger, G. Obminski, J. R. Sutton, and N. L. Jones (1988). Role of lungs and inactive muscle in acid-base control after maximal exercise. J. Appl. Physiol. 65: 2090-2096
16. Linossier, M. T., C. Denis, D. Dormois, A. Geyssant, and J. R. Lacour (1993). Ergometric and metabolic adaptation to a 5-s sprint training programme. Eur. J. Appl. Physiol. 67: 408-414
17. Loy, S. F., and L. D. Segel (1988). 31P-NMR of rat thigh muscle: effect of sprint and endurance training. Med. Sci. Sports Exerc. 20: S65
18. MacDougall, J. D., G. R. Ward, J. R. Sutton, and D. G. Sale (1977). Muscle glycogen repletion following high-intensity intermittent exercise. J. Appl. Physiol. 42: 129-132
19. McKenna, M. J., G. J. F. Heigenhauser, R. S. McKelvie, J. D. MacDougall, and N. L. Jones (1997). Sprint training enhances ionic regulation during intense exercise in men. J. Physiol. (Lond.) 501: 687-702
20. McKenna, M. J., T. A. Schmidt, M. Hargreaves, L. Cameron, S. L. Skinner, and K. Kjeldsen (1993). Sprint training increases human skeletal muscle Na+-K+-ATPase concentration and improves K+ regulation. J. Appl. Physiol. 75: 173-180
21. Nevill, M. E., L. H. Boobis, S. Brooks, and C. Williams (1989). Effect of training on muscle metabolism during treadmill sprinting. J. Appl. Physiol. 67: 2376-2382
22. Phillips, S. M., H. J. Green, M. A. Tarnopolsky, G. J. F. Heigenhauser, and S. M. Grant (1996). Progressive effect of endurance training on metabolic adaptations in working skeletal muscle. Am. J. Physiol. 270 (Endocrinol. Metab. 33): E265-E272
23. Roberts, A. D., R. Billeter, and H. Howald (1982). Anaerobic muscle enzyme changes after interval training. Int. J. Sports Med. 3: 18-21
24. Sale, D. G., J. D. MacDougall, I. Jacobs, and S. Garner (1990). Interaction between concurrent strength and endurance training. J. Appl. Physiol. 68: 260-270
25. Saltin, B. K., K. Nazar, D. L. Costill, D. L. Stein, E. Jansson, B. Essen, and P. D. Gollnick (1976). The nature of the training response: peripheral and central adaptations to one-legged exercise. Acta Physiol. Scand. 96: 289-305
26. Slievert, G. G., R. D. Backus, and H. A. Wenger (1995). The influence of a strength-sprint training sequence on multi-joint power output. Med. Sci. Sports Exerc. 27: 1655-1665
27. Spriet, L. L., M. I. Lindinger, R. S. McKelvie, G. J. F. Heigenhauser, and N. L. Jones (1989). Muscle glycogenolysis and H+ concentration during maximum intermittent cycling. J. Appl. Physiol. 66: 8-13
28. Wibom, R., E. Hultman, J. Johansson, K. Matherei, D. Constantin-Teodosiu, and P. G. Schantz (1992). Adaptation of mitochondrial ATP production in human skeletal muscle to endurance training and detraining. J. Appl. Physiol. 73: 2004-2010